ROLE OF AUXIN AND POLYAMINES IN ADVENTITIOUS ROOT FORMATION
AT THE BASE OF MUNG BEAN CUTTINGS
S. NAG, K SAHA AND M.A. CHOUDHURI• Botany Department, Burdwan University, Burdwan 713104
Received on28 Oct., 1998, Revised on 4 Aug., 1999
SUMMARY
Indole-3-acetic acid (IAA) and indole-3-butyric acid (IBA) at the cone. of 10-s M stimulated adventitious root formation at the base of stem cuttings of mung bean (Vigna radiata L. cv. 105). IBA was more effective than IAA in this respect. Treatment for shorter duration in either IAA (4 days) or IBA (3 days) produced better root development than their continuous treatment (12 days) and spermine (10-5 M for 1 day) for shorter duration markedly improved adventitious root formation,
whereas spermidine had the least effect on adventitious root formation. Treatment of stem cuttings with specific inhibitors of ornitheine decarboxylase (ODC) and arginine decarboxylase (ADC) of putrescine biosynthesis viz., a.-difluoromethyl ornithine (DFMO) and a.-difluoromethyl arginine (DFMA), respectively and the counteraction of putrescine on the inhibitory effect of these chemicals on rooting indicated that the putrescine is involved in the formation of adventitious root and that the ODC pathway is the preferred pathway for putrescine biosynthesis which is implicated in adventitious root formation in mung bean cuttings.
Key words: Auxin, polyamines, mung bean, adventitious root.
INTRODUCTION
The proper formation of adventitious roots at the base of stem cuttings is an important developmental phenomenon in the growth and survival of stem cuttings. It is interesting from the point of view of plant development that it involves the initiation of several new meri:.tematic areas in the differentiated tissues of stem cuttings and their subsequent development into mature root structures. This process appears to be governed by an array of endogenous physiological factors (Hess, 1962, Davis and Haissig, 1994; Jackson, 1986; Davis eta/., 1988; Gaspar eta!., 1997). Among the endogenous factors, auxin is
generally credited as the prime trigger for root initiation (Haissig, 1974; Torrey, 1986; Gaspar and Hofinger, 1988; Bellamine eta! .. 1998). Polyamines (PAs), another
group of endogenous plant grov.1h regulators, are also implicated in rooting of stem cuttings (Jarvis eta!.. 1983:
*
Author for correSJX?)ldenceJndwn.J. Plant Phys10l.. Vol. 4. l'?. 4, (0-'.S.) pp. 247-253 (Oct-D-.;c .. 1999)
Walker et al.. 1985; Sankhla and Upadhyaya, 1988;
Kevers et al .. 1997; Ballester et al., 1999). Although PAs
have been implicated in adventitious root formation under in vitro and in vivo conditions, there is a great deal of controversy regarding the exact role played by PAs in adventitious root formation at the base of stem cuttings and its relationship with auxin (Friedman et
aL
1982. 1985;Jarviseta/., l983;SankhlaandUpadhyaya, 1988)The present study was, therefore. undertaken to investigate the role of both auxins and PAs in adventitious root fom1ation at the base of mung bean cuttings. The study also included the analysis of pretreatment time, in auxin or PAs, reqtmed by the cuttings for the development
of root primordia and primary roots in cuttings. In addition, continuously for a period of 12 days and the rooting attempts were also made to elucidate the involvement of performance was recorded in each case.
P A in root initiation and development using specific inhibitors of P A biosynthesis and estimating rooting performance of cuttings.
MATERIALS AND METHODS
Mung bean seeds (Vigna radiata L. cv. 105) were surface sterilized with 4% sodiumhypochlorite (w/v) before germination and seedlings raised in a temperature (24
±
1 °C} and light ( 16 hours photoperiod at 220 j..Lmoles m·2s-1 at 400-700 run) controlled growth chamber up to 6days. Six-day-old seedlings were excised 3 em below the cotyledonary node, the cotyledons were removed and the resulting stem cuttings (consisting of hypocotyl and the intact epicotyl with a pair of primary leaves and apical buds) were used in rooting experiments.
Freshly prepared hypocotyl cuttings were dipped into glass beakers containing 50 ml distilled water (control) or test solutions included : the auxin derivatives viz. indole-3-acetic acid (IAA, I0-5 and I0-6 M),
indole-3-butyric acid (IBA, 1
o-s
and 10-6M), and polyamines viz.,putrescine ( 10-4 and 1
o-s
M), spennidine ( 1 o-~ and 10-9M)and spem1ine ( 1
o-s
and 1 Q-6 M). The results presented hereincluded only the effective optimum cone. of these substances such as IAA ( 1
o-s
M), IBA ( 1o-s
M), putrescine ( 1o-"
M), spennidine ( 1 Q-9 M) and spem1ine ( 1 0-5 M) . Thecuttings were initially pretreated in each of the above chemicals at the optimum cone. for different durations ( l, 2, 3 or 4 days) and transferred to distilled water to
In all the above experiments, the treated cuttings were kept in the growth chamber under same temperature and light conditions at which the seedlings were raised. After the stipulated periods, the cuttings were taken out and the following observations were made to characterize rooting performance: number of root primordia formed, number and length of primary root and secondary root per cutting.
The experiments were repeated at least three times and statistically analysed by using CRD factorial design. (Panse and Sukhatme, 1967).
RESULTS AND DISCUSSION
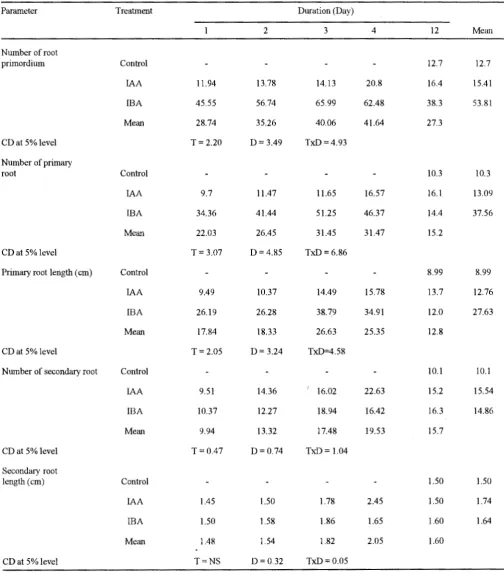
The perusal ofthe data included in Table I. indicated that both the IAA and IBA treatment had significant effect on different characteristics of rooting of cuttings of Vigna radiata over control, except the secondary root length. Though IAA treatment produced significantly higher number of secondary root (15.54), the number of root primordium (53.81), number of primary root (37.56) and primary root length (27.63 em) were found significantly greater with IBA treatment. Treatment duration upto 4 days increased all the growth parameters significantly. but a sharp decline in root growth was observed during 12 days treatment. The effect oftreatment duration from 1 to 3 days was at par in all the gro\\-th parameters except the number of root primordium and number of secondary root. Further, it was noted (Table I)
complete 12 days. In another set of experiments, the thattheinteractioneffectofgrowthregulatorandtreatment cuttings were continuously treated with each test solution duration was statistically significant. Effect of IBA at 3 for 12 days to ascertain the effective time of action of days duration gave the significantly best result in most of
auxins and polyamines in adventitious root fom1ation.
In a separate set of experiments, the cuttings were treated with an inhibitor of putrescine biosynthesis, a-difluoromethyl ornithine (DFMO, IO-', 10-4 & I0-5 M) or
a~difluoromethyl arginine (DFMA, I0-3
, I0-4 & l()-5 M
ci ther alone or in presence of putrescine (1 0-4 M)
248
the cases whereas IAA at 4 days duration showed bL·t · effect on secondary root length (2.45 em). lt is clear lll~tt
IBA was more effective in improving rooting perfom1ance than IAA in mung bean. One of the reasons might be ascribed to greater susceptibility of IAA to the attack of IAA-oxidase systems than IBA. The occurrence of maximum root primordia and primary roots of cuttings
TABLE I : Eflect of different duratwns of IAA and IBA (lo-s M) treatment on rooting of cuttings of Vigna radiata L
Parameter Treatment Duration (Day)
2 3 4 12 Mean
Number of root
primordium Control 12.7 12.7
IAA 11.94 13.78 14.13 20.8 16.4 15.41
IBA 45.55 56.74 65.99 62.48 38.3 53.81
Mean 28.74 35.26 40.06 41.64 27.3
CD at 5% level T= 2.20 D= 3.49 TxD=4.93
Number of primary
root Control 10.3 10.3
IAA 9.7 11.47 11.65 16.57 16.1 13.09
IBA 34.36 41.44 51.25 46.37 14.4 37.56
Mean 22.03 26.45 31.45 31.47 15.2
CD at 5% level T = 3.07 D = 4.85 TxD=6.86
Primary root length (em) Control 8.99 8.99
IAA 9.49 10.37 14.49 15.78 13.7 12.76
IBA 26.19 26.28 38.79 34.91 12.0 27.63
Mean 17.84 18.33 26.63 25.35 12.8
CD at 5% level T= 2.05 D= 3.24 TxD=4.58
Number of secondary root Control 10.1 10.1
lAA 9.51 14.36 16.Q2 22.63 15.2 15.54
IBA 10.37 12.27 18.94 16.42 16.3 14.86
Mean 9.94 13.32 17.48 19.53 15.7
CD at 5% level T = 0.47 D=0.74 TxD = 1.04
Secondary root
length (em) Control 1.50 1.50
IAA 1.45 1.50 1.78 2.45 1.50 1.74
IBA 1.50 1.58 1.86 1.65 1.60 1.64
Mean 1.48 1.54 1.82 2.05 1.60
CD at 5% level T=NS D = 0.32 TxD=0.05
treated with either auxin for a period of 4 days and not beyond that suggests that the auxin active stage of rooting (Hartmann et al., 1993) in mung bean is extended up to 4 days which is followed by an auxin-inactive stage or even auxin-inhibitive stage if the duration of auxin treatment was extended up to 12 days. The results also reveal that prior to emergence of roots, there must be an optimum level of auxin for the induction of meristematic activity and beyond an optimum level, the auxin might be inhibitory to this meristematic activity at the base of the cuttings.
The data of Table II revealed that the effect of PUT and SPM was at par on number of root primordium (17. 99 and 17.69, respectively), but both had significantly greater effect than SPD. This is quite in agreement with the observations of Jarvis et al. (1983) in mung bean. Further, it was noted that SPD did not show any significant effect over control on number of root primordium. Though PUT always produced significantly better growth, SPD and SPM produced similar effect on number of primary root (11.35 & 11.15, respectively) and number of secondary root (9.28 & 8.93 respectively). But SPM had significantly greater effect than ~PD in case of primary root length ( 11.03 & 9.17 em) and secondary root length (1.49 & 0.94 em). It was interesting to note that SPD and SPM had lower effect on number of secondary root and secondary root length than the control. Number of root primordium and number of primary root decreased with the increase of durations of treatments and a drastic reduction was noted at 12 day treatment. No significant change in primary root length and number of secondary root was observed upto 4 days but at 12 days a decrement of these values was observed. Treatment duration had no significant effect on secondary root length. Again, it was found (Table II) that the interaction effect of chemical treatments and durations was statistically significant in all the cases except the number of primary root. Among the treatments, PUT at 3 days produced the maximum effect in case of number of root primordium (22.34) primary root (17. 75) and primary root length (19 .02 em), whereas SPM exerted its effect earlier than 3 days i.e., earlier than PUT. The effect of PUT at 12 days was found
250
to be greatest on number of secondary root (13.9) and secondary root length (3 .25 em) whereas such duration of treatment with SPM produced inhibitory effect. Thus both PUT and SPM exerted their effects on rooting process earlier than auxin as evident from Tables I and II. Thus it can be concluded that early phase of adventitious root formation (possibly inductive phase) required high levels of PAs, particularly PUT and/or SPM in mung bean.AlthoughFriedmaneta/., (1982)foundnopromotive effects of PAs on rooting of cuttings, our observations lend support to those ofJarvis eta/., (1983) who reported promotive effects of SPM and inhibitory effects of SPD in rooting of mung bean cuttings.
From the Table III it is observed that PUT+ DFMA produced significantly better result i.e. lower inhibitory effect on number of primary root (9 .2) number of secondary root (8.68) and secondary root length (14.3 em). Though the inhibitory effect of PUT+ DFMO and PUT+ DFMA was almost same on primary root length (8.57 & 7. 99 em respectively), the number of root primordium was higher (10.50) with PUT+ DFMO treatment only. DFMO and DFMA treatment alone gave higher inhibitory effect (i.e. lower growth value) than in the presence of PUT. All the above treatments, however, could not produce root growth values which remained below that of control. All the growth paranteters increased sharply with the lowering of concentration from 10-3 M to 10-5 M. The concentration of
10-3 M and 1
o-
4 M had no significant variation in effect onprimary root length.It was also noted (Table III) that interaction effect of inhibitor and concentration of inhibitors was found statistically significant in all the growth parameters except primary root length. PUT + DFMA at l0-5 M concentration gave significantly higher
value (i.e. lesser inhibition) of number of root primordium (12.26), number of primary root (10.54) and number of secondary root (9.85). But PUT+ DFMO and PUT+ DFMA at 10-5 M concentration gave significantly at par
effect (1.81 em and 1.65 em, respectively) in case of secondary root length. DFMO and DFMA alone provided more inhibitory effect i.e. lower value of root gro-wth than in the presence of PUT. The results thus suggest that PUT plays an important role in the process of adventitious root
TABLE II: Eflect of different durations of putrescine (PUT, 104M), supermidine (SPD, 1 0·9 M) and spermine (SPM, 1 0·5 M) treatments on
rooting of cuttings of Vigna radiata L.
Parameter Treatment Duration (Day)
2 3 4 12 Mean
Number of root
primordium Control 12.7 12.7
PUT 17.08 17.28 22.34 20.46 12.8 17.99
SPD 13.06 13.54 13.83 13.98 8.4 12.56
SPM 28.2 21.16 17.06 13.62 8.4 17.69
Mean 19.45 17.33 17.74 16.02 9.87
CD at 5% level T = 1.47 D = 1.90 TxD=3.29
Number of primary
root Control 10.3 10.3
PUT 15.09 15.68 17.75 17.71 10.8 15.41
SPD 11.46 12.23 12.51 12.94 7.6 11.35
SPM 15.79 12.29 10.86 9.21 7.6 11.15
Mean 14.12 13.40 13.71 13.29 8.67
CD at 5% level T = 1.76 D= 2.27 TxD=NS
Primary root length (em) Control 8.99 8.99
PUT 15.76 16.57 19.02 17.77 11.35 16.10
SPD 9.21 9.46 10.79 10.99 5.40 9.17
SPM 14.62 13.98 10.25 9.31 7.0 11.03
Mean 13.20 13.34 13.35 12.69 7.92
CD at 5% level T = 0.50 D= 0.65 TxD=l.l2
Number of secondary root Control 10.1 10.1
PUT 9.48 9.71 10.52 9.85 13.9 10.69
SPD 10.73 10.21 9.55 9.89 6.0 9.28
SPM 9.05 9.66 9.79 10.12 6.0 8.93
Mean 9.75 9.86 9.95 9.95 8.63
CD at 5% level T = 0.45 D = 0.58 TxD = 1.0
Secondary root
length (em) Control 1.5 1.5
PUT 1.21 1.28 1.33 1.28 3.25 1.67
SPD 1.07 1.06 0.91 0.99 0.66 0 ')4
SPM 1.2 1.74 1.82 1.97 0.7 1.49
Mean 1.16 1.36 1.35 1.41 1.54
CD at 5% level T = 0.26 D=NS Tx D= 0.58
TABLE III: Effect of continuous treatment ofDFMO and DFMA alone or in presence of putrescine (104M) on rooting of cuttings of Vigna
radiata L.
Parameter Treatment Concentrations (M)
10-3 10-4 10-5 Mean
Number of root primordium Control 12.7
DFMO 7.04 7.28 9.05 7.79
DFMA 5.76 8.72 11.01 8.50
PUT+DFMO 8.29 11.07 12.13 10.50
PUT+DFMA 7.08 11.37 12.26 10.24
Mean 7.04 9.61 11.11
CD at 5% level T =0.1
c
= 0.08 TxC=0.17Number of primary root Control 10.3
DFMO 5.74 6.02 7.65 6.47
DFMA 4.98 6.02 8.53 6.51
PUT+DFMO 7.76 8.22 9.15 8.38
PUT+DFMA 7.25 9.81 10.54 9.20
Mean 6.43 7.52 8.98
CD at 5% level T = 0.51
c
= 0.43 Tx C = 0.89Primary root length (em) Control 8.99
DFMO 6.38 6.4 7.31 6.70
DFMA 2.63 5.85 6.78 5.09
PUT+DFMO 8.3 8.53 8.88 8.57
PUT+DFMA 7.4 8.17 8.41 7.99
Mean 6.18 7.24 7.85
CD at 5% level T= 1.45
c
= 1.25 TxC=NSNumber of secondary root Control 10.1
DFMO 0.20 4.12 7.75 4.02
DFMA 6.85 7.64 7.80 7.43
PUT+DFMO 3.07 4.88 6.46 4.80
PUT+DFMA 7.14 9.06 9.85 8.68
Mean 4.32 6.43 7.97
CD at 5% level T = 1.06
c
= 0.92 TxC=1.84Secondary root length (em) Control 1.5
DFMO 0.31 0.64 1.23 0.73
DFMA 0.87 1.33 1.45 1.22
PUT+DFMO 0.35 1.2 1.81 1.12
PUT+DFMA 1.19 1.45 1.65 1.43
Mean 0.68 1.16 1.54
CD at 5% level T = 0.12
c
= 0.10 Tx C = 0.20fonnation in mung bean cuttings (Biondi et a!., 1990). The greater inhibitory effect of DFMO than that of DFMA suggests that the ODC pathway of PUT biosynthesis had a greater role in mung bean adventitious root formation than ADC pathway.
ACKNOWLEDGEMENT
The authors are grateful to Prof. A. Altman, the Hebrew University of Jerusalem, Israel for gifting us DFMO and DFMA.
REFERENCES
Ballester A, San-Jose M. C., Vidal N., Fernandez-Lorenza JL. and Vietez A M. (1999). Anatomical and biochemical events during in vitro rooting of microcuttings from juvenile and mature phases of chestnut. Ann. Bot. 83 : 619-62 9.
Bell amine J., Penel C., Greppin H. and Gaspar T. ( 1 998 ). Confirmation ofthe role ofauxinand calcium in the late phases of adventitious root formation. Plant Growth Regul. 26: 191-194.
Biondi,S., Diaz, T., Iglesias, I., Gamberinig, G. andBagni,N. (1990). Polyamines and ethylene in relation to adventitious root formation in Prunus avium shoot cultures. Physiol. Plant., 78 : 474-483.
Davis, T.D. and Haissig, B.E. (1994 ). In: Biology ofRoot Formation. (T.D. Davis, and B. E. Haissig, eds. ), Plenum Press, New York.
Davis, T.D. andHaissig, S.E. andSankhla,N. (1988). In:Adventitious Root Formation in Cuttings (T.D. Davis, B.E. Haissig and N. Sankhla, eds.) Advances in Plant Sciences Series, Vol. II. Dioscorides Press. Portland, Oregon.
Friedman, R., Altman, A and Bachrach, U. (1982). Polyamines and root formation in mung beans hypocotyl cuttings. I. Effects of exogenous compounds and changes in endogenous polyamine content. Plant Physiol., 70 : 844-848.
Friedman, R., Altman, A and Bachrach, U. (1985). Polyamines and root formation in mung beans hypocotyl cuttings. II. Incorporation of precursors into polyamincs. Plant Physiol., 78 . 80-83.
Gaspar, T. and Hofinger, M. ( 1988). Auxin metabolism during adventitious rooting. In : Adventitious Root Formation in
JndianJ. Plant Physiol., Vol. 4. No.4, (N.S.) pp. 247-253 (Oct.-Dec., 1999)
Cuttings. (T.D. Davis, and B.E. Haissig, and N. Sankhla, eds).
Advances in Plant Sciences Series, Vol. II. Dioscorides Press, Portland, Oregon. pp. 117-131.
Gaspar, T., Kevers, C. and Hausman, J.F. (1997). Indissociable chief factors in the inductive phase of adventitous rooting. In : Biology of Root Formation and Development. (A Altman and Y. Waisel eds.). Plenum Press, New York pp. 55-63.
Haissig, B.E. (1974). Influence of auxins and auxin synergists on adventitious root primordium initiation and development. New Zealand J. For. Sci., 4 : 311-323.
Hartmann, H.T., Kester, D.E. and Davies, F.T. (1993). Plant Propagation-Principles and Practices. 5th edn. Prentice Hall of India Pvt. Ltd. New Delhi. pp. 199-255.
Hess, C.E. ( 1962 ). Characterization of the rooting co factors extracted from Hedera helix L. and Hibiscus rosa-sinensis. Proc. 16th Inter. Herb. Cong. pp. 382-88.
Jackson, M.B. (1986). New Root Formation in Plant and Cuttings (M.B. Jackson, ed.) Martinus NijhoffPubl., Dordrecht, USA
Jarvis, B.C.,Shannon,P.R.M. and Yasmin,S. (1983). Involvement of polyamines with adventitious root development in stem cuttings of mung bean. Plant Cell Physiol., 24 : 677-683.
Kevers, C., Bringaud, C., Hausman, J.F. and Gaspar, T. (1997). Putrescine involvement in the inductive phase of walnut shoots rooting in vitro. Saussurea 28: 47-57.
Panse, V.G. and Sukhatme, P.T. (1967). Statistical Methods for Agricultural Works. 2rid edn. Indian Council of Agricultural Research, New Delhi.
Sankhla, N. and Upadhyaya, A (1988).• Polyamines and adventitious root fonnation. In : Adventitious Root Fonnation In Cuttings. (T. D. Davis, and B. E. Haissig, and N. Sankhla, eds. ). Advances in Plant Sciences Series, Vol. II. Dioscorides Press. Portland, Oregon. pp. 202-131.
Torrey, J.G. (1986). Endogenous and exogenous influences on the regulation oflateral root fonnation. In: New Root Formation m Plant and Cuttings (M.B. Jackson ed. ). Martin us Nijhoff Pub!., Dordrecht, USA pp. 32-66.
Walker, M.A., Roberts, D.R., Shih, C.Y. andDumbrof{ E. F. (1985). A reqmrement for polyamines during cell divis10n phase of radicle emergence in seeds of Acer saccharum. Plant Cell Physiol., 26: 967-971.