Abstract
Cardiomyopathies are rare but serious disorders that affect the heart muscle and require lifetime treatment in patients surviving to adulthood. Mutations in cardiac
sarcomere components lead to the development of symptomatic cardiomyopathies such as cardiac hypertrophy; however, it has not yet been assessed whether the
misexpression of non-cardiac sarcomere paralogs in the developing heart may also contribute to the development of early onset cardiomyopathies. Ablation of Chd4, the catalytic subunit of the transcriptional repressive Nucleosome Remodeling and
Deacetylase (NuRD) complex, leads to the misexpression of fast skeletal and smooth muscle isoforms of sarcomeric proteins in the developing murine heart. Utilizing this model of non-cardiac myofibril paralog misexpression, it is possible to assess whether the misexpression of these non-cardiac isoforms leads to impaired cardiac muscle function. Through fluorescent labeling of fast skeletal troponin I2 (fsTnI2), α-actinin, and smooth muscle myosin heavy chain (SMMHC), we found that misexpression of non-cardiac sarcomeric paralogs disrupts non-cardiac sarcomere formation, contributing to impaired heart function and embryonic lethality early in gestation. Wild-type sarcomeres normally exclude non-cardiac isoforms during formation, but following loss of CHD4-mediated transcriptional repression, developing sarcomeres incorporate misexpressed SMMHC and exhibit disorganized structure. Our results suggest that the incorporation of non-cardiac myofibril isoforms in developing cardiac sarcomeres contributes to sarcomere malformation and the development of cardiomyopathies. Identifying these functional requirements of CHD4 in proper heart development will lead to further advancements in the understanding and future treatment strategies for patients with mutations that disrupt this process.
Introduction
Cardiomyopathies are rare but serious disorders that impair the ability of the heart to circulate blood throughout the body. Affecting at least 1 in every 100,000 children in the world, cardiomyopathies require lifetime treatment in the 60% of patients that survive into adulthood, making this group of diseases an important focus of heart development research.1 It has been shown that mutations in cardiac myofibril isoforms
the development of symptomatic cardiomyopathies.2,3,4 However, it has not yet been
assessed whether misexpression of non-cardiac myofibril isoforms in the developing heart may also contribute to the development of cardiomyopathies.
The Nucleosome Remodeling and Deacetylase (NuRD) complex is known to interact with several cardiac transcription factors to regulate gene expression in the developing heart; ablation of these interactions leads to aberrant gene expression and altered cardiac function.5,6,7,8 The NuRD complex regulates chromatin organization, gene
transcription (typically through repression), and developmental signaling pathways.9 The
NuRD complex was the first protein complex found to contain a dual enzymatic activity, containing an ATP-dependent helicase subunit (CHD3/4), which allows it to move and restructure nucleosomes, and a histone deacetylase (HDAC) subunit.9
This study focuses on the core catalytic subunit of the complex, the
chromodomain helicase DNA binding protein 4 (CHD4). Through ATP hydrolysis, CHD4 moves along chromatin and repositions nucleosomes to influence gene target
transcription at the epigenetic level.9 CHD4 is evolutionarily conserved in vertebrate
animals and plants, underscoring its importance as a functional ATPase.9 It has been
found that CHD4 does not bind DNA in a sequence specific manner. Rather, CHD4 and the NuRD complex are located to gene-specific targets through association with tissue-specific transcriptional co-factors, complementing its function as a transcriptional repressor.10
Previous lab work has shown that the conditional knockout of Chd4 in the
Cre-recombinase drivers, driven by the Nkx2-5 locus, a known cardiac transcription factor expressed only in the developing heart, or a transgene construct containing the cardiac troponin T2 promoter (Tnnt2) driving CRE expression. RNA sequencing data revealed a misexpression of Tnnc2, Tnnt3, and Tnni2 (fast skeletal troponin subunits), Acta1 (fast skeletal α-actin), and Myh11 (smooth muscle myosin heavy chain) in Chd4 cardiac null hearts at E10.5, indicating that CHD4 is required to repress expression of these non-cardiac myofibril isoforms in the developing heart.11
At this stage in the developing heart, cardiac sarcomeres are rapidly forming and arranging themselves across cardiomyocytes to produce symmetrical contraction that leads to controlled, unidirectional blood flow.12 The sarcomere is the basic contractile
unit in a cardiomyocyte. Developing myofibrils, or contractile threads in cardiomyocytes, new sarcomeres consist of parallel arrays of 1 μm long thin filaments of actin that intersperse between aligned 1.6 μm long thick filaments of myosin.12 The thin actin
filaments are linked together at the Z-disk and bind together with the scaffolding protein α-actinin.12 Threads of tropomyosin line the actin thin filaments and are interspersed
with troponin complexes, the site of calcium binding during contraction. The cardiac homologs of actin, troponin, and myosin are important in cardiac sarcomere structure and function.
There are three types of muscle tissue present in the body: skeletal, smooth, and cardiac. The different muscle tissues share structural and functional proteins in the same family, such as myosin, but the specific myofibril isoform is specific to the shape and function of each tissue type.13,14 Striated sarcomere structures are present in
muscle are under involuntary control while skeletal muscle is controlled voluntarily. Myosin, important for contraction, has varying isoforms per muscle type that are
adapted for the specific mechanical function of each type.15 The sarcomeric isoforms, in
skeletal and cardiac fibers, have 80% sequence similarity to each other, but the smooth muscle isoform is only 40% similar to the skeletal and cardiac sarcomeric isoforms.15
Cardiac and skeletal muscle fibers utilize the same mechanism of excitation-contraction coupling with calcium binding, but differentially express specific isoforms of the troponin complex unique to the muscle type.13,16 The differences in structure and functionality of
each muscle type underscore the importance of repressing the transcription of non-cardiac myofibril isoforms in the developing heart through normal CHD4 function.
Here, we demonstrate that in Chd4 cardiac null hearts, the atypical expression of skeletal and smooth muscle myofibril isoforms interferes with the proper formation of adjacent sarcomeres into a symmetrical array. This work further supports the argument that misexpression of non-cardiac myofibril isoforms and sarcomere disorganization in developing cardiomyocytes results in inadequate heart contraction and embryonic lethality, thus presenting an essential role for repression of these alternative muscle subunits during cardiac development.
Generation of CHD4 Conditional Knockout Mice
Female Chd4fl/fl mice were crossed with male Chd4fl/+;Nkx2-5Cre/+ mice to generate the
control, (Chd4fl/fl), and CHD4 cardiac null, (Chd4fl/fl;Nkx2-5Cre/+) embryos. Male Chd4fl/ +;TgTnnt2cre mice were crossed to female Chd4fl/fl mice as an independent
Cre-recombinase driver. (Jackson Laboratory No: 024240)
Genotyping
Mice were genotyped using PCR. Chd4 F primer seq: 5' TCCAGAAGAAGACGGCAGAT 3' and R primer seq: 5'
GTCCTTCCAAGAGAGAGCAAG 3'. Cre recombinase F primer seq: 5'
GTCCCCTTCTCCATCTCCAG 3' and R primer seq: 5' GCAAACGGACAGAAGCATTT 3'. PCR results were resolved using gel electrophoresis in a 1% agarose gel with a 1 kb+ ladder (Invitrogen).
Tissue Preparation
At stage E10.5, embryos were fixed in 4% PFA for 16 hours. They were then washed in 15% Sucrose/1X PBS overnight followed by a wash in 30% sucrose/1X PBS overnight to increase tissue density. Embryos were then embedded in TissueTek OCT media and stored at -80°C.
Embryos were sectioned using a CryoStar NX70 cryostat (n=3 per genotype) at a thickness of 10 μm. Antigen retrieval was performed in sodium citrate buffer (10 mM, 0.05% Tween 20, pH 6) for 20 minutes for antigen retrieval and blocked for 1 hour in 10% fetal bovine serum (FBS), 1% TritonX-100, and 1X phosphate buffered saline (PBS). Sections were incubated with primary antibody overnight at 4°C in wash buffer of 1% FBS, 0.1% TritonX-100, and 1X PBS. Sections were incubated in secondary
antibodies for 30 minutes at room temperature and nuclei were stained using DAPI-PBS (200 ng/mL). Primary antibodies used include sarcomeric α-actinin (Sigma A7732), tropomyosin (DSHB CH1), smooth muscle myosin heavy chain (Abcam ab53219), and fast skeletal troponin I2 (Abcam ab183508). Secondary antibodies used include a goat α-mouse IgG (H+L) 488 (Molecular Probes A11001), a goat α-mouse IgG1 546 (MP
A21123), and a goat α-rabbit IgG (H+L) 647 (MP A21245).
Imaging and Image Analysis
Sections were imaged using an Olympus BX61 microscope with the DAPI, TxRed, and Cy5 fluorescent channels at 4X and 40X. Images were analyzed in FIJI (NIH). Three 40X regions from each ventricle were analyzed for a measure of sarcomeric
disorganization. A vector identification system was used (n=25 per 40X region in three different embryos of each genotype) and all vectors were classified as outlined in Ang Y-S et al., 2011.17 Sections were also imaged using a Zeiss 700 confocal microscope
Statistics
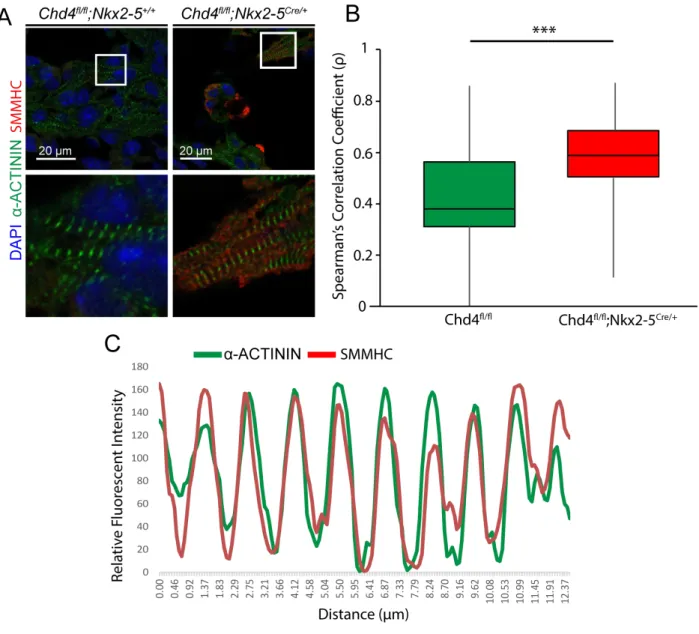
A Chi-squared test was performed on the sarcomere classifications for each ventricle of the control and CHD4 cardiac null embryos. To determine intercalation, a Spearman’s correlation test was used on ranked fluorescent values between α-ACTININ and
SMMHC expression along the length of each sarcomere. A p-value was obtained using a two-tailed Student’s t-test.
Results
CHD4 is required for repression of non-cardiac myofibril isoforms
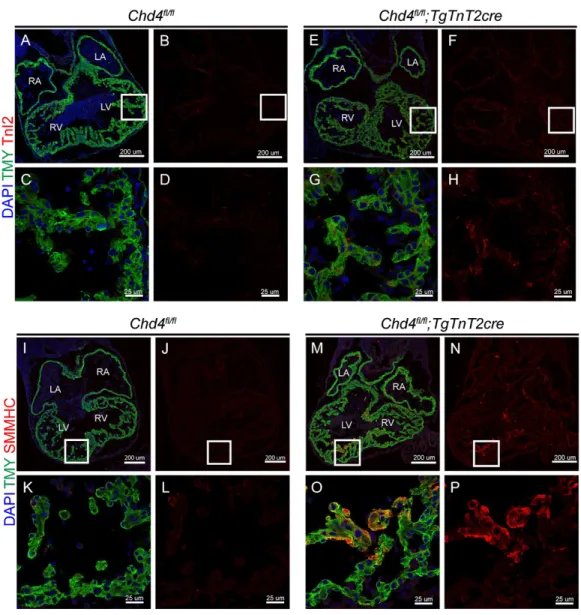
Figure 1. CHD4 is required to repress the expression of fast skeletal Troponin I2 (fsTnI2) and smooth muscle myosin heavy chain (SMMHC) in the myocardium of the developing heart at (E)10.5. a-h, High diffuse expression of fsTnI2 throughout the myocardium in the CHD4 conditional knockout (Chd4fl/fl;Nkx2-5Cre/+) (e-h) as compared
to a wild-type (WT, Chd4fl/fl) control (a-d). i-p, High, localized expression of SMMHC in
regions of the myocardium in the Chd4 conditional knockout (i-l) as compared to a WT control (m-p).
Ablation of Chd4 is Sufficient to Disrupt Normal Cardiac Sarcomere Formation
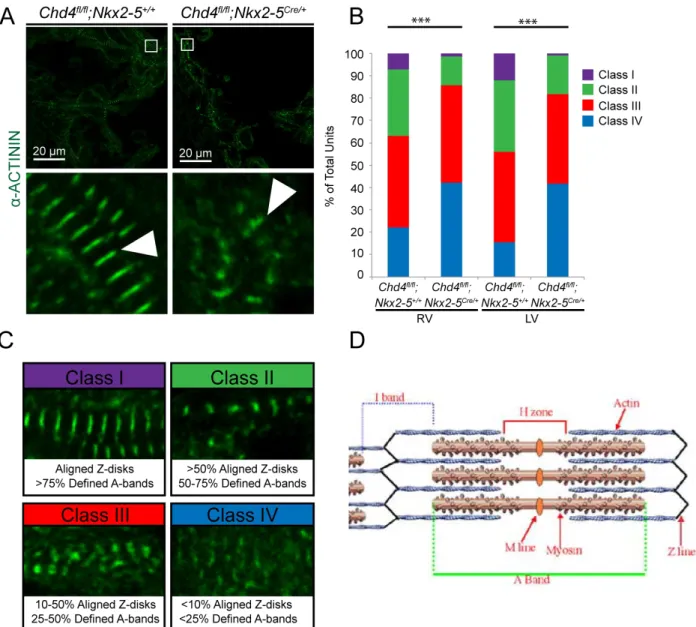
fluorescently labeling α-actinin, an essential structural protein found in the Z-line of sarcomeres (Figure 2A). For an embryo of each genotype, we conducted three independent experiments and analyzed 3 regions in each ventricle by drawing 25 vectors that provided a representative sample of that region. Sarcomere disorganization was scored using a classification system based on Z-disks alignment and A-band definition (Figure 2C,D). Chd4 cardiac null embryos show a greater degree of
sarcomere disorganization compared to controls as qualitatively characterized by Z-disk alignment and A-line visibility (Figure 2A). Quantitatively, Chd4 cardiac null embryos showed a significant increase in the number of disorganized sarcomeres when
Figure 2. Repression of non-cardiac myofibril isoforms is required for the proper development and patterning of cardiac sarcomeres. a, Loss of CHD4 disrupts normal sarcomere organization, visualized through the fluorescent labeling of α-ACTININ in the Z-line. b, Significant changes in the categorization of cardiac sarcomeres between the WT control and Chd4 conditional knockout in the right and left ventricles, (n=3
Misexpressed Smooth Muscle Myosin Heavy Chain Intercalates into Nascent Cardiac Sarcomeres
Discussion
The NuRD complex is known to interact with cardiac transcription factors in
transcriptional repression.5,6 Consequently, we have shown that the NuRD complex
Acknowledgements
I would like to thank Carrie Wilczewski and Dr. Frank Conlon for their support and guidance throughout the entirety of my project. I would also like to thank Haley Davies for her assistance with experiments and the generation of ideas. I would also like to express my gratitude towards Pablo, Kristen, and Vicky at MSL for their training and assistance in obtaining my confocal images.
References
1. Lipshultz SE, et al. (2013) Pediatric cardiomyopathies: causes, epidemiology, clinical course, preventative strategies and therapies. Future Cardiology 9(6):817-848.
2. Morita H, et al. (2008) Shared Genetic Causes of Cardiac Hypertrophy in Children and Adults. New England Journal of Medicine 358(18):1899-1908.
3. Kamisago M, et al. (2000) Mutations in Sarcomere Protein Genes as a Cause of Dilated Cardiomyopathy. New England Journal of Medicine 343(23):1688-1696. 4. Tayal U, Prasad S, & Cook SA (2017) Genetics and genomics of dilated cardiomyopathy and systolic heart failure. Genome Medicine 9(1):20.
5. Kaltenbrun E, et al. (2013) A Gro/TLE-NuRD corepressor complex facilitates Tbx20-dependent transcriptional repression. J Proteome Res 12:5395-5409.
8. Garnatz AS, et al. (2014) FOG-2 Mediated Recruitment of the NuRD Complex Regulates Cardiomyocyte Proliferation during Heart Development. Development Biology 395:50-61.
9. Allen HF, Wade PA, & Kutateladze TG (2013) The NuRD architecture. Cell Molecular Life Sciences 70(19):3513-3524.
10. Bouazoune K, et al. (2002) The dMi-2 chromodomains are DNA binding modules important for ATP-dependent nucleosome mobilization. Journal of Embryology 21:2430-2440.
11. Gómez-del Arco P, et al. (2016) The Chromatin Remodeling Complex Chd4/NuRD Controls Striated Muscle Identity and Metabolic Homeostasis. Cell Metabolism 23:881-892.
12. Gregorio CC & Antin PB (2000) To the heart of myofibril assembly. Trends in Cell Biology 10:355-362.
13. Yang Z, et al. (2009) Differences between Cardiac and Skeletal Troponin Interaction with the Thin Filament Probed by Troponin Exchange in Skeletal Myofibrils. Biophysical Journal 97(1): 183-194.
14. Craig R & Woodhead JL (2006) Structure and function of myosin filaments. Current opinion in structural biology 16(2): 204-212.
15. Walklate J, Ujfalusi Z, & Geeves MA (2016) Myosin isoforms and the