Copyright 0 1988 by the Genetics Society of America
Mutational Analysis of the Open Reading Frames in the Transposable
Element IS1
Michael Jakowec,* Pierre Prentki,* Michael Chandler? and David J. Galas*
*Department of Molecular Biology, University of Southern Calvornia, Los Angeles, Calijornia 90089-1340, and ?Centre de
Recherche de Biochimie et Ginitique Cellulaires du Centre National de la Recherche Scientijique, 31062 Toulouse Cedex, France
Manuscript received December 1, 1987 Revised copy accepted June 1, 1988
ABSTRACT
IS1 is one of the smallest transposable elements found in bacteria (768 bp). It contains eight overlapping open-reading-frames (ORFs) greater than 50 codons, designated insA to insG and insB’. To determine which of the ORFs actually code for proteins involved in transposition, we have introduced amber codons into each ORF by site-directed mutagenesis which make neutral changes in the overlapping ORFs. Each mutant IS1 was then tested for its ability to mediate cointegrate formation in Su+ and Su- backgrounds. The mutant elements were also tested for trans-complementation in an IS1 -free Salmonella background. Our results show that the products of the insA and insB genes are the only ones essential for cointegrate formation. We suggest that other ORFs may, however, encode accessory proteins.
T
HE insertion sequence ISI is a small transposableelement (768 bp) which can both transpose and
induce plasmid fusions (cointegrates) (CHANDLER and
GALAS 1984). ISI can also form compound transposon
in which an otherwise nontransposable DNA segment
is flanked by two copies of ISI in opposite orientation. Understanding the molecular mechanisms by which ISI transposes requires the identification of the pro- teins involved in this process. Like several insertion sequences, notably IS2, IS4, IS5 and IS903 [for re- view, see RAK and VON REUTERN ( 1 984), GALAS and
SMITH (1984); also D. J. GALAS and M. CHANDLER
(unpublished data)], ISI has several overlapping read- ing frames (ORFs) on both strands, potentially capable of encoding proteins greater than 50 amino acids in length (an arbitrary cutoff). The eight ORFs in ISI are designated insA, B, B’, C, D, E, F and G. Using insertions, deletions, and chimeric DNA constructions between ISI s from other enteric bacteria with variant sequences, as well as mutagenesis, to disrupt the
ORFs, MACHIDA et al. ( 1 982) and MACHIDA, MACHIDA
and OHTSUBO (1984) concluded that insA and insB
are essential for cointegrate formation, and that all others are not. However, because of multiple muta- tions within each ORF and some of the overlapping
ORFs, only the conclusion that insA encodes an essen-
tial product appears unambiguous.
T o investigate the possible role of each of the po- tential encoded polypeptides in cointegrate formation, we interrupted each of the eight ORFs with amber codons. T h e mutations were designed to affect one ORF at a time, leaving all (or most) other overlapping
ORFs unaltered with respect to amino acid coding.
This set of mutant ISIs were tested for their ability
Genetics 1 2 0 47-55 (September, 1988)
to form cointegrates in Escherichia coli in the absence and in the presence of amber suppressors. Since many E. coli strains carry chromosomal copies of IS2 (NY-
MAN et al. 1981; DEONIER 1987), the mutants were
also examined in a Salmonella typhimurium strain which lacks chromosomal copies of ISI, to rule out possible transcomplementation effects of the genomic copies.
Our results identify the reading frames insA and
insB as specifying the only ISI-encoded proteins es-
sential for cointegrate formation in E. coli. In addition, we found that nonsense mutations in insE (and per- haps insG) reduce cointegrate formation in Salmo- nella, suggesting that one or both of these may specify a protein whose function is accessory to the transpo- sition process, and can be provided in trans from the chromosome.
MATERIALS AND METHODS
Bacteria: The bacterial strains used in this study and their sources are shown in Table 1. The plasmids pBR322 (SUT-
CLIFFE 1978, PEDEN 1983); pZ152 (ZACURSKY and BERMAN
1984); pTC72.1, pTC72.2, pTC72.3 and pTC72.24
(PRENTKI et al. 1986) were previously described. The con- struction of the artificial transposon (“Qon”) is described in PRENTKI et al. 1987. All strains were grown in LB Broth and plated on LB Agar (MILLER, 1972). Media were supple- mented, where needed, with tetracycline (25 gg/ml), kana- mycin (50 Pglrnl), streptomycin (100 Pg/ml), spectinomycin (25 Pg/ml) and ampicillin (50 gg/ml).
TABLE 1 Bacterial strains
M. JakoM
Strains Genotype Source
E. coli
XAC F’/(Alac pro) ara nalA thi MILLER (1972)
str’ riff” argEam sup-
XA102 F’/(Alac pro) ara metB nalA MILLER (1972) riff” argEam thi supE (Su2)
XA103 F’/(Alac pro) ara metB nalA MILLER (1972)
riff“ argEam thi supF (Su3)
BW313 dut- ung- F’/lysA SACHER and STRAUSS (1983) HBlOl hsdS20 recA13 rpsL20(strA) Laboratory
collection
S. typhimurium
RMlO thyA deo pSLT- (no F-inhib- MAUER et al.
~~~~ ~
iting plasmid) (1984)
plasmid carrying the KmR marker (CHANDLER and GALAS 1983), were crossed with a suitable recipient, selecting for transfer of KmR (conjugation frequency) or KmR
+
TcR (IS1mediated cointegrates). The ratio of KmR + TcR/KmR was used to determine the frequency of cointegrates in the population.
DNA methods: Small and large scale plasmid DNA prep- arations were obtained using the alkaline lysis (MANIATIS, FRITSCH and SAMBROOK 1982) and “cleared lysate” (CLEW- ELL and HELINSKI 1969) methods, respectively. Restriction endonucleases were purchased from New England Biolabs or Boehringer Mannheim, and used according to the man- ufacturers’ recommendations. Transformations of plasmid DNA into E. coli (MANDEL and HIGA 1970) and Salmonella (LEDERBERG and COHEN 1974) were carried out as de- scribed. Oligonucleotides for site-directed mutagenesis and for DNA sequencing were synthesized on an Applied Bio- systems 381A DNA Synthesizer. DNA sequencing was car- ried out using the chain termination method of SANGER, for double-stranded DNA templates (ZAGURSKY et al. 1985).
Plasmid construction: The vector used as the template for mutagenesis (pMJ1) was constructed as follows: plasmid pTC72.24, a pBR322 derivative carrying an insertion of
IS1 at position 4212 (PRENTKI et al. 1986), was digested with EcoRI and ScaI. The smaller fragment containing IS1
was purified and ligated to the larger fragment of a ScaI-
EcoRI digest of pZ152 which carries the M13 origin of replication (ZAGURSKY and BERMAN 1984). The resulting plasmid contained the M13 origin and a copy of IS1 (Figure
The plasmid vector pMJ4 is a derivative of plasmid pTc72.24 (see above) in which the IS1 is rendered inactive due to site-directed mutations at both the left-end (GG to CT) and the right-end (CC to GA) of the inverted repeats. This plasmid is unable to mediate cointegrates (Table 3).
The plasmid pMJ25 was constructed to determine the frequency of IS1 transposition. The artificial transposon (“Qon”) (PRENTKI et al. 1987) containing the ends of IS1
flanking the StrR/SpcR antibiotic marker, was inserted into the EcoRl site of pMJ4 (see above). This element is able to transpose when complemented in cis by IS1 at a frequency comparable to that of wild-type IS1 (Table 5 ) .
Site-directedmutagenesis: Single-stranded template DNA was isolated by infection of plasmid-carrying strains with M13 phage as described (ZAGURSKY and BERMAN 1984). Single-strand plasmid DNA was prepared in E. coli BW3 13 (Ung- Dut-) to permit the incorporation of uracil residues 1 )-
iec al.
in the DNA (KUNKEL 1985). The mutagenesis protocol using synthetic oligonucleotides (of length 24-28 nucleotides) (KUNKEL 1985) yielded mutants at frequencies from 15 to 90% of the transformants. The occurrence of second-site mutations during the mutagenesis procedure could pose serious problems for the interpretation of our results. Therefore, we have attempted to assess the frequency of such mutations and taken measures to assure the validity of our results. Following the polymerization-ligation step in the mutagenesis of pMJl, an aliquot of the DNA was trans- formed into HBlOl. Several hundred TcR colonies were selected and then screened for sensitivity to ampicillin. The overall average rate of Ap5 among the TcR colonies was about 1.8%. We estimate that the frequency of mutations in the @-lactamase gene could be as high as 4-6%. Since this gene is roughly the size of IS1 this seems to be a reasonable estimate for the rate of second-site mutations in IS]. The region immediately surrounding the directed mutation (50- 75 nucleotides) was sequenced in each case and we found no nondirected changes. T o minimize the likelihood of an undetected mutation affecting our results, we used at least two, independent, sequenced mutants to assay the effects of the mutation, thus making it very unlikely that we could be observing an effect of a second site mutation. All independ- ent isolates behaved identically in each case.
RESULTS
T h e potential role of IS2 ORFs in encoding poly- peptides required for cointegrate formation was in- vestigated by interrupting each of the eight ORFs with amber codons. T h e mutations left most overlap- ping reading frames’ coding capacity unaffected (Ta- ble 2). T h e parent plasmid used in all mutagenesis experiments was pMJl (Figure 1).
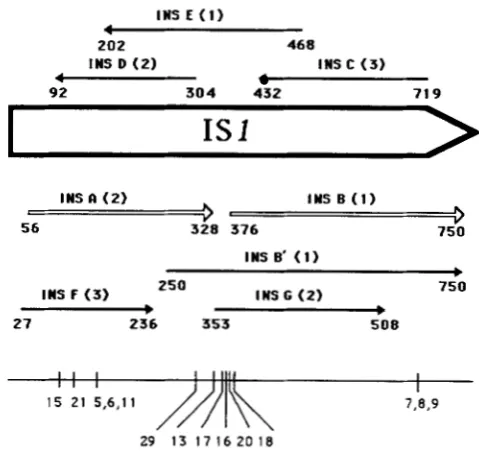
InsA: The ORF designated insA (Figure 2) begins with the codon GTG at position 56 of ISI, terminates with TAA at position 332 in frame 2, and can code for a protein of 92 amino acids. An amber mutation was created in insA by changing C to T at position
179 in a Gln codon. Simultaneously, an A to G trans- ition destroying a unique PstI site and creating a new PvuII site was introduced at position 175 (Figure 3) to allow screening of potential mutants by restriction endonuclease analysis. (The mutations were con- firmed by DNA sequencing. T h e high proportion of mutants obtained (30-go%), however, allowed us to identify them directly by sequencing in subsequent experiments.) Both of the insA mutations in this plas- mid designated pMJ5, leave the overlapping ORF insD unaltered with respect to amino acid coding. TO rule o u t effects on gene expression or cointegrate formation not directly related to the amber codon, a neutral mutation was made which leaves both insA and insD unaltered in amino acid coding. This was made by changing A to G at position 175 (the site of the amber mutation in pMJ5) to generate pMJ6.
IS1 Open Reading Frames 49
TABLE 2
Effect of directed mutations on each ORF
ORF Affected
Plasmid
(pMJ) A B C D E F G B’ A-B inter
5 am * N-S
21 am I-L I-s
6 * * N -S
7 am
*
8
9
*
am28 am
*
** *
11
*
am C-Y13
*
am v-L+
15
*
H-Y am16 N-Y am
*
+
17 F-L K-R am
+
18 * am
+
20 N-L S-R am
+
29
*
*
am~~ ~~~ ~~~
The plasmids are indicated by their number (omitting the pMJ for brevity), the ORFs are indicated by the single letter designation as shown in Figure 2. The type of mutation produced in the ORF is indicated as follows: am = amber mutation,
*
= a neutral codon change (same amino acid as wild type, but different codon), capital= a missense mutation which changes the encoded amino acid from the first (wild type) to the second indicated amino acid (single letter amino acid code).
+
(in last column) = mutations which affect the region between insA and insB.oriM13 I S 1
* * 1
i ; R pMJ25
FIGURE 1 .-Plasmid structures. Plasmid pMJi is a derivative of pBR322 carrying an insertion of IS1 at coordinate 4212 in pTC72.24 and a 455 bp fragment with the origin of replication of phage M13. Plasmid pMJ4 is a derivative of pMJl in which both ends of IS1 have been inactivated by mutations in their first 2 bp (asterisks); pMJ25 carries the artificial IS1 transposable element “Ron” (PRENTKI et al. 1987) in the EcoRI site of pMJ4.
The plasmids pMJ5, pMJ6, and pMJ2l were trans- formed into E. coli strains XAC (Su-), XA102 ( S U ~ ) , and XAlO3 (Su3) and the target plasmid, pOX38Km, was crossed into each strain. The cointegrate forma- tion frequency of each mutant IS2 was then deter- mined by the “mating out” assay (see MATERIALS AND
METHODS). The results are shown in Table 3. Both insAam mutations (pMJ5 and pMJ21) reduced the ability of IS2 to mediate cointegrate formation to a frequency of less than
7
X lo-’ cointegrates per transconjugant in XAC (Su-). Wild-type IS2 (pMJ1)INS E ( 1 ) 4
202 468
INS 0 (2)
-
INS C (3)9 2 3 0 4
4 3 2 71 9
IS
1
INS fi (2) INS B ( 1 )
56 328
3,
376 750INS 6’ ( 1 )
b INS F ( 3 ) 250 INS C ( 2 ) 750
b
27
-
236 353 508I 1 I I I I I I I
I 1 I I
15 21 5,6,11 7B,9
29 13 I 7 16 20 18
FIGURE 2.-The open reading frames of ISI. The arrows indi- cate the ORFs of IS1 consisting of greater than 50 amino acid residues, and point in the direction of transcription. The numbers at the ends of the arrows indicate the IS1 coordinates of the extremities of the ORF. The reading frames are indicated in parentheses. Frame 1 starts with coordinate 1. The positions of the mutations discussed in this paper are indicated at the bottom of the Figure.
produced cointegrates at a 100-fold higher frequency (6 X 10”) in the same background while both pBR322 and pMJ4 (containing an IS2 inactivated by mutations at the ends of each inverted repeat; see MATERIALS AND METHODS and Figure 3) did not generate cointe- grates at a detectable frequency (<4 X 1 O-’, the lower limit of detection). The ability of pMJ5 to mediate cointegrate formation was restored to approximately 10% of the wild-type level (Table 3) in Su2 (insertion of Glu) or Su3 (insertion of Tyr) strains. This result indicates that the amber codon at position 179 in insA disrupts a protein-coding reading frame essential for cointegrate formation and that InsA is functional with either Tyr or Gln at this position. This is not the case with the amber mutation carried by pMJ21; the Su3 (Tyr) suppressor restores the wild-type protein se- quence and also the ability to form cointegrates while the Su2 (Gln) suppressor does not. This result indi- cates that the Tyr residue specified by the codon at position 145 is important to the structure and/or function of the InsA protein. The results with pMJ5 and pMJ21 also imply that InsA cannot be provided in trans from genomic copies of ISZ.
The neutral codon changes in both insA and insD (pMJ6) not only fail to reduce cointegrate formation in the Su- background but actually increase its fre- quency (Table 3) relative to wild-type IS2 by sixfold.
ZnsB: InsB begins with an ATG codon at position
M. Jakowec et al.
I N S F- ALA I L L SER 4 I N S A- ARG TYR LEU
+
CGCTATCTCTG GCGATAGAGAC
+ALA ILE, GLU ”INS D
”-
D
WILD TYPE
I N S A
-
GLN LEU GLNWILD TYPE
A
CAACTGCAG
GTTGACGTC
t LEU GLN LEU ”INS D
”-
I N S A - G L N LEU STOP
C LEU CLN LEU
-
I N S D_”
I I
175 179
I N S F
-
ALA SER SER+
”
-INS A- ” ARG STOP
+
CGCTAGCTCTG
GCGATCGAGAC
c
I
”-
+ALA LEU GLU- I N S D
1 45 INS A AMBER
p M J 5 INS A AMBER
p M J 2 1
I N S A
-
GLN LEU GLN 4I N S A- ARG STOP ”3
TCGGTAACCTCG AGCCATTGGAGC
”- + ARG TYR GLY
-
INS E EWILD TYPE INS D AMBER
p M J l 1
C STOP LEU - I N S D
- -
I I
175 178
I N S A- ARG STOP
STOP GLY SER ”INS E
I
328 I N S A- GLN LEU GLN 4
”-t LEU GLN LEU
-
I N S DI
175
INS E AMBER p M J I 3
INS A/D NEUTRAL p M J 6
I N S F- TRP TRP cvc
GGGTGGTGCG
CCCACCACGCINS B
-
ARG TYR THR jCGATATACG GCTATATGC
”-
+
TYR VAL-
INS C”
WILD TYPE
B
WILD TYPE
t- HIS HIS
-
INS D ”INS B
-
ARG STOP+
INS F AMBER p M J 15
INS B AMBER
p M J 7 + TVR HIS
-
I N S D”
I
107
t TYR
”
lYAL
-
INS6 1 2
INS B - E TYR THR 4
+ STOP VAL
-
INS C”
I
609INS 8’- ILE VAL
*
I N S G
-
SEP SEP SEEAGTAGCAGA TCATCGTCT
+-
ASP ASP ”INS E“
INS C AMBER
p M J 9 F
WILD TYPE
INS B\C NEUTRAL I N S 8- ARC TYR THR 4
p M J 8 INS 8’- I L L YAL 4
INS 6- SER STOP 4
”
t- TVR ASP “INS E ”
I
363
t TYR YAL
-
I N S C”
I
609
INS G AMBER p M J l 6
C 322 343
”.) inSA 7 36 1 373
,-)
i n s BAAACTCAGGCCG ~AGTCGGTAACCTCGCGCATA~AGCCGGGCAGTGACGTC~TCGTCTGCGCG~AAATGGACGAA
1
L
T
T-
T T T A G T A G ptlJ29 ptlJ17 ptlJ20 pMJlB
IS1 Open Reading Frames 5 1
TABLE 3
Activities of alleles of IS1
(suppressor background) Cointegrate frequency
Plasmid Mutation su- su2 su3
pBR322 <0.05 (4) <0.01 (4) CO.01 (4) pMJl None 1 .OO (8) 1 .OO (6) 1 .OO (4) pMJ4 Inactive ends <0.05 (2) <0.003 (2) C0.004 (2) pMJ5 A amber <0.012 (4) 0.059 (5) 0.102 (6)
pMJ2 1 ( 1 79) <0.010 (6) <0.001 (6) 0.180 (6) pMJ6 A amber 6.43 (3) 2.24 (3) 1.81 (3) pMJ7 (145) <0.010 (10) CO.001 (12) 0.219 (10) pMj8 A/D neutral
(175) (61 3) (609) B amber B/C neutral pMJ9 C amber pMJl1 (609) pMJ 13 D amber pMJ15 (175) pMJ16 E amber pMJl7 (328) pMJ 18 F amber pMJ20 ( 1 07) pMJ29 G amber
B’ amber B’ amber B’ amber B‘ amber 1363) (343) (373) (361) (322)
1.45 (4)
3.22 (6) 6.40 (6) 1.22 (3) 0.72 (3) 1.77 (3) 0.28 (4) 0.05 (4) 0.15 (4) 0.10 (3)
0.619 (6) 2.29 (2)
0.638 (4) 1.2 1 (4) 1.05 (6) 1.49 (3) 0.124 (4) 0.171 (3) 0.235 (3) 2.47 (3) 1.66 (2) 0.87 (3) 0.056 (4) 0.092 (3) 0.051 (3) 0.007 (4) 0.013 (4) 0.019 (4) 0.0 19 (3) 0.182 (3)
Logarithmic means of multiple measurements (the number of which is indicated in parentheses after each entry) were normalized within each strain against pMJ 1 (wild-type IS1 which has cointegrate frequencies of 0.60, 9.05, and 7.14 in the Su-, Su2, and Su3 backgrounds, respectively). The numbers in parentheses after the mutation designations are the coordinates (relative to the left end o f ISI) of the base-pair changed in each mutation (the first base- pair changed if more than one was altered). Cointegrate formation assay is described in MATERIALS A N D METHODS.
(Figure 2). An amber mutation was created by chang- ing T to G at position 6 12, at Tyr codon, (pMJ7). T h e change is neutral with respect to amino acid coding in the overlapping ORF insC (opposite in direction to insB). A second mutation in insB was made by chang- ing A to G at position 609 (pMJ8) which is neutral in its amino acid coding effects on both insB and insC (Table 2).
ISI s containing an insBam mutation were unable to mediate cointegrate formation in the Su- background (Table 3). This ability was restored for pMJ7 in the Su3 background to a level approximately 20% that of wild type. Suppression by Su3 (Tyr) restores the wild- type protein sequence. No restoration of cointegrate forming activity was observed in the Su2 (Gln) back- ground. T h e amber mutations examined here clearly
disrupt a required sequence in which the Tyr residue plays an important role. However, since insB is also part of a larger ORF, insB’ (Figure l), it was necessary to examine the possibility that insB’ rather than insB encodes the required protein.
ZnsB’: The ORF insB’ is a 5‘ extension of insB
which overlaps insA (Figure 2). It begins with the codon ATG at position 250, terminates at position 75 1 with TAA, and may encode a protein of 167 amino acids. Four amber codons were introduced separately into the amino terminal segment of the insB‘ reading frame, three within the “intergenic re- gion” between insA and insB and a fourth in the overlapping region of insA and insB’ but not affecting insA. T h e mutations are, in order: pMJ29, a C to T transition at position 322 within a Gln codon; pMJ17, a C to T transition at position 343 within a Gln codon; pMJ20, replacement of the Ile codon ATC by TAG at position 361-3; and pMJl8, replacement of a Gln codon GAA (at position 373-5) by TAG (Figure 2). In the absence of suppressors, all insB’am mutants mediated cointegrate formation at a reduced fre- quency but significantly higher than that mediated by the insA and insB amber mutants (Table 3). A re- duced frequency was also observed in the Su2 and Su3 backgrounds. Several of the resulting cointe- grates were confirmed by restriction analysis (data not shown).
T h e results of these experiments shown that InsB’ is not an essential protein for cointegrate formation. T h e mechanism leading to the observed reduction in cointegrate formation by the insB’ amber mutations is as yet unclear (see DISCUSSION).
Other ORFs: IS2 has five additional ORFs (other
than insA, insB and insB’) which may encode proteins of length greater than 50 amino acids. These ORFs were designated in&, D, E, F and insG (Figure 1). None of the amber mutants in these ORFs eliminated the ability of IS1 to mediate cointegrate formation in the Su- E. coli background (Table 3) indicating that these ORFs do not produce a protein essential to cointegrate formation in E. coli.
Mutations in insC and insD showed an increase in cointegrate formation of at least fourfold compared to wild-type ISI. T h e reason for this is not clear. It is possible that insC and insD produce proteins that act to reduce the observed cointegrate populations, as has been suggested (MACHATTIE and JACKOWSKI 1977; REIF and ARBER 1980; BIEL and BERG 1984; BRAEDT 1985; AHMED 1986; BRAEDT 1988). Alterations in codon usage in insA, regulation of insA or insB expression, or regulation of another ISI protein could also account for this effect.
M. Jakowec
TABLE 4
Effect of suppression on cointegrate frequency
Strain (frequency X 10‘) Plasmid su- s u 3 s u 2
PMJ~ 0.60 (8) 9.05 (6) 7.14 (4) pTC72.24 0.18 (3) 0.53 (4) 1.14 (3) pTC72.3 0.32 (6) 0.33 (7) 0.22 (3) pTC72.1 0.22 (2) 0.72 (2) 0.46 (3) pTC72.2 0.69 (2) 0.53 (3) 0.93 (3) Using the suppressor-carrying strains shown in Table 1 , the cointegrate frequency was measured for the four ISI-carrying plas- mids discussed in the text. The entries are the logarithmic means of multiple measurements. The numbers in parentheses indicate the number of independent assays.
TABLE 5
Transposition and cointegrate frequency in E. coli and
S. typhimurium
Transposition Cointegration ( S F R ) (Spc’+TcR)
E. coli HBlOl 1.66 X (6) 3.94 X (6) S. typhimurium 5.60 X (5) 1.31 X (6) The frequencies are expressed as logarithmic means. The num- ber of independent assays is indicated in parentheses.
ited by the wild type ISI, suggesting a role of insE in the suppressor effect (see below).
Since the apparent, nonessential nature of these ORFs could simply be due to trans complementation of the mutant alleles from genomic copies of ISI present in E . coli K12, cointegrate formation was examined in an ISI-free background (see below).
Cointegrate formation in
S.
typhimurium: South-ern blotting using an ISI-specific probe demonstrated that S. typhimurium strain RMlO has no resident ISI (data not shown).
T o ensure that ISI acts in Salmonella as it does in
E. coli, we compared the frequency of both cointegrate formation and transposition of wild-type ISI in these strains. The plasmid pMJ25 (Figure l), which carries an ISI inactive for transposition due to mutant in- verted repeats (see MATERIALS AND METHODS) and the
artificial transposon “Qon” (PRENTKI et al. 1987) con- taining synthetic inverted repeats of ISI flanking the StrR/SpcR antibiotic marker and the target plasmid pOX38Km were introduced into the S. typhimurium
strain RM 10 and the frequency of both transposition (SpcR
+
KmR) and the cointegrate formation (SpcR+
T c R+
KmR) was determined by the mating-out assay using E . coli HBlOl R i p NalR. When complementedin cis by ISI the “Qon” is able to mediate both coin- tegrate formation and transposition at frequencies similar to wild-type ISI (Table 4).
Each plasmid containing ISI amber mutations and the target plasmid pOXKm38 were introduced into strain RMlO and the cointegrate formation was de- termined as described above (Table 6 ) . Wild-type ISI
TABLE 6
Cointegrate formation in S. typhimurium
Relative
frequency
pBR322 co.0 12
Plasmid Mutation IS1 cointegrate
(6)
PMJ~ None 1
.oo
(4)
pMJ4 IR-L and IR-R C0.015
(2)
pMJ5 A amber C0.015
(3)
pMJ7 B amber c0.021
(3)
pMJ9 C amber 2.94
(14)
PMJ~ 1 D amber 1.18
(2)
PMJ~ 3 E amber 0.01 5
(7)
pMJ15 F amber 0.97
(4)
pMJ16 G amber 0.22
(2)
pMJ18 B’ amber 0.50
(4)
The frequency of cointegrates in the population is expressed as the logarithmic mean of multiple measurements and normalized against wild type ISI on pMJl (3.3 X cointegrates/transcon- jugant in the population). The number of independent assays is
shown in parentheses.
(pMJ 1) showed a cointegrate frequency of 3.25 X 10” cointegrates per transconjugant in this strain. The inactive IS2 with altered inverted repeats (pMJ4) did not mediate cointegrates at a detectable level (<5 X
1 O-’ in the population). This represents the detection limit of the assay. The relative activities of the mutants were essentially the same as in E . coli Su- strain, with two exceptions. Although mutations in insE and insG
had no effect on cointegrate formation in E. coli, they induce strong reductions in S. typhimurium. This sug- gests that these functions are not essential but do stimulate transposition and can be provided in trans
from genomic copies of IS1 carried by the E . coli host. While the insE amber mutation (pMJ13) does not affect insG, the insG amber mutation (pMJ16) also introduces a missense mutation in insE. The present data, while suggesting that both insE and insG may make accessory (nonessential) proteins, are also con- sistant with the hypothesis that insE alone encodes such a protein. Since an indirect effect of the muta- tions on gene expression in Salmonella cannot be ruled out, these influences cannot be considered firm at the moment.
Increased cointegrate formation frequency in s u p
pressor background: A comparison of the activities
IS1 Open Reading Frames 53
(Table 4): a 1 O-20-fold increase in frequency of coin- tegrate formation in the Su2 and Su3 background was seen compared to the Su- background. T o determine whether the phenomena is due to the presence of the M13 origin of replication in pMJ1, the behavior of the parent plasmid, pTC72.24, was investigated. This plasmid displays a reduced frequency of cointegrate formation in the Su- strain and, although some en- hancement is observed in the Su+ strains, its magni- tude is significantly reduced compared to pMJl (Table
4). Plasmid pTC72.3, with IS2 inserted at the same position as pTC72.24 but in the opposite orientation, does not show enhanced cointegrate formation in the suppressor background. Plasmid pTC72.1 (IS2 in- serted at position 4177 of pBR322) also exhibits a small suppressor enhancement while pTC72.2 (IS2 inserted at the same site but opposite orientation as pTC72.1) does not (Table 4). These results suggest that the suppressor effect may be amplified by the presence of the phage M 13 origin of replication by an unknown mechanism, and is also affected by the ori- entation of IS1 in pBR322. The Su+ and Su- cells harboring the ISI-containing plasmids were examined for an increase in plasmid dimers which could effect the cointegrate frequency. No dimers were observed in either strain background (data not shown).
DISCUSSION
We have created amber stop codons in each of the eight ISI ORFs. In at least one instance for each ORF, all overlapping ORFs were left unaltered with respect to amino acid coding. The mating-out assay was used to analyze the effect of each IS2 mutant in both Su+ and Su- E . coli backgrounds. We show that two cis- acting proteins encoded by the ORFs insA and insB are essential for ISI-mediated cointegrate formation. A third, probably trans-acting, protein encoded by the insE ORF, may act as an accessory protein.
We have shown that insA and insB are essential for cointegrate formation. Previous work by MACHIDA et al. (1 982) used insertions and deletions to disrupt the ORFs within IS2 to conclude that insA and insB are essential. Their study, however, did not rule out the possible disruption of ORFs overlapping insA and insB which may encode essential or accessory proteins or elimination of essential sequences not encoding proteins in deletion mutants. A subsequent study (MACHIDA et al. 1984) employed chimeric DNA con- structions between IS2s with variant sequences from other enteric bacteria, as well as mutagenesis to intro- duce mutations into the various ORFs. An IS2 carry- ing an amber within insA was only active in a s u p pressor strain, showing that this ORF does encode an essential product. However, analysis of the other
ORFs (including insB) was ambiguous because of the
multiple mutations within each ORF and some of the overlapping ORFs.
Amber codons at positions 145 (Tyr) or 179 (Gln) of insA reduced the cointegrate frequency to an un- detectable level. Suppression of the amber at position 179 in Su2 and Su3 restored cointegrate forming ability to a level 10% that of wild-type IS1 in the same background. This indicates that the amber mutation disrupts a sequence that encodes a polypeptide and that InsA protein is able to function with either Gln or Tyr at this position. The amber at position 145 was suppressed by Su3 (Tyr) but not Su2 (Gln), indicating the importance of this Tyr residue to the function of InsA protein. The Tyr at position 145 is part of a region of InsA protein which has some similarity to the catalytic site of E. coli DNA gyrase. In gyrase, the
Tyr residue forms a covalent bond with DNA as an intermediate in the catalytic pathway (HOROWITZ and WANC 1987). If the analogous Tyr residue in InsA protein plays a similar role, changing it to Gln may eliminate this active-site catalytic function and destroy InsA proteins’ ability to mediate cointegrate forma- tion. The Gln-substituted InsA protein is indeed in- active.
InsA protein has been overproduced using a
T7
expression vector and has been shown to bind specif- ically to the ends of IS1 (ZERBIB et al. 1987). The InsA protein sequence shows the presence of a helix-turn- helix motif, characteristic of DNA binding proteins (PAW and SAUER 1984), near the carboxy terminus. Site-directed mutagenesis of this region of InsA pro- tein is currently underway to identify the sequence- specific binding determinants.
ZnsB encodes a protein essential for cointegrate forming ability since an amber at position 612 (Tyr)
was suppressed by Su3 (Tyr) but not Su2 (Gln). Ge- nomic copies of IS2 do not complement ambers in either insA or insB indicating that both proteins act in cis. This was also shown by the inability of genomic or plasmid bound IS2 encoded proteins to mediate transposition of an ‘ ‘ Q O ~ ” (artificial transposon) pre-
M. et
binding site (Shine-Dalgarno) preceding insB. This alteration may reduce the expression of InsB protein which was shown to be essential for cointegrate for- mation. An alternate explanation is that maximal expression of insB may require some translation through insB’ which is reduced by the amber muta- tions. T h e intergenic region between insA and insB contains extensive potential secondary structure which could play a role in the control of insB expres- sion. These mutations may also disrupt a transcription terminator identified in this region (PRENTKI et al.
1986; HUBNER, IIDA and ARBER 1987; S. Hollings- head, personal communication). In addition, the insA- insB intergenic region is spanned by the ORF insE which has not been shown to encode a protein but could possibly do so (see below). T h e intergenic region is probably involved in the control of gene expression in IS1 which makes the analysis of mutations in this region complicated.
ISI has five additional ORFs potentially capable of encoding proteins greater than 50 amino acids. In E .
coli, none of the amber mutations in in&, D, E, F, or insG significantly reduced the frequency of cointe- grate formation. However, not all mutations were completely neutral in their effect. In the Su- back- ground, a sixfold increase in cointegrate frequency compared to wild-type IS1 was seen with both the insA/insD neutral and the insD amber mutations and a threefold increase for the insC amber mutation. These mutations, like insB’, may exert their effect by altering expression of the essential proteins InsA and InsB by changing their codon usage, mRNA stability, or transcription or translation efficiency.
To rule out possible trans-complementation by ge- nomic copies of ISI, the activity of insC, D, E, F and insG amber mutants was examined in S. typhimurium which does not carry genomic copies of ISl. Wild- type IS1 transposition and cointegrate formation fre- quency was similar in both S. typhimurium and E . coli. In S. typhimurium, these ORF amber mutations, with the exception of those in insE and insG, did not show strongly reduced cointegrate forming ability. T h e insG amber mutation creates a missense in insE sug- gesting that insE, not insG, may encode a trans-acting accessory protein.
In E . coli Su+ cointegrate formation frequency in- creased 10-20-fold compared to Su-. A similar obser- vation was made by MACHIDA et al. (1 984); the reason is still unclear. Our results suggest that both the orientation of IS1 and the presence of the M 13 origin may account for this phenomenon. One possibility is that the anti-tet promoter of pBR322 (BROSIUS, CATE and PERLMUTTER 1982) may transcribe into IS1 and, in conjunction with the Su+ genotype, affect the expression of ZnsA, ZnsB, or another regulatory pro- tein. T h e insE amber mutation and several insB’
ambers (which cause missense mutations in insE) do not display the suppressor effect. This may suggest the possible existence of a functional polypeptide en- coded by the insE ORF.
We have identified the insA and insB gene products as the two essential ISI-encoded proteins which me- diate cointegrate formation. It is entirely possible that other ORFs encode proteins involved in some func- tion of ISI, but are not essential for cointegrate for- mation. While the InsA protein has been shown to bind specifically to the ends of ISI (ZERBIB et al. 1987), the full function of InsA protein is presently unknown and the function of the InsB protein is entirely un- known. Experiments designed to determine these functions can now proceed with the knowledge that no other IS1 proteins are required.
We wish to thank J. MILLER, R. J. ZAGURSKY and R. MAURER for bacteria strains, MINH-HA PHAM for excellent technical assist- ance, and GISELLE PETZINGER-JAKOWEC and BRUCE TETER for suggestions on the manuscript. We have benefitted from useful discussions of this work with M. SUSSKIND. This work was supported by National Institutes of Health grant A11936 to D. G. and a Natural Sciences and Engineering Research Council of Canada Postgraduate Fellowship to M. J.
L I T E R A T U R E C I T E D
AHMED, A., 1986 Evidence for replicative transposition of T n 5 and Tn9. J. Mol. Biol. 191: 75-84.
BIEL, S. W., and D. E. BERG, 1984 Mechanism of IS1 transposition in E. coli: choice between simple insertion and cointegration. Genetics 108: 319-330.
BRAEDT, G . , 1985 Recombination in recA cells between directed repeats of insertion sequence ISZ. J. Bacteriol. 162: 529-534. BRAEDT, G., 1988 Different reading frames are responsible for
ISZdependent deletion and recombination. Genetics 1 1 8
BROSIUS, J., R. L. CATE and A. P. PERLMUTTER, 1982 Precise location of two promoters for the fl-lactamase gene of pBR322. J. Biol. Chem. 257: 9205-9210.
CHANDLER, M., and D. J. GALAS, 1983 Cointegrate formation mediated by Tn9. 11. Activity of IS1 is modulated by external DNA sequences. J. Mol. Biol. 170: 61-91.
CHANDLER, M., and D. J. GALAS, 1984 Studies on the transposition of ISZ, pp. 53-77. In: Bacterial Plasmids Edited by D. HELINSKI, S. COHEN and D. CLEWELL. Plenum Press, New York.
CLEWELL, D., and D. HELINSKI, 1969 Supercoiled circular DNA- protein complex in Escherichia coli: induced conversion to an open circular DNA form. Proc. Natl. Acad. Sci. USA 62: 1 159-
1 166.
DEONIER, R. C., 1987 Location of native insertion sequence ele- ments. pp. 982-989. In: Escherichia coli and Salmonella typhi- murium, Edited by F. C. NEIDHARDT. American Society for Microbiology, Washington D.C.
GALAS, D. J., and M. CHANDLER, 1982 Structure and stability of Tn9 mediated cointegrates: Evidence for two pathways of transposition. J. Mol. Biol. 1 5 4 245-272.
GALAS, D. J., and T . F. SMITH, 1984 The relationship between codon boundries and multiple reading frame preferences: Cod- ing organization of bacterial insertion sequences. Mol. Biol. Evol. 1: 260-268.
HOROWITZ, D. S., and J. C. WANC, 1987 Mapping the active site tyrosine of Escherichia coli DNA gyrase. J. Biol. Chem. 262: 5339-5344.
IS1 Open Reading Frames 55
HUBNER, P., S. IIDA and W. ARBER, 1987 A transcriptional ter- minator sequence in the procaryotic transposable element ISI. Mol. Gen. Gen. 206 485-490.
KUNKEL, T. A., 1985 Rapid and efficient site specific mutagenesis without phenotypic selection. Proc. Natl. Acad. Sci. USA 82:
LEDERBERG, E. M., and S. N. COHEN, 1974 Transformation of
Salmonella typhimurium by plasmid deoxyribonucleic acid. J. Bacteriol. 1 1 9 1072-1074.
MACHATTIE, L. A., and J. B. JACKOWSKI, 1977 Physical structure and deletions effects of the chloramphenicol resistance element T n 9 in phage lambda. pp. 2 19-228. In: DNA Insertion Elements, Plasmids and Episomes, Edited by A. J. BUKHARI, J. A. SHAPIRO and S. L. ADHYA. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
MACHIDA, Y., C. MACHIDA and E. OHTSUBO, 1984 Insertion se- quence ISI encodes two structural genes required for its trans- position. J. Mol. Biol. 177: 229-245.
MACHIDA, Y., C. MACHIDA, H. OHTSUBO and E. OHTSUBO,
1982 Factors determining frequency of plasmid cointegration mediated by insertion sequence ISI. Proc. Natl. Acad. Sci. MANDEL, M., and A. HIGA, 1970 Calcium dependent bacterio-
phage DNA infection. J. Mol. Biol. 52: 154-162.
MANIATUS, T., E. F. FRISCH and J. SAMBROOK, 1982 Molecular Cloning. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
MAURER, R., B. C. OSMOND, E. SHEKHTMAN, A. WONG and D. BOTSTEIN, 1984 Functional interchangabilities of DNA rep- lication genes in Salmonella typhimurium and Escherichia coli
demonstrated by a general complementation procedure. Ge- netics 108: 1-23.
MILLER, J. H., 1972 Experiments in Molecular Genetics. Cold Spring
N Y M A N , K., K. NAKAMURA, H. OHTSUBO and E. OHTSUBO,
488-492.
USA 7 9 277-281.
Harbor Laboratory, Cold Spring Harbor, N.Y.
1981 Distribution of the insertion sequence ISI in Gram- negative bacteria. Nature 3 8 9 609-6 12.
PABO, C. O., and R. T. SAUER, 1984 Protein-DNA recognition. Annu. Rev. Biochem. 53: 293-321.
PEDEN, K. W. C., 1983 Revised sequence of the tetracycline- resistance gene of pBR322. Gene 2 2 275-280.
PRENTKI, P., B. TETER, M. CHANDLER and D. GALAS,
1986 Functional promoters created by the insertion of trans- posable element ISI. J. Mol. Biol. 191: 383-393.
PRENTKI, P., M. H. PHAM, P. GAMAS, M. CHANDLER and D. J. GALAS, 1987 Artificial transposable elements in the study of the ends of insertion sequence ISI. Gene 61: 91-101.
RAK, B., and M. VON REUTERN, 1984 Insertion element IS5 contains a third gene. EMBO J. 3: 807-8 1 1.
REIF, H. J., and W. ARBER, 1980 Analysis of transposition of
I S I h n and its relatives. Cold Spring Harbor Symp. Quant. Biol. 45: 40-43.
SANGER, D., and B. STRAUSS, 1983 Insertion of nucleotides o p posite apurinic/apyrimidinic sites in deoxyribonucleic acid dur- ing in vitro synthesis: uniqueness of adenine nucleotides. Bio- chemistry 22: 4518-4526.
SUTCLIFFE, J. G., 1978 Complete nucleotide sequence of the Esch- erichia colt plasmid pBR322. Cold Spring Harbor Symp. Quant. Biol. 43: 77-90.
ZAGURSKY, R. J., and M. L. BERMAN, 1984 Cloning vectors that yield high levels of single-stranded DNA for rapid sequencing. Gene 27: 183-191.
ZAGURSKY, R. J., K. BAUMEISTER, N. LOMAX and M. L. BERMAN,
1985 Rapid and easy sequencing of large linear double stranded DNA and supercoiled plasmid DNA. Gene Anal. Techn. 2: 89-94.
ZERBIB, D., M. JAKOWEC, P. PRENTKI, D. J. GALAS and M. CHAN- DLER, 1987 Expression of proteins essential for IS1 transpo- sition: Specific binding of InsA to the ends of ISI. EMBO J. 6
3163-3169.