0095-1137/07/$08.00⫹0 doi:10.1128/JCM.01012-06
Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Clonal Complexes of
Campylobacter jejuni
Identified by Multilocus
Sequence Typing Are Reliably Predicted by Restriction Fragment
Length Polymorphism Analyses of the
flaA
Gene
䌤
Steven P. Djordjevic,
1* Leanne E. Unicomb,
2Penelope J. Adamson,
3† Lance Mickan,
4Rosa Rios,
5and the Australian Campylobacter Subtyping Study Group‡
Department of Primary Industries, Elizabeth Macarthur Agricultural Institute, Camden, New South Wales,1OzFoodNet,
Hunter New England Population Health, Wallsend, New South Wales, and the National Centre for Epidemiology and Population Health, Australian National University, Canberra,2Flinders Medical Centre, Bedford Park,
South Australia,3Institute of Medical and Veterinary Science, Adelaide, South Australia,4and
Microbiological Diagnostic Unit, Parkville, Victoria,5Australia
Received 15 May 2006/Returned for modification 11 July 2006/Accepted 7 October 2006
Multilocus sequence typing (MLST) has provided important new insights into the population structure of
Campylobacter jejuniand is rapidly becoming the gold standard for typing this species. However, the
method-ology is comparatively costly and slow to perform for the routine surveillance testing of large numbers of isolates required by public health laboratories. Restriction fragment length polymorphism analysis of theflaA gene (RFLP-flaA) and sequencing of the variable region in theflalocus (SVR-fla) were compared to MLST to determine if a low cost alternative could be found that reliably predicts clonal lineage (as determined by MLST). An isolate ofC. jejunifrom each of 153 patients from New South Wales, Australia, collected sequen-tially over a period of 30 months from 1999 to 2001 and comprising 40 sequence types (ST) from 15 clonal complexes (CC) was examined. Of 15 CC, 12 were represented by more than one isolate and a predominant RFLP-flaAtype was found for 10 (83%). Of these, seven (70%) correctly predicted the predominant MLST CC with a probability of >0.8. Of 40 STs detected, 19 were reported for the first time, 9 of which were represented by more than one isolate. Eight of these were represented by a single RFLP-flaAtype. Only two of eight major SVR-flatypes were able to predict CC with a probability of >0.8, indicating thatflaA-RFLP is a more reliable predictor of CC than SVR-flaand thus offers an alternative to MLST for use in routine surveillance.
Despite being significantly underreported,Campylobacter in-fections represent one of the most common causes of acute bacterial enteritis in humans worldwide (33). In Australia, Campylobacteris the most commonly notified pathogen (19), and the number of cases per year has continued to increase since notifications were first recorded (Communicable Dis-eases Australia, NNDSS notifiable disease data 2006; http: //www1.health.gov.au/cda/Source/CDA-index.cfm).
Efforts have recently focused on determining important risk factors for Campylobacter infection to guide interventions aimed at reducing disease burden. Such risk factors are com-monly determined for other pathogens through investigations
of outbreaks; however, despite the large number of Campy-lobacternotifications, outbreaks are rarely detected. Case con-trol studies to determine risk factors for infection have iden-tified consumption of chicken, exposure to animals, and consumption of contaminated water as significant (1, 4, 16). A meaningful typing system that could be applied to Campy-lobacterisolates as they arrive in the public health laboratory could aid outbreak detection and help identify common sources of infection.
Numerous typing strategies, including pulsed field gel elec-trophoresis (PFGE), PCR-restriction fragment length poly-morphism analysis of flagellin genes (RFLP-fla), sequencing of
* Corresponding author. Mailing address: Department of Primary Industries, Elizabeth Macarthur Agricultural Institute, Camden, New South Wales, Australia. Phone: 246406426. Fax: 11 61 246406384. E-mail: [email protected].
† Present address: Academic Unit of Molecular Vascular Medicine, The LIGHT Laboratories, Clarendon Way, University of Leeds, Leeds LS2 9JT, United Kingdom.
‡ S.P.D., L.E.U., P.J.A., L.M., and R.R. are members of the Aus-tralian Campylobacter Subtyping Study Group. Other members (listed in alphabetical order) include Penny Adamson (Flinders Medical Cen-tre, South Australia), Kellie Cheung (Institute of Clinical Pathology and Medical Research, Westmead, New South Wales), Barry Combs (Department of Human Services, Adelaide, South Australia), Craig Dalton (Hunter New England Population Health, Newcastle, New South Wales), Robyn Doyle (Institute of Medical and Veterinary Sci-ence, Adelaide, South Australia), John Ferguson (Hunter New En-gland Health Service, Newcastle, New South Wales), Lyn Gilbert (In-stitute of Clinical Pathology and Medical Research, Westmead, New
South Wales), Rod Givney (Department of Human Services, Adelaide, South Australia), David Gordon (Flinders Medical Centre, Bedford Park, South Australia), Joy Gregory (Department of Human Services, Melbourne, Victoria), Geoff Hogg (Microbiological Diagnostic Unit, University of Melbourne, Parkville, Victoria), Tim Inglis (Division of Microbiology & Infectious Diseases, PathWest, Nedlands, Western Aus-tralia), Peter Jelfs (Institute of Clinical Pathology and Medical Research, Westmead, New South Wales), Martyn Kirk (OzFoodNet, Canberra, Australian Capital Territory), Karin Lalor (Department of Human Ser-vices, Melbourne, Victoria), Jan Lanser (Institute of Clinical Pathology and Medical Research, Westmead, New South Wales), Lyn O’Reilly (Di-vision of Microbiology & Infectious Diseases, PathWest, Nedlands, West-ern Australia), Minda Sarna (Department of Health, Perth, WestWest-ern Australia), Hemant Sharma (Hunter New England Health Service, New-castle New South Wales), Helen Smith (Queensland Health Scientific Services, Coopers Plains, Queensland), and Mary Valcanis (Microbiolog-ical Diagnostic Unit, University of Melbourne, Parkville, Victoria).
䌤Published ahead of print on 8 November 2006.
102
on May 16, 2020 by guest
http://jcm.asm.org/
the short variable region of theflalocus (SVR-fla), ribotyping, multilocus enzyme electrophoresis, multilocus sequence typing (MLST), randomly amplified polymorphic DNA, and ampli-fied fragment length polymorphism have been employed to examine epidemiological relationships between isolates within the species Campylobacter (reference 35 and references therein). PFGE and amplified fragment length polymorphism are understood to provide excellent discrimination between isolates ofCampylobacter, and PFGE is often regarded as the “gold standard” for tracing outbreaks (9, 10, 27). MLST cata-logues sequence variation among genes encoding proteins that are essential for metabolic function (so-called “housekeeping genes”), defining each isolate in terms of its sequence type (ST), which then can be grouped into clonal complexes (CCs) that represent lineages presumed to derive from a common ancestor (6). MLST studies of Campylobacter jejuni demon-strate that the organism is genetically diverse, predominantly as a result of frequent intra- and interspecies recombination, within a weakly clonal population structure (7, 16, 31). It is increasingly evident that the CC, as defined by MLST, is an epidemiologically relevant unit for both long- and short-term investigations of C. jejuni epidemiology (16). Furthermore, MLST has been successfully used to compare populations ofC. jejunifrom veterinary, human, and environmental sources and has highlighted meat-producing practices as reservoirs for pathogenic campylobacters that infect humans as well as iden-tification of species-restricted clones (6, 16). MLST is highly reproducible, portable, and easy to interpret, has an interna-tional nomenclature, and thus, is rapidly becoming the method of choice for large-scale molecular epidemiological studies of C. jejuniand otherCampylobacterspp.
Despite its many advantages, MLST is expensive to perform, and the equipment and expertise required to interpret data are not readily available to many clinical and veterinary laborato-ries that would be required to type a large number of isolates on an on-going basis. To more frequently identify outbreaks, to clarify infection routes, and to refine case control study anal-yses, a practical yet meaningful alternative to MLST is re-quired. Previous studies suggest that RFLPs withinflaAgene sequences (flaA-RFLP) alone provide sufficient discrimination for its use as a subtyping method forC. jejuni and Campy-lobacter coli(10). Despite the observation that recombination rates inC. jejunimay potentially have an adverse impact on the reliable interpretation of flaA-RFLP data (13, 34), several studies show that the majority ofC. jejuniandC. coliisolates are genetically stable over long periods (2, 10, 11, 23, 25) and that susceptibility to genome rearrangement may be strain specific. These observations are largely consistent with data from large-scale comparative genomic studies that were com-pared to the C. jejuni NCTC11168 genome sequence (26) showing that significant regions of the C. jejunigenome are stable (32) but that as much as 21% of genes in NCTC11168 may be dispensable (8). More recently, an interlaboratory eval-uation of three flagellin PCR/RFLP methods for typing C. jejuni and C. colishowed that analysis of the full flaAgene using the restriction endonuclease DdeI was an appropriate method for standardization, as demonstrated by 100% inter-laboratory reproducibility (12). Collectively, these studies sug-gest that RFLP-flaAis a discriminatory, reliable, and relatively inexpensive typing tool forCampylobacter. However,
longer-term studies require typing systems that can be used by mul-tiple laboratories to systematically examine the epidemiology ofCampylobacterinfection and detect outbreaks and common sources of sporadic infection.
The aim of this study was to determine whether RFLP-flaA typing could be a useful and cost-effective alternative to MLST for potential use in routine surveillance ofCampylobacter iso-lates. To best assess RFLP-flaA typing as an alternative to other more expensive and comparatively technically demand-ing methodologies, we used isolates that were collected se-quentially over a 30-month period from a public laboratory that would reflect isolates from mandatoryCampylobacter no-tifications. RFLP-flaAtyping was compared to MLST for con-gruence and for the ability to predict MLST-derived clonal complexes. The time period studied also offered the opportu-nity to explore the stability of theflaAlocus and determine whether this is a limitation of the method for longer term epidemiological studies.
MATERIALS AND METHODS
Study population.A case control study was conducted in the Hunter Health Area of New South Wales, Australia, between January 1999 and July 2001. Two
pathology laboratories supplied reports on subjects withCampylobacterdetected
in stool samples, and a total of 318 patients were recruited. A total of 240
Campylobacterisolates were collected from the cases identified through one laboratory, and 171 of the isolates were stored and remained viable.
Molecular and biochemical tests used to confirm species identity.Isolates from diarrheal stool cultures were stored and subsequently tested for species by hippurate hydrolysis and PCR as described previously (3, 29). Template DNA for PCR was prepared using Instagene matrix as outlined in the manufacturer’s instructions (Bio-Rad, California). PCR amplifications were performed in a
50-l volume containing 10 mM Tris-HCl (pH 8.3), 1.5 mM MgCl2, 50 mM KCl,
200M concentrations of each deoxynucleoside triphosphate (dNTP), 0.2M
concentrations of each primer, 1.25 U ofTaqDNA polymerase (Roche
Diag-nostics, Castle Hill, Australia), and 1l of Instagene prepared DNA. All PCR
experiments were performed on a PC-960G gradient thermal cycler or PC-960 and FTS thermal cyclers (Corbett Research, Australia), and the amplification products were analyzed on 1% agarose gels. This enabled classification of the
isolates as eitherC. jejunior non-C.jejuni. Of the 171 isolates, 153 were found
to beC. jejuni, and these isolates were included in this study.
Typing methods.MLST was performed according to the method of Dingle et al. (7), with modified amplification conditions (denaturation at 94°C [1 min], annealing at 50°C [1 min], and extension at 72°C [1 min]) as described by Mickan et al. (18). Sequences were assigned allele numbers and the isolates were as-signed their sequence types by interrogation of the Campylobacter MLST data-base (http://campylobacter.mlst.net/).
RFLP-flaAwas undertaken by digestion of the PCR product with a single
enzyme (DdeI) (RFLP-flaA[s]) based on the method described previously (20) or
with two restriction enzymes (PstI and EcoRI) (RFLP-flaA[d]) according to the
method of Wassenaar and Newell (35). Typing by determination of the gene
sequence of theflaAgene short variable region (SVR-fla, base positions 145 to
600) was performed according to the method described by Meinersmann et al. (17). Sequences were assigned a type by submission to the website http: //outbreak.ceid.ox.ac.uk/campylobacter.
Riboprinting (automated ribotyping [RT]), pulsed field gel electrophoresis (PFGE), and Laboratory of Enteric Pathogens (LEP) serotyping were also ap-plied to isolates, as described by O’Reilly et al. (24).
Data analysis. (i) Typing assignment.All experimental data were imported
into a BioNumerics database. The gel images from RFLP-flaA(s) and
RFLP-flaA(d) analyses were imported as raw TIFF images. The BioNumerics software
was used to analyze banding patterns using the Dice band matching coefficient and unweighted-pair group method using average linkages with a position
tol-erance of 1% and an optimization of 1%. The RFLP-flaA(s) and RFLP-flaA(d)
patterns were grouped into subtypes when the banding patterns were visually indistinguishable.
(ii) Comparison of methods.Methods were compared for congruence. To
compare MLST with RFLP-flaA(s), a predominant RFLP-flaA(s) type was
as-signed, defined as the RFLP-flaA(s) type representingⱖ50% of the isolates
on May 16, 2020 by guest
http://jcm.asm.org/
within that CC (with⬎1 isolate) and/or whereⱖ50% of the ST with a CC were
represented by a predominant RFLP-flaA(s) type; the latter definition was
re-quired where a CC comprised several ST containing only a single isolate. The proportion of correct (successful) predictions of the predominant CC was
cal-culated for each of the major RFLP-flaA(s) types as (number of isolates of
predominant CC for RFLP-flaA[s]x)/(total number of isolates of RFLP-flaA[s]x).
Predominant SVR-flatypes were similarly assigned.
Statistical analysis.We aimed to measure the degree of congruence of
RFLP-flaA(s) and RFLP-flaA(d) using Cramer’s V statistic calculated using SPSS
ver-sion 12.0. This statistic assumes that the marginal differences in rows are similar to those for columns, an assumption that is unlikely to be met with these data but would result in a smaller V statistic. Furthermore, Cramer’s V assumes that the change from one type to another is equally likely. Despite these limitations, Cramer’s V determines an approximate measure of association between two methods with different levels of discrimination, which is likely to be conservative. The discrimination index (DI) was determined using the method of Hunter and Gaston (15).
RESULTS
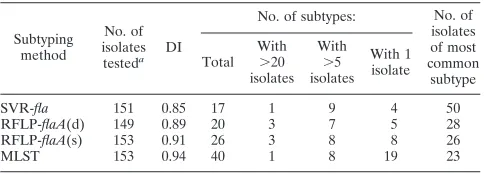
Comparison of ability of typing methods to detect STs.Of the 171 isolates collected, 153 were confirmed asC. jejuni, and these were examined using RFLP-flaA(s), RFLP-flaA(d), SVR-fla, and MLST. The distribution of subtypes using each method is given in Table 1. Of the four methods, MLST was the most discriminatory, identifying 40 ST (Table 1) which could be grouped into 14 clonal complexes (18) and having a DI of 0.94. There were 25 isolates represented by five ST for which a clonal complex could not be assigned (18). In an attempt to seek alternative methods to MLST, congruence (with MLST), ability to predict clonal complexes, and evidence of genetic stability were examined for flaA(s), RFLP-flaA(d), and SVR-fla. An expected high level of congruence was detected between RFLP-flaA(s) and RFLP-flaA(d) (V⫽ 0.84, P ⬍ 0.001); thus, RFLP-flaA(s) only was analyzed for congruence, ability to predict MLST CCs, and genetic stability.
Congruence of RFLP-flaAtyping and SVR-fla with MLST.
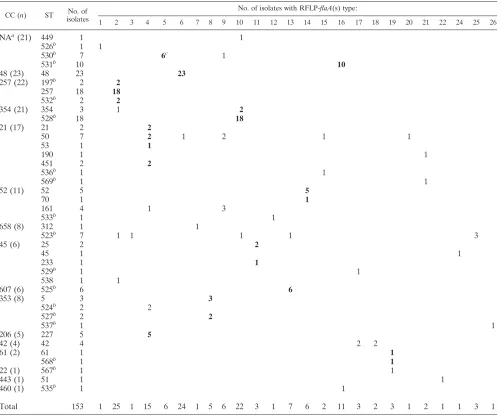
Of the 14 CCs, there were 12 with more than one isolate, and a predominant RFLP-flaA(s) type was found for 10 (83%) (Table 2). Within CCs, a high degree of correlation (ⱖ95%) between predominant RFLP-flaA(s) types was found, particu-larly for CCs 48, 257, and 354, the most common CCs (Table 2). More than one RFLP-flaA(s) type was detected for STs 530, 354, 50, 161, 523, and 42, with the greatest variation for ST 50. RFLP-flaA(s) types 2, 4, 6, 8, 9, 10, 13, 16, 17, and 19 were detected among more than one CC (Table 2).
Of the 40 STs detected among 153 isolates, there were 19 that had not previously been described (STs 197, 523 to 538, and 567 to 569) (18). Of these, 10 were represented by more than a single isolate, 8 of which (80%) were represented by a single RFLP-flaA(s) type (STs 197, 524, 525, 531, 532, and 535). Of the remaining two STs, 6 of 7 ST 530 isolates were represented by the predominant RFLP-flaA(s) type and 3 of 7 ST 523 isolates were represented by a predominant RFLP-flaA(s) type (Table 2).
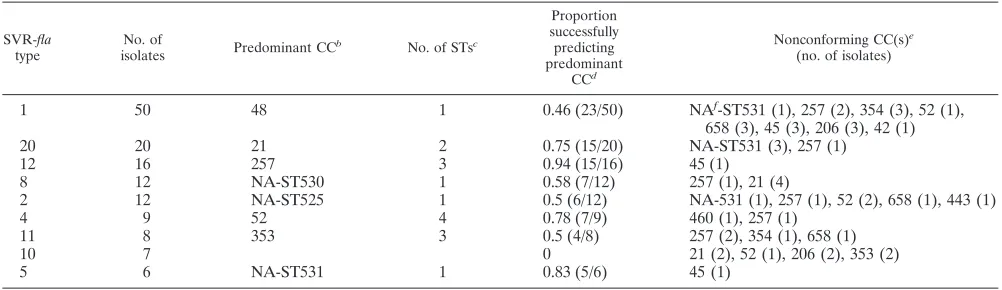
SVR-fla has been considered a candidate typing method with potential for public health application. A high degree of congruence was found for CCs 48, 354, and 257, and all CCs had a predominant SVR-flatype. More than one SVR-flatype was detected for eight STs (STs 531, 257, 354, 528, 50, 161, 523, and 42). Of the 10 newly described STs represented by more than one isolate, 6 possessed a single SVR-flatype (525, 530, 197, 532, 524, and 527).
Predicting MLST clonal complexes.The ability of the major RFLP-flaA(s) types to predict CCs is outlined in Table 3. There were nine RFLP-flaA(s) types which comprised at least five isolates [or nonassigned STs for 2 RFLP-flaA(s) types]. Of these, the proportion of correct predictions of the predominant CC ranged from 0.47 to 1.0. There were seven RFLP-flaA(s) types (78%) where the probability of predicting the predomi-nant CC was⬎0.8. Two RFLP-flaA(s) types (types 14 and 9) predicted the same CC (CC 52).
SVR-fla was also examined for its ability to predict CCs. There were only two (22%) of the nine major SVR-flatypes where the proportion that successfully predicted the predom-inant CC was⬎0.8 (Table 4).
Genetic stability of RFLP-flaA(s). The Campylobacter ge-nome is considered to be highly variable, with horizontal trans-fer of genetic material occurring frequently in some regions of the chromosome compared with regions displaying remarkable stability (8). Assuming isolates representing a single MLST ST are clonal, we defined genetic instability surrounding theflaA locus as the presence of two or more RFLP-flaA(s) types within an ST. Twelve of the 15 CCs identified in this study comprised more than one isolate, 10 of these had a predomi-nant RFLP-flaA(s) type and 6 (50%) possessed RFLP-flaA(s) types other than the predominant one (Table 2). Among these six CCs, three showed a difference in RFLP-flaA(s) that was consistent with a difference in ST (CCs 45, 52, and 353). To determine if isolates belonging within each of the remaining three CCs (CCs 21, 52, and 354) that possessed more than one RFLP-flaA(s) type had undergone recombination in regions of the genome outside theflaAlocus, we applied several widely used typing strategies (RT, PFGE, LEP serotyping) to examine their genetic relationships. For CC 21 (ST 50), 5 of 9 ST-50 isolates had a nonpredominant RFLP-flaA(s), and this dif-ference was also reflected in RFLP-flaA(d), RT, and PFGE (Table 5). For CC 52 (STs 161 and 533), differences in all methods were detected, with the exception of serotyping. For CC 354 (ST 354) only 1 of 21 isolates differed by RFLP-flaA(s), and this difference was also reflected by SVR-flaand RFLP-flaA(d) as well as PFGE; however, the RT result was consistent with the other 21 isolates (Table 5).
DISCUSSION
Public health surveillance requirements of a typing method for Campylobacter include the ability to (i) detect potential outbreaks during routine typing, (ii) distinguish isolates
repre-TABLE 1. Comparison of the frequency and distribution ofC.jejunisubtypes
Subtyping method
No. of isolates
testeda DI
No. of subtypes: No. of
isolates of most common
subtype Total
With
⬎20 isolates
With
⬎5 isolates
With 1 isolate
SVR-fla 151 0.85 17 1 9 4 50
RFLP-flaA(d) 149 0.89 20 3 7 5 28
RFLP-flaA(s) 153 0.91 26 3 8 8 26
MLST 153 0.94 40 1 8 19 23
aNumbers of isolates used to compared subtyping methods vary slightly
be-cause the occasional isolate failed to be revived when dispatched to the testing laboratory.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:3.585.301.543.89.176.2]sentative of a known outbreak from sporadic isolates, (iii) determine an outbreak vehicle (such as food or environmental sources), and (iv) detect common sources of infection using case control studies. Although MLST was originally applied to examine the population structure ofC. jejuni, numerous stud-ies have shown it to also have merit as a subtyping method for various Campylobacter spp. and it is a reliable predictor of clonality (7). It is also considered to be resilient to the effects of genetic instability (6) and has been successfully used for public health purposes to type outbreak-related compared to nonrelated Campylobacter isolates (5, 27). Routine public health surveillance ofCampylobacterrequires a high-through-put, low-cost method with a level of discrimination that is capable of distinguishing outbreak isolates (clonally related) from those occurring sporadically (5).C. jejuniundergoes fre-quent intra- and interspecies recombination, and typing
[image:4.585.44.546.80.497.2]strat-egies that are sensitive to changes in genome structure (inver-sions, duplications, deletions, and insertions) are affected by these events. PFGE is considered a gold standard typing tool for tracing outbreaks ofC. jejuni, and in this regard, it is highly discriminatory. However, our study shows that PFGE gener-ates multiple patterns for isolgener-ates residing within a CC and within STs (Table 5 and data not shown) and was not a reliable predictor of CC. Limitations of PFGE for the routine surveil-lance ofC. jejunihave been reported by others (14). Thus, the comparisons undertaken in this study did not aim to determine the most discriminatory method but instead examined whether two RFLP-flaAallele typing methods would offer an alterna-tive to MLST for routine public health surveillance purposes. Among the 153C. jejuni isolates used in our study, 40 ST were identified among 15 CC; a clonal complex could not be assigned to 19 isolates comprising 4 STs (18). Twelve of the 15 TABLE 2. Correlation of multilocus sequence typing clonal complexes with RFLP-flaA(s) types
CC (n) ST No. of
isolates
No. of isolates with RFLP-flaA(s) type:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 24 25 26
NAa(21) 449 1 1
526b 1 1
530b 7 6c 1
531b 10
10
48 (23) 48 23 23
257 (22) 197b 2
2
257 18 18
532b 2
2
354 (21) 354 3 1 2
528b 18
18
21 (17) 21 2 2
50 7 2 1 2 1 1
53 1 1
190 1 1
451 2 2
536b 1 1
569b 1 1
52 (11) 52 5 5
70 1 1
161 4 1 3
533b 1 1
658 (8) 312 1 1
523b 7 1 1 1 1 3
45 (6) 25 2 2
45 1 1
233 1 1
529b 1 1
538 1 1
607 (6) 525b 6 6
353 (8) 5 3 3
524b 2 2
527b 2
2
537b 1 1
206 (5) 227 5 5
42 (4) 42 4 2 2
61 (2) 61 1 1
568b 1 1
22 (1) 567b 1 1
443 (1) 51 1 1
460 (1) 535b 1 1
Total 153 1 25 1 15 6 24 1 5 6 22 3 1 7 6 2 11 3 2 3 1 2 1 1 3 1
a
NA, clonal complex unavailable (not yet assigned).
b
Sequence types only reported for Australian isolates, as of 14 March 2006.
c
The predominant RFLP-flaA(s) type for each clonal complex is designated by boldface type.
on May 16, 2020 by guest
http://jcm.asm.org/
CC were represented by more than one isolate, and a predom-inant RFLP-flaA type was found for 83%. Seven of the 10 (70%) RFLP-flaAtypes successfully predicted the predomi-nant CC⬎0.8 of the time. SVR-flawas examined as a potential predictor of CCs. A large proportion of isolates (33%) typed as SVR-fla type 1 and the finding that only 2 of 9 (22%) major SVR-fla types were able to predict CC with a probability of ⬎0.8 suggested that RFLP-flaAis a more reliable predictor of CC than SVR-fla.
Data presented here suggest that the lower-cost RFLP-flaA(s) is an alternative typing methodology that is capable of reliably predicting MLST CC. However RFLP-flaA(s) has its limitations. Most notable is the need for standardization and subsequent interpretation of resultant electrophoretic pat-terns, similar to that required for PFGE. Since completing this study, we have applied RFLP-flaA(s) to the typing of more than 600 clinical isolates ofC. jejunicollected from and typed in different regions of Australia. Standardization of the method appeared to have been achieved, as evidenced by
tection of five of the six predominant RFLP-flaA(s) types de-scribed in this study in all five states of Australia with the typing conducted in four laboratories (L. E. Unicomb, L. C. O’Reilly, M. D. Kirk, R. J. Stafford, H. V. Smith, N. G. Becker, M. S. Patel, G. L. Gilbert, and the Australian Campylobacter Subtyping Study Group, unpublished results). Our observa-tions are consistent with others who reported that RFLP-flaA(s) can be standardized with 100% interlaboratory repro-ducibility (12).
[image:5.585.43.543.81.216.2]The validity of RFLP-flaAhas been called into question due to significant levels of recombination that is a characteristic of theC. jejunipopulation structure. Despite this, RFLP-flaAhas been shown to be a reliable typing tool for epidemiological investigations, and groupings determined using this approach correlate well with PFGE, ribotyping, and random amplified polymorphic DNA typing strategies (10, 21, 22, 30). We exam-ined our data for evidence of recombination affecting theflaA locus. Assuming that isolates belonging to a particular ST are clonal, more than one RFLP-flaA(s) per ST was taken as an TABLE 3. Ability of the majoraRFLP-flaA(s) types to predict MLST CCs
RFLP-flaA(s)
type No. of isolates Predominant CC
b No. of STsc
Proportion successfully predicting predominant
CCd
Nonconforming CC(s)e
(no. of isolates)
2 25 257 4 0.88 (22/25) 354 (1), 658 (1), 45 (1)
6 24 48 1 0.96 (23/24) 21 (1)
10 22 354 2 0.91 (20/22) NAf-ST449 (1), 658 (1)
4 15 21 7 0.47 (7/15) 206 (5), 52 (1), 353 (2)
16 11 NA-ST530 1 0.91 (10/11) 460 (1)
13 7 607 1 0.86 (6/7) 658 (1)
14 6 52 4 1.0 (6/6)
9 6 52 0.5 (3/6) NA-ST530 (1), 21 (2)
8 5 353 3 1.0 (5/5)
a
RFLP-flaA(s) types with⬎5 isolates (n⫽121).
b
The most common CC within an RFLP-flaA(s) type.
c
Number of STs corresponding to CC.
d
Numbers given in parentheses are numbers of isolates of respective RFLP-flaA(s) type correlating with predominant CC/total.
e
The number of isolates for each CC is given in parentheses.
f
NA, not assigned.
TABLE 4. Ability of the majoraSVR-flatypes to predict MLST CCs
SVR-fla
type
No. of
isolates Predominant CC
b No. of STsc
Proportion successfully predicting predominant
CCd
Nonconforming CC(s)e
(no. of isolates)
1 50 48 1 0.46 (23/50) NAf-ST531 (1), 257 (2), 354 (3), 52 (1),
658 (3), 45 (3), 206 (3), 42 (1)
20 20 21 2 0.75 (15/20) NA-ST531 (3), 257 (1)
12 16 257 3 0.94 (15/16) 45 (1)
8 12 NA-ST530 1 0.58 (7/12) 257 (1), 21 (4)
2 12 NA-ST525 1 0.5 (6/12) NA-531 (1), 257 (1), 52 (2), 658 (1), 443 (1)
4 9 52 4 0.78 (7/9) 460 (1), 257 (1)
11 8 353 3 0.5 (4/8) 257 (2), 354 (1), 658 (1)
10 7 0 21 (2), 52 (1), 206 (2), 353 (2)
5 6 NA-ST531 1 0.83 (5/6) 45 (1)
aSVR-flatypes with⬎5 isolates (n⫽140).
bThe most common CC within an SVR-flatype.
cNumber of STs corresponding to CC.
dNumbers given in parentheses are numbers of isolates of respective RFLP-flaA(s) types correlating with predominant CC/total.
eThe number of isolates for each CC is given in parentheses.
fNA, not assigned.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:5.585.42.543.531.676.2]indication of possible genetic rearrangement, and this was identified for 12 of 121 isolates (10%). Although it is conceiv-able that point mutations may contribute to this variability, other typing methodologies (Table 5) suggested that recombi-nation was not confined to theflaAlocus and that RFLP-flaA is able to discriminate within a CC. In one MLST study ofC. jejuni, sequence variation due to recombination was estimated to be 50 times greater than that contributed by mutation (28). Despite this, genetically stable RFLP-flaA(s) types were de-tected [RFLP-flaA(s) types 14 and 8] throughout the study period of 30 months from one geographic region. Moreover,
a subsequent national study described above has detected 20 of the 26 RFLP-flaA(s) types described in this study in at least one of the five states included (Unicomb et al., unpub-lished results), demonstrating RFLP-flaA(s) stability over a period of up to 44 months, an observation consistent with other studies (10).
[image:6.585.45.539.79.552.2]While MLST is likely to remain the method of choice for distinction ofCampylobacter subtypes, until it can be under-taken a lower cost, using readily available equipment with minimal labor requirements, it is unlikely to be used for rou-tine public health surveillance. RFLP-flaA(s) typing offers a TABLE 5. Analysis ofC. jejuniisolates with RFLP-flaA(s) types noncongruent with predominant RFLP-flaA(s) MLST pairsa
CC
Identification of nonconforming
RFLP-flaA(s) type
ST
RFLP-flaA(s) type(s)
SVR-fla
type(s)
RFLP-flaA(d) type(s)
RT PFGE type(s) LEP serotype(s)b
257 Predominant
RFLP-flaA(s) type
197, 257, 532 2 1, 4, 8, 12 11, 17 15, 28 7, 31, 40, 41, 42, 64, 70 11, 13, 14, 31, 50
F128 527 8 11 15 4 8 18
F212 527 8 11 15 — 8 60
354 Predominant
RFLP-flaA(s) type
354 10 1, 11, 20 8, 13, 18 22 1, 30, 32, 48, 55, 60 18
F066 354 2 37 12 22 39 18
21 Predominant
RFLP-flaA(s) type
21, 50, 53, 451 4 1, 8, 10 5 5, 9, 10, 30 10, 61 UT, 1
F536 50 6 1 1 7 10 UT
F014 50 9 8 1 — 65 1
F251 50 9 8 1 7 6 UT
F239 50 15 — 5 — — —
F051 50 20 1 20 8 — UT
F304 190 21 1 — — — UT
F079 536 15 10 3 8 14 UT
F112 569 21 1 3 5 — UT
52 Predominant
RFLP-flaA(s) type
52, 70 14 4 7 11, 16 33, 34, 35, 37, 58 11, 13, 31
F025 161 4 10 5 12 45 11, 13, 31
F004c 161 9 4 3 2 44 13, 31
F451 161 9 2 3 24 46 11, 31
F501 161 9 2 3 — 46 31
F537c 533 12 1 15 21 27 UT
658 Predominant
RFLP-flaA(s) type
523 25 1, 71, 90 17 29 7, 56, 66 UT
F100 312 7 1 1 16 38 18
F509 523 2 71 11 22 21 UT
F044 523 3 1 4 23 20 UT
F270 523 10 11 8 22 18
F037 523 13 2 5 17 25 50
NA-ST530 Predominant
RFLP-flaA(s) type
530 5 8 5, 6 19, 20 26, 28 12, 23, 37
F459c 530 9 8 1 — 69 12, 37
45 Predominant
RFLP-flaA(s) type
25, 233 11 1 10 14 4, 59 UT
F377 45 24 5 2 13 19 27
F381c 529 17 9 16 13 52 UT
F458 538 2 12 11 — — 21
353 Predominant
RFLP-flaA(s) type
5 8 11 15 4 8, 62 3, 60
F055 524 4 10 5 6 2 37
F380 524 4 10 5 6 25 UT
F364 537 26 11 20 — — UT
42 Predominant
RFLP-flaA(s) type
42 17 9 16 25 3, 51 UT, 23
F063 42 18 9 5 25 10 UT
F064 42 18 1 13 25 22 UT
aBoldface type indicates typing differences from predominant RFLP-flaA(s) types. —, typing not performed.
bUT, untypeable strains.
cIsolates from infections acquired during travel overseas.
on May 16, 2020 by guest
http://jcm.asm.org/
medium- to high-throughput alternative to MLST for these purposes, but further subtyping would be required for some types, especially when outbreaks were suspected, and in these cases, methods such as MLST or PFGE could be considered. Finally, our data suggest that RFLP-flaA(s) not only reliably predicts MLST CC but is also capable of identifying isolates of C. jejunithat require further subtyping.
ACKNOWLEDGMENTS
This study was funded under the OzFoodNet program of work, which is an initiative of the Australian Government Department of Health and Ageing. Hunter New England Population Health is a unit of the Hunter New England Area Health Service, supported by fund-ing from NSW Health through the Hunter Medical Research Institute. We thank Wendy Forbes for excellent technical assistance in per-forming RFLP-flaA(d). We are also grateful for the assistance from Michael Hornitzky for providing expertise and equipment for the rou-tine culture ofC. jejuni.
REFERENCES
1.Abuoun, M., G. Manning, S. A. Cawthraw, A. Ridley, I. H. Ahmed, T. M. Wassenaar, and D. G. Newell.2005. Cytolethal distending toxin
(CDT)-negativeCampylobacter jejunistrains and anti-CDT neutralizing antibodies
are induced during human infection but not during colonization in chickens.
Infect. Immun.73:3053–3062.
2.Burnens, A. P., J. Wagner, H. Lior, J. Nicolet, and J. Frey.1995. Restriction fragment length polymorphisms among the flagellar genes of the Lior
heat-labile serogroup reference strains and field strains ofCampylobacter jejuni
andC. coli. Epidemiol. Infect.114:423–431.
3.Burnett, T. A., M. A. Hornitzky, P. Kuhnert, and S. P. Djordjevic.2002.
SpeciatingCampylobacter jejuniandCampylobacter coliisolates from poultry
and humans using six PCR-based assays. FEMS Microbiol. Lett.216:201–
209.
4.Butzler, J. P.2004.Campylobacter, from obscurity to celebrity. Clin.
Micro-biol. Infect.10:868–876.
5.Clark, C. G., L. Bryden, W. R. Cuff, P. L. Johnson, F. Jamieson, B. Ciebin, and G. Wang.2005. Use of the oxford multilocus sequence typing protocol and sequencing of the flagellin short variable region to characterize isolates
from a large outbreak of waterborneCampylobactersp. strains in Walkerton,
Ontario, Canada. J. Clin. Microbiol.43:2080–2091.
6.Dingle, K. E., F. M. Colles, R. Ure, J. A. Wagenaar, B. Duim, F. J. Bolton, A. J. Fox, D. R. Wareing, and M. C. Maiden.2002. Molecular
characteriza-tion ofCampylobacter jejuniclones: a basis for epidemiologic investigation.
Emerg. Infect. Dis.8:949–955.
7.Dingle, K. E., F. M. Colles, D. R. Wareing, R. Ure, A. J. Fox, F. E. Bolton, H. J. Bootsma, R. J. Willems, R. Urwin, and M. C. Maiden.2001. Multilocus
sequence typing system forCampylobacter jejuni. J. Clin. Microbiol.39:14–23.
8.Dorrell, N., J. A. Mangan, K. G. Laing, J. Hinds, D. Linton, H. Al-Ghusein, B. G. Barrell, J. Parkhill, N. G. Stoker, A. V. Karlyshev, P. D. Butcher, and B. W. Wren.2001. Whole genome comparison ofCampylobacter jejuni hu-man isolates using a low-cost microarray reveals extensive genetic diversity.
Genome Res.11:1706–1715.
9.Fitzgerald, C., L. O. Helsel, M. A. Nicholson, S. J. Olsen, D. L. Swerdlow, R. Flahart, J. Sexton, and P. I. Fields.2001. Evaluation of methods for
sub-typingCampylobacter jejuniduring an outbreak involving a food handler.
J. Clin. Microbiol.39:2386–2390.
10.Fitzgerald, C., K. Stanley, S. Andrew, and K. Jones.2001. Use of pulsed-field gel electrophoresis and flagellin gene typing in identifying clonal groups of
Campylobacter jejuniandCampylobacter coliin farm and clinical
environ-ments. Appl. Environ. Microbiol.67:1429–1436.
11.Hanninen, M. L., M. Hakkinen, and H. Rautelin.1999. Stability of related
human and chickenCampylobacter jejunigenotypes after passage through
chick intestine studied by pulsed-field gel electrophoresis. Appl. Environ.
Microbiol.65:2272–2275.
12.Harrington, C. S., L. Moran, A. M. Ridley, D. G. Newell, and R. H. Madden.
2003. Inter-laboratory evaluation of three flagellin PCR/RFLP methods for
typing Campylobacter jejuni and C. coli: the CAMPYNET experience.
J. Appl. Microbiol.95:1321–1333.
13.Harrington, C. S., F. M. Thomson-Carter, and P. E. Carter.1997. Evidence
for recombination in the flagellin locus ofCampylobacter jejuni: implications
for the flagellin gene typing scheme. J. Clin. Microbiol.35:2386–2392.
14.Hedberg, C. W., K. E. Smith, J. M. Besser, D. J. Boxrud, T. W. Hennessy, J. B. Bender, F. A. Anderson, and M. T. Osterholm.2001. Limitations of
pulsed-field gel electrophoresis for the routine surveillance ofCampylobacter
infections. J. Infect. Dis.184:242–244.
15.Hunter, P. R., and M. A. Gaston.1988. Numerical index of the discrimina-tory ability of typing systems: an application of Simpson’s index of diversity.
J. Clin. Microbiol.26:2465–2466.
16.Manning, G., C. G. Dowson, M. C. Bagnall, I. H. Ahmed, M. West, and D. G. Newell.2003. Multilocus sequence typing for comparison of veterinary and
human isolates ofCampylobacter jejuni. Appl. Environ. Microbiol.69:6370–
6379.
17.Meinersmann, R. J., L. O. Helsel, P. I. Fields, and K. L. Hiett.1997.
Discrimination of Campylobacter jejuni isolates by fla gene sequencing.
J. Clin. Microbiol.35:2810–2814.
18.Mickan, L., R. Doyle, M. Valcanis, K. E. Dingle, L. Unicomb, J. Lanser, and the Australian Campylobacter Subtyping Study Group.Multilocus sequence
typing ofCampylobacter jejuniisolates from New South Wales, Australia.
J. Appl. Microbiol., in press.
19.Miller, M., P. Roche, K. Yohannes, J. Spencer, M. Bartlett, J. Brotherton, J. Hutchinson, M. Kirk, A. McDonald, and C. Vadjic.2005. Australia’s notifi-able diseases status, 2003 annual report of the National Notifinotifi-able Diseases
Surveillance System. Commun. Dis. Intell.29:1–61.
20.Nachamkin, I., K. Bohachick, and C. M. Patton.1993. Flagellin gene typing ofCampylobacter jejuniby restriction fragment length polymorphism
analy-sis. J. Clin. Microbiol.31:1531–1536.
21.Nachamkin, I., H. Ung, and C. M. Patton.1996. Analysis of HL and O
serotypes ofCampylobacterstrains by the flagellin gene typing system. J. Clin.
Microbiol.34:277–281.
22.Nielsen, E. M., J. Engberg, V. Fussing, L. Petersen, C. H. Brogren, and S. L.
On.2000. Evaluation of phenotypic and genotypic methods for subtyping
Campylobacter jejuniisolates from humans, poultry, and cattle. J. Clin.
Mi-crobiol.38:3800–3810.
23.On, S. L., E. M. Nielsen, J. Engberg, and M. Madsen.1998. Validity of
SmaI-defined genotypes ofCampylobacter jejuniexamined by SalI, KpnI, and
BamHI polymorphisms: evidence of identical clones infecting humans,
poul-try, and cattle. Epidemiol. Infect.120:231–237.
24.O’Reilly, L. C., T. J. J. Inglis, L. Unicomb, and the Australian Campy-lobacter Subtyping Study Group.2006. Australian multicentre comparison
of subtyping methods for the investigation ofCampylobacterinfection.
Epi-demiol. Infect.134:768–779.
25.Owen, R. J., K. Sutherland, C. Fitzgerald, J. Gibson, P. Borman, and J. Stanley.1995. Molecular subtyping scheme for serotypes HS1 and HS4 of
Campylobacter jejuni. J. Clin. Microbiol.33:872–877.
26.Parkhill, J., B. W. Wren, K. Mungall, J. M. Ketley, C. Churcher, D. Basham, T. Chillingworth, R. M. Davies, T. Feltwell, S. Holroyd, K. Jagels, A. V. Karlyshev, S. Moule, M. J. Pallen, C. W. Penn, M. A. Quail, M. A. Rajan-dream, K. M. Rutherford, A. H. van Vliet, S. Whitehead, and B. G. Barrell.
2000. The genome sequence of the food-borne pathogen Campylobacter
jejunireveals hypervariable sequences. Nature403:665–668.
27.Sails, A. D., B. Swaminathan, and P. I. Fields.2003. Utility of multilocus sequence typing as an epidemiological tool for investigation of outbreaks of
gastroenteritis caused byCampylobacter jejuni. J. Clin. Microbiol.41:4733–
4739.
28.Schouls, L. M., S. Reulen, B. Duim, J. A. Wagenaar, R. J. L. Willems, K. E. Dingle, F. M. Colles, and J. D. A. Van Embden.2003. Comparative
geno-typing ofCampylobacter jejuniby amplified fragment length polymorphism,
multilocus sequence typing, and short repeat sequencing: strain diversity,
host range, and recombination. J. Clin. Microbiol.41:15–26.
29.Sharma, H., L. Unicomb, W. Forbes, S. Djordjevic, M. Valcanis, C. Dalton, and J. Ferguson.2003. Antibiotic resistance inCampylobacter jejuniisolated from humans in the Hunter Region, New South Wales. Commun. Dis. Intell.
27:(Suppl.)S80–S88.
30.Stanley, J., D. Linton, K. Sutherland, C. Jones, and R. J. Owen.1995.
High-resolution genotyping ofCampylobacter coliidentifies clones of
epide-miologic and evolutionary significance. J. Infect. Dis.172:1130–1134.
31.Suerbaum, S., M. Lohrengel, A. Sonnevend, F. Ruberg, and M. Kist.2001.
Allelic diversity and recombination inCampylobacter jejuni. J. Bacteriol.
183:2553–2559.
32.Taboada, E. N., R. R. Acedillo, C. D. Carrillo, W. A. Findlay, D. T. Medeiros, O. L. Mykytczuk, M. J. Roberts, C. A. Valencia, J. M. Farber, and J. H. Nash. 2004. Large-scale comparative genomics meta-analysis of Campy-lobacter jejuniisolates reveals low level of genome plasticity. J. Clin.
Micro-biol.42:4566–4576.
33.Tompkins, D. S., M. J. Hudson, H. R. Smith, R. P. Eglin, J. G. Wheeler, M. M. Brett, R. J. Owen, J. S. Brazier, P. Cumberland, V. King, and P. E. Cook.1999. A study of infectious intestinal disease in England:
microbio-logical findings in cases and controls. Commun. Dis. Public. Health.2:108–
113.
34.Wassenaar, T. M., B. N. Fry, and B. A. van der Zeijst.1995. Variation of the
flagellin gene locus ofCampylobacter jejuniby recombination and horizontal
gene transfer. Microbiology141:95–101.
35.Wassenaar, T. M., and D. G. Newell.2000. Genotyping ofCampylobacter
spp. Appl. Environ. Microbiol.66:1–9.