Vol.33,No.1 JOURNALOFVIROLOGY, Jan. 1980,p.250-255
0022-538X/80/01-0250/06$02.00/0
ev
2,
aGenetic Locus
Containing Structural Genes for
Endogenous Virus, Codes for Rous-Associated Virus Type
0Produced by Line
72
Chickenst
S. M.ASTRIN,'* L. B.CRITTENDEN,2 ANDE. G. BUSS3
The InstituteforCancerResearch,Philadelphia, Pennsylvania 19111,1 U.S. Department of Agriculture
Science andEducationAdministrationRegional Poultry Research Laboratory, East Lansing, Michigan
48823,2andDepartment of Poultry Science, PennsylvaniaState University, University Park,Pennsylvania
16802
ev2 isoneofseven recentlydescribed geneticloci ofchickens which contain
structural genes for endogenous virus. ev 2 is present exclusively in line 72
chickens,aninbred strain of whiteLeghorns which is homozygousfor thecapacity
to produce Rous-associated virus type 0 (RAV-0), a subgroup E virus. This
phenotype is knownasV+and hasbeen assignedagenetic allele designated
V-E7.The segregationofev2wasfollowed inageneticcrossin which the V-E7+
phenotype was also segregating. The progeny of the cross were analyzed for
endogenousviral lociby cleavage of embryo DNA with restriction endonuclease SstI, electrophoretic separation oftheresulting fragments, andidentificationof
bands containing viral sequencesby hybridization of the DNAto radiolabeled
viral RNA. Fourendogenousviralloci,ev 1,ev2,ev4, andev5,wereidentified
intheprogenyof thecross.One of theprogenycontainednodetectable
endoge-nousviralsequences. ev1,ev4,andev5werepresent inprogenyof boththe
V-E7+andV-E7-phenotypes. ev2waspresentexclusivelyinprogenyof the V-E7+ phenotype,and allV-E7+progenycontainedev2.Inaddition,oneof the V-E7+
progeny containedonly ev2. Fromthese data,we concludethat ev2 codes for
RAV-0 virusproduced bythecellsof line72 chickens.
Line 72 is an inbred line ofwhite Leghorns
which is geneticallyhomozygous for the V-E7
allele, an allele controlling the production of Rous-associated virus type 0 (RAV-0), a sub-group E endogenous retrovirus (6, 8, 13, 19).
Recently, line 72 chickens have been shownto
containendogenousviralgeneticinformationat twospecific chromosomal sites of residence (1). The genes at these twosites have been
desig-natedendogenous viral loci1and2(abbreviated
ev 1 and ev2). ev 1 was found in all chickens
tested, butev2wasfoundexclusivelyinline72
chickens. This finding suggests that ev 2 may code for RAV-0 virusproduced by the cellsof
these chickens.
To testthishypothesis, wehavefollowedthe
segregation ofev 2 ingenetic crosses involving
the progeny of line 72 chickens. The progeny,
which wereheterozygousfor theproduction of
RAV-O, were mated with nonproducer (V-E-) chickens, and the resulting embryos were
as-sayedfor thecapacitytoproduceRAV-O(V-E7+ phenotype) and for the presence of ev 2. We found an exact correlation between the
segre-gation of the V-E7+ phenotype and the
segre-t Journal series no.5870 of the Pennsylvania Agricultural ExperimentStation.
gation ofev 2; in addition, one V-E7+ embryo containedonlyev2. Weinterpretthesefindings
toindicate thatev 2 codes forRAV-Oin line 72
chickens.
MATERIALS AND METHODS
Chickens. Line 72 isan inbred line of white
Leg-hornsof theV+ phenotype which is maintained at the
Regional PoultryResearchLaboratory, East Lansing,
Mich. (6). Hens of the V-E-phenotypewereobtained
fromSPAFAS, Inc., Norwich, Conn. V-E- henswere
negative for viralgroup-specific antigen(gs) and viral
envelopeprotein(chf).Individualmatingsweremade
between birds in individual cagesby artificial
insemi-nation.
Cells and cell DNAs. Chicken embryo cultures
were prepared from 11-day-old white Leghorn
em-bryosasdescribedbyRubin(16).Cellsweretypedfor
production ofRAV-0byinfectingamixture ofBryan
RSV-transformed quail cells and turkey cells with
culture supernatant.SubgroupE virusreleasedbythe
quail-turkey cell mixture wasassayed for focus
for-mationonsubgroupE-susceptible chickencells(5).
The sires, dams, and progeny embryos were not
susceptibletoinfectionby subgroupEvirus.DNA was
prepared from cultured cells or erythrocytes as
de-scribedbyVarmusetal.(18).Erythrocyteswere
iso-lated from heparinized whole blood by differential
centrifugation. 250
on November 10, 2019 by guest
http://jvi.asm.org/
Enzymes, gels, andtransfer of DNA to
nitro-cellulose filters. RestrictionendonucleaseSstI was
purchased from Bethesda Research Laboratories.
DNAs at aconcentrationof250,ug/mlweredigested
by using 250 U of endonuclease perml at 37°C for 4 h.
Digestion was monitoredby the additionof SV40 or
A DNA to a portion of the digestion mixture and
visualization of the products, after digestion and elec-trophoresis, by staining with ethidium bromide. Elec-trophoresis of digested DNAs was carried out in 1%
agarose (Sea Kem) slab gelsasdescribed by Ketner
andKelly (10). A 25-,tg amountof DNA was run in
eachwell. DNAwastransferredtonitrocellulose filters
(HAWPOO010; Millipore Corp.) by the method of
Southern (17) as modified by Ketner andKelly (10).
Hybridization and autoradiography.
3P-la-beled RAV-2 RNAwasusedas aprobe for endogenous
viral sequences. RAV-2 RNA has greater than 80%
sequencehomology withthe RNA of RAV-0 (9, 11).
RAV-2 RNA was chosenfor these studies since
RAV-2grows to high titer whichfacilitates the preparation
of the hybridization probe. RAV-2-infected cell
cul-tures werelabeled with 32P as described by Parsons et
al. (12), using 1.5 mCi of[32P]phosphate (carrier-free,
NewEngland Nuclear Corp.) per ml of culture fluid.
Virus wascollectedandpelletedat 24-hintervals, and
70S RNA was purified by pronase-sodium dodecyl
sulfate (SDS)treatmentfollowedbysucrosegradient
sedimentation asdescribed by Robinson et al. (15).
Purified 70S RNA washybridized at 70°C to DNA
immobilized onnitrocellulose filtersin amixture
con-taining4xSSC(lx SSC is0.15MNaClplus 0.015 M
sodiumcitrate,pH 7.4),500tigof carrier wheatembryo
RNA perml,0.1%SDS, and 32P-labeled 70S RAV-2
RNA(specificactivity,1 x 107to2 x 107cpm/,ug) at 0.3,ug/ml.The salt and temperature conditions chosen
forhybridizationarethosedescribedbyBaludaetal.
(3). The filters wereincubated in hybridization
niix-turesfor10 to 20h,washed with2xSSC,treatedwith
pancreaticRNAase (20pg/mn in2xSSC for0.5h at
37°C), washed with4x SSC and 0.1% SDS at70°C,
and dried.Hybrid bandsweredetectedby
autoradiog-raphy at -70°C by using Kodak XR-5 film and a
DupontQuanta IIintensifyingscreen.
RESULTS
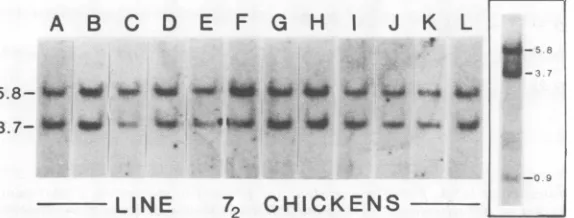
Endogenousviralloci in line 72chickens.
A
B
C
D
E
F
G
H
DNA was extracted from erythrocytes of 26 birds of line 72 and cleavedwith restriction
en-donuclease SstI. This enzyme was chosen be-cause it produces a single major fragment for each site of residence ofendogenous viral genes in the chicken genome(1). DNA fragmentswere
fractionated by gelelectrophoresis, denatured in situ, and transferredtonitrocellulose filters(17).
Endogenous viral genes were detected by hy-bridization with 32P-labeled70SRNA extracted
from virions of RAV-2. Bands containing viral sequences were visualized by autoradiography. All 26 DNAs produced identical patterns with fragments of 5.8 x 106, 3.7 x 106,and 0.9x 106 daltons. Of the 26patterns, 12 are shown inFig.
1, lanes A through L. After a long
autoradi-ographic exposure, a third band at 0.9 x 106
daltonscould be seen in every well (inset, Fig.
1). The band at 5.8x 106daltons has previously
been identified as representing a single site of residence of endogenous viral genes and has beendesignated ev1.Thebandsat3.7 x106 and 0.9 X 106 daltons represent a second site of residence, designated as ev 2 (1). Digestion of the DNAs withHindIIIwasusedtoconfurm the presence of ev 1 and ev 2 in these six DNAs. HindIII cleavage producedbands at 8.5 x 106,
3.2 x 106, 2.4 x 106, 1.9 x 106, and 1.0 x 106 daltons (data notshown). The 3.2 x 106, 1.9 x
106, and 1.0x 106 bandsrepresent ev 1,and the 8.5and 2.4 bands arecharacteristic ofev2 (1).
The 26 line 72 chickens that weanalyzedwere
specifically chosen to be representative of the
gene poolpresent in thebreeding stocks of line
72.Wecan,therefore,conclude fromourresults
that line 72 chickens contain onlytwo loci for endogenous viral genes, loci ev 1andev 2.Two roosters (Fig. 1, lanes A and B) were chosen
from the group of 26birdsanalyzed tobe used
assires in themating experiments.
Segregation ofgenotype andphenotype
in backcross matings. Two line 72 roosters wereeachmated withtwodifferent hens of the
I
J
K
ip
L
5.8-_%W 00
4_0
am_ 000,*W.
--w.-3.7-b
W o l".,%WIwo
iw
_0. -w _.NLINE
72
CHICKENS
_-5 8
_ II
- 9
-O g
FIG. 1. SstIdigestionof line 72chicken DNAs andidentification of endogenousviralsequences. A marker
ofEcoRI-digested 32P-labeledADNAwasusedtodeterminemolecularweights(whichareshown x10').The
insertattherightshowsalongautoradiographicexposureofthesamplein lane A.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.504.104.391.513.622.2]252 ASTRIN, CRITTENDEN, AND BUSS
V-E-phenotype.Progeny from each of the
mat-ingsweregrown tomaturity,andonemale from eachwasselected foruse as asire inabackcross. Eachof these four F-1 roosters wasmatedto a
V-E-gs-chf-hen, and10to 12progenyembryos fromeachmatingwereanalyzedforproduction
of RAV-0 and for endogenous viral locias de-scribed above.
TheSstI patterns for thesire, dam,and prog-enyfor each of the fourmatings areshown in Fig. 2 through 5. Various combinations of four bands (12 x 106, 5.8 x 106, 5.4x 106,and3.7 x
106 daltons) canbe seenin the patterns. After longautoradiographicexposures, afifth bandat 0.9 X 106 daltons could beseen always
accom-panying the 3.7 x 106 molecular weight band (data not shown). As described above, the 5.8
X 106 daltonband represents ev 1; the 3.7 and 0.9molecularweightbandsrepresentev2.The bandsat5.4x106and12x 106daltonsrepresent twoother distinct sites of residence of viralgenes
(1) that have been designated ev 4 and ev 5,
respectively. ev4 and ev5 have been observed
tosegregateinaMendelianmanner(2).
Appro-priate DNAs have been cleaved with BamHIto
confimnthepresenceofev4andev5(seelegends
toFig.2and4).
Inmating1 (Fig.2), the sire carriedev 1and
ev 2 (Fig. 2, laneA) and the dam carried ev 1
and ev 5 (Fig. 2, laneB). Of the 11progeny, 9
inheritedev1 (Fig.2, lanesC, D, E, F, G, H, I,
K, and L), fourprogeny inherited ev5 (Fig. 2,
lanes F, G, L, andM), andfour inherited ev2
(Fig.2,lanesD,F, L,andM).One of theprogeny
(Fig. 2, laneJ)appearstolack
endogenous
viral loci. To ensure that therewasindeed DNA inwellJ, thegelswerestained withethidium bro-mide before transfer of the DNAto
nitrocellu-lose filters and the DNAwasvisualized with a
UV lamp.Well J
appeared
tocontainanornalamount ofDNA.Inaddition,this DNA
sample
was analyzed asecond time and again showed nohybridization. The frequencies ofoccurrence
of individual loci in these eleven progeny are
consistent with the dam beingheterozygous for
ev1andev5andthe sirebeing heterozygous for
ev 1 andev2.Inthiscase, 1/16oftheprogeny
wouldbe expectedtolack endogenousviralloci. All theprogenythatinheritedev 2(Fig.2,lanes
D, F, L, and M)were of the V-E7+phenotype.
The remaining progeny were of theV-E- phe-notype. Thus, ev 2 segregated with theV-E7+
phenotype ineveryinstanceinthismating.
Inmating2 (Fig. 3), the sire carried ev 1 and ev 2 (Fig. 3, lane A) and the dam carriedev 1
and ev 5 (Fig. 3, lane B). Of the 10 progeny,8
inherited ev 1 (Fig. 3, lanes C,D, E, F, I, J, K,
andL),7progenyinheritedev 2(Fig.3,lanesD,
E,G, H,I,K, and L), and threeprogenyinherited
ev 5 (Fig. 3, lanesH, I, and J).As inmating 1,
these frequencies are consistent with the dam
being heterozygous forev 1andev 5 and the sire
being heterozygous for ev 1 and ev 2. All the progenythatinherited ev 2(Fig. 3,lanes D,E,
G,H, I, K, and L)wereofthe V-E7+ phenotype, whereas the remaining progenywere ofthe
V-E-phenotype. In the case of the V-E7+ embryo
whosepatternis shown inFig. 3laneG, ev 2is
the only locus present. Therefore, the RAV-0
produced by the cells of this embryo mustbe
codedfor byev 2.Thesegregation ofev 2with theV-E7+phenotype in the otherprogeny sup-portsthis conclusion.
In matings 3 and 4 (Fig. 4 and 5), the sires
carried ev 1 and ev 2 (A lanes) and the dams carriedev 1andev 4(Blanes). All of theprogeny
of bothmatings inherited ev 1. Four of the 11 progenyofmating3inheritedev 4(Fig.4,lanes E, F,K, andM), asdid 3ofthe 12 progeny of
mating4(Fig.4,lanesE,I, and K). Nineprogeny
of mating 3 and seven progeny of mating 4
inheritedev 2(Fig.4,lanesC, D,E, F, G, I, J, L,
A B C D E F G H u K
LM
A N_hi _
4-S-_
to_1
*
PROG E NY
-FIG. 2. SstIdigestionof DNAsfrom the sire, damandprogenyotmating no. 1.Molecularweignts(xiu)
areshown. AmarkerofEcoRI-cleaved32P-labeledA DNA isshown inLane N. +,Embryo wasof the V'
phenotype;-, VWphenotypeembryo. Assays forRAV-Oproduction wereperformed as described in the text.
BamHIdigestionofthe DNA shown in lane Byielded bandsat 7.8 x 106,3.2 x106, 1.0 x 106,and 0.7 x106
daltons(datanotshown). This is the expectedpatternforaDNA containing only ev 1 and ev 5 (1). The sire
(laneA)isanF-I betweenamalehomozygous forev1andev2and afemale heterozygousfor ev 4.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.120.409.497.599.2]VOL. 33,1980
A B C D E F G H
_;~~~~~~~~*,O*
J K L M}13 7t
ev 5)
12-ev 1) 5.8- _ i_ ii ev 2) 3.7-to
t~~~~~~~~~~~~~~~~
we i4Iiit-33
-21
v + + -+4 ±
--1-1
+
PROGENYFIG. 3. SstIdigestion ofthe DNAsfromthesire, dam, and progenyof matingno.2. Molecularweights
(x106) are shown. A markerof EcoRI-cleaved32P-labeledA DNA is shown in lane M. +,Embryowasofthe
V+ phenotype; -, V-phenotype. The sire(lane A)isanF-I betweenamalehomozygous forevI andev2 and
afemale heterozygous forev 4.
A
B
C D E
F G
H
I
J
K
L
M
ev 1
-ev 4
%a -ev 2
+ t t -4- - + - + +
[image:4.504.96.384.72.178.2]d
-o
PROGENY
FIG. 4. SstIdigestionofDNAsfromthesire, dam, and progenyof matingno.3.Molecularweights(x106)
wereestimatedby usingamarkerofEcoRI-cleaved32P-labeled A DNA (not shown). +,EmbryowasoftheV+
phenotype; -, V-phenotype. BamHIdigestionofthe DNA shown in lane Byielded bandsat4.5x106 3.2x
106, 1.0x106,and0.7x106 daltons(datanotshown).This is theexpectedpatternforaDNAcontaining only
ev 1 andev4(1). The sire(lane A) isanF-i between amale homozygous forev 1 andev2andafemale
homozygousforev 1.
A B C D E F G H I J K L M N
5.8_- u_ "
5.4-3.7
-ev 1
- I..
VW. to*
-oo$
-111
evev 441.-,: -ev 2
Ce""
- + --f
+ + + - t -+
t --
-PROGENY
FIG. 5. SstIdigestionofDNAsfromthesire, dam,and progenyofmatingno. 4.Molecularweights(x106)
wereestimatedby usingamarkerof EcoRI-cleaved32P-labeledX DNA (not shown). +, Embryo was of the V+
phenotype; -, V-phenotype. The sire(lane A) isanF-I betweenamalehomozygous forev Iandev 2anda
female homozygousforev1andheterozygous forev4andev 5.
andM;Fig.5,lanesE,G,H,I,J, L, and N). All the progeny thatinherited ev 2 wereof the V+
phenotype; the remaining progeny were of the V- phenotype. Thus in all 45 progeny, ev 2
segregated with theV+phenotype. In addition, in oneof theV+progeny, ev 2 wasthe only locus present. We conclude that ev 2 codes for the virusproduced bycells ofline 72 chickens.
DISCUSSION
Recently, the identification of seven genetic
loci containingstructuralgenesforendogenous virus was reported (1). Two of these loci, ev2
andev7,arefoundexclusivelyinembryosofthe
V-E7+ or V-15+ phenotypes, respectively, ev 2
beingcharacteristic of line72 embryosandev7
5.8=ImF s _ %W
3.7-eV 2 253
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.99.387.235.354.2] [image:4.504.86.387.427.529.2]254 ASTRIN, CRITTENDEN, AND BUSS
being characteristic of line 15B embryos. The virus produced by line 72cellsis a subgroup E
retrovirusdesignatedas RAV-0 (5, 19), whereas
line 15Bcellsproducenoninfectiousvirus
parti-cles of unknownsubgroup (14). We have recently documented the existence of three additional
loci inchickens, one of which, ev 10, is
charac-teristic ofV-E+ embryos oflines1514 and C (L.
B. Crittenden and S. M. Astrin, manuscript in
preparation). Lines 1514 and C produce a sub-group Evirussimilar butnotidenticalto RAV-0 (P. Tsichlis and J. Coffin, unpublished data). Thus there appear tobe atleastthree distinct types of virus produced by thesethree pheno-typesof V+ embryo, each of which is associated with the presence of a distinct genetic locus containingendogenous viralsequences. One in-terpretation of thisrelationship isthateachof these loci codes for the virus produced by the cells containing that locus.Alternatively,
corre-lation between locus andphenotype could sim-ply reflect the fact that the chickens inwhich they are observed are genetically related. In experiments reported herewepresentevidence thatev 2 codes for RAV-0produced by cells of line72embryos. Notonly didev 2segregatewith
the V-E7+ phenotype in everyinstance infour
matingexperiments, butoneof theV-E7+ prog-enycontainedonlyev 2.Thusev 2mustcontain the genetic information for the virus produced by the cells of thisembryo.
Virus produced by cells of line 72 embryos
grows to a high titeronly after induction with BUdRorafter co-cultivation with cells
suscep-tibletosubgroup Evirus (7, 13). Therefore, ev 2mustnormallybeexpressedatonlyaverylow level. Recentexperiments ofCooperand
Silver-man (4), using line 100 chickens (a subline of line 72), indicate thepresenceofregulatory
se-quences,adjacenttotheviralsequences, which
repress transcription. Their data showed that theDNAofsubgroupE-resistantV-E+ cellswas
infectious in a transfection assay only after shearing to the approximate size of the virus
genome. Since the results presented here indi-cate that ev2 codesfor the virus produced by
cellsofline72 chickens, thefindingsofCooper
and Silverman implythat ev 2transcription is
regulated by linkage to cis-acting control
ele-ments. Furtherworkwill benecessaryto
deter-mine the nature and mode of action of these
control elements.
One of the progeny in the experiments
re-ported here lacked detectableendogenousviral genes (Fig.2,laneJ).We haveanalyzed
endog-enousviralsequences inover500chickens, and
this is the firstexampleofanindividual
appar-entlylackingthese genes. We haverepeatedthe
J. VIROL.
mating which produced this embryo and have succeeded in hatching a rooster which has no
detectable endogenous viral sequence (la).
Theseresultsindicatethatthe endogenous viral genes perform no essential function in the
chicken, atleasttothe age of7months,which
isthe present age ofourrooster. Chickens lack-ing endogenous viralgenesshould prove useful
inassessing the role of thesegenesinontogeny,
oncogenesis, and disease.
ACKNOWLEDGMENTS
Wethank DebraKeefer,Joanna Fang, FrancesGulvas, and Deborah Eagen forexcellent technical assistance.
This work wassupportedby grant VC232 from the Amer-icanCancerSociety, grant PCM 76-09721 from the National ScienceFoundation,grants CA06927andRR05539 from the National Institutes ofHealth,Interagency agreement Y01-CP-40214 with the Division ofCancer Cause and Prevention, National CancerInstitute, and by an appropriation from the Commonwealth ofPennsylvania.
LITERATURE CITED
1. Astrin, S. M. 1978. Endogenous viral genes of the white Leghorn chicken: common site of residence and sites associated withspecific phenotypesof viral gene expres-sion.Proc. Natl. Acad. Sci.U.S.A.,75:5941-5945. la.Astrin, S. M., E. G. Buss, and W. S. Hayward.1979.
Endogenous viral genes are nonessential in the chicken. Nature(London) 282:339-341.
2. Astrin, S. M., and H. L.Robinson. 1979. Gs, an allele of chickens forendogenousavianleukosis viralantigens, segregates with ev 3, a genetic locus that contains structural genes for virus. J. Virol. 31:420-425. 3. Baluda, M. A., M. Shoyab, P. D.Markham, R. M.
Evans, and W. N. Drohan. 1974. Base sequence com-plexity of 35S avian myeloblastosis virus RNA deter-minedby molecularhybridizationkinetics. ColdSpring HarborSymp. Quant.Biol. 39:869-874.
4. Cooper,G.M.,and L. Silverman.1978.Linkageof the
endogenous avianleukosis virus genome of
virus-pro-ducing chicken cells to inhibitory cellular DNA
se-quences.Cell 15:573-577.
5. Crittenden, L. B., D. A.Eagen, and F. A. Gulvas.
1979.Assays forendogenouslymphoid leukosis viruses andchickenhelper factor with RSVC-1 cell lines. Infect. Immun.24:379-386.
6. Crittenden,L.B.,J. V.Motta,and E. J. Smith.1977.
Genetic control of RAV-0production in chickens.
Vi-rology76:90-97.
7. Crittenden,L.B.,E. J.Smith,F. A.Gulvas,and H.L.
Robinson. 1979.Exogenousvirusexpressionin chicken lines maintainedattheRegionalPoultryResearch Lab-oratory.Virology95:434-444.
8. Crittenden,L. B.,E.J.Smith,R.A.Weiss,and P. S.
Sarma.1974.Hostgene controlofendogenousavian leukosisvirusproduction. Virology57:128-138.
9. Fujita,D.J.,J.Tal,H. E.Varmus,and J. M.Bishop.
1978.envgene ofchicken RNAtumorviruses. Extent of conservationincellular and viral genomes. J.Virol. 27:465-474.
10.Ketner,G.,andT. J.Kelly,Jr.1976.Integratedsimian virus40sequences in transformed cell DNA: analysis usingrestrictionendonucleases.Proc.Natl.Acad.Sci. U.S.A.73:1102-1106.
11.Neiman,P.E., S.Das, D.MacDonnell, andC.
Mc-Millin-Helsel. 1977. Organization of shared and un-sharedsequencesin thegenomesofchickenendogenous andsarcomaviruses.Cell11:321-329.
on November 10, 2019 by guest
http://jvi.asm.org/
12. Parsons,J.T.,J. M.Coffin,R.K.Haroz,P. A. Brom-ley, andC. Weissman. 1973.Quantitative determina-tion and location of newly synthesized virus-specific ribonucleic acid in chicken cells infected with Rous sarcoma virus.J.Virol. 11:761-774.
13. Robinson, H. L. 1978. Inheritance and expression of chicken genes thatarerelated to avianleukosis sarcoma virus genes.Curr. Top. Microbiol. Immunol. 83:1-36. 14. Robinson, H. L., C. A.Swanson,J. F.Hruska,andL.
B.Crittenden.1976.Virology69:63-74.
15. Robinson, W. S.,A. Pitkanen, and H. Rubin. 1965. Thenucleic acid of the Bryan strain of Rous sarcoma virus: purification of the virus and isolation of the
nucleic acid. Proc.Natl. Acad. Sci. U.S.A. 54:137-144. 16. Rubin, H. 1957. Interactions between Newcastle disease virus (NDV),antibody, and cell. Virology 4:533-562. 17. Southern, E. M. 1975. Detection of specific sequences
among DNAfragments separated bygel electrophore-sis.J. Mol. Biol. 98:503-517.
18. Varmus, H.E., and J. M.Bishop.1973.Appearance of virus-specific DNA in mammalian cells following trans-formation by Rous sarcoma virus. J. Mol. Biol. 74:613-626.
19.Vogt, P.K., and R.Friis. 1971. An avian leukosis virus related toRSV(0):properties and evidence for helper activity. Virology 43:223-234.
on November 10, 2019 by guest
http://jvi.asm.org/