Copyright ©) 1991, AmericanSociety for Microbiology
Contribution of
NF-KB and
Spl
Binding
Motifs to the
Replicative
Capacity of
Human
Immunodeficiency Virus
Type 1: Distinct
Patterns
of
Viral Growth Are Determined by T-Cell Types
ELIZABETH K. ROSS,1 ALICIA J. BUCKLER-WHITE,2 ARNOLD B. RABSON,1t GEORGE ENGLUND, AND MALCOLM A. MARTIN1*
Laboratory of Molecular Microbiology, National Institute of Allergy andInfectious Diseases, Bethesda, Maryland
20892,1
andDivision ofMolecular Virology andImmunology, Georgetown University Schoolof Medicine,
Rockville, Maryland208542 Received 13 February1991/Accepted9May 1991
Startingwith areplication-incompetentmolecular clone ofhumanimmunodeficiency virustype1,lackingall the NF-KB and Spl binding sites present in the native long terminal repeat (LTR), proviruses containing reconstructed LTRs with individual or combinations of NF-KB and Spl elementsweregeneratedand evaluated fortheir capacity to produce virus progeny following transfection-cocultivation. Virusstocks obtained from theseexperiments exhibitedacontinuum ofreplicativecapacitiesindifferenthuman T-celltypesdependingon whichelement(s) was present in the LTR. For example, inexperiments involving proviral clones with LTRs containing one ortwoNF-KBelements (and no Spl bindingsites), ahierarchyofcellularpermissivitytovirus replication (peripheral blood lymphocytes = MT4 > H9 > CEM > Jurkat) was observed. Of note was the associated emergence of second-site LTR revertants which involved an alteration of the TATA box. These results suggest that the human immunodeficiency virus type 1 LTR possesses functional redundancy which ensures virus replication in different T-cell types and is capable of changing depending on the particular combination oftranscriptional factorspresent.
The retroviral long terminal repeat (LTR), generated dur-ing the early phase of virus infection by reverse transcrip-tion, contains several cis-acting elements that influence viral geneexpression. These include the transcriptional promoter and start site, the polyadenylation signal and addition site, and binding motifs for transcriptional activators. In some retroviral subfamilies, the LTR also contains targets for virus-encoded transactivator proteins.
During the past few years, the structure and function of the human immunodeficiency virus (HIV) LTR has been intensively investigated. Regulatory motifs (e.g., Spl, TATA)have been identified which bind cellular proteins that are constitutively expressed in most cells (7, 11, 20); these sequences appear to be critical for basal-level HIV gene
expression.OtherLTRelements (e.g., NF-KB and NFAT-1) may interact with factors synthesized only in activated T
lymphocytes (6, 21, 28, 37), thereby limiting vigorous virus
replication to a small subset of circulating peripheral blood
lymphocytes(PBLs) invivo. Still other regions (e.g., TAR) are unique to HIV and are required for high levels of LTR-directed gene activity (4, 8, 13, 18, 36, 41). Thus, the
interaction of virus-encoded (3, 40) and cellular regulatory proteins with one another and with the HIV LTR in both resting and activated human lymphocytes has the potential toeffect acontinuum of replicative capacities.
Todate, most of our knowledge of HIV LTR structure and function comes from studies of small DNA constructs con-taining the LTR linked to reporter genes and tested in HeLa cellsorin humanT-cell lines (2, 5, 15, 28, 32, 38). Although this approach has helped to elucidate the mechanism by which the HIV LTR is activated, the interaction of
virus-*Correspondingauthor.
tPresent address:Center for Advanced Biotechnology and Med-icine, Piscataway, NJ 08854.
encoded and cellular proteins with cis-acting regulatory
elements duringa productive infectioncannotbe evaluated in such systems. We (23) and others (14, 24) have electedto evaluateLTRfunction in the context of HIVreplicationand have previously reported that some of the cis-acting ele-ments may be dispensable, depending on the cell type infected. Forexample, the deletion of both NF-KB elements only modestly affects HIV replication in PBLs or in CEM andMT4cells (23). In contrast, mutation ofSpl motifscan drasticallyreducereplicativecapacityinsomehuman T-cell cultures. Inparticular,thedeletion of all threeSplelements either eliminated orgreatly reduced the capacity of HIV to infect CEM cells (two successes in fiveindependent exper-iments); replication of this mutant in PBLs and MT4 cells was only slightly reduced (30). Finally, the simultaneous elimination of the two NF-KB and all three Spl elements resulted in a provirus exhibiting no detectable biological activity (23); in this report, we refer to this irreversibly replication-incompetent proviral DNA aspNL-ZERO.
To understand more fully the contribution of individual LTR components to the replicative process and to the
complexinteractions of cellular and viralproteinswith their LTR targets, we analyzed the biological activity of pNL-ZEROinto which different combinations of NF-KB andSpl cis-acting elementswere added. Our results suggest a hier-archy of cellularpermissivity to virus infectionwhich very likely reflects the relative abundance oftranscriptional reg-ulatory factors and the presence of specificHIV LTRmotifs.
MATERIALS ANDMETHODS
Construction ofproviral DNAs containing LTR mutations and generation of virus stocks. pNL-ZERO is a replication-incompetentderivative of the infectiouscircularlypermuted proviralclonepILIC(23); pNL-ZERO lacks the two NF-KB 4350
on November 10, 2019 by guest
http://jvi.asm.org/
and three Spl sites and was previously designated pTA. pNL-ZERO DNA was digested with BamHI and SphI to generate a 2.1-kb fragment containing the LTR. This
frag-ment was subcloned into M13 and subsequently used to produce the reconstructed LTRs, shown in Fig. 1, by oligonucleotide-directed mutagenesis (47) as modified by
Kunkel(22). Full-lengthcircularlypermutedproviralDNAs were reconstituted by ligating the LTR-containing 1.7-kb
XhoI-SphI fragment from M13 recombinants to a similarly
digested7.4-kb cleavage productofpNL-ZERO. Following the verification of the mutations prepared in vitro by
nucle-otidesequenceanalysis(34), thereplicative capacities ofthe
different LTR mutants were evaluated by transfecting a derivative of A3.01 (9)CEMcells, designated 12D7 (gener-ously providedbyGuido Poli, Laboratory of Immunoregu-lation, National Institute of Allergy and Infectious Diseases) with 3 ,ug of DNA (per 6 x 106 cells) in the presence of DEAE-dextran as previously described (23). Forty-eight hours later, thetransfected cells werecocultivated with2 x 106 MT4(25)cells and monitored for virus production by a
32P-reversetranscriptase (RT) assay (46). Virus stocks were
prepared from the supernatants of transfection-cocultures
when RT levels exceeded 3,500
32p cpm/,lp.
Infections. PBLs, the A3.01 (9)clonal derivative ofCEM
cells, MT4, H9 (31), and JE6.1 (45), a clonal lineof Jurkat cells (2 x 106 cells of each type), were infected with
equivalentamounts of virus (300,000 cpm of32P-RT activi-ty). Priortoinfection, PBLs werestimulated with phytohe-magglutinin (0.25 ,ug/ml) for 3 days; upon infection, the
PBLs were maintained in medium containing 10%
interleu-kin 2. Infected cells were maintained in T25 Teflon flasks (Costar)in RPMI 1640medium(10 ml)containing 10%fetal calf serum, and progenyvirion production was detected by
RT activity.
PCR analysis of genomic RNA.Aliquots(100 ,ul)ofculture supernatants from selected infections were digested with
RNase-free DNase (0.05 U; PromegaBiotec) for 15 min at
37°C and then incubated for10min at68°Ctoinactivatethe DNase. First-strand cDNA synthesis from viral RNA was
performed as described by Morgan et al. (26). The cDNAs werethen amplified in aPerkin-Elmer/Cetus DNAThermal
Cycler with thefollowing primers: (i)a plus-sense oligonu-cleotide spanning positions 9073 to 9092
(HIVBRU
coordi-nates[27])with 9nucleotidesatits5' endbearinganEcoRI site (5'-ATCGAATTCTGGAAGGGCTAATTCACTCC-3'; gift from Jonathan Silver), and (ii)aminus-sense oligonucle-otide spanning positions 9504to9521 with12nucleotidesat
its 5' end bearing botha PstI and an RsaIsite (5'-AACTG
CAGGTACCAGGCAAAAAGCAGCTGC-3').
The amplifi-cation protocol consisted of denaturation at 94°C for 30 s,annealing at
60°C
for 30 s, andextension at72°C for2 min.Digestion of the polymerase chain reaction (PCR) product with
EcoRI
and PstI, ligation intopIBI30
(International Biotechnologies, Inc.), transformation of Escherichia coliDH5-aMCR (Bethesda Research Laboratories), and selec-tion of recombinants by colony hybridization by using a
5'-end-labeled
oligonucleotide probe spanningpositions 143 to 182 of theHIVNL4-3
(1, 27) LTR were all done aspreviously described (33). Hybridization-positive
recombi-nants were sequenced by the method ofSangeretal. (34).
RESULTS
We previously described the construction ofa circularly permuted infectious molecular clone of HIV containing a
single LTR which was used to investigate the role(s) of
NF-KBand
Spl
elementsduring
virusreplication
in human T cells (23). Viruses in which the two NF-KB binding sites were deletedreplicated efficiently
in phytohemagglutinin-stimulatedhumanPBLs, aswellas in MT4 and CEMcells. HIVpreparations containing
one or twomutatedSpl
bind-ing
sites were alsoreplication
competent but exhibited variablegrowth kineticsdependingonthe cell typeinfected. Deletion of all threeSpl
sites resulted in virus whichreplicated
efficiently
in MT4 cellsand in PBLs butinconsis-tently
andwithdelayed
kinetics inCEMTcells unlessthey
were exposed to tumor necrosis factor
alpha (30).
In con-trast, the deletion of68bp
ofU3 sequencesencompassing
the two NF-KB and the three
Spl
sitesgenerated
an HIVprovirus
which failed toreplicate following
transfection-cocultivation. Thisreplication-defective
HIVprovirus,
des-ignated pNL-ZERO,
wasused ina series ofexperiments
as a basal-levelproviral
DNA target to which NF-KB orSpl
siteswere added eitherindividually
orincombination.Generation ofmutant LTR HIV-1 stocks by transfection-cocultivation.
Circularly permuted
HIVproviruses
contain-ing
asingle wild-type
orreconstructed LTR werereleased fromplasmid
vectorsfollowing digestion
with BamHI andligated
to regenerate "conventional"full-length
proviral
DNAs
(i.e.,
containing
atleasttwoLTRs).
Aclonedderiv-ative ofthe
CD4+
T-cell lineA3.01,
designated 12D7,
was thentransfected withtheligated proviral
DNAsasdescribedinMaterialsandMethods.
Forty-eight
hoursfollowing
trans-fection,
the 12D7cellswerecocultivated with MT4cells andprogenyvirus
production
wasthen monitoredby
RTassay over a51-day
period. Supernatants
were collected at thepeak
ofRTactivity
and served as virus stocks for subse-quentinfectivity
studies.As
expected,
transfection-cocultivation withwild-type
HIV
proviral
DNA resulted inrapid
virusproduction
(first
detected 4 to 5
days later),
whereaspNL-ZERO
failed toreplicate.
As indicated inFig.
1, virusproduction
wasobserved
following
transfection-cocultivation ofproviral
DNAs
containing
one or two NF-KBelements,
although
replication
oftheformer(pNL-lKB)
wasmarkedly
delayed
(first
detectedonday 27).
ClonedHIVproviruses
containing
two orthree
Spl
sites(pNL-2Spl
andpNL-3Spl)
or asingle
pair
ofNF-KB andSpl
sitesseparated by
a mutatedinter-vening
NF-KB site(pNL-KB/Spl-R)
allreplicated
following
transfection-cocultivation but with kinetics somewhat
slower than that of wild type.
Interestingly,
thepNL-KB/
Spl-J
provirus
containing juxtaposed
NF-KB andSpl
single
elements
replicated nearly
asefficiently
aswild-type
HIV(data
notshown).
Ofthe mutated HIV LTRs evaluatedby
transfection-cocultivation, only
thepNL-lSpl
proviral
con-struction,
which contains thesingle Spl
elementSpl(I),
failed to
replicate
in threeindependent
experiments
lasting
51
days,
2.5months,
and 4months,
respectively.
Infection of PBLs and human T-cell lines with HIV-1
derivatives containing NF-KB or
Spl
motifs in theirrecon-structed LTRs.
Although
the transfection-cocultivationex-periments
described above indicated which ofthe NF-KB/Spl
insertions intothepNL-ZERO
LTRrestoredreplicative
capacity,
how these elements influenced virusinfectivity
required
additionalinvestigation.
Infections werethereforeperformed
employing
virusstocksgenerated
fromthetrans-fection-cocultures referred to in
Fig.
1. The various T-cell lineswere infectedwithequivalent
amountsofwild-type
or LTRmutantvirus(3
x 10532P-RTcpm,approximately
equal
to a
multiplicity
of infection of0.001forwild-type
virus),
andsubsequent
progeny virusproduction
wasmonitoredby
RTactivity.
on November 10, 2019 by guest
http://jvi.asm.org/
| U3 |R |U5
NF-kB Spi TATA
-341 -186 -106 -56 -28 + 1 +19 +42
IZ7
Z7W1
*---- 11
1
NF-kB(II) NF-kB(I) Spi (III) Sp1 (II) SpI(I)
TACAAGGGACTTTCCGCTGGGGACTTTCCAGGGAGGCGTGGCCTGGGCGGGACTGGGGAGTGGCGAGCCCTC
pNL-lSpl TAC
B pNL-2Spl TAC
pNL-3Spl TAC
TGGGGAGTGGCGAGCCCTC
C pNL-kB/Spl-J TAC
pNL-kB/Spl-R TACAAGGGACTTTCCGCTGCTCACTTTCCAGGGAGGCGTGGCC
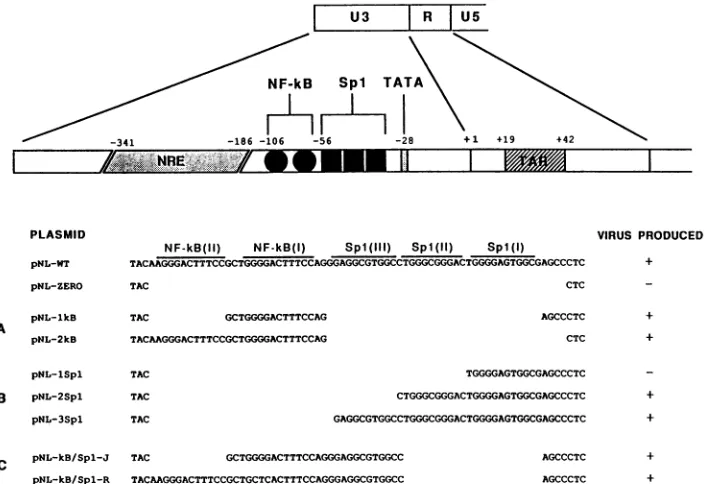
FIG. 1. Structure of reconstructed HIV LTRs associated with full-length proviral DNAs. The positions of several cis-acting regulatory elements within the HIV-1 LTRareindicated diagrammatically at thetop. The nucleotide sequencesofa72-bp region encompassingthe
NF-KB andSpl motifs of the wild-type LTR (pNL-WT)aswellaseight reconstructed LTRsareshown below. The nucleotide substitution
in pNL-KB/Spl-R, previously reported (28) to inactivate the NF-KB binding site, is underlined. Virus production following
transfection-cocultivation is indicated in the columnatthe right.
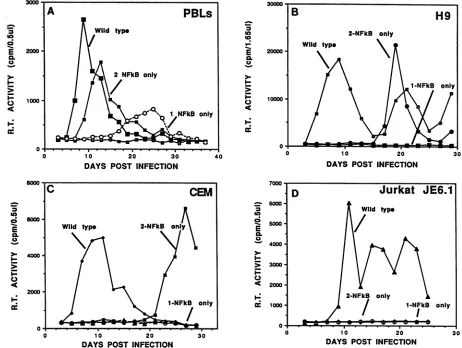
Inthefirstgroupof experiments, PBLs and different T-cell
lines (MT4, CEM [A3.01 clone], H9, and Jurkat [clone JE6.1])wereinfected withmutantHIVpreparations contain-ingone or twoNF-KB elements (andnoSpl motifs). Three
different replicative phenotypes were observed. The first, seenwithPBLsorMT4cells,wascharacterized by delayed
replication asthe number of NF-KB siteswasreduced from two to one (Fig. 2A). InH9 (Fig. 2B) and A3.01 (Fig. 2C)
cells, however, HIV with the single NF-KB(I) element failed to replicate; virus containing twoNF-KB sites in its
recon-structed LTR replicated slowly compared with wild-type HIV, generating virus progeny2.5to4weeks postinfection. Notably, infection ofA3.01 cells with HIVcontaining only twoNF-KBelements resulted in virusproductioninonlyone
ofthreeindependent experiments (the onedepicted in Fig.
2C) monitored for periods of up to 42 days. The third replicative phenotype was observed in Jurkat cells and is
shown in Fig. 2D. Inthis case, virus withLTRs containing oneortwoNF-KBelements failedtoreplicate during nearly 4weeks of observation.
A similar approach was followed to monitor the role of Splmotifsonvirus infectivity. Sincenoviruswasgenerated
following transfection-cocultivation of pNL-lSpl, the infec-tivity studies were limited to virus stocks containing two
[Spl(I) plus Spl(II)] or three [Spl(I) plus Spl(II) plus
Spl(III)] Spl motifs as shown in Fig. 1. In all cell types
tested, the presence of a second Spl element restored
infectivity. In some cells, HIVs with reconstructed LTRs containing two or threeSpl binding sites had similar repli-cation kinetics (PBLs in Fig. 3A), while in others, virus harboring only two Spl elements (H9 cells in Fig. 3B) reproducibly exhibited retarded growth properties.
Infectivity of HIV preparations with reconstructed LTRs containing combinations of NF-KB and Spl elements. As
shown in Fig. 2 and 3, duplications of either the NF-KB or
Spl element frequently resulted in enhanced HIV replica-tion. Giventheproximityof theNF-KBandSplmotifs inthe wild-type LTR configuration, we wondered whether the observed augmented replicative capacity was limited to
combinations of the sameelement. Toassessthepossibility
that NF-KB andSpl elements mightfunction ina
combina-torial fashion,wegeneratedaprovirus containingLTRswith
single copiesof both motifs. In the five human T-cell systems evaluated,this virus
(HIVNL-KB,sp1_J)
replicatedasrapidlyormore rapidly than HIV preparations with LTRs harboring duplications or triplications of the NF-KB or Spl elements (datanotshown).Thisphenomenonwasparticularly striking
inJurkat cellsinwhichHIV withtwoNF-KB elementsfailed to replicate during a 25-day observation period (Fig. 4A). Furthermore,theNL-KB/Spl-Jvirusreplicatedwith kinetics similar to that ofwild-type HIV in most of the cell lines tested(Fig. 4B).
We wondered whether the efficient viral replication di-rected by an LTR containing a pair of NF-KB and Spl
elements reflected theirjuxtaposition or simply their
pres-ence in the reconstructed LTR. Consequently, a different LTRcontainingthesinglefunctionalNF-KB(II)andSpl(III) elements spatially separated from one anotherby a
muta-genized (28) NF-KB(I)motif(seetheNL-KB/Spl-R provirus inFig.1)wasexamined for itsabilitytosupportaproductive
virus infection. In H9 cells, the replicative capacityof this viruswas similar tothat of virus containing thejuxtaposed NF-KB(I) andSpl(III) bindingsites(Fig. 5). Thus, an 11-bp
separationof functional NF-KBandSpl sites didnot
appre-ciably affectinfectivity; virions with this LTRconfiguration replicated more rapidly than those containing only two NF-KB ortwoSpl elements.
Replicative capacity of HIV proviral DNA containing a
PLASMID
pNL-WT
A
pNL-ZERO TAC
pNL-lkB TAC GCTGGGGACTTTCCAG pNL-2kB TACAAGGGACTTTCCGCTGGGGACTTTCCAG
VIRUS PRODUCED
CTC
-AGCCCTC +
CTC +
AGCCCTC
+
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.143.495.73.315.2]Wild type
PBLs
IFkB only
10 20 30
DAYS POST INFECTION
U2
(0 I.
-E
I-5:
0
40
DAYS POST INFECTION
8000
go 0. U
E
0
F-C.)
'a
2
U!
3.
I-0
--
so
' - 0 10 20 300 10 20 30
DAYS POST INFECTION DAYS POST INFECTION
FIG. 2. Infections ofhuman T cells withwild-typeHIVor mutantHIVderivativeswith LTRscontainingone ortwoNF-KBandnoSpl elements. Activated PBLs orH9, CEM, or Jurkat cells were infected with approximately 3 x 105 32P-RT cpm ofthe indicated virus preparations obtained by transfection-cocultivation. VirusproductionwasmonitoredbyRTassay.
Wild type PBLs
0
-2-Spl
1000 p
Mock
0 I-10 20 30
0 10 2003
B
to
E
a
3.- I-0
ci
8000
DAYS POST INFECTION DAYS POST INFECTION10 20 30
FIG. 3. Infections of human T cells withwild-typeHIVormutantHIV derivatives with LTRscontainingtwoorthreeSplandnoNF-KB elements. Activated PBLsor H9 cells were infected withthe indicated virus preparationsasdescribed in Materials and Methods. Virus
productionwasmonitoredbyRTassay.
I0
a
E
C.
a
3..
5:
I-0
2000-
1000-0
A
to
E
C)
S
0. a
I-0
A
0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.66.528.74.422.2] [image:4.612.91.526.518.690.2]B
a.
0
20 30 10 20
DAYS; POST IN.FECTION
[image:5.612.109.534.72.243.2]DAYS POST INFECTION
FIG. 4. Infections of human T cells with wild-type HIV or mutant HIV derivatives with LTRs containing pairs of homologous or
heterologous cis-acting elements. Jurkat orCEM cells were infected with the indicated virus preparations as described in Materials and
Methods.Virus productionwasmonitoredby RTassay.
single enhancer-activator element.The results shown in Fig. 4 and 5 suggest that paired NF-KB and Spl elements are
acting in a combinatorial manner since HIV preparations
withduplications of thesameelements exhibitedanimpaired
replication phenotype in some cell lines. Moreover, the
transfection-cocultivation results presented in Fig. 1 indi-cated that the single Spl motif Spl(I) failed to restore replication competence to pNL-ZERO, suggesting that a
single Splelement couldnotdirect virus production unless it
was associated with either a second Spl element or an
NF-KB element. These conclusions all assume that the
biological activity of each of the single NF-KB or Spl
elements canbe predicted fromthe data showninFig. 1.
However, since the sequences of the differentNF-KB or
Spl motifsin the HIV-1LTRarenotidentical,weundertook a series of experiments to evaluate more critically the
functionalcontribution of individual enhancer-activator ele-mentsin thereconstructed LTRs. Toward this end, proviral DNAs containing only single NF-KB or Spl elements were
Wild type H9
Juxtaposed
6000~- y NFkB/Spl
E Separated
CL ~~~~~NFkB/Spl
>'4000
o- , ... .~~- k
0 10 20 30
DAYS POST INFECTION
FIG. 5. Infectionof H9 cells with HIV containingan LTRwith
single separated NF-KB and Spl elements. H9cells wereinfected
with theindicated viruspreparations containing pairs of homologous or heterologous motifs. Virus production was monitored by RT
assay.
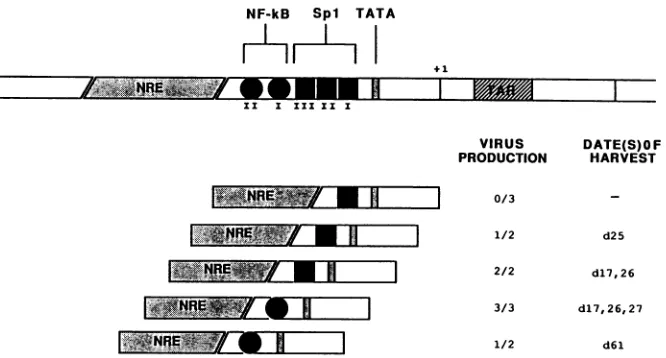
assayed for virus production by transfection-cocultivation. AsshowninFig. 6,and inagreementwith the datapresented in Fig. 1, a proviral clone containing only the Splelement Spl(I) failed toreplicate in threeindependent experiments. In contrast, the presence of the single Spl motif Spl(III) restored the capacitytoproducevirus. Splelement Spl(II) had an intermediate effect in these assays: in one
experi-ment, progeny virus was detected by day 25, while in a
second experiment, none was produced after 77 days. A
similar disparity was also observed when the two NF-KB elementswereexamined individually. NF-KB(I)was associ-ated with virus production in three separate experiments, whereas NF-KB(II) restored replicationcompetenceinonly
one oftwo independent experiments. In view of the very
long time(61 days) beforeprogenyvirions weredetected in
this latter experiment, the replication of NF-KB(II) most likely reflects a reversion event similarto that observed in the LTR sequences of other late-arising HIV variants (see nextsection). It isourcurrentbelief thatNF-KB(II) byitself cannot restore replicative capacitytothepNL-ZEROLTR.
Second-site reversions whichappearinreconstructed LTRs following virus infection of human PBLsor T-cell lines. We previously reported that second-site revertants of replica-tion-defective HIV mutants may emerge during long-term tissue culture infections(46). Consequently,theappearance
ofprogenyvirionsatverylatetimesfollowingHIV infection inthisstudyraised thepossibilitythatacompensatoryLTR alteration was responsible for the infectivity observed. We therefore focused attention onthegroup ofproviruses
con-taining only NF-KB elements in their reconstructed LTRs since they exhibited a spectrum of replicative capacities related to the cell type infected. As indicated in Fig. 2, a
hierarchy of cellular permissiveness was observed when HIVharboring onlyone ortwoNF-KB elementswas
exam-ined. PBLs and MT4cellswerethemostpermissive, allow-ingHIV containing onlya single NF-KB motifto replicate, whereas constructs containing one or twoNF-KB elements completely failed to replicate in Jurkat cells. H9 and CEM cells exhibited an intermediate phenotype: replication of virus witha single NF-KB sequence wasrestrictedin these
cells, but HIV containing both NF-KB elements was infec-tious intwooftwoexperiments inH9 cellsandone(shown
in Fig. 2C) of threeexperiments in CEM cells.
A
7 6000 E 5000'
a.
400
4-
I.-2000
C 1000'
JURKAT JE6.1
Wild type
2-Spl only A\/,\Juxtaposed
d
VCS \ ^/NFkB/SpI
IL L @ 2-NFkB only
-- L -6 -.
10
-..Iir. . . -A
aA
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.85.291.507.669.2]RECONSTRUCTED HIV LTRs 4355
NF-kB Spl TATA
+1
II
xs~~~~~~IU
I 1111 IxDATE(S)Ox
VIRUS DATE(S)0OF
PRODUCTION HARVEST
0/3
1/2 d25
2/2 d17,26
3/3 d17,26,27
[image:6.612.141.474.72.251.2]1/2 d61
FIG. 6. Replication of HIV-1 proviruses withreconstructed LTRs containing single NF-KBorSplbindingmotifs following
transfection-cocultivation. Thestructure of theHIV-1 LTRisshown diagrammatically atthetop. Inthe reconstructed LTRs,afilledsquareorcircle
indicates thepresenceofanindividualSplorNF-KB motif, respectively. NRE denotesthe negative regulatoryelement. The frequencyof
virus production(numberofRT-positivetransfections-cocultivations pernumber ofindependent experiments)and the day of virusharvest
areindicated in the columns ontheright.
Toassesswhetherthe LTRs might have undergone alter-ation during these virus infections, we performed "RNA
PCR" on thegenomic RNAs isolated from particular
sam-ples ofprogeny virions. These samples included (i)H9 cells
infected with
HIVNL-2KB
(twoNF-KBandnoSplelements),(ii) CEM cells infected with
HIVNL-2KB
(twoNF-KBand noSpl elements), and(iii) PBLs infected withHIVNLlKB(one
NF-KB andnoSpl elements).Theresultantamplified cDNA
wascloned, andthe 5' 445bpofthe U3 portionof the LTR were sequenced. In all cases, the original deletions
gener-atedin vitro remained intact.Furthermore, asshown in
Fig.
7, none of the eight cDNA clones derived from H9 cells infected with pNL-2KB contained additional changes in a
regionof the LTRextendingfromtheNF-KB elementstothe TATAA site. In contrast, 8 of 10 PCR clones of virus recovered from CEM cells infected with
HIVNL-2KB
con-NF-KB(II) NF-KcB(Q) Spl(il) Spl(II) Spl(l) TATAA
pNL4-3 ACAAGGGACT TTCCGCTGGG GACTTTCCAG GGAGGCGTGGCCTGGGCGGG ACTGGGGAGTGGCGAGCCCTCAGATGCTGC ATATAAG H9 2tNFkB ...
.... .. ... ...
CEM 2NFkBN ...
...
.-.-PB1N**skB*s
...----..-.--*-
---PBNk3.--- ----...
*. e----...
...
.T .T
.T
...
.T .T .T
.T
.T .A.T
...T
.A.T .A.T
.A.T
.A.T
..T
...T
.A.T
A.T
.-.A....
...
...A.. .
...
...
....
*... *... ...
...e
...
...A....
... ...
FIG. 7. PCRanalysis ofaportion of the U3 region of HIV LTRmutantslacking Spl bindingmotifs. The RNA associated with the cell-free
virus emerging from H9orCEM cellsinfected withHIVNL-2,B(Fig.1)orPBLsinfected with HIVNL1KB (Fig. 1)was reversetranscribedand amplifiedbyPCRasdescribedinMaterialsandMethods. The nucleotidesequencesof individual clonesweredetermined andcomparedwith thatoftheparental wild-typeHIVNL4-3shownatthetop.Identical nucleotidesareindicatedbythedots;dashes denote deletednucleotides. Nucleotide substitutionsareindicated.
A-; VOL.65, 1991
--- --- --- ---
---
--- -- --- ---
-- ---
--- ---
---
--- --- --- --- --- --- --- --- ---
--- --- --- ---
on November 10, 2019 by guest
http://jvi.asm.org/
tainedaC-to-T
change
atnucleotide-30(27),
2bp
upstream from the TATAA site. Inaddition,
this same substitution wasobservedin all 10 clonesarisingfrom the PBLinfection with HIVNL 1KB virus and wasaccompanied
by a secondchange (T
to A) at nucleotide -32 in 8 of these 10 clones. Some of theamplified
LTRclones hadnoalterations in otherregions
of the LTR; in the othersexamined,
a maximum of twoadditionalchanges
perLTRwerenoted,
noneof which were clustered in aspecific region.
DISCUSSION
In our
previous studies,
a series of deletion and pointmutationswere introduced into the HIV LTR to
study
the rolesof the NF-KB andSpl
binding
sites and the TARregion
in virus
production
(23).Deletion of thetwoNF-KBsites did not render theresulting
proviral
DNAreplication
incompe-tent. The removal of the three
Spl binding
sites withpreservation
ofthe NF-KBbinding
sites resulted in aviruscapable
ofreplicating
in humanPBLsbutnotincellslacking
NF-KB
binding activity
(30).Furthermore,
thesimultaneousexcision of both NF-KB and all three
Spl binding
siteseliminatedthe
capacity
of HIVtoreplicate
inhumanTcells. These observations suggested that discretebinding
sites in the LTRfunctionasmodularunits,
called enhansons(10, 29,43),
which canfunctionally
substitute for one another and whoseimportance
toviralreplicationvaries in differentcelltypes
depending
on the nature of cellulartranscriptional
factors present in those cells. In the presentstudies,
weattempted
to define minimal sequencesrequired
to support viralreplication
in a variety of human T lymphocytes and furthercharacterizedthenatureofthe enhansons present in the HIVpromoter-enhancer
sequences.Our efforts to determine minimal
promoter-enhancer
se-quencesrequired
forviralreplication
wereundertakenusingareconstructive
approach.
Theinsertion ofthesingle
NF-KB elementNF-KB(I)
intopNL-ZERO
was sufficientto restore HIVreplication
in CEM-MT4cocultures;
in contrast,NF-KB(II)
didnot restorereplication
competence. InanalogousLTR reconstruction
experiments involving
individualSpi
motifs,
acontinuum ofreplicative capacities
was observed:Spl(III)
>Spl(II)
>Spl(I).
This latter result is consistentwith reports
showing
that theSpl(I)
element has loweraffinity
forSpl protein (20)
andfor factorspresentin crude nuclearextractsof humanlymphocytes
(35)than doeseitherelement IIorIII. The
ability
of HIVpreparations
containingonly
Spl(III)
to infect human T cells is currently under evaluation.Asneitherthe NF-KBnor
Spl binding
sitesareabsolutelyrequired
forvirusreplication
in PBLs(aslongasthe otheris presentin the LTR), the marked conservation ofthe pres-ence and arrangement of these two types ofcis-acting
sequences in all known HIV-1 isolates is puzzling. The
experiments
shown inFig.
4 and 5 suggest that thearrange-mentand
spacing
ofadjacent
NF-KBandSpl bindingsites in thenativeconfiguration
ofthe HIV-1 LTR contribute moresignificantly
to virus replicationthan duplications ortripli-cations of either element. In this regard, we previously reported that point mutations affecting theadjacent NF-KB and
Spl
elements resulted inasignificantreductionof virusreplicative
capacity
(23). Thus, two independent studies suggest the existence ofafunctional interaction(s) between these two different cis-acting elements and the proteins whosefunctions theymediate.Notably, NF-KB binding sitesare
frequently
situated adjacent to binding sites forconsti-tutive cellular
transcriptional
factors. Onestriking exampleis in the promoter of the cytomegalovirus immediate-early gene, in which a series of NF-KB binding sites are inter-spersed with the binding sites for NF-1 (12, 16). When these elementswereplacedadjacenttoeachother, 5' tothewhey acidic protein promoter, a synergistic enhancement of pro-moteractivitywasobserved, suggestingthat in thiscontext, NF-KB wasabletointeract with factorsbindingtothe NF-1 sitetoaugmenttranscription (17). Likewise, similar studies have been performed to study the interactions between
adjacentbindingsites forSpland OTF-1. AnSplelement in combination withahigh-affinityOTF-1 siteaugmented
tran-scription
2-fold; however,whenplacednext to alow-affinityOTFsite, the Spl site increased transcription 20-fold (19). Thus, as both NF-KB and Spl caneach interact with other
transcription factors to augment transcription, it is not
surprising
thattheymayinteract with each other in the HIV LTRto promoteviralreplication.An interesting aspect of this analysis is the apparent
hierarchyof cellularpermissivenessfor HIV infectionseen best with proviruses containing only one or two NF-KB elements. The order observed(PBLs = MT4> H9> CEM > Jurkat cells) is consistent with our previous findings (PBLs= MT4>CEM)withanHIV LTRmutantcontaining a wild-type arrangement of NF-KB motifs but lacking all three Spl sites and presumably reflects various levels of
cellular factors
capable
ofbinding
toNF-KB(30).
Asshown in Fig. 2 of this study, virus containing asingle
NF-KB elementwascapableofinfectingPBLs and MT4 cells (data notshown)exclusively.
At theother extreme, HIV withtwo NF-KB elements failed to replicate in Jurkat cells. This continuum of cellularsusceptibilitywasalso reflected in the number and typesofsecond-siterevertants whichappearedduring
infection. H9 cellsconsistently supported replicationof HIV with two NF-KB elements; progeny virions from suchaninfection containedanunaltered LTR
(Fig.
7).This same virus inoculum produced progenyvirions in onlyone of threeindependent
infections ofCEM (A3.01) cells. PCRanalysis
revealedaconsistent second-site mutationin 8 of 10 cDNA clonesarising
from this single positive experimentthatwas localizedtothe TATAregion ofthe LTR.Finally,
the HIV construction with a single NF-KB binding site
(NL-1KB) replicated slowly in PBLs but, in the process,
acquired
twochanges
also situatedimmediately
upstreamof the TATA box. Thus, a selection for virionscontaining
a similar TATAmutationis operativeinbothCEMcells andPBLs, possibly
reflecting
the 40-bp deletionencompassing
the three Spl sites which effectively places the NF-KB(I)
motif 13
bp
upstreamofthe TATAbox. This TATA altera-tion is consistent with reports describing the restricted interaction between a cell-specific enhancer and discreteVIRUS
UILD TYPE
CE11REUERTRNT
POLREUERTRNT
TATR BOX
TGCTGCF|iG
TGCTG ATATAG
TGCRG TATAG
FIG. 8. TATAboxrevertants which appearfollowing infection of CEM and activated PBL cells with HIV containing only an
NF-KBmotif(s)in theNF-KB/Spl domain.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.373.520.588.687.2]TATA sequences (39, 42, 44). Perhaps the alteration of the wild-type HIV TATAA box to TATATAA is necessary for functionalinteraction with the dislocated NF-KB element(s). It is worth noting that the base change common to both second-site revertant viruses extended the TATA box two nucleotides in the 5' direction relative to that of the parental virus pNL4-3 (Fig. 8). Since the TATA element positions where RNA polymerase II initiates RNA synthesis, this change could shift the transcriptional start site from its normal location at the U3/R border. Experiments to ascer-tain whether the transcriptional start site has been altered in the LTR revertant viruses and to assess the effects of such TATA changes on HIV replication are presently in progress.
ACKNOWLEDGMENT
The following reagent was obtained from the AIDS Research and Reference Reagent Program, Division of AIDS, National Institute of Allergy and Infectious Diseases: Jurkat clone E6-1 from the Amer-ican Type Culture Collection.
REFERENCES
1. Adachi, A., H. E. Gendelman, S. Koenig, T. Folks, R. Willey, A. Rabson, and M. A. Martin. 1986. Production of acquired immu-nodeficiency syndrome-associated retrovirus in human and non-human cells transfected with an infectious molecular clone. J. Virol. 59:284-291.
2. Ahmad, N., and S. Venkatesan. 1988. Nef protein of HIV-1 is a transcriptional repressor of HIV-1 LTR. Science 241:1481-1485.
3. Arya, S. K., C. Guo, S. F. Josephs, and F. Wong-Staal. 1985. Trans-activator gene of human T-lymphotropic virus type III (HTLV-III). Science 229:69-73.
4. Berkhout, B., and K.-T. Jeang. 1989. trans activation of human immunodeficiency virus type 1 is sequence specific for both the single-stranded bulge and loop of the trans-acting-responsive hairpin: a quantitative analysis. J. Virol. 63:5501-5504. 5. Berkhout, B., R. H. Silverman, and K.-T. Jeang. 1989. Tat
trans-activates the human immunodeficiency virus through a nascent RNA target. Cell 59:273-282.
6. Durand, D. B., J.-P. Shaw, M. R. Bush, R. E. Replogle, R. Belagaje, and G. R. Crabtree. 1988. Characterization of antigen receptor response elements within the interleukin-2 enhancer. Mol. Cell. Biol. 8:1715-1724.
7. Dynan, W. S., and R. Tjian. 1983. The promoter-specific tran-scription factor Spl binds to upstream sequences in the SV40 early promoter. Cell 35:79-87.
8. Feng, S., and E. C. Holland. 1988. HIV-1 tat trans-activation requires the loop sequence within tar. Nature (London) 334: 165-167.
9. Folks, T., S. Benn, A. Rabson, T. Theodore, M. D. Hoggan, M. Martin, M. Lightfoote, and K. Sell. 1985. Characterization of a continuous T-cell line susceptible to the cytopathic effects of the acquired immunodeficiency syndrome (AIDS)-associated retro-virus. Proc. Natl. Acad. Sci. USA 82:4539-4543.
10. Fromental, C., M. Kanno, H. Nomiyama, and P. Chambon. 1988. Cooperativity and hierarchical levels of functional orga-nization in theSV40 enhancer. Cell 54:943-953.
11. Garcia, J. A., D. Harrich, E. Soultanakis, F. Wu, R. Mitsuyasu, and R. B. Gaynor. 1989. Human immunodeficiency virus type 1 LTR TATA and TAR region sequences required for transcrip-tional regulation. EMBO J. 8:765-778.
12. Ghazal, P., H. Lubon, B. Fleckenstein, and L. Hennighausen. 1987. Binding of transcription factors and creation of a large nucleoprotein complex on the human cytomegalovirus en-hancer. Proc. Natl. Acad. Sci. USA84:3658-3662.
13. Guyader, M., M. Emerman, P. Sonigo, F. Clavel, L. Montagnier, and M. Alizon. 1987. Genome organization and transactivation of the human immunodeficiency virus type 2. Nature (London) 326:662-669.
14. Harrich, D., J. A. Garcia, R. Mitsuyasu, and R. B. Gaynor. 1990. TAR independent activation of the human
immunodefi-ciency virus in phorbolesterstimulatedTlymphocytes. EMBO J. 9:4417-4423.
15. Harrich, D., J.A.Garcia, F. K. Wu, R. Mitsuyasu, J.Gonzales, and R. B. Gaynor. 1989.Role ofSpl-bindingdomains in in vivo transcriptionalregulation of the humanimmunodeficiencyvirus type 1 long terminalrepeat. J.Virol. 63:2585-2591.
16. Hennighausen, L., and B. Fleckenstein. 1986. Nuclear factor 1 interacts withfiveDNAelements in the promoterregion ofthe humancytomegalovirus major immediateearlygene.EMBO J. 5:1367-1371.
17. Hennighausen, L., P. A. Furth, and C. W. Pittius. 1989. KB elements strongly activate gene expression in non-lymphoid cells andfunction synergistically withNF1 elements. Nucleic AcidsRes. 17:8197-8206.
18. Jakobovits,A., D. H.Smith, E. B.Jakobovits,and D.J.Capon. 1988. Adiscrete element 3'ofhumanimmunodeficiencyvirus 1 (HIV-1) and HIV-2 mRNA initiation sites mediates
transcrip-tional activation by an HIV trans activator. Mol. Cell. Biol. 8:2555-2561.
19. Janson, L., and U. Pettersson. 1990. Cooperative interactions between transcription factorsSplandOTF-1.Proc.Natl.Acad. Sci. USA87:4732-4736.
20. Jones, J. A., J. T.Kadonga, P. A. Luciw, andR. Tijan. 1986. Activation of the AIDS retrovirus promoter by the cellular transcriptionfactor, Spl. Science 232:755-759.
21. Kawakami, K., C. Schneidereit, and R.G. Roeder. 1988. Iden-tification andpurificationofahumanimmunoglobulin enhancer-binding protein (NF-KB) that activates transcription from a
humanimmunodeficiency virus type 1 promoter in vitro. Proc. Natl. Acad. Sci. USA 85:4700-4704.
22. Kunkel, T. A. 1985.Rapid andefficientsite-specificmutagenesis without phenotype selection. Proc. Natl. Acad. Sci. USA 82:488-492.
23. Leonard, J., C.Parrott, A. J. Buckler-White, W.Turner,E. K.
Ross, M. A.Martin, and A. B. Rabson. 1989. The NF-KBbinding
site in the human immunodeficiency virus type 1 long terminal repeat are not required for virus infectivity. J. Virol. 63:4919-4924.
24. Lu, Y., M. Stenzel, J. G. Sodroski, and W. A. Haseltine. 1989. Effects of longterminalrepeatmutationsonhuman immunode-ficiency virus type 1 replication. J. Virol.63:4115-4119. 25. Miyoshi, I., H.Taguchi, I. Kubonishi, S.Yoshimoto, Y.Ohtsuki,
F. Shiraishi, and T. Akagi. 1982. Type c virus-producing cell linesderived from adult T-cell leukemia. GannMonogr. 28:219-228.
26. Morgan, R. A., K. Cornetta, and W. F. Anderson. 1990.
Appli-cations of thepolymerase chain reactionin retroviral-mediated gene transfer and theanalysis ofgene-markedhuman TILcells. Hum.Gene Ther. 1:135-149.
27. Myers, G., A. B. Rabson, J. A. Berzofsky, T. F.Smith, and F.
Wong-Staal(ed.). 1990. Human retroviruses and AIDS 1990, p. I-A-64. Theoretical Biology and Biophysics, Los Alamos Na-tionalLaboratory, Los Alamos, N.Mex.
28. Nabel, G., and D. Baltimore. 1987. An inducible transcription
factoractivates expression ofhumanimmunodeficiencyvirus in Tcells. Nature(London) 326:711-713.
29. Ondek, B., L. Gloss, and W. Herr. 1988. The SV40 enhancer contains two distinct levels oforganization. Nature (London) 333:40-45.
30. Parrott, C., T. Seidner, E. Duh, J.Leonard, T. S.Theodore,A.
Buckler-White, M.A.Martin, andA. B.Rabson. 1991. Variable role of the long terminal repeat Spl-binding sites in human immunodeficiency virus replicationin Tlymphocytes. J. Virol. 65:1414-1419.
31. Popovic, M., M. G. Sarnagadharan, E.Read, andR. C. Gallo. 1984. Detection, isolation, and continuous production of cyto-pathic retroviruses (HTLV-III) from patients with AIDS and pre-AIDS. Science 224:497-500.
32. Rosen, C. A., J. G. Sodroski, and W. A. Haseltine. 1985. The location ofcis-acting regulatory sequences in the human T cell lymphotrophic virus type III (HTLV-III/LAV) long terminal repeat. Cell 41:813-823.
33. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular
on November 10, 2019 by guest
http://jvi.asm.org/
cloning: alaboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
34. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
35. Seidner, T. J., and A. B. Rabson. Unpublished data.
36. Selby, M. J., E. S. Bain, P. A. Luciw, and B. M. Peterlin. 1989. Structure,sequence,and position ofthe stem-loopintar deter-minetranscriptional elongation by tat through the HIV-1 long terminalrepeat. Genes Dev. 3:547-558.
37. Shaw, J.-P., P. J. Utz, D. B. Durand, J. J. Toole, E. A.Emmel, andG. R.Crabtree. 1988. Identificationofaputativeregulator of earlyTcellactivation genes. Science 241:202-205.
38. Siekevitz, M., S. F. Josephs, M. Dukovich, N. Peffer, F. Wong-Staal, and W. C. Greene. 1987.Activationof the HIV LTR by T cellmitogens and the trans-activator protein of HTLV-1. Sci-ence238:1575-1578.
39. Simon, M. C., T. M.Fisch, B. J. Benecke, J. R. Nevins, and N. Heintz. 1988.Definition ofmultiple, functionally distinct TATA elements, one of whichisa target in the hsp7Opromoter for ElA regulation. Cell 52:723-729.
40. Sodroski, J., C. Rosen, F. Wong-Staal, S. Z. Salahuddin, M. Popovic, S. Arya, R. C. Gallo, and W. A. Haseltine. 1985. Trans-acting transcriptional regulation of human T-cell leuke-mia virustype III longterminal repeat. Science 227:171-173. 41. Sodroski, J. G., C. A. Rosen, F. Wong-Staal, R. Patarca, and
W. A. Haseltine. 1985. Location of the trans-activating region on the genome of human T-cell lymphotropic virus type III. Science 229:74-77.
42. Struhl, K. 1986. Constitutive and inducible Saccharomyces cerevisiae promoters:evidencefor two distinct molecular mech-anisms. Mol. Cell. Biol. 6:3847-3853.
43. Tanaka, M., U.Grossniklaus, W. Herr, and N. Hernandez.1988. Activation ofthe U2 snRNA promoter by the octamer motif definesanewclassofRNApolymeraseIIenhancer elements. GenesDev.2:1764-1788.
44. Wefald, F. C., B. H. Devlin, and R. S. Williams. 1990. Func-tional heterogeneity of mammalian TATA-box sequences re-vealed byinteraction withcell-specificenhancer. Nature (Lon-don) 344:260-262.
45. Weiss, A. L., R. L. Wiskocil, and J. D. Stobo. 1984. Therole of T3 surface molecules in the activation of human T cells: a two-stimulus requirementfor IL 2 production reflects events occurringat apre-translational level.J. Immunol. 133:123-128. 46. Willey, R., D. H. Smith, L. A. Lasky, T. S. Theodore, P. L. Earl, B. Moss, D. J. Capon, and M. A. Martin. 1988. In vitro mutagenesis identifiesaregion within the envelopegeneofthe humanimmunodeficiency virus that is criticalforinfectivity. J. Virol. 62:139-147.
47. Zoller, M. J., and M. Smith. 1983. Oligonucleotide-directed mutagenesis: asimple methodusing twooligonucleotide prim-ersandasingle-stranded DNAtemplate. DNA 3:479-488.