in root and nodule development
Karsten Oelkers
A thesis submitted for the degree of
Doctor of Philosophy
of The Australian National University
College of Medicine, Biology and Environment
Division of Plant Science, Research School of Biology
Declaration of originality
I declare that this thesis reports the results of my own original work, performed while
being enrolled in the Doctor of Philosophy program at The Australian National
University, that no part has been previously accepted or presented for the award of
any degree from any university, and that, to the best of my knowledge, no material
previously published or written by any other person is included, except where due
acknowledgement is given.
I am most grateful to my mentor Ulrike Mathesius for the opportunity to pursue this thesis project under her supervision and in her laboratory at The Australian National University, and to attend several national and international conferences enabling me to meet some of the most inspiring people in the plant sciences. Also, I would like to express my gratitude for her personal encouragements and motivation as well as her patience and calmness. In my thesis project Uli gave me the freedom that I wanted, but also the support that I needed, and I acknowledge her active involvement and many suggestions throughout the doctoral candidature as well as her expert correction and proof-reading of this thesis.
My co-supervisor Peter Gresshoff is acknowledged for his genuine, father-like support, interest in and promotion of the thesis project, and for inviting me to oral presentations at annual symposia of the Australian Research Council Centre of Excellence for Integrative Legume Research as a graduate student among more senior academic staff. I thank Georg Weiller Michael Djordjevic and Nijat Imin, who were cooperative advisors.
Nicolas Goffard and Tancred Frickey are credited for advice in the bioinformatics (Chapters Two and Four), motivation for and assistance in learning PERL, decreasing the steep slope of the LINUX learning curve, unforgettable surfing trips and the scientific inspiration shaping my evolution to become a professional geek.
Emlyn Williams and Terry Newman from the ANU Statistical Consulting Unit assisted with statistical analysis required for Chapter Three. Virginie Mortier, Annick De Keyser, Sofie Goormachtig and Marcelle Holsters hosted me at the PSB/VIB in Ghent/Belgium for three weeks and I acknowledge their cooperation teaching me in situ hybridisations (Chapter Four). I owe special thanks to Laure Yassin for her voluntary help in scoring the first round of the peptide assay (Chapter Three). I enjoyed many inspiring conversations with members of Bany Pogson's lab, particularly with Matthew Gordon, who gave me expert advice on taming Roche's LightCycler (Chapter Four) as well as Gonzalo Estavillo, who recommended
acknowledged for sharing their excellent RNA extraction protocol (Chapter Four).
The numerous present and past colleagues and visitors of the Mathesius Lab (Quinton Pratt, Qui
Yu Peter Dung, Jin Jian, Samira Hassan, Melinda Aprelia, Chooi Hua Goo, Corinna Paeper,
Giel van Noorden, Flavia Pellerone, Anton Wasson, Britta Winterberg, Tursun Kerim, Fay
Khaw, Diana Kirk, Sofia Costa, Paul Wiley and particularly Belinda Schiller as well as
Cassandra Harris) are appreciated for the working atmosphere in the lab, scientific input during
lab meetings and for actively helping to measure peptide bioassays (Chapters Three and Four).
Farzaana Thanweer and Bee Doolan were valued office colleagues and enlivened the daily
routine. Also, I thank the general and academic staff of the School of Biochemistry and
Molecular Biology/Research School of Biology for providing a productive framework, and
making the School an inspiring place to work.
My trusted friends Joana Moscoso, Klementina Kakar, Dana Rich, Andrew Elborn, Andrew
Haese, Jaime Swift, Kelly Roe, Ryan Taylor and Amir Hadad have been close to my heart and I
am grateful for their support over the years. Also, I would like to acknowledge my friends from
Burton and Garran Hall, University House, Graduate House, Fellows Garden, the ANU Scuba
Club and the Canberra Handball Club for a lot of good memories.
Above all, I am grateful to my parents Beatrice and Hans-Joachim, and my brothers Nils and
Rufus for their unconditional love and support in every respect beyond the boundaries of
geography.
°C degrees Celsius
ACR4 A. thaliana crinkly-like 4 ANOVA analysis of variance
Mb mega basepair minutes
mRNA messenger ribonucleic acid
ANU The Australian National Mt
University
AON autoregulation of nodulation n AP alkaline phosphatase N
Medicago truncatula
sample size
molar equivalents per litre
At Arabidopsis thaliana NCBI national centre for biotechnology information ATP AU BLAST BMM BMRF Bn bp cDNA cf. adenosine 5'-triphosphate absorption unit
basic local aligment and search tool Bergensens medium Biomolecular Facility Brassica napus basepair complimentary deoxyribonucleic acid compare nd ND nm NOD modified
Resource nork
nr ORF Os not determined nodule density nanometre nodule number
nodulation receptor kinase
non-redundant open reading frame
Oryza sativa
CLE CLAVATA3/Endosperm Surrounding Reagion
CLV CLAVATA
CN cyst nematode
cp crossing point
crn coryne ct cycle threshold Da Dalton
DEPC diethylpyrocarbonate DF degree of freedom
DIG digoxigenin
dmi doesn't make infection DNA deoxyribonucleic acid Dnase deoxyribonuclease dNTP deoxy
dpi days after inoculation dT deoxythymine E estimate, efficiancy, expect
pH Pi POD PSI-BLAST potentia Hydrogenii isoelectric point seed pod
position specific iterative basic
local alignment search tool qRT-PCR quantitative real time
polymerase chain reaction R reverse/antisense RAM root apical meristem RELGRO relative root grwth REST Relative RKN root-knot nematode RLK receptor-like kinase RNA ribonucleic acid Rnase ribonuclease RO root organ number rpm revolutions per minute RT room temperature
RT-PCR reverse transcriptase polymerase chain reaction
e.g. exempli gatia
EDTA ethylenediaminetetraacetic acid
ENOD early nodulin gene
s second SAM shoot apical meristem
region
EST expressed sequence tag Std
et al. et alii/et aliae sunn
F forward/sense symrk
g gravity force/gram TA
GAPDH Glycerine aldehyde 3- Taq phosphate dehydrogenase
gDNA genomic deoxyribonucelic TDIF
acid
GERM germination root length TE
GFP green fluorescent protein TIGR
Gm Glycine max Tm
GRO root growth Tris
h hour UBPL7
harl hypernodulation abberant UTP
root 1
Hg Heterodera glycines vs.
HMM hidden Markov model WOX
i.e. id est wus
ISH in situ hybridisation
KAPP kinase-associated GSP
protein prhopshatase
kb kilobasepair HK standard
super nummeric nodules
symbiosis receptor kinase
transcript assembly
DNA Polymeraae from Termophilus
aquaticus
tissue differentiation factor
Tris-EDTA
the institute for genome research
melting temperature
tris(hydroxymethyl)aminomethane
Ubiquitin Protein Ligase 7
versus
wuschel-related homeobox
wuschel
gene-specific primer
LAT lateral root number LSD least significant difference LB Lysogeny Broth
LRJD lateral root density
LysM m
lysine motif meter
Single letter code Three letter code Amino acid
A Ala Alanine
B Asx Aspartic acid or Asparagine
C Cys Cysteine
D Asp Aspartic Acid
E Glu Glutamic Acid
F Phe Phenylalanine
G Gly Glycine
H His Histidine
I lie Isoleucine
K Lys Lysine
L Leu Leucine
M Met Methionine
N Asn Asparagine
P Pro Proline
Q Gin Glutamine
R Arg Arginine
S Ser Serine
T Thr Threonine
V Val Valine
W Trp Tryptophan
X Xaa Any
Y Tyr Tyrosine
Abbreviation Base or Nucleotide
A Adenine
C Cytosine
G Guanine
T (or U) Thymine (or Uracil)
N Any
Genetic nomenclature
Genetic nomenclature is held according to Medicago truncatula and plant science
community conventions (VandenBosch and Frugoli, 2001, Leyser and Day, 2003,
Meinke and Koornneef, 1997).
Wild type allele, phenotype, mutation or gene SUNN-4
Mutant allele, phenotype, mutation or gene sunn-4
Wild type protein SUNN-4
The hypothesis of the thesis project postulates that CLV3/ESR-like (CLE) signalling
peptides are involved in the control of nodule number. At the start of the project, only
seven CLE members from legumes had been identified. To characterize CLE domain
proteins functionally and to test for their involvement in the autoregulation of
nodulation it was necessary to identify more members from this CLE family.
More than 100 new CLE signalling peptides were identified in the first part of the
project, including five protein sequences containing multiple, regularly arranged CLE
motifs. It was possible to cluster the CLE domain proteins into 13 groups based on
pairwise similarities in the primary CLE motif. In addition, secondary motifs were
identified, which could lead to the discovery of recognition sites for processing
peptidases. The groupings of the CLE motifs correlate with known biological functions
of CLE signalling peptides and are analogous to previously established groupings based
on phylogenetic analysis and ectopic overexpression studies. The biological function of
two predicted CLE signalling peptides was tested in Medicago truncatula in vivo using
a peptide bioassay. These peptides inhibited the cell differentiation at root apical and
lateral root meristems in a manner consistent with functional predictions based on other
CLE signalling peptides clustering in the same groups.
In the second part of the thesis, the possible involvement in nodulation of a selection of
identified CLE signalling peptides was tested in peptide bioassays using two Medicago
truncatula genotypes, wild type as well as the supernodulating mutant sunn, by
supplying the growth medium with synthetic CLE peptides. Initial pilot tests involving
root number were observed in response to peptide treatment, but were not statistically
significant. Nevertheless, two lateral root architecture phenotypes "spiky root" and
"melty root" were observed and described. There are indications for a common peptide
motif involved causing the phenotypes. The initially promising results of specific
effects of CLE peptides on nodule number were not repeatable with statistical
significance. Generally, variation in the number of nodules could be detected in the
Petri dish-based peptide bioassay and did not allow a measurement of the effects of
individual CLE signalling peptides.
Several protein sequences containing multiple CLE domains were identified in the first
part of the project. CLE68 is such a multi-CLE domain protein encoded by Medicago
truncatula. To ensure that CLE68 is encoded as predicted and not an artefact of
erroneous genome annotation the genomic locus was sequenced. Quantitative real-time
polymerase chain reaction was used to demonstrate that CLE68 is expressed in vivo.
The functional ortholog of Arabidopsis thaliana CLV3 in Medicago truncatula is
described in detail and expression is analyzed. Examination of the genomic locus
demonstrates that CLE68 is a paralog of the functional ortholog of CLV3 in Medicago
truncatula. Peptide bioassays demonstrate that CLE68 is unique, as it contains CLE
peptides with differential effects when ectopically applied in a root apical meristem
(RAM) peptide bioassay. Members of the CLE signalling peptide family containing
several CLE domains within a single protein precursor instead of commonly a single
1. Karsten Oelkers, Nicolas Goffard, Georg F. Weiller, Peter M. Gresshoff, Ulrike
Mathesius, Tancred Frickey "Bioinformatic analysis of the CLE signalling peptide
family" BMC Plant Biology (2008) 8:1
2. Karsten Oelkers, Nicolas Goffard, Georg F. Weiller, Peter M. Gresshoff, Ulrike
Mathesius, Tancred Frickey "Erratum: Bioinformatic analysis of the CLE signalling
Declaration of originality I Acknowledgements II Abbreviations IV Abstract X Publications XII
Chapter One: General introduction 1
1.1. Plant genomes and plant signal transduction 1 1.1.1. Peptide hormones and biologically active peptides 2
1.1.1.1. Plant signalling peptides 3 1.1.1.1.1. CLE signalling peptides 4
1.1.1.1.1.1. The CLA VA TA genes of Arabidopsis thaliana 7
1.1.1.1.1.2. Discovery and identification of CLE signalling peptides 7 1.1.1.1.1.3. Primary sequence characteristics of the CLE family 10 1.1.1.1.1.4. The receptor-binding of CLE signaling peptides 10 1.1.1.1.1.5. Posttranslational processing: cleavage and covalent modifications 10
1.1.1.1.1.6. Physiological function of CLE members in plants 11
1.1.1.1.1.7. Embryo Surrounding Region genes 12 1.1.1.1.1.8. The stem cell niches around CLAVATA3 and CLE40 12
1.1.1.1.1.9. CLE signalling peptides and biotechnological applications 15
1.1.2. Receptor-like kinases 16 1.1.2.1. Modes of action of plant receptor-like kinase 16
1.1.2.2. Plant receptor-like kinase family 17 1.1.2.3. Plant leucine-rich repeat receptor-like kinases 20
1.2. Nitrogen and symbiosis 21 1.2.1. The model legume Medicago truncatula 22
1.2.2. The root nodule symbiosis 23 1.2.3. Essential steps for the formation of nodules 24
1.2.3.1. Plant flavonoids induce bacterial Nod factor biosynthesis 25
1.2.3.4. The development of a nodule 31 1.2.4. Phytohormones in nodule formation 32
1.2.5. Restriction of nodule formation 36 1.2.6. Leucine-rich repeat receptor-like kinases in nodule development 38
1.3. The evolutionary emergence of the rhizobia-legume symbiosis 39
1.3.1. Arbuscular mycorrhiza 39 1.3.2. Plant nematode interactions 40 1.3.2.1. Nematode parasitism and root nodule symbiosis 40
1.3.2.2. Nematode CLE signalling peptides 41 1.3.2.3. Nematode resistance locus rhgl 42 1.4. Is there a role for CLE signalling peptides in symbiosis? 43
1.4.1. Hypothesis and aim of the thesis project 45
1.4.2. Research proposal 45
Chapter Two: Bioinformatic analysis of the CLE signalling peptide family 47
2.1. Abstract 47 2.2. Introduction 48
2.2.1. Experimental approach 49 2.3. Identification of CLE signalling peptides 52
2.3.1. Protein sequence database from various plants 52
2.3.2. Query sequences 52 2.3.3. Motif search in protein sequence databases 53
2.3.4. Sequence analysis 61 2.4. CLE members with multiple and regularly arranged CLE domains 61
2.5. Sequence analysis and identification of novel sequence characteristics 65 2.6. Biological function of identified CLE signalling peptides in M. truncatula 70
2.7. Erratum 74 2.8. Discussion 76
treatment 81
3.1. Abstract 81 3.2. Introduction 82
3.2.1. Experimental approach 83 3.3. Pilot peptide bioassays on nodulation 87
3.3.1. Interim conclusion I 93 3.4. Peptide map and selection of sequences 94
3.5. Screen for CLE peptides involved in root nodulation and architecture 97
3.5.1. Design of experiments, terms and treatments 97
3.5.2. Data analysis 99 3.5.2.1. Incubator bias 100 3.5.2.2. Qualitative and quantitative effects summary 102
3.5.2.3. Root length 103 3.5.2.4. Lateral root number 103
3.5.2.5. Nodule number 104 3.5.2.6. Assessment of response variable adjustment 105
3.5.3. Interim conclusion II 111 3.6. Variation of nodule number in Petri dish based bioassays 111
3.6.1. Interim conclusion III 114 3.7. Qualitative observations of changes in lateral root architecture 115
3.7.1. Attempts of automated detection of root architecture 116
3.7.2. Interim conclusion IV 117
3.8. Discussion 120
Chapter Four: CL(u)Es on multi-CLEs: Origin and function of proteins
containing multiple CLV3/ESR-motifs 122
4.1. Abstract 122 4.2. Introduction 122
4.2.1. Identification of MTCLE68 and M/CLV3 123 4.3. Sequence similarity between CLV3s and M/CLE68 124
4.4. Sequencing of the MtCLE68 genomic locus 130
4.5. Expression analysis of MtCLV3 and MtCLE68 131
4.5.2. Quantitative real-time PCR 132
4.5.2.1. Method establishment 132 4.5.2.2. Relative quantification results 134
4.5.3. In situ detection of MtCL V3 transcripts 138
4.6. Motif similarity and peptide assay 140
4.7. Discussion 143 4.7.1. Evidence for biological functions of multi-CLE domain proteins 143
4.7.2. The genetic origin of MtCLE68 as a paralog of MtCL V3 144
4.7.3. The biological function of MfCLE68 144 4.8. Accessions, nucleic acid and protein sequences 147
4.8.1. M. truncatula CLE68 147 4.8.2. M. truncatula CLAVATA3 148
Chapter Five: General discussion 149
Chapter Six: Material and methods 152
6.1. Biological sequence resources 152 6.2. Motif search methodology 153 6.3. Biological material 155
6.3.1. Plant material 155 6.3.2. Seed propagation and green house plant growth 155
6.3.3. Bacterial strains 156
6.4. Growth media 156 6.5. Seed germination 158 6.6. Controlled environment incubator 159
6.7. Plant growth 160 6.8. Plant peptide treatment 160
6.8.1. Peptide synthesis 160 6.8.2. Peptide stock solutions 161 6.8.3. Time points of peptide treatment and inoculation with S. meliloti 162
6.9. Plant measurements 162 6.10. Design of experiments and statistical analysis 163
6.13. Extraction of nucleic acids 164 6.13.1. gDNA extraction 164 6.13.2. Total-RNA extraction 165 6.14. DNase treatment of RNA samples 167 6.15. RNA Clean-up after DNase treatment 167
6.16. cDNA synthesis 167 6.17. Oligonucelotide primer 167
6.17.1. Sequence design 167 6.17.2. Oligonucelotide primer list 168
6.18. Polymerase chain reaction 170 6.18.1. Standard amplification of DNA 170
6.18.2. Quantitative real-time polymerase chain reaction 171
6.18.2.1. Setup of qRT-PCR 171 6.18.2.2. Preparation of qRT-PCR 172 6.18.2.3. Analysis of qRT-PCR 174 6.19. Agarose gel-electrophoresis 175
6.19.1. Gel extraction 175 6.19.2. Sequencing 175
6.20. RNA in situ hybridization 175
6.20.1. Tissue preparation and pre-treatment 176
6.20.2. RNA probe synthesis 176 6.20.3. Tissue pre-treatment 177 6.20.4. Probe hybridization and detection 177
1.1. Plant genomes and plant signal transduction
There are elementary differences in the plasticity of development and basic metabolism
between the autotrophic, immobile lifestyle of plants and the mobile heterotrophic
animals. Thus, plant genomes are required to encode a more versatile genetic
framework to enable the adaption over a wider range of constantly changing
environmental conditions than animal genomes (Leyser and Day, 2003). These aspects
are also manifested in signal transduction networks, which are fundamentally different
between plants and animals, although sharing the same underlying molecular
mechanisms (Cock, 2002). Nevertheless, detailed knowledge in plant signal
transduction compared with animals is still in its infancy (Becraft, 2002). This could
partially be due to the medical relevance but also the occurrence of neural networks
with electrical signal transmission and centralized signalling organs in animals such as
hormonal glands (Busch and Benfey, 2010). Therefore, research in plant signal
transduction has traditionally been focussed on plant growth regulators or classic
phytohormones originating from secondary metabolism, which include auxins,
cytokinins, ethylene, jasmonic acid, gibberellins, abscisic acid, brassinosteroids,
polyamins, salicylic acid, strigolactones and flavonoids. For a discussion of classic
phytohormones, the reader is referred to the corresponding section reviewing their
involvement in nodule formation. However, advances in molecular biology and
particularly the completion of several plant genome sequence projects have established
an invaluable resource that gives insight into the fundamental differences in the
plasticity and regulation of multicellular development of plants compared with animals
Lijsebettens give an excellent overview over the history of technical advances in
combination with milestone achievements in plant developmental biology (van
Montagu, 2005, van Lijsebettens and van Montagu, 2005). The introduction to Chapter
Two will give some examples of genome sequence facilitated identification of
developmentally relevant gene families.
1.1.1. Peptide hormones and biologically active peptides
Peptide signalling occurs in many physiological processes across all domains of life.
Examples outside the plant kingdom include venomous peptides (scorpion, spider,
snake, sea anemone), defence peptides (antibiotic, antifungal) and numerous others. For
an extensive overview the reader is referred to the "Handbook of Biologically Active
Peptides" (Kastin, 2006), which reviews biologically active peptides across all
kingdoms of life. Historically, insulin has not only been a vanguard in the research area
of peptide hormones, but also land marking milestone achievements in the scientific
field of biology. Insulin was naturally isolated as the first peptide hormone by John
James Richard McLeod, Charles Best and Frederick Banting (Banting and Best, 1922).
Furthermore, it was the first determined protein sequence (Stretton, 2002, Sanger and
Tuppy, 1951b, Sanger and Tuppy, 1951a, Sanger and Thompson, 1953b, Sanger and
Thompson, 1953a), but also the first determined protein crystal structure (Hodgkin,
1971) as well as first protein to be artificially produced (Zahn, 2000) and the first
recombinantly engineered gene (Ullrich et al., 1977).
Generally, biologically active peptides can have various modes of actions
(antagonistic/inhibitory, agonistic/stimulatory) origins (ribosomal, de nonol
non-ribosomal synthesis) and targets, such as the producing cells themselves or
(endocrine, exocrine). Originally, this classification (greek Kpivto/crino: to judge) was
conceived for the description of secreted, glandular compounds in bodily fluids denoted
as hormones (greek op(iovr|/ormoni: impulse, onset) (Kastin, 2006). Possibly due to this
origin of the definition, the terminology in the literature referring to biologically active
peptides in the kingdom Plantae as "plant peptides", "plant signalling peptides" or
"plant peptide hormones" is inhomogeneous. Some authors seem to avoid the term
"peptide hormones" intentionally, possibly to emphasize the fundamental differences
with animals and lack of classic endocrine hormone glands in plants.
1.1.1.1. Plant signalling peptides
Under the lead of Clearance A. Ryan at Washington State University, SYSTEMIN was
the first bioactive signalling peptide identified from plants (Schaller and Ryan, 1994).
The range of plant signalling peptides has been extended recently. Currently, the plant
peptides known to the author include SYSTEMIN (Schaller and Ryan, 1994),
PHYTOSULFOKINE (PSK) (Matsubayashi et al., 2002), CLV3/ESR-LIKE (CLE)
(Cock and McCormick, 2001a), RAPID ALKALINISATION FACTOR (RALF)
(Pearce et al., 2001b), S-LOCUS CYSTEIN RICH PEPTIDE (SCR/SP11) (Schopfer et
al., 1999a), ^/PEPl/PROPEP (Huffaker et al., 2006, Yamaguchi et al., 2006),
POLARIS (Farrokhi et al., 2008), ROTUNDOFOLIA4/DEVIL1 (RTF/DVL1) (Wen et
al., 2004), INFLORESCENCE DEFICIENT IN ABSCISSION FACTOR 1 (IDA1)
(Butenko et al., 2009), GIBBERELLIC ACID-STIMULATED ARABIDOPSIS 4
(GASA4) (Roxrud et al., 2007), EARLY NODULIN40 (ENOD40) (Imaizumi-Anraku
et al., 2000, Vleghels et al., 2003, Campalans et al., 2004, Wan et al., 2007) and
EPIDERMAL PATTERNING FACTOR (EPF) (Hara et al., 2007, Hunt and Gray,
1.1.1.1.1. CLE signalling peptides
CLAVATA3/EMBRYO-SURROUNDING REGION (CLV3/ESR)-like sequences are
grouped to the CLE protein family and postulated to release CLE signalling peptides
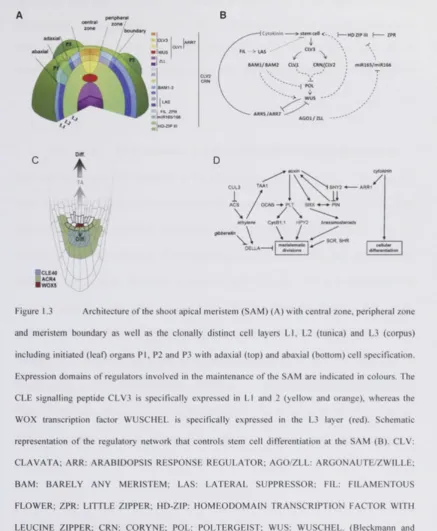
after posttranslational cleavage (Figure 1.1, Figure 1.2).
1 I t » 30 4 0 » S t 7 0 SO 9 0 1 0 0 1 1 0 1 2 0 1 2 4 I I I I I I I I I i I I i I
N O n C F L f l A X F L T P R V M I l t l X F S G P S T S S M Q O F H P M M S P I RSSHE R E F n S t C T B V P T r . s W l H H W ! n L T S £ n S A F S V L L I L L I F C N S - G F C I A G 6 L L N F T S S A S S f l K N l S l l SSNPELE OMfGVPSGflHPlHNR
t1GtHSSTISSSSSSSSSFFUSRFLRHLLVHGl.VSI.LVLCSlGSGEGTIHPriO-USQERV>CHER VVGROKPV D R B E U B T m i S m V P H G P O P I H H R R B G H S G R P P G f l n i n > K « N I I R E X r a j M I T l l S H I S S S S F L F S R F F F R V L W V G F V C F K V F S S t V S 6 6 6 T 6 a T I l l M I S £ E T U O { P g 8 V V V « n ( S V Y V I M M S L O n m ^ ^
n ^ K S S I I L L L L T I L L S V n O f l T I S R W S V T H V T L V O M E R R S t r r L S S n F ^ T S f l r L T S L M W r G W W r ^ H V V P V ^ P l H H
n M Y T T D O V O V Y V l V I V L C I F F S T L Q f l R S l . B I l H P L I H r a i D S R S I . L Q K l . R I H I S I M C O V HO ISGOKLSPIKPOPgNNGKSPPIIK NEHSMCVPISKIGl I H I f l F S I F F H S P H f l R R l E G G S M O S I l R L l f f LMVORIMIKRS RTDLEDKRVPGORLSPGGPHWIHH
l . . f « . l h k , k r . v f . G r * r . H M . r
h y d r o p h o b i c r e g i o n I variable region I CLE m o t i f I variable
Figure 1.1 Multiple sequence alignment of the seven CLE signalling peptide precursor protein sequences known from legumes (M. truncatula (Mi) and G. max (Gm)) at the start of the thesis project (Cock and McCormick, 2001b). The sequence identifier contains the corresponding EST TC number as well as their CLE sequence identifier (CLE number). Key elements of the protein precursor, which enable the bioinformatic identification of CLE signalling peptides, are illustrated below the protein sequence alignment. The sequence similarity within the CLE domain-containing protein family is restricted to the CLE motif of approximately 14 amino acids length, which is located at or near the C-terminus. Furthermore, a characteristic N-terminal secretion signal containing predominantly hydrophobic amino acids is contained in family members. In the central variable region and the dispensable C-terminal variable region sequence similarity cannot be observed.
CLE sequences Transcription Translation Precursor protein Translocated Precursor Cleavage I
Processed Precursor I Cleavage II Processed Precursor II posttranslational modification
CLE motif
CLE gene
CLE domain protein
PreproCLE peptide
ProCLE peptide
PreCLE peptide (premature)
CLE peptide (active)
S?
CLE
CLE
C L E | |
A
Receptor-binding
Figure 1.2 Diagram of CLE signalling peptide processing, maturation and nomenclature according to the definitions of the International Union of Pure and Applied Chemistry (IUPAC) and the International Union of Biochemistry and Molecular Biology (IUBMB) for " m o t i f ' , "domain", sequence" and "consensus". CLE domain proteins are translated to form the preproCLE signalling peptide precursor molecule. Via intermediate steps, cleavage and posttranslational activation, active CLE signalling peptides emerge from the precursor to trigger cellular responses upon receptor binding.
While this thesis project was underway, substantial progress has been made in the area
of CLE signalling peptides, particularly concerning the intricate biochemical
involvement of CLV3 in the regulation of shoot apical meristem (SAM) differentiation
in A. thaliana, and TRACHEARY ELEMENT DIFFERENTIATION FACTOR (TDIF)
in the control of vascular cell differentiation. Also, a CLAVATA-like receptor module
was identified that regulates stem cell differentiation at the root apical meristem (RAM).
For example, the CLV3 and TDIF peptides were detected in situ for the first time (Ito et
2006), direct interaction of the CLV3 peptide with the CLV1 receptor was demonstrated
(Ogawa et al., 2008), and details emerged on further CLAVATA receptor complex
components, such as CORYNE/SUPPRESSOR OF LLP1 2 (CRN/SOL2)
(Casamitjana-Martinez et al., 2003, Miwa et al., 2008, Bleckmann et al., 2009, Miiller et al., 2008).
Furthermore, CLAVATA-like receptors (ARABIDOPSIS CRINKLY-LIKE 4, ACR4)
and CLE signalling peptide (CLE40) involvement was demonstrated at the RAM (De
Smet et al., 2008, Hobe et al., 2003, Stahl et al., 2009). In addition, a receptor
(PHLOEM INTERCALATED WITH XYLEM, PXY/PUTATIVE TDIF RECEPTOR,
TDR) and signalling peptide (TDIF/CLE41) responsible for the differentiation of
procambial cells into xylem and phloem in Zinnia elegans and A. thaliana were
identified (Whitford et al., 2008, Hirakawa et al., 2008, Etchells and Turner, 2010,
Sieburth, 2007).
The introduction to Chapter Three will give a brief account of the evidence that was
available for the action of CLV/ESR-like sequences as signalling peptides prior to in
situ detection of CLE signalling peptides and demonstration of the direct CLV3-CLV1
1.1.1.1.1.1. The CLAVATA genes of Arabidopsis thaliana
The clavata-phenotype of A. thaliana was first analyzed by Leyser and Furner (1992).
Characteristic for this phenotype is the fasciation of the SAM (latin fasciculum: band,
ribbon), which includes altered phyllotaxis and broadening, flattening and eventually
bifurcation due to an increase of the SAM in size (Leyser and Furner, 1992, Leyser and
Day, 2003, Fletcher et al., 1999a). Several independent alleles resulting in
clavata-phenotypes had been identified. Thus there was strong evidence for action of
independent CLAVATA genes (clvl, civ2, and civ3) acting in the same signalling
pathway. Positional cloning revealed the identity of CLV1 as a LRR-RLK (Clark et al.,
1997), CLV2 as a receptor-like protein missing a kinase domain (Jeong et al., 1999) and
CLV3 as a 96 amino acid protein (Clark et al., 1995). Initially, no proteins were known
to share homology with CLV3. Overexpression of CLV3 in A. thaliana hampered the
initiation of organs at the SAM after emergence of the first leaves. These analyses
uncovered a signalling pathway required for limitation stem cell numbers at the SAM
and thus SAM maintenance.
1.1.1.1.1.2. Discovery and identification of CLE signalling peptides
In 2001, Cock and McCormick described the CLE protein family for the first time shortly after the publication of the A. thaliana genome sequence. The identification
relied on iterative protein database searches using position-specific iterative BLAST
(PSI-BLAST). The authors were able to identify 39 CLV3-related protein sequences
(Cock and McCormick, 2001a).
The first sequences that were described to later belong to the CLE family were initially
Z mays endosperm (Bonello et al., 2000b, Opsahl-Ferstad et al., 1997) and their mRNA constitutes the major proportion of the mRNA in the that region. The first description of the c/<2vata-phenotype in A. thaliana predates the identification of ESR genes (Leyser and Furner, 1992). Also, the best described member of the CLE family at the time of discovery had been CLV3, which is required for limiting the number of stem cells at the SAM and forms the paradigm of plant LRR-RLK signalling. Thus, the name of the CLE sequence family is derived from CLAVATA3 (CLV3) and EMBRYO-SURROUNDING REGION (ESR) amalgamated in the prefix CLE.
CLE/Synonym Number Identification First report
ESRal, ESRlg2 3 cDNA sequencing (OpsahlFerstad et al., 1997)
ESR3g2
CLAVATA3 1 Positional cloning (Clark et al., 1995, Fletcher et
al., 1999b, Brand et al., 2000)
CLE1-CLE39 39 PSI-BLAST (Cock and McCormick, 2001b)
CLE40 1 blastn (Sharma et al., 2003)
HgCLE 1 MEME/MAST (Olsen and Skriver, 2003b)
HgSYV46
5«CLE19 1 Differential display
reverse transcriptase
PCR
(Fiers et al., 2004b)
CLE41-CLE45 5 tblastn (Strabala et al., 2006)
FON4 1 Positional cloning (Chu et al., 2006b)
OyCLE42- 10 tblastn (Chu et al., 2006b)
OsCLE51
ZhCLE/TDIF 1 Mass spectrometry (Ito et al., 2006, Kondo et al.,
2006)
1.1.1.1.1.3. Primary sequence characteristics of the CLE family
The designation CLE testifies the discovery of the first identified biological sequences
carrying a characteristic C-terminal protein motif. This characteristically conserved
CLE motif spans 12-20 residues (Chapter Two). A hydrophobic signal peptide is
located at the N-terminus and required for the translocation to the apoplast in order for
CLV3 to function (Rojo et al., 2002). Between the N-terminal hydrophobic region and
the C-terminus CLE motif sequence conservation can generally not be observed
(Chapter Two). Thus, the interlaying region is regarded as variable. Precursor protein
sequences of CLE signalling peptides have predominantly, but not exclusively, basic
isoelectric points. The majority of precursor proteins are between approximately 50 and
120 amino acids long (Figure 1.1, Figure 1.2).
1.1.1.1.1.4. The receptor-binding of CLE signaling peptides
Ogawa and colleagues (2008) determined the specific binding of CLV3 by the CLV1
receptor with a dissociation constant of 17.5nM (Ogawa et al., 2008). Mutated CLV3
peptides corresponding to the genomic mutations in clv3-l and cl3-5 alleles as revealed
by positional cloning (Fletcher et al., 1999a) were unable to bind the CLV1 receptor.
1.1.1.1.1.5. Posttranslational processing: cleavage and covalent
modifications
The posttranslational cleavage of preproCLE signalling peptides determining the length
of the active peptides is unknown for most CLE signalling peptides. Considering the
available reports, there are controversies regarding CLV3, which has been reported in
glycosylated peptide carrying an arabinose residue (Kondo et al., 2006, Ohyama et al.,
2009). TDIF has been identified as 12 amino acid peptide (Ito et al., 2006). Two
peptidases have been postulated to be involved in the cleavage of the peptide precursor.
• i 2+
Mutagenesis and map-based cloning identified the A. thaliana SOLI Zn
-carboxypeptidase, as a suppressor of the CLE 19 overexpression phenotype
(Casamitjana-Martinez et al., 2003). Ectopic overexpression of the A. thaliana subtilase
yl/SBT5.4 produced developmental defects resembling a clavata phenotype (Liu et al.,
2009). However, biochemical evidence, such as in vitro cleavage of peptides is not
available yet.
The oligopeptide transporter from A. thaliana, OTP6 has been identified and
characterization in Xenopus laevis oocytes revealed a broad spectrum of peptide
transport, including CLE signalling peptides. The authors speculate about an
involvement of this peptide transporter in nodulation (Pike et al., 2009).
1.1.1.1.1.6. Physiological function of CLE members in plants
CLE domain proteins have been identified in monocotyledonous and dicotyledonous
species across the plant kingdom and are part of the essential developmental toolkit of
plants. CLE signalling peptides act in close collaboration with homeotic genes of the
WUSCHEL homeobox (WOX) gene family to limit stem cell differentiation in the
primary growth axis of plants (Floyd and Bowman, 2007, van der Graaff et al., 2009). The CLE family is plant specific with the exception of CLE domain proteins encoded
by plant-parasitic nematodes. Nematodes have presumably acquired a plant signal
through horizontal gene transfer in order to induce feeding site establishment in the
1.1.1.1.1.7. Embryo Surrounding Region genes
CLE members originating from embryo surrounding region genes were the first
described members of the CLE family. These genes are specifically expressed in the
embryo surrounding region of Z mays endosperm in vicinity of the developing embryo
(OpsahlFerstad et al., 1997) and their mRNA constitutes the major proportion of mRNA
of this region (Bonello et al., 2000a). Initially eight gene candidates (ESRlgl, ESRlg2,
ESR2gl, ESR2g2, ESR3g2, ESRlcl, ESR2cl, ESRal) were sequenced from isolated
mRNA, out of which three highly homologous proteins (ESR1 ESR2 ESR3) were
predicted (OpsahlFerstad et al., 1997). The biological function of these genes remains
unknown, but it is likely that their role is related to endosperm or embryo development.
1.1.1.1.1.8. The stem cell niches around CLAVATA3 and CLE40
The shoot apical meristem (SAM) and root apical meristem (RAM) are the two centres,
which control plant stem cell differentiation, growth and development in the primary axis of plants, giving rise to all aboveground and belowground tissues respectively
(Leyser and Day, 2003) (Figure 1.3A and Figure 1.3C). Longitudinally, the SAM is
organized into an outer layer (tunica), which in turn can be separate into the two
clonally distinct cell layers (L1 and L2), and an inner cell layer (corpus, L3). Radially,
the SAM consists of central zone (CZ), peripheral zone (PZ) and boundary (Leyser and
Day, 2003).
The A. thaliana SAM contains a pool of undifferentiated stem cells in the outermost cell
layers of the central zone. A signalling loop connects these stem cells of the central
zone with the underlying organizing center. Expression of the homeotic WOX family
transcription factor WUSCHEL (WUS) in the organizing center (Laux et al., 1996)
CLV3 (Clark et a l | 1995, Fletcher et al., 1999a). Binding of CLV3 to its receptor CLV1
(Clark et al., 1997) leads to repression of WUS transcription. Cells that exit this stem
cell niche become recruited into differentiation pathways, and form the plant organs,
such as leaves. The functional orthologs of the CLV-WUS pathway have been identified
in further model systems such as O. sativa FLORAL ORGAN NUMBER 1 (FON1),
FON2/FON4, FON1 SPARE 1 and FON1-LIKE CLE PROTEIN (FCP) (Suzaki et al.,
2004, Suzaki et al., 2006, Chu et al., 2006a, Suzaki et al., 2008, Suzaki et al., 2009) and
Z. mays THICK TASSEL DWARF 1 (TD1), CRINKLY-like 4 (CR4) and
FASCIATED EAR 2 (FEA2) (Cao et al., 2005, Bommert et al., 2005) . Further
peripheral zona / j /boundary m
B
JC L V ;
FIL ZPR
HNR166/166
CLV2 CRN
—> stem cell \
fe i
CIV3 .
f \
BAM1/BAM2 CLV.l CRN/.CLV2
ARRS/ARR7
AGOl/ZU
mfR165/miR266
[image:34.504.35.473.60.591.2]D
HISTONE ACETYL TRANSFERASE GCN5; PLT: PLETHORA; HPY: HIGH PLOIDY; CYCB1: CYCLIN B l ; BRX: BREVIS RADIX; PIN: PIN-LIKE FORMED; SHY: SHORT HYPOCOTYL; SCR: SCARECROW; SHR: SHORTROOT; ARR: ARABIDOPSIS RESPONSE REGULATOR; TA: transit-amplifying cells; Diff.: cell differentiation (Stahl and Simon, 2009). Figure 1.3A and Figure 1.3B are adapted from Bleckmann and Simon (2009); Figure 1.3C, and Figure 1.3D are adapted from Stahl and Simon (2009).
1.1.1.1.1.9. CLE signalling peptides and biotechnological applications
Plant growth factors and regulators in the widest sense, i.e. classic phytohormones and
plant signalling peptides resemble bona fide targets for molecular design of plant
architecture yielding improved, potentially ideal, crops (Wang and Li, 2008). There are
at least two specific examples of attempts to patent CLE signalling peptides and their
nucleotide or protein precursor sequences and the use of these molecules for biotechnological, commercial application for altering plant development (Liu et al.,
2001, Bruce et al., 2003). Potentially, peptides could be applied to fields, soils or seeds
among others. In return, this would have a beneficial outcome on various stages of plant
development, such as germination synchronization, inhibition of pre-harvest sprouting,
enlarged fruit size, increased seed number or changes to root architecture. However, the
ethics of patents covering biological sequences of natural origin evolved through
Chapter One
1.1.2. Receptor-like kinases
The detail of knowledge available of animal signal transduction is not comparable with
plant signal transduction. However, this is in stark contrast to the number of
receptor-like kinases (RLKs) and receptor-receptor-like cytoplasmic kinases (RLCKs, RLKs lacking
extracellular domains) encoded by plant genomes compared to animal genomes
(Becrafit, 1998, Becraft, 2002, Shiu and Bleecker, 2001a, Shiu and Bleecker, 2001c).
Generally, RLKs and RLCKs of plants belong to a large, monophyletic gene family of
more than 600 members, representing almost 2.5% of the protein coding sequences of
the entire Arabidopsis thaliana genome (Shiu and Bleecker, 2001a, Shiu and Bleecker,
2001c). There seem to be shared features in plant and animal RLK structures and signal
transduction pathways, but generally the extracellular domains of plant RLKs are
different from animal receptors (Torii, 2000, Cock, 2002, Tichtinsky et al., 2003).
Exceptions are leucine-rich repeat RLKs (LRR-RLKs), which resemble the TOLL-like
receptor (TLR) family of animals (Gay and Gangloff, 2007, Torii, 2004). Based on their
primary sequence similarities as well as secondary and tertiary structure characteristics,
plant RLKs are grouped into 15-25 subfamilies (Figure 1.4) (Becraft, 1998, Becraft,
2002). LRR-RLKs represent the largest receptor subfamily in the A. thaliana genome
with more than 200 members (Torii, 2004).
1.1.2.1. Modes of action of plant receptor-like kinase
Generally, RLKs contain a protein kinase that is modulated, usually by trans- or
autophosphorylation, in response to a stimulus, i.e. the perception of an extracellular
ligand such as a phytohormone or signalling peptide. RLKs are made up of an
extracellular receptor domain (ectodomain), transmembrane domain(s) and an
intracellular, cytoplasmic kinase domain (Becraft, 2002). Usually the amino-terminus
(N-terminus) is located extracellularly, while the carboxyl-terminus (C-terminus) is
located within the cell membrane (Tichtinsky et al., 2003). Different mechanisms exist
how RLKs transduce extracellular signals. The dogma is that RLKs appear as inactive
monomers in the membrane and dimerize in response to ligand binding, bringing each
other's cytoplasmic domains in proximity, and allowing activation by
autophosphorylation. The phosphorylation results in the activation of the intracellularly
located kinase domain, which transphosphorylates adaptor proteins that represent the
first component of the intracellular signalling cascade (Becraft, 2002, Becraft, 1998).
Other hypotheses predict that they occur as multimers in their inactive form, some show
intracellular kinase activity. The actual mode of action for an RLK must be determined
individually. Concerning their target phosphorylation sites, RLKs can be classified
either as receptor tyrosine kinase or as serine/threonine kinase receptors (Becraft, 2002,
Becraft, 1998). The evolutionary emergence of single specificity receptor tyrosine
kinases and the presence of such kinases in plants are disputed. There is evidence for
the occurrence of protein tyrosine phosphorylation in Archaeae (Kennelly and Potts,
1996, Smith et al., 1997). Also, findings from protist Choanoflagellata and the amoeba
Dictyostelium suggest that receptor tyrosine kinases might have arisen before the
divergence of plants, metazoans and fungi (King and Carroll, 2001). However, these
authors contradict themselves about the evolutionary presence of protein tyrosine
kinases in plants (King and Carroll, 2001, King et al., 2003)
1.1.2.2. Plant receptor-like kinase family
Based on their ectodomains receptor-like kinases are grouped into 15-25 subfamilies,
Signal/TM — Chitinase |
DUF26 | EGF repeat • B-lectin | C-lectin |
L-lectin | LRR • LysM • PAN | Proline-rich |
Thaumatin TNFR repeat •
U-box 9 Ser/Thr kinase |
Unknown <
Figure 1.4 Families of receptor-like cytoplasmic kinases (RLCK) and receptor-like kinases (RLK) in plants and schematic overview of their domain structure (Shiu and Bleecker, 2001a). The grey line represents the plasma membrane except for TAK, which is localized to the thylakoid membrane. The domains above the grey line are the putative extracellular domains with signal sequences. The area below the grey line represents cytoplasmic domains. The signal peptides are presumably absent in mature proteins and are displayed to demonstrate their presence in the RLKs. These representative RLKs are numbered as follows: 1, A5gl2000; 2, NAK; 3, TAK; 4, PERK; 5, RKF3; 6, CrRLKl 7, LRK10; 8, At5g56890; 9, Xa21; 10, CLAVATA1; 11, BRI1; 12, TMKL1; 13, Atlg53340; 14, TMK1; 15, LRRPK; 16, SERK; 17, Atlg52310; 18, At3g26700; 19, WAK1; 20, PR5K; 21, LecRKl; 22, RKF2; 23, SRK; 24, CRINKLY4 and 25, CHRK1. TM, transmembrane domain; DUF, domain of unknown function; EGF, epidermal growth factor; B-lectin, agglutinin; C-lectin, C- type lectin; L-lectin, legume lectin; LRR, leucine-rich repeat; LysM, lysin motif; PAN, plasminogen/apple/nematode protein domain; TNFR, tumor necrosis factor receptor. The figure is adapted from Shiu and Bleecker (2001a).
ft
1 2 3 4 S 6 7 B 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
I I I I
1.1.2.3. Plant leucine-rich repeat receptor-like kinases
The plant LRR-RLK family comprises the largest subfamily of transmembrane
receptors in plants with several hundred members. Only in few cases an assignment of
ligand-receptor pairs was possible and little knowledge is available of their signalling
pathways (Matsubayashi, 2003b, Torii, 2004). The LRR motif itself is a 20-29 amino
acid long sequence with the consensus "LXXLXLXX(N/C)XL" forming a
horseshoe-shaped three-dimensional structure (Kobe and Deisenhofer, 1994). The LRR motif
sequence is present in proteins with diverse functions but appears to have a primary
function in the formation of a versatile structural framework for protein-protein
interactions (Kobe and Deisenhofer, 1995, Kobe and Deisenhofer, 1994, Kobe and
Kajava, 2001).
One aspect of plant biology, in which LRR-RLKs play important roles are
plant-microbe interactions. Both, parasitic and symbiotic associations of plants with
microorganisms provide examples of LRR-RLK involvement. In this thesis, the
symbiosis between plants of the legume family and their symbiosis with nitrogen fixing
bacteria is the central focus. The interaction and its importance for agriculture is
1.2. Nitrogen and symbiosis
In most farming areas nitrogen is the primary growth limiting macronutrient of plants.
Modern agriculture disperses large quantities of nitrogen in form of synthetic
nitrogen-containing fertilizers into the soil to promote plant growth. This is an artificial and
anthropogenic remedy, with widespread ecological consequences (Drinkwater et al.,
1998). The nitrogen contained in synthetic fertilizers originates from molecular nitrogen
and hydrogen gas, which are the starting materials for industrial nitrogen fixation, the
Haber-Bosch process (Haber, 1918, Bosch, 1931, Bergius, 1931). Biologically, various
metabolic pathways can supply nitrogen to the avail of organisms. An important source
of nitrogen and factor in the biogeochemical nitrogen cycle is the natural ability of
symbiotic nitrogen fixing bacteria (diazotrophs) to form root nodule symbioses.
Phylogenetically, root nodule symbioses are restricted to the Rosid I clade of
angiosperms (Sprent, 2007). The most important symbioses include actinorhizal
symbioses of plants from the order Rosales, Fagales and Cucurbitales with bacteria
from the genus Frankia (Actinorhiza), and symbioses of leguminous plants (order
Fabales) and the single non-legume genus Parasponia (family Ulmaceae) with bacteria
from the genera Azorhizobium, Allorhizobium, Bradyrhizobium, Mesorhizobium,
Rhizobium and Sinorhizobium (collectively referred to as rhizobia). Both forms of root
nodule symbioses rely on the ability of the bacterial symbionts to perform an
energetically unfavourable, endergonic enzymatic reaction employing nitrogenase to
convert the inert, molecular, atmospheric nitrogen to bioavailable nitrogen (Sprent,
Thus, if the soil is deficient in nitrogen-containing compounds, these specific plants are
able to engage in a unique symbiotic association in order to overcome shortages in
nitrogen. This leads to the organogenesis of a specific symbiotic, organ-like structure in
the plant, the root nodule, which accommodates the symbiosome (Spaink, 1995).
During the establishment of this process, the bacteria differentiate into a symbiotic
form, called bacteroids. In return for obtained nitrogen, the plant provides the bacteria
in the symbiosome with carbon compounds from photosynthesis such as homocitrate
(Hakoyama et al., 2009). Furthermore, the plant maintains optimal conditions for
nitrogenase activity by reducing the oxygen partial pressure actively through
biosynthesis of leghaemoglobin (Garrocho-Villegas et al., 2007, Ott et al., 2009).
The symbiotic interaction has increasingly fascinated scientists, Kutschera et al. and
Ngai review interesting historical aspects (Kutschera and Niklas, 2009,
McFall-Ngai, 2008).
1.2.1. The model legume Medicago truncatula
The model system used throughout this study is the leguminous plant Medicago
truncatula also referred to as "barrel medik" due to the barrelled shape of its seedpods.
Maureira-Butler et al. (2008) provide a selection of seedpod images. M. truncatula is
an omni-mediterranean plant, grown as annual forage legume and a near relative of
alfalfa {Medicago sativa), which represents the world's economically most important
forage legume (Maureira-Butler et al., 2008). M. truncatula is self-fertile and its
genome, unlike the complex genome of many other legume species, is diploid, having
eight pairs of chromosomes. M truncatula has been selected as a model legume because
of its small genome size, short seed-to-seed generation time and abundant seed set (May
Sinorhizobium meliloti, has been fully sequenced (Galibert et al., 2000). S. meliloti
infects M. truncatula with high host specificity, making the S. meliloti-M. truncatula
system a paradigm for the study of root nodule symbioses. Although, it was recently
shown that the host plant is poorly matched with its symbiont strain S. meliloti 1021
commonly used in laboratories throughout the research community. Growth of M
truncatula in symbiosis with S. meliloti 1021 under nitrogen-free conditions leads to
sub-optimal nitrogen nutrition and nitrogen stressed plants. As a better matching
symbiont Sinorhizobium medicae WSM419 has been recommended, also having a fully
sequenced genome (Terpolilli et al., 2008, Moreau et al., 2008). M. truncatula Gaern.
cv. Jemalong A17 is the wild type representing the model legume plant forming
indeterminate nodules. Lotus japonicus on the other hand is a model for determinate
nodule formation. In indeterminate nodules the nodule meristem persists, unlike the
mechanism of determinate nodule formation, where the tissue matures synchronously
and no meristematic tissue remains (Spaink, 1995).
1.2.2. The root nodule symbiosis
One motivation to study this symbiosis is to extend it to non-legume plants, which are
unable to form symbioses with diazotrophic bacteria. If the genetic framework
necessary for this symbiosis could be introduced to agronomically important
non-legumes such as Oryza sativa (rice), Zea mays (corn) or Triticum aestivum (wheat) the
inert nitrogen of the earth atmosphere could be converted to bioavailable nitrogen, which would reduce the need for costly and polluting nitrogen fertilizers (Hurtley and
Marx, 2004). However, so far, the signal transduction pathway that controls root
nodulation in legumes is only partly known. This thesis aims at deciphering components
1.2.3. Essential steps for the formation of nodules
The essential steps for the development of functional root nodules and concomitant
morphological changes of the plant vary substantially among various legume species
(Sprent, 1989). The following part of the introduction will give a brief overview over
the signal exchange and involved components in the case the indeterminate nodule
organogenesis of M. truncatula, as host legume and S. meliloti, as bacterial symbiont.
Essential steps of the rhizobia-legume symbiosis involve pre-infection (multiplication
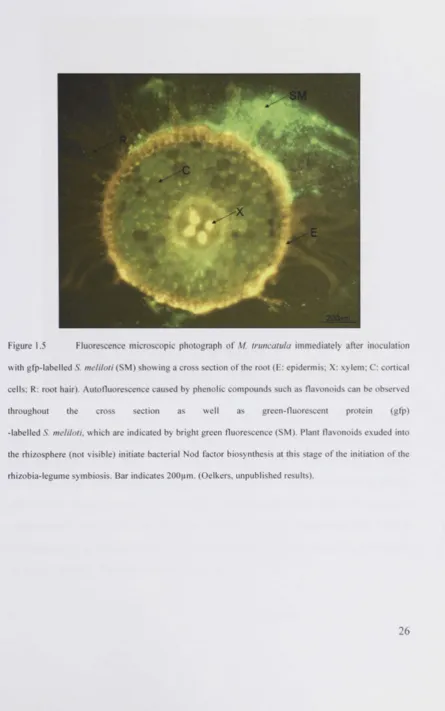
of bacteria, colonisation of the root, bacterial attachment to the root) (Figure 1.5),
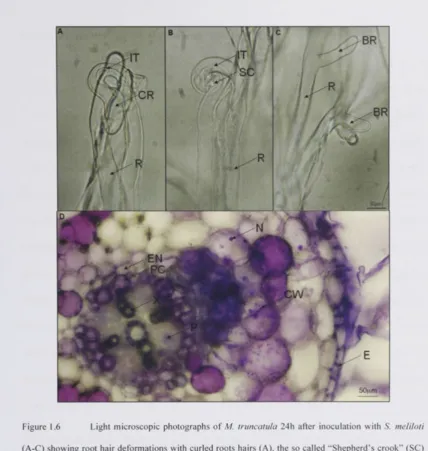
infection (root hair curling, branching, infection) (Figure 1.6A-C), nodule formation
(cortical cell division and differentiation) (Figure 1.6D) and nodule functioning
(nitrogenase facilitated nitrogen fixation, leghaemoglobin biosynthesis, nodule
persistence) (Finan et al., 1985, Schultze and Kondorosi, 1998, Spaink, 1995).
In the past years research in the field was focused on decoding the complex molecular
conversation between bacteria and host plants required for the establishment of the
symbiosis. More recently, essential components of the pathway of root nodule initiation
were identified, uncoupling the essential steps between symbiotic bacteria and host
plant (Finan et al., 1985). The identification of components relied on the analysis of
mutants, which were derived from chemical (ethylmethane sulfonate) or radiation
(y-ray, fast neutron bombardment) mutagenesis (Stougaard, 2001). The mutants presented
altered nodulation phenotypes, e.g. non-nodulating (deficient in nodulation: nod") or excessively nodulating, ("supernodulating", "hypernodulating": nod++). The screening
of mutations using predominantly positional cloning resulted in the discovery of novel
Due to the amount of literature available on the subject, some details of nodule
formation have been omitted. The reader is referred to excellent recent reviews
(Oldroyd and Downie, 2008, Crespi and Frugier, 2008, Jones et al., 2007, Den Herder
and Parniske, 2009).
1.2.3.1. Plant flavonoids induce bacterial Nod factor biosynthesis
Legume plants exude flavonoids into the rhizosphere (Figure 1.5). These phenolic
compounds act as potent inducers of bacterial gene expression (Peters and Long, 1988,
Redmond et al., 1986). The Nod box mediates the bacterial perception of flavonoids.
Nod D which is a lysR-family transcriptional activator, existing in all rhizobia species
in three isoforms, interacts with flavonoids directly and activates in response to
flavonoid binding the expression of Nod, Nol and Noe genes in bacteria. Nod genes
encode enzymes that synthesize so-called nodulation factors (Nod factors), which are
essential for the establishment of the symbiosis. Most Nod genes are located on the
essentially required rhizobial sym-plasmid, only few are located in the bacterial genome
Figure 1.6 Light microscopic photographs of M. truncatula 24h after inoculation with S. meliloti
1.2.3.2. Structure and biosynthesis of Nod factors
The rhizobial response to plant flavonoid secretion and recognition is Nod factor
biosynthesis. Nod factors are oligosaccharide-based molecules with an attached lipid
moiety, lipochitin oligosaccharides. They represent the most important host specificity
determining signal molecule in the rhizobia-legume symbiosis (Hirsch, 1992, Spaink,
1995, Spaink, 2000, Spaink, 2004). Quite a number of Nod genes are involved in the
formation and processing of Nod factors. The backbone is synthesized by Nod A (chitin
transferase), Nod B (chitin synthase) and Nod C (chitin deacetylase). Further
modifications on the resulting three to five residue long (3-1.4-linked
N-acetyl-D-glucosamine backbone are performed in a species-specific manner and catalysed by
products of other Nod genes. Examples of such modifying enzymes are Nod L encoding
an acetyl transferase or Nod F and Nod E encoding ketoacetyl synthases. Numerous
strain-specific chemical decorations of Nod factors are possible, such as the attachment
of sulphate, methyl, acetyl and carbamoyl groups, or glycerol, fucose and arabinose
residues (Spaink, 2000).
1.2.3.3. Nod factors determine host specificity and induce nodule formation
There are several facts that indicated the existence of high-affinity receptors in the plant
for the recognition of Nod factors. Their action depends on the defined chemical
decoration of the P-1.4-N-acetyl-D-glucosamine backbone. They act at low, picomolar
concentrations, can mimic symptoms of nodule formation without the presence of
bacteria, and are unable enter plant cells due to their amphiphilic character (Spaink,
1995). Recent studies based on nodulation deficient mutants identified novel plant
and Frugier, 2008). Radutoiu and colleagues conclusively demonstrated the host
specific effect of Nod factors by introducing receptors required for Nod factor
perception of M. truncatula in L. japonicus, thus extending the symbiotic range of L.
japonicus from Mesorhizobium loti to Sinorhizobium meliloti (Radutoiu et al., 2007).
The Nod factor-dependent pathway of root nodule formation is the prevalent signalling
pathway for nodule initiation and establishment of the rhizobia-legume symbiosis.
Alternative pathways have been developed by photosynthetic Bradyrhizobia, which can
establish the symbiosis in a Nod factor independent pathway (Giraud et al., 2007).
There are further modes of interaction, which are not as well understood as the
interaction between M. truncatula and S. melioti (Sprent, 2007, Elliott et al., 2007,
Phenotypical Mutant _ k ' L . „ c
, f . Gene product Plant species Reference
detect allele
t \ /! d i v 7 • (Madsen et al., 2003)
nfrl/nfrS LysM-RLK L.japomcus ( R a d u t o i u e t al., 2003) Nod factor nfp LysM-RLK M. truncatula (Amor et al., 2003) perception symlO LysM-RLK P. sativum (Madsen et al., 2003)
1 LysM-RLK P. sativum ,T. . , -,aa~,n
V™2 (LYK3, LYK4) M truncatula (LlmpenS * 2 ° ° 3 )
dmi2 LRR-RLK M. truncatula (Endre et al., 2002)
symrk LRR-RLK L. japonicus (Capoen et al., 2005)
nork LRR-RLK M. sativa (Endre et al., 2002) Calcium flux syml9 LRR-RLK P. sativum (Stracke et al., 2002) root hair dmil ion channel M. truncatula (Ane et al., 2004) deformation
pollux ion channel (Imaizumi-Anraku et al.,
pollux ion channel L. japonicus
2005)
ion channel L. japonicus (Imaizumi-Anraku et al., castor ion channel L. japonicus
2005)
dmi3 CCaMK M. truncatula (Levy et al., 2004)
nsp I GRAS-family TF M. truncatula (Smit et al., 2005) Calcium spiking nsp2 GRAS-family TF M. truncatula (Kalo et al., 2005)
nin Putative TF L. japonicus (Schauser et al., 1999)
sym35 Putative TF P. sativum (Borisov et al., 2003)
hcl Unknown M. truncatula (Wais et al., 2000b)
I in cerberus
(Yano et al., 2009,
I in
cerberus Ubox/WD40 M. truncatula Kuppusamy et al., 2004,
Root hair curling
I in cerberus
Kiss et al., 2009) Infection thread crinkle
(Tansengco et al., 2003) formation Ljsym79 Unknown L. japonicus (Tansengco et al., 2003) alb Unknown L. japonicus (Imaizumi-Anraku et al.,
alb Unknown L. japonicus
2000)
harl CLVl-like
L. japonicus (Krusell et al., 2002) Ljsym78 LRR-RLK L. japonicus (Nishimura et al., 2002b)
sym29 CLVl-like
LRR-RLK P. sativum (Krusell et al., 2002) Nodule number nark/nts
CLVl-like
LRR-RLK G. max (Searle et al., 2003) control
sunn CLVl-like
LRR-RLK M. truncatula (Schnabel et al., 2005a)
sickle EIN2 M. truncatula (Penmetsa et al., 2003b)
klavier Unknown L. japonicus (Oka-Kira et al., 2005)
astray
Ljsym77 bZIP TF L. japonicus (Nishimura et al., 2002c)
Table 1.2 A selection of plant signalling components required for the initiation of nodule organogenesis and references for their identification in Pisum sativum, Glycine max, Medicago truncatula, Lotus japonicus and Medicago sativa (Crespi and Frugier, 2008, Stacey et al., 2006, Den Herder and Parniske, 2009). nfr: Nod factor receptor; nfp: Nod factor perception; sym: symbiosis; nsp:
nodule signalling pathways; dmi: doesn't make infection; symrk: symbiosis receptor kinase; nork:
[image:50.504.12.471.3.737.2]1.2.3.4. The development of a nodule
The external application of Nod factors onto plant roots leads to membrane
depolarisation. Calcium influx, calcium efflux and a potassium efflux seem to be
responsible for this membrane depolarisation, which is the first indicator of symbiotic
engagement. Within minutes after Nod factor exposure a calcium spiking in root hair
cells can be observed (Miwa et al., 2006, Kosuta et al., 2008, Hazledine et al., 2009).
Calcium spiking plays an important role in gene regulation and leads to the activation of
the nodule initiation signalling cascade (Wais et al., 2000a, Oldroyd and Downie, 2006).
After few hours (1-3 Oh) root hairs deform, which can be observed as root hair curling,
branching, and swelling. Bacteria are trapped in curled root hairs ("Shepherd's crook"),
which are then invaded by bacteria forming so-called infection threads, a tubular
structure within the root hair (Figure 1.6A-C). Synchronously with the penetration of
bacteria into the root through root hairs and within 18-30h after exposure to Nod
factors, asymmetric cell division in inner cortical cell layers can be observed as the first
sign of nodule organogenesis. The nodule primordium is only found at certain positions
in the inner root cortex, at sites opposing protoxylem poles (Figure 1.6D, for a
comparison of root architecture in a cross section see Figure 1.5). On the genetic level
expression of early nodulin (ENOD) genes occurs. Finally, through the formation of
cytoplasmatic bridges and cell wall penetration in infection threads bacteroids invade
the multiplying cortical cells (Sprent, 1989, Hirsch, 1992, Finan et al., 1985, Crespi and
1.2.4. Phytohormones in nodule formation
The formation of nodules is tightly integrated into plant development and there are
strong similarities between symbiotic root nodule development and the development of
innately induced organs, such as lateral roots (Hirsch and Fang, 1994, Hirsch et al.,
1997). In fact, the direct involvement of all the classic phytohormones in the formation
or control of root nodules has been demonstrated. In detail, this includes ethylene
(negative regulator) (Oldroyd et al., 2001, Penmetsa and Cook, 1997, Penmetsa et al.,
2008), auxin (positive regulator) (Mathesius et al., 1998, van Noorden et al., 2006,
Hirsch et al., 1989, Huo et al., 2006), flavonoids (positive regulator) (Wasson et al.,
2006, Subramanian et al., 2009), abscisic acid (negative regulator) (Ding et al., 2008,
Liang et al., 2006), cytokinin (positive regulator) (Gonzalez-Rizzo et al., 2006,
Mathesius et al., 2000, Frugier et al., 2008, Murray et al., 2007, Tirichine et al., 2007),
jasmonic acid (positive regulator) (Sun et al., 2006, Kinkema and Gresshoff, 2008),
brassinosteroids, gibberellins (negative regulator) (Ferguson et al., 2005, Maekawa et
al., 2009), polyamines (Terakado et al., 2006, Efrose et al., 2008) and salicylic acid
(negative regulator) (Peleg-Grossman et al., 2009). For a review see Ferguson and
Mathesius (2003). It has been shown that there is extensive crosstalk among
phytohormones for the coordination of development, i.e. in the differentiation of
vascular tissues (Dettmer et al., 2009), shoot branching (Ferguson and Beveridge, 2009)
and nodule development (Gonzalez-Rizzo et al., 2006, Sun et al., 2006, Ferguson and
Beveridge, 2009, Ferguson and Mathesius, 2003). Thus, the detailed roles of individual
phytohormones are complex, as they rarely act isolated, much rather in finely tuned and
orchestrated manner through interlocked signalling networks (Dettmer et al., 2009).
paragraph, long-distance shoot-root interactions are important for the signal integration
of plant growth regulators in the regulation of multicellular development, particularly
with regard to the control of nodule number (Beveridge et al., 2007). Also, one has to
differentiate between local as well as systemic effects of plant hormones in their
involvement in nodulation (van Noorden et al., 2006). Due to this tight integration into
essential plant developmental processes, it has been more difficult to genetically
uncouple nodulation specific from native plant developmental actions of
phytohormones than it has been to genetically uncouple bacterial from plant effectors
and responses (as discussed for flavonoids and Nod factors earlier). However, the
elusive roles and recruitment of phytohormones actions for the development of
symbiotic root nodules begin to emerge.
Besides phytohormones, microRNAs (miRNAs) have also been shown to be regulators
specifically involved in nodule formation (Combier et al., 2006, Boualem et al., 2008,
H ™
iv!|f .c FTL • I f * ' vi? iL J H '
I I I : I a :
i
mI S U N N 4 S 5 7 * I P 1 1 8
71 TT~
l i F M l
| p s
I h a r J - 4
_p106
| i fI S
Arabidopsis thaliana 15222877 CLVHike Brassica napus 31540632 CLV1-like Glycine max 7329122