SIV responses during early SIV infection.
N
atural killer (NK) cell reactivity against virus-infected cells is influenced by the balance of opposing signals mediated through the diverse family of activating and inhibitory killer-cell immunoglobulin-like receptors (KIRs) that recognize major his-tocompatibility complex (MHC) class I molecules (1). Therefore, KIRs likely modulate NK cell responses against human immuno-deficiency virus type 1 (HIV-1)-infected cells. Epidemiologic studies have shown that the presence or absence of particular KIR genes and the extensive allelic variability of KIRs affect HIV-1 disease outcome (2–10). The effects of KIR-expressing NK cells during acute HIV-1 infection, however, remain unclear.We have previously shown that genetic variation of KIRs is another aspect of KIR diversity affecting an AIDS virus infection. In that report, we demonstrated that copy number variation (CNV) of the activatingKIR3DHgene family was associated with lower simian immunodeficiency virus (SIV) replication during acute SIV infection inMamu-A*01⫺Indian-origin rhesus ma-caques that express restrictiveTRIM5alleles (11). In the present studies, we demonstrate an association between CNV of the other known activating KIR gene family in Indian-origin rhesus ma-caques,KIR2DL4(12–16), and the loss of CD4⫹T cells in acutely SIVmac251-infectedMamu-A*01⫺rhesus macaques, with higher
KIR2DL4copy numbers being associated with a less severe CD4⫹
T-cell depletion. Although KIR2DL4 contains a long cytoplasmic tail that is characteristic for inhibitory KIRs (17–19), binding through KIR2DL4 stimulates NK cells to produce cytokines and chemokines (20–24). Thus, KIR2DL4 is a predominantly activat-ing KIR in humans (15, 23, 24). In the present study, higher
KIR2DL4 copy numbers were also associated with increased
gamma interferon (IFN-␥) production by stimulated cytokine-producing NK cell subsets, thereby indicating thatKIR2DL4CNV may affect the production of a crucial cytokine in the antiviral immune response following SIV infection.
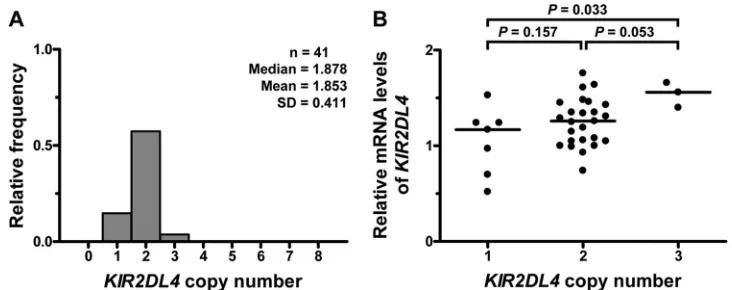
KIR2DL4CNV was evaluated in 41Mamu-A*01⫺Indian-origin
rhesus macaques by quantitative real-time PCR (qPCR) with a
KIR2DL4-specificTaqManprimer-probe set, 5=-GTTTCCTCATT
AGCCCTGTGACTGCA-3=, 5=-TTGCTGGGTGCCGACCACTC AGT-3=, and 5=-FAM (6-carboxyfluorescein)-TGCAGGGACCT ACAGATGTCGAGTTTTTCATCCGCACTCC-BHQ (black hole quencher)-3=, using genomic DNA samples.STAT6served as a
reference control gene (25). Absolute copy numbers were quanti-fied by generating standard curves using six serial log dilutions (108to 103copies) of plasmids containing the specific sequence to be amplified in the individual qPCRs and determiningKIR2DL4
copy numbers by normalizing againstSTAT6copies. Using this assay,KIR2DL4copy numbers varied between 1 and 3 copies per cell (Fig. 1A).KIR2DL4copy numbers were not associated with copy numbers ofKIR3DHthat were assessed in the same cohort (data not shown) using a previously published assay (11), indicat-ing that the two quantitative assays specifically measure copy numbers ofKIR2DL4orKIR3DH, respectively. This finding was further confirmed by sequencing of the qPCR amplicons (data not shown). Then, the effect ofKIR2DL4copy numbers onKIR2DL4
mRNA expression was determined by sorting peripheral blood CD14⫺ CD16⫹NK cells from 35 uninfected rhesus macaques using immunomagnetic beads. In these cells, relativeKIR2DL4
mRNA expression was measured using the KIR2DL4-specific qPCR assay described above using total RNA and analyzed using the⌬⌬CTmethod (26,27). RelativeKIR2DL4mRNA levels were
significantly higher in animals with threeKIR2DL4copies than in animals with oneKIR2DL4copy (Kruskal-Wallis test,P⫽0.033) (Fig. 1B), thereby indicating that higherKIR2DL4copy numbers result in increasedKIR2DL4transcript levels.
To assess the impact ofKIR2DL4CNV on SIV pathogenesis in rhesus macaques during primary SIV infection, we evaluated plasma SIV RNA levels using an ultrasensitive branched-DNA amplification assay (Bayer Diagnostics, Berkeley, CA) and the loss of total peripheral blood CD4⫹T cells and central memory (CM) CD4⫹T cells during primary SIV infection by flow cytometry in 41, 22, or 15Mamu-A*01⫺Indian-origin rhesus macaques, re-spectively. While there was no correlation betweenKIR2DL4CNV
Received19 October 2012Accepted19 February 2013
Published ahead of print28 February 2013
Address correspondence to Jörn E. Schmitz, [email protected].
† Deceased.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.02949-12
on November 7, 2019 by guest
and viral load at either peak, measured on day 14 post-SIV chal-lenge (Fig. 2A), or set point of the SIV infection, measured on day 70 post-SIV challenge (data not shown), we observed a signifi-cantly larger CD4⫹T-cell depletion in animals with lowKIR2DL4
copy numbers in a relatively small cohort of 22 Mamu-A*01⫺
rhesus macaques on day 14 following SIV infection (Mann-Whit-ney U test,P⫽0.03) (Fig. 2B). To assess whether expression of restrictiveTRIM5alleles contributed to this effect, we grouped the
Mamu-A*01⫺rhesus monkeys into two cohorts: one of animals
expressing only the restrictiveTRIM5alleles 1 to 5 and one of animals expressing at least one of the permissiveTRIM5alleles 6 to 11. There was no difference between loss of CD4⫹T cells between these two groups (Mann-Whitney U test,P⫽0.875) (data not shown). In contrast, when we further subgrouped these animals according to the number ofKIR2DL4copies that they have, we observed a strong trend of less CD4⫹T-cell depletion in animals with twoKIR2DL4copies than in animals with oneKIR2DL4copy in animals expressing TRIM5 alleles 1 to 5 (Mann-Whitney U test,
P⫽0.06) (data not shown), indicating that the expression of particularTRIM5alleles is not underlying the association between
KIR2DL4CNV and CD4⫹T-cell depletion. Unfortunately, there
were not enough animals with oneKIR2DL4copy (n⫽1) in the cohort expressingTRIM5alleles 6 to 11 to allow for statistical analysis (data not shown). Furthermore, a trend toward an asso-ciation between higherKIR2DL4copy numbers and smaller de-cline of CD4⫹T cells was observed on days 21 and 28 post-SIV infection (data not shown). In contrast, the loss of total peripheral CD4⫹T cells on day 70 following SIV infection (data not shown) and the decline of total circulating CM CD4⫹T cells on day 28 following SIV infection were not associated withKIR2DL4CNV (Fig. 2C). These findings may suggest thatKIR2DL4CNV influ-ences CD4⫹T-cell decline during acute SIV infection.
Since the allelic diversity of KIR genes has been shown to affect SIV replication (28,29), we assessed whether particularKIR2DL4
alleles might influence the effect ofKIR2DL4CNV on SIV patho-genesis. In Indian-origin rhesus macaques, two distinctKIR2DL4
forms,KIR2DL4.1andKIR2DL4.2, differ in their intracellular sig-naling motifs (16). These structural differences might result in altered NK cell reactivity mediated through theseKIR2DL4 allo-types, thereby potentially affecting the association between
FIG 1Distribution ofKIR2DL4copy numbers andKIR2DL4mRNA expression levels in Indian-origin rhesus macaques. (A) Copy numbers ofKIR2DL4genes were estimated in 41Mamu-A*01⫺rhesus macaques by quantitative real-time PCR (qPCR) using genomic DNA samples. The median, mean, and standard deviation (SD) ofKIR2DL4copy numbers are shown. (B) ForKIR2DL4mRNA levels, CD14⫺CD16⫹NK cells from uninfected rhesus macaques were isolated from peripheral blood mononuclear cells of 35 uninfected rhesus macaques by immunomagnetic bead separation. Total RNA from these cells was extracted, and KIR2DL4mRNA levels were determined using qPCR. Relative RNA expression was quantified using the 2⫺⌬⌬CTmethod (26,27). Rhesus macaques were grouped
into animals with 1, 2, or 3 copies ofKIR2DL4. Horizontal bars indicate medians. Analysis was performed using the Kruskal-Wallis test with Dunn’s multiple comparison test.
FIG 2KIR2DL4copy numbers and the loss of CD4⫹T cells on day 14 following SIV infection are associated inMamu-A*01⫺Indian-origin rhesus macaques. KIR2DL4copy numbers were determined in 15 to 41 SIVmac251-infectedMamu-A*01⫺rhesus macaques using qPCR, and the animals were subdivided into groups according toKIR2DL4copy number. In these groups, the association betweenKIR2DL4copy numbers and the clinical outcome following SIV infection was evaluated by measuring plasma SIV RNA levels (A) and the loss of total circulating CD4⫹T cells (B) and central memory (CM) CD4⫹T cells (C) on day 14 (peak) following SIVmac251 infection. Comparisons were analyzed using the Mann-Whitney U test for two groups and the Kruskal-Wallis test for three groups. Horizontal bars indicate medians.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:2.585.108.474.66.211.2] [image:2.585.59.524.547.666.2]KIR2DL4 CNV and CD4⫹ T-cell depletion. Expression of
KIR2DL4.1 and KIR2DL4.2 alleles was assessed in 41
Mamu-A*01⫺rhesus macaques by PCR. The presence ofKIR2DL4.1 al-leles was defined using 5=-TGTGATTAGGTACTCGGTGGCC-3=
(primer F1) and 5=-AGATTCCAYCTGCTGGTACATTG-3=
(primer R1). Due to high sequence homology betweenKIR2DL4
andKIR3DHalleles in the amplified region, bothKIR2DL4.1and
KIR3DHalleles were amplified using these primers. PCR products
were then ligated into the pGEM-T Easy backbone (Promega, Madison, WI). Plasmids containingKIR2DL4.1inserts were dis-tinguished fromKIR3DH-containing plasmids based on a splice site mutation in intron 8 (16) by PCR using 5=-CCCTCCTCTCC CCAGC-3= and R1 above, resulting in a KIR2DL4.1-specific 362-bp fragment. The presence ofKIR2DL4.2alleles can be dis-tinguished fromKIR2DL4.1allotypes by three deletions in nucle-otide positions 1052, 1106, and 1107 (16) and was assessed by PCR using F1 above and 5=-CAGCTGCTGGCACTGGA-3=, resulting in a 965-bpKIR2DL4.2-specific fragment. We observed that 27% of the animals expressed onlyKIR2DL4.1alleles, 39% expressed
onlyKIR2DL4.2alleles, and 34% expressed bothKIR2DL4.1and
KIR2DL4.2alleles (Fig. 3A). Then, we divided our cohort into
animals having one, two, or threeKIR2DL4gene copies. While
KIR2DL4.1and KIR2DL4.2 were fairly equally distributed
be-tween animals with two and those with threeKIR2DL4copies, 88% of animals with one KIR2DL4 copy expressed only
KIR2DL4.2alleles (Fig. 3B). We then assessed whether this
un-equal distribution ofKIR2DL4alleles contributed to the associa-tion betweenKIR2DL4CNV and CD4⫹T-cell depletion during acute SIV infection. In fact, animals that expressed only
KIR2DL4.1alleles lost significantly less CD4⫹T cells than did
animals that expressed onlyKIR2DL4.2alleles or bothKIR2DL4.1
andKIR2DL4.2alleles (Kruskal-Wallis test with Dunn’s multiple
comparison,P⫽0.049 andP⫽0.02, respectively) (Fig. 3C), while the loss of circulating CM CD4⫹T cells was not associated with the presence of particularKIR2DL4alleles (Fig. 3D). These find-ings suggest that lowKIR2DL4copy numbers are associated with a higher frequency ofKIR2DL4.2alleles and that the presence of
KIR2DL4.2alleles is associated with a more severe CD4⫹T-cell
decline.
We then evaluated whetherKIR2DL4CNV influences NK cell functionality. Since previous reports showed that KIR2DL4 stim-ulates potent IFN-␥production and only weak cytotoxicity in NK cells (15,22,23), we assessed the effect ofKIR2DL4CNV on IFN-␥ production inin vitro-stimulated NK cells from uninfected and SIVmac251-infected rhesus macaques on day 28 postinfection us-ing flow cytometry. Rhesus NK cells were defined as CD3⫺ CD8␣⫹NKG2A⫹(30,31), and IFN-␥secretion was assessed fol-lowing a 6-hour stimulation with K562 cells in three NK cell sub-sets defined by their expression of CD16 and CD56: CD16⫹, CD56⫹, and double-negative (DN) NK cells. Since these NK cell subpopulations differ in their main effector functions, with CD16⫹NK cells mediating strong cytotoxicity, CD56⫹NK cells producing mainly cytokines, and DN NK cells mediating cytotox-icity and producing large amounts of cytokines (31), an effect of
KIR2DL4copy numbers on NK cell function might occur only in
particular NK cell subsets. In uninfected rhesus macaques, there was no association betweenKIR2DL4CNV and IFN-␥production following stimulationin vitroin any of the three NK cell subsets (Fig. 4A). On day 28 post-SIV infection,KIR2DL4CNV was not associated with IFN-␥production in CD16⫹NK cells (Fig. 4B). In contrast, higherKIR2DL4copy numbers were significantly asso-ciated with increased IFN-␥production in DN NK cells
(Mann-FIG 3Distribution ofKIR2DL4alleles in Indian-origin rhesus macaques and their association with CD4⫹T-cell depletion. TwoKIR2DL4allotypes,KIR2DL4.1 andKIR2DL4.2, were distinguished based on a complex mutation in exon 9 of theKIR2DL4genes (16) in 41Mamu-A*01⫺rhesus macaques by PCR. The frequency of animals expressing onlyKIR2DL4.1alleles (black bars), onlyKIR2DL4.2alleles (gray bars), or bothKIR2DL4.1andKIR2DL4.2alleles (white bars) was evaluated in the entire cohort (A) and in cohorts of rhesus macaques that were grouped into animals with 1, 2, or 3KIR2DL4copies (B). Losses of total circulating CD4⫹T cells (C) and central memory (CM) CD4⫹T cells (D) on day 14 postinfection were compared betweenMamu-A*01⫺rhesus macaques expressing onlyKIR2DL4.1alleles (black circles), onlyKIR2DL4.2alleles (gray circles), or bothKIR2DL4.1andKIR2DL4.2alleles (white circles). Horizontal bars indicate medians. Data were analyzed using the Kruskal-Wallis test with Dunn’s multiple comparison.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.138.449.64.288.2]Whitney U test,P⫽0.01) (Fig. 4B), in a small cohort of 11 rhesus monkeys with only three of these animals having one copy of
KIR2DL4. Unfortunately, there were insufficient CD56⫹NK cells
to allow for a similar analysis. Furthermore, on day 28 post-SIV infection, all animals with oneKIR2DL4 copy expressed only
KIR2DL4.2alleles, while all rhesus macaques with twoKIR2DL4
copies expressed at least oneKIR2DL4.1allele (data not shown), thereby suggesting that the presence of particularKIR2DL4alleles might contribute to the effect ofKIR2DL4CNV on IFN-␥release from NK cells during acute SIV infection. These findings suggest that highKIR2DL4copy numbers are associated with increased IFN-␥production in stimulated cytokine-producing NK cell sub-sets during primary SIV infection.
In the present study, we show that higherKIR2DL4copy num-bers were associated with a less severe CD4⫹T-cell depletion and an increased IFN-␥production in stimulated NK cell subsets in
Mamu-A*01⫺ rhesus macaques during primary SIV infection.
This might demonstrate that genetic variability of the activating
KIR2DL4influences CD4⫹T-cell levels and the function of
par-ticular cytokine-producing NK cell subsets, thereby further em-phasizing the importance of KIR expression in modulating NK cell responses during the earliest phase of SIV infection.
Very few studies have evaluated KIR CNV, and to date, no study has quantifiedKIR2DL4CNV.KIR2DL4duplications have, however, been implied in transcriptional analyses in rhesus ma-caques and humans, with up to three different KIR2DL4 se-quences reported in a single subject (16,32,33). In line with these reports, 1 to 3KIR2DL4copies per cell were measured in the present study. This low copy number variation is also consistent with a study quantifying copy numbers of another KIR with 2 D-domains,KIR2DS2, ranging from 0 to 2 copies per cell (34).
Despite a low range of CNV, higherKIR2DL4copy numbers were associated with a smaller CD4⫹T-cell depletion. Our data
further suggested that the exclusive presence ofKIR2DL4.1alleles might have contributed to this better preservation of CD4⫹T cells, while animals that expressed at least oneKIR2DL4.2allele lost significantly more CD4⫹T cells during acute SIV infection. Consistent with this finding, a previous study documented an as-sociation between the presence of particularKIR2DL4alleles and plasma viral load levels in SIV-infected rhesus macaques (29). Furthermore, it was demonstrated that distinct humanKIR2DL4
alleles affected KIR2DL4 surface expression and function differ-ently (35). RhesusKIR2DL4.2alleles have a deletion in bp 1052 of exon 9 that results in the loss of a serine/threonine kinase site in the translated protein (36).KIR2DL4.2 alleles might therefore transmit weaker intracellular signals than doKIR2DL4.1alleles, as previously described for humanKIR2DL4alleles (35,37), poten-tially resulting in less efficient KIR2DL4.2-mediated NK cell re-sponses against target cells.
In addition to having an association with CD4⫹T-cell num-bers,KIR2DL4CNVs were also associated with NK cell function-ality in the current studies, with highKIR2DL4copy numbers being associated with increased IFN-␥production in DN NK cells, a major cytokine-producing NK cell subset (31). This suggests that higherKIR2DL4expression may have resulted in increased activation of particular NK cell subsets that produce cytokines. This finding presented here is consistent with previous reports describing KIR2DL4 as predominantly inducing cytokine produc-tion and only little cytotoxicity (20–24). Furthermore, it is possi-ble that this enhanced NK cell functionality affects SIV disease outcome. In fact, functional in vitro studies were reported in which the protective effects ofKIR3DS1/HLA-B Bw4-80I combi-nations on delaying progression to AIDS and lowering HIV-1 rep-lication may be due to stronger NK cell-mediated viral inhibition and increased degranulation and IFN-␥secretion starting in the early phases of HIV-1 infection (4). In contrast, some studies also
FIG 4Secretion of IFN-␥inin vitro-stimulated NK cell subsets from rhesus macaques with different numbers ofKIR2DL4copies. Peripheral blood mononuclear cells from uninfected (A) and SIVmac251-infected rhesus macaques on day 28 postchallenge (B) were stimulated with K562 cells for 6 h at an effector-to-target-ratio of 10:1. IFN-␥release following stimulation was measured in CD16⫹, CD56⫹, and double-negative (DN) NK cell subsets. Insufficient numbers of CD56⫹ NK cells were acquired to allow an analysis. Horizontal bars indicate medians. Analysis was performed using the Mann-Whitney U test.
on November 7, 2019 by guest
http://jvi.asm.org/
was observed in a relatively small cohort of 11 rhesus monkeys, with three of these animals having one copy ofKIR2DL4. While the observation thatKIR2DL4CNV may influence the function-ality of NK cells in such a small cohort might indicate its biological significance, a larger cohort needs to be investigated to further explore this finding. Furthermore, the association between
KIR2DL4CNV and CD4⫹T-cell decline with a concurrent lack of
an association with CM CD4⫹T-cell loss may be explained by the small cohort size.
Taken together, the data presented in these studies provide further evidence that NK cells play a critical role in affecting SIV disease course and strongly support the importance of activating KIRs in modulating NK cell responses during SIV infection. The present studies add a novel aspect to the effects of KIR diversity in an AIDS virus infection with the numbers of activatingKIR2DL4
copies being associated with CD4⫹T-cell decline and function of particular NK cell subsets during acute SIV infection and may thereby suggest that altered NK cell reactivity may be the under-lying mechanism of the association betweenKIR2DL4CNV and a better preservation of CD4⫹T cells.
ACKNOWLEDGMENTS
Research reported in this publication was supported by the National In-stitute of Allergy and Infectious Diseases of the National InIn-stitutes of Health and by the Center for HIV/AIDS Vaccine Immunology, grant number U01-AI067854-07.
The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. We thank the NIAID Vaccine Research Center, Dan Barouch, and James Whitney for providing us with the rhesus macaque peripheral blood mononuclear cells and So-Yon Lim for help with theTRIM5typing.
REFERENCES
1.Vilches C, Parham P.2002. KIR: diverse, rapidly evolving receptors of innate and adaptive immunity. Annu. Rev. Immunol.20:217–251. 2.Martin MP, Gao X, Lee JH, Nelson GW, Detels R, Goedert JJ,
Buch-binder S, Hoots K, Vlahov D, Trowsdale J, Wilson M, O’Brien SJ, Carrington M.2002. Epistatic interaction between KIR3DS1 and HLA-B delays the progression to AIDS. Nat. Genet.31:429 – 434.
3.Qi Y, Martin MP, Gao X, Jacobson L, Goedert JJ, Buchbinder S, Kirk GD, O’Brien SJ, Trowsdale J, Carrington M.2006. KIR/HLA pleiotro-pism: protection against both HIV and opportunistic infections. PLoS Pathog.2:e79. doi:10.1371/journal.ppat.0020079.
4.Alter G, Martin MP, Teigen N, Carr WH, Suscovich TJ, Schneidewind A, Streeck H, Waring M, Meier A, Brander C, Lifson JD, Allen TM, Carrington M, Altfeld M.2007. Differential natural killer cell-mediated inhibition of HIV-1 replication based on distinct KIR/HLA subtypes. J. Exp. Med.204:3027–3036.
5.Barbour JD, Sriram U, Caillier SJ, Levy JA, Hecht FM, Oksenberg JR.
2007. Synergy or independence? Deciphering the interaction of HLA class I and NK cell KIR alleles in early HIV-1 disease progression. PLoS Pathog.
3:e43. doi:10.1371/journal.ppat.0030043.
number variation of KIR genes influences HIV-1 control. PLoS Biol.
9:e1001208. doi:10.1371/journal.pbio.1001208.
10. Carrington M, Alter G. 2012. Innate immune control of HIV. Cold Spring Harbor Perspect. Med. 2:a007070. doi:10.1101/cshperspect .a007070.
11. Hellmann I, Lim SY, Gelman RS, Letvin NL. 2011. Association of activating KIR copy number variation of NK cells with containment of SIV replication in rhesus monkeys. PLoS Pathog.7:e1002436. doi:10.1371 /journal.ppat.1002436.
12. Hershberger KL, Shyam R, Miura A, Letvin NL.2001. Diversity of the killer cell Ig-like receptors of rhesus monkeys. J. Immunol.166:4380 – 4390.
13. Yusa S, Catina TL, Campbell KS.2002. SHP-1- and phosphotyrosine-independent inhibitory signaling by a killer cell Ig-like receptor cytoplas-mic domain in human NK cells. J. Immunol.168:5047–5057.
14. Faure M, Long EO.2002. KIR2DL4 (CD158d), an NK cell-activating receptor with inhibitory potential. J. Immunol.168:6208 – 6214. 15. Kikuchi-Maki A, Yusa S, Catina TL, Campbell KS.2003. KIR2DL4 is an
IL-2-regulated NK cell receptor that exhibits limited expression in hu-mans but triggers strong IFN-gamma production. J. Immunol.171:3415– 3425.
16. Blokhuis JH, van der Wiel MK, Doxiadis GG, Bontrop RE. 2009. Evidence for balancing selection acting on KIR2DL4 genotypes in rhesus macaques of Indian origin. Immunogenetics61:503–512.
17. Fry AM, Lanier LL, Weiss A.1996. Phosphotyrosines in the killer cell inhibitory receptor motif of NKB1 are required for negative signaling and for association with protein tyrosine phosphatase 1C. J. Exp. Med.184: 295–300.
18. Bruhns P, Marchetti P, Fridman WH, Vivier E, Daeron M. 1999. Differential roles of N- and C-terminal immunoreceptor tyrosine-based inhibition motifs during inhibition of cell activation by killer cell inhibi-tory receptors. J. Immunol.162:3168 –3175.
19. Long EO.1999. Regulation of immune responses through inhibitory re-ceptors. Annu. Rev. Immunol.17:875–904.
20. Cooper MA, Fehniger TA, Caligiuri MA.2001. The biology of human natural killer-cell subsets. Trends Immunol.22:633– 640.
21. Cooper MA, Fehniger TA, Turner SC, Chen KS, Ghaheri BA, Ghayur T, Carson WE, Caligiuri MA.2001. Human natural killer cells: a unique innate immunoregulatory role for the CD56(bright) subset. Blood97: 3146 –3151.
22. Rajagopalan S, Fu J, Long EO.2001. Cutting edge: induction of IFN-gamma production but not cytotoxicity by the killer cell Ig-like receptor KIR2DL4 (CD158d) in resting NK cells. J. Immunol.167:1877–1881. 23. Kikuchi-Maki A, Catina TL, Campbell KS.2005. Cutting edge: KIR2DL4
transduces signals into human NK cells through association with the Fc receptor gamma protein. J. Immunol.174:3859 –3863.
24. Rajagopalan S, Bryceson YT, Kuppusamy SP, Geraghty DE, van der Meer A, Joosten I, Long EO.2006. Activation of NK cells by an endocy-tosed receptor for soluble HLA-G. PLoS Biol.4:e9. doi:10.1371/journal .pbio.0040009.
25. Degenhardt JD, de Candia P, Chabot A, Schwartz S, Henderson L, Ling B, Hunter M, Jiang Z, Palermo RE, Katze M, Eichler EE, Ventura M, Rogers J, Marx P, Gilad Y, Bustamante CD.2009. Copy number varia-tion of CCL3-like genes affects rate of progression to simian-AIDS in rhesus macaques (Macaca mulatta). PLoS Genet.5:e1000346. doi:10.1371 /journal.pgen.1000346.
26. Livak KJ, Schmittgen TD.2001. Analysis of relative gene expression data
on November 7, 2019 by guest
http://jvi.asm.org/
using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods25:402– 408.
27. Pfaffl MW.2001. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res.29:e45. doi:10.1093/nar/29.9.e45. 28. Bostik P, Kobkitjaroen J, Tang W, Villinger F, Pereira LE, Little DM,
Stephenson ST, Bouzyk M, Ansari AA.2009. Decreased NK cell fre-quency and function is associated with increased risk of KIR3DL allele polymorphism in simian immunodeficiency virus-infected rhesus ma-caques with high viral loads. J. Immunol.182:3638 –3649.
29. Chaichompoo P, Bostik P, Stephenson S, Udompunturuk S, Kobkit-jaroen J, Pattanapanyasat K, Ansari AA.2010. Multiple KIR gene poly-morphisms are associated with plasma viral loads in SIV-infected rhesus macaques. Cell. Immunol.263:176 –187.
30. Webster RL, Johnson RP.2005. Delineation of multiple subpopulations of natural killer cells in rhesus macaques. Immunology115:206 –214. 31. Reeves RK, Gillis J, Wong FE, Yu Y, Connole M, Johnson RP.2010.
CD16- natural killer cells: enrichment in mucosal and secondary lym-phoid tissues and altered function during chronic SIV infection. Blood
115:4439 – 4446.
32. Williams F, Maxwell LD, Halfpenny IA, Meenagh A, Sleator C, Curran MD, Middleton D.2003. Multiple copies of KIR 3DL/S1 and KIR 2DL4 genes identified in a number of individuals. Hum. Immunol.64:729 –732. 33. Traherne JA, Martin M, Ward R, Ohashi M, Pellett F, Gladman D, Middleton D, Carrington M, Trowsdale J.2010. Mechanisms of copy
number variation and hybrid gene formation in the KIR immune gene complex. Hum. Mol. Genet.19:737–751.
34. Gaudieri S, DeSantis D, McKinnon E, Moore C, Nolan D, Witt CS, Mallal SA, Christiansen FT.2005. Killer immunoglobulin-like receptors and HLA act both independently and synergistically to modify HIV dis-ease progression. Genes Immun.6:683– 690.
35. Goodridge JP, Witt CS, Christiansen FT, Warren HS.2003. KIR2DL4 (CD158d) genotype influences expression and function in NK cells. J. Immunol.171:1768 –1774.
36. Blokhuis JH, van der Wiel MK, Doxiadis GG, Bontrop RE.2010. The mosaic of KIR haplotypes in rhesus macaques. Immunogenetics62:295–306. 37. Witt CS, Martin A, Christiansen FT.2000. Detection of KIR2DL4 alleles by sequencing and SSCP reveals a common allele with a shortened cyto-plasmic tail. Tissue Antigens56:248 –257.
38. Jennes W, Verheyden S, Demanet C, Menten J, Vuylsteke B, Nkengasong JN, Kestens L.2011. Low CD4⫹T cell counts among African HIV-1 infected subjects with group B KIR haplotypes in the absence of specific inhibitory KIR ligands. PLoS One6:e17043. doi:10.1371/journal.pone.0017043.
39. O’Connell KA, Han Y, Williams TM, Siliciano RF, Blankson JN.2009. Role of natural killer cells in a cohort of elite suppressors: low frequency of the protective KIR3DS1 allele and limited inhibition of human immuno-deficiency virus type 1 replication in vitro. J. Virol.83:5028 –5034. 40. Hosmalin A, Lebon P. 2006. Type I interferon production in
HIV-infected patients. J. Leukoc. Biol.80:984 –993.