0022-538X/06/$08.00⫹0 doi:10.1128/JVI.00006-06
Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Effect of Apolipoprotein E on the Cerebral Load of Latent
Herpes Simplex Virus Type 1 DNA
Javier S. Burgos,†* Carlos Ramirez,† Isabel Sastre, and Fernando Valdivieso*
Departamento de Biologı´a Molecular and Centro de Biologı´a Molecular Severo Ochoa (CSIC-UAM),
Universidad Auto´noma de Madrid, Madrid, Spain
Received 2 January 2006/Accepted 10 March 2006
Herpes simplex virus type 1 (HSV-1) is neurotropic and enters a latent state lasting the lifetime of the host. This pathogen has recently been proposed as a risk factor for Alzheimer’s disease (AD) in conjunction with apolipoprotein E4 (ApoE4). In a murine acute infection model, we showed that viral neuroinvasiveness depends directly on the overall ApoE dosage and especially on the presence of isoform ApoE4. If an interaction between ApoE and HSV-1 is involved in AD, it may occur during latency rather than during acute infection. Certainly, ApoE plays an important role in late-onset AD, i.e., at a time in life when the majority of people harbor HSV-1 in their nervous system. In the present work, wild-type, APOE knockout, APOE3, and APOE4 transgenic mice were used to analyze the influence of the ApoE profile on the levels of latent virus DNA. The knockout mice had significantly lower concentrations of the virus in the nervous system than the wild-type mice, while the APOE4 mice had very high levels in the brain compared to the APOE3 animals. ApoE4 seems to facilitate HSV-1 latency in the brain much more so than ApoE3. The APOE dosage correlated directly with the HSV-1 DNA concentration in the brain, strengthening the hypothesis that HSV-1, together with ApoE, might be involved in AD.
Most cases of Alzheimer’s disease (AD) (⬎95%) are late
onset, are of the sporadic type, and appear to be associated with no known genetic markers (22). A variety of potential risk factors, both genetic and environmental, have been examined, but the etiology and pathogenesis of sporadic AD are still poorly understood. The idea that both kinds of factors interact and contribute to the development of AD is gaining wide acceptance. Although many proposed causes have been stud-ied, only two factors are widely accepted as being associated with nonfamilial disease: age (16, 19, 35, 38) and the possession
of the apolipoprotein Eε4 allele (APOE4) (4, 41, 50).
Apolipoprotein E (ApoE), a major component of very-low-density lipoproteins, may play an important modulatory func-tion in the central nervous system (CNS): in the peripheral nervous system, several apolipoproteins are involved in lipid transport, but in the CNS, there is less redundancy (32, 40). ApoE is involved in the mobilization and redistribution of lipids and cholesterol during neuronal growth and repair (12, 42), the long-distance systemic and cerebrospinal transport of lipids (17), and the promotion of synaptic plasticity following neuronal injury (12). The human APOE gene has three
com-mon alleles,ε2,ε3, andε4, which encode the ApoE2, ApoE3,
and ApoE4 protein isoforms, respectively (43). Sequencing studies have shown ApoE3 and ApoE4 to differ by just a single amino acid (Cys or Arg at residue 112, respectively) (48, 52). APOE3 is the most common allele. APOE4 has been associ-ated with a high risk for AD in many studies, irrespective of race or geographical location (10, 17, 28, 31, 37). Nevertheless,
the APOE4 allele alone is neither necessary nor sufficient for the development of AD (13, 44).
Although the involvement of an infectious agent in the eti-ology of AD is far from fully demonstrated, herpes simplex virus type 1 (HSV-1) has recently been implicated in the patho-genesis of this disease (22, 25, 34). Several coincidences rein-force this association: HSV-1 is neurotropic (in line with the neurological nature of AD), is ubiquitous in the human pop-ulation (36), and infects and causes pathological changes in the brain regions mainly affected by AD (1, 2). One of the impor-tant features of this virus is its ability to establish a latent infection and to reside in the nervous system over the lifetime of the host. A prominent characteristic of the virus is its ability to reactivate from latency and cause viral shedding and/or recurrent disease. HSV-1 has been found in the brains of many patients with AD but has also been found in normal brains of elderly people (25, 26), suggesting that HSV-1 infection is not independently associated with AD (22). However, while the incidence of AD is not increased in those with HSV-1 DNA or the APOE4 allele alone (25), it is highest in the carriers of this allele who also harbor HSV-1 DNA in the CNS.
Using a murine model of acute hematogenous HSV-1 infec-tion, we previously reported that viral neuroinvasion was re-duced in mice lacking ApoE compared to wild-type mice and that the ApoE dose was directly linked to the HSV-1 DNA concentration detected in the nervous system (8). In a further study, we showed that during acute infection, ApoE4 was more efficient than ApoE3 in promoting colonization of the brain by HSV-1 (comparisons were made at the time when this organ presented the highest viral loads) (7). If an interaction between ApoE and HSV-1 has anything to do with AD, this may occur during latency.
HSV-1 DNA may be present in the majority of elderly hu-man brains (26). Almost all adults show evidence of having
* Corresponding author. Mailing address: Lab CX340, Centro de Biologı´a Molecular, Universidad Auto´noma de Madrid, Cantoblanco, 28049 Madrid, Spain. Phone: 34 91 4978471. Fax: 34 91 4974870. E-mail for Javier S. Burgos: [email protected]. E-mail for Fernando Valdivieso: [email protected].
† J.S.B. and C.R. contributed equally to this work.
5383
on November 8, 2019 by guest
http://jvi.asm.org/
been infected by HSV-1, its genome residing latently in the trigeminal ganglia and in the CNS of both healthy individuals and those suffering from neurological disease (14). The trigem-inal ganglion is the primary site of HSV-1 latency, although other sites including the sensory neurons (33) and other sen-sory ganglia such as the nodose ganglion of the vagus nerve (18), the dorsal root ganglia (20), the sympathetic ganglia (51), and the brain may be involved (3, 47). The persistence of HSV-1 in nonneuronal tissue has also been suggested but re-mains controversial (33).
Latency is a poorly understood phenomenon, and the host factors that contribute to its establishment and maintenance are far from fully known. The present work describes how the ApoE profile affects the levels of latent HSV-1 and shows that ApoE4 in particular is associated with increased amounts of latent virus in the brain. The relationship between these two risk factors for AD could be of great importance in the patho-genesis of this disease.
MATERIALS AND METHODS
Inoculation and dissection.All experiments were performed in accordance with the guidelines of the 1986 European Community Animals Act (Scientific Procedures). All animals underwent a period of quarantine. Strict precautions were taken to prevent contamination during inoculation and dissection.
The animals used were 37 14-week-old C57BL/6 female mice: 19 wild-type mice (Harlan, Barcelona, Spain), 6 APOE knockout mice, 6 APOE4 human transgenic mice, and 6 APOE3 human transgenic mice. All mice were marked, examined, and analyzed individually. APOE knockout mice (B6.129P2-Apoetm1UncN11), generated using a protocol described previously by Piedrahita et al. (39), were obtained from Taconic M&B (Bomholtrej, Denmark). To compare the isoform-specific effects of ApoE3 and ApoE4 on HSV-1 latent infection, mice transgenic for human APOE3 (B6.129P2-ApoeTm1(APOE3)Mae N8) and APOE4 (B6.129P2-ApoeTm1(APOE4)Mae
N8) were purchased from the same supplier. These mice were generated by replacing their APOE gene with the corresponding human homolog as described previously by Knouff et al. and Sullivan et al. (30, 49). Such animals are useful for in vivo studies of the human ApoE isoforms since the replacement of the mouse APOE does not alter the murine regulatory sequence. Consequently, the mice express the human ApoE isoforms with tissue distribu-tions and levels very close to those of endogenous murine ApoE.
HSV-1 was propagated and titrated by plaque assay in confluent monolayers of Vero cells (8). HSV-1 strain KOS (kindly supplied by L. Carrasco) was used in all experiments. Female mice were intraperitoneally inoculated with doses of 105
, 106
, or 107
PFU of virus suspension as previously described (6). The mouse strain and the female gender were selected in order to compare the results with previously published data (6–8). The female gender was also used because this sex shows greater viral infectivity than males (6). An extra set of mock-infected animals was used as controls. Since overt infection subsides 2 weeks after inoc-ulation and all HSV-1 present is in a latent state by 4 weeks after inocinoc-ulation (33), the mice were sacrificed at 37 days postinfection (when latent infection was assured), and their organs were carefully removed and frozen at⫺70°C. The HSV-1 DNA concentrations in whole blood, the adrenal glands, the gonads, the spinal cord, the trigeminal ganglia, and the brain were then determined. For more precise analyses, the brain samples were subdivided into four regions: the midbrain (including the midbrain and nearby structures such as the pons, the me-dulla, and the superior and inferior colliculi), the ventricles (including the third and lateral ventricles, the thalamus, the hypothalamus, the preoptic area, and the striatum), the cerebral cortex (including the cortex; the temporal, frontal, pari-etal, and occipital lobes; the hippocampus; the corpus callosum; and the olfactory bulbs), and the cerebellum.
HSV-1 DNA quantification in tissue homogenates.DNA from homogenized samples was extracted by conventional methods (DNeasy 96 tissue kit [QIAGEN Sciences, MD] or QIAamp 96 DNA blood kit [QIAGEN GmbH, Hilden, Ger-many]). The concentration of HSV-1 DNA in different organs was then quanti-fied by real-time quantitative PCR with an ABI Prism 7900HT SD system (Applied Biosystems) using a Custom TaqMan assay (a specific assay for a sequence belonging to the US12 viral gene). Reactions were performed under universal conditions using TaqMan Universal PCR master mix (Applied Biosys-tems). The exact same PCR protocol was used to quantify the mouse genomic
DNA using an Assay-On-Demand probe specific for the GAPDH
(glyc-eraldehyde-3-phosphate dehydrogenase) housekeeping gene (item no.
Mm99999915_g1; Applied Biosystems). This generated an amplicon of 107 bp from the GenBank accession number NM_001001303.1 transcript. An appropri-ate concentration range of virus was used for the optimization of the standard curve; the viral DNA concentration was expressed in terms of viral copy num-bers. PCR calibration was performed by amplification of the GAPDH house-keeping gene from a concentration range of mouse genomic DNA (results are expressed as nanograms of host DNA). Viral DNA loads were also corroborated by amplification of the thymidine kinase viral gene and the-actin housekeeping gene by real-time PCR using a LightCycler rapid thermal cycler (Roche Diag-nostics Ltd., Lewes, United Kingdom) and a LightCycler FastStart DNA Master SYBR Green I kit (catalog no. 3 003 230; Roche, Germany) as previously described (6–9). Each experiment was performed in triplicate. Melting curve analyses, agarose and acrylamide gel electrophoresis, restriction analysis, and nested PCR confirmed the specificity of the products (5, 9).
Statistical analysis.Fisher’s exact test was used to compare viral DNA con-centrations. Significance was set at aPvalue of⬍0.05.
RESULTS
Effect of viral inoculation dose on latent HSV-1 DNA levels.
To evaluate the influence of the injected viral dose on the final concentrations of latent viral DNA, wild-type female mice
were inoculated with between 105and 107PFU. Figure 1 shows
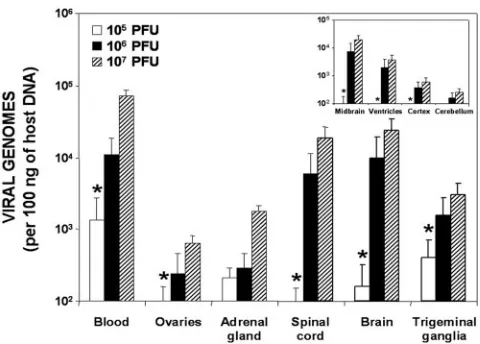
[image:2.585.302.542.67.239.2]the viral DNA load of several organs in relation to the admin-istered dose. The HSV-1 DNA concentration was always di-rectly dependent on the inoculation dose, with a similar pat-tern of infection seen for all doses analyzed. Independent of the dose, the organs with the highest HSV-1 DNA levels were the nervous system (spinal cord, brain, and trigeminal ganglia, where the virus establishes latency) and the blood (where amounts of virus produced during latency eventually arrive from the trigeminal ganglia) (6). The viral DNA levels of the four brain regions analyzed (Fig. 1, right inset) were also in-oculation dose dependent. The midbrain had the highest viral DNA load for every dose tested, followed by the ventricles,
FIG. 1. Quantification of viral DNA loads during latency with re-spect to the viral dose injected into wild-type mice. The bar graph represents the viral copy number detected in each organ expressed on a logarithmic scale. The inset shows the areas of the brain analyzed. Values are the means⫾standard errors of the means (SEM) of the quantity of viral DNA (expressed as viral genomes and normalized with respect to the quantity of mouse genomes in 100 ng of host DNA). Fisher’s exact test was used to compare the HSV-1 DNA concentra-tions achieved with the different doses (always compared to a dose of 106PFU) (
*
P⬍0.05).on November 8, 2019 by guest
http://jvi.asm.org/
cortex, and cerebellum. This order was consistent in all
exper-iments. For further experiments, an inoculation dose of 106
PFU per animal was selected.
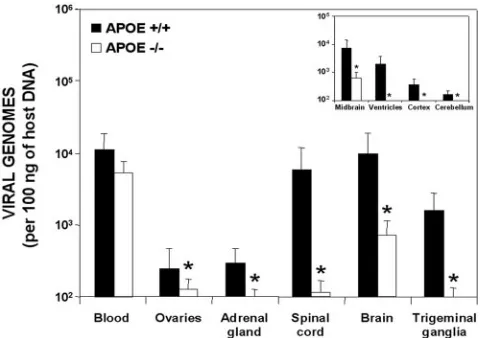
Effect of ApoE on HSV-1 latent infection.At 37 days postin-fection, HSV-1 was preferentially detected in the nervous
sys-tem and in the whole blood of the wild-type (or APOE⫹/⫹)
mice; the ovaries and adrenal glands showed lower levels of viral DNA. When DNA loads in the different organs of the
APOE⫹/⫹ and APOE knockout (or APOE⫺/⫺) mice were
compared, those of the wild-type mice showed significantly higher concentrations, except for whole blood (Fig. 2). In both backgrounds, the organ with the highest viral DNA load after the blood was the brain, indicating preferential viral tropism towards this organ and the establishment of latency in the CNS. The differences between HSV-1 DNA concentrations in
APOE⫹/⫹ and APOE⫺/⫺ mice were dramatic in the brain
(wild-type-to-knockout ratio, 13.7;P⬍0.001), spinal cord
(ra-tio, 52.1;P⬍0.001), and trigeminal ganglia (ratio, 23.7;P⬍
0.001). In the dissected brains of APOE⫹/⫹mice, the order of
viral DNA concentrations was midbrain, ventricles, cortex, and
cerebellum. APOE⫺/⫺mice had detectable levels of viral DNA
only in the midbrain but at considerably lower concentrations
(ratio, 11.7;P⬍0.001), and no viral DNA was detected in the
distal regions of their brains (Fig. 2, inset). The concentra-tions in the ovaries and adrenal glands, although very low,
were significantly different (P⬍ 0.05) in the wild-type and
knockout mice.
Effect of ApoE isoform on HSV-1 latent infection.Striking isoform-dependent differences were seen in the viral DNA concentrations of the infected organs (Fig. 3). In general, higher viral DNA loads were detected in the organs of APOE4 mice than in the organs of APOE3 mice. The blood and the nervous system were the sites where the virus was preferen-tially detected. The nervous system viral DNA concentrations showed significant genotype-dependent differences; the brain
APOE4-to-APOE3 viral DNA ratio was 13.6 (P⬍0.001), that
of the spinal cord was 180.7 (P ⬍ 0.001), and that of the
trigeminal ganglion was over 200 (P⬍0.001). Differences in
brain region viral DNA loads were found between APOE4 and APOE3 mice (Fig. 3, inset). HSV-1 DNA levels in the ventri-cles, cortex, and cerebellum of APOE3 mice were undetect-able, and only low levels were found in the midbrain of APOE3
mice compared to those of APOE4 mice (ratio, 12.7; P ⬍
0.001). However, no significant differences were seen between these groups with respect to blood and adrenal gland viral DNA loads. In conclusion, the present data show that HSV-1 DNA levels during viral latency in the brain are more than 10 times higher in the APOE4 background than in the APOE3 background. Interestingly, the differences seen between the APOE4 and APOE3 mice were comparable to those observed
between APOE⫹/⫹ and APOE⫺/⫺ animals (compare Fig. 2
and 3). The human APOE3 genetic background seemed to reduce the establishment of latency in a manner similar to the lack of APOE. Finally, in the majority of organs, the APOE4 and wild-type mice differed very little with respect to HSV-1 DNA concentrations. Moreover, the order of viral DNA levels in the different brain regions of these mice was identical.
DISCUSSION
[image:3.585.44.284.69.238.2]HSV-1, one of the most commonly encountered pathogens, has been related to several neurological diseases, especially AD (23, 25, 34). Infection with this virus, which can on occa-sion cause long-term neurodevelopmental damage in neo-nates, is characterized by the pathogen entering a latent state in the nervous system that lasts the lifetime of the host. The foremost site for this latent infection is the trigeminal ganglia, a location very close to the brain region where the protein tangles associated with AD first appear. Interestingly, HSV-1
FIG. 2. Quantification of viral DNA levels in relation to APOE dosage. Black bars represent the wild-type mice, and white bars rep-resent the APOE knockout group (all mice were inoculated with 106 PFU of HSV-1). The inset shows the areas of the brain analyzed. Values are the means⫾SEM of the quantity of viral DNA (expressed as viral genomes and normalized with respect to the quantity of mouse genomes in 100 ng of host DNA). Fisher’s exact test was used to compare the values for the different APOE dosage groups (
*
P⬍0.05).FIG. 3. Quantification of HSV-1 DNA concentrations related to the ApoE isoform. Black bars represent the APOE4 group, and white bars represent the APOE3 group (all animals were injected with 106 PFU of HSV-1). The inset shows the areas of the brain analyzed. Values are the means⫾SEM of the quantity of viral DNA (expressed as viral genomes and normalized with respect to the quantity of mouse genomes in 100 ng of host DNA). Fisher’s exact test was used to compare the values for the different ApoE isoform groups (
*
P⬍0.05).on November 8, 2019 by guest
http://jvi.asm.org/
[image:3.585.305.538.71.236.2]DNA is detectable in regions of the brain that are affected by this disease (2).
We previously reported that acute infection of the brain by HSV-1 is influenced by the ApoE dosage and isoform (6–8). However, if HSV-1 is truly involved in AD, it may be associ-ated with latency rather than acute infection. The establish-ment of the latent state is regulated by viral and host factors (both cellular and immune), not all of which are known or understood (15); the present results suggest, however, that the ApoE dosage and isoform play a critical role.
HSV-1 becomes latent in the neurons of both the peripheral nervous system and the CNS (11). Although the trigeminal ganglia represent the primary site where HSV-1 resides in the latent form (33), other locations have been documented, in-cluding the nodose ganglion of the vagus nerve (18), the dorsal root ganglia (20), the sympathetic ganglia (51), and the brain (3, 47). Thus, the spinal cord and brain could act as reservoirs of latent virus. Most, if not all, human brains harbor latent viral genomes (14, 26, 27), but their quiescence or, conversely, their ability to cause harm may depend on their exact location, the immune status of the host, and their copy number (in which ApoE could play an important role). It has been shown that the HSV-1 genome copy number is positively correlated with the ability to reactivate in vivo in neurons of the trigeminal ganglia (46). After reactivation, recurrent disease may develop in neu-rons harboring the latent virus; in fact, HSV-1 has been impli-cated in necrotic and apoptotic neural cell death in the trigem-inal ganglia of acutely infected experimental animals (29, 53). Recently, we provided direct evidence that HSV-1 reactivates molecularly in the hippocampal neurons (5); this could be the cause of the local degenerative neuropathological complex that characterizes AD.
Possession of the APOE4 allele has been associated with the development of AD (10, 45). However, this alone is neither essential nor sufficient; interaction with another factor appears to be necessary. Itzhaki et al. previously showed that the rela-tive risk of developing AD for those posirela-tive for HSV-1 DNA in the brain and who carried an APOE4 allele was considerably higher than that for individuals with only one (or none) of these factors (22, 24, 25). In addition, Itabashi et al. previously showed that patients with AD were more likely to be positive for both HSV and the APOE4 allele than for one factor alone (21). These results suggest that the combination of these two factors is strongly associated with the development of AD. However, the latter studies do not show how this ApoE iso-form influences infection or predisposes those infected to AD, and until now, it was unclear whether a direct correlation between viral DNA levels in the brain and the possession of the APOE4 allele existed. ApoE4, perhaps unlike other ApoE isoforms, may increase the risk of AD by increasing HSV-1 viral loads in the nervous system.
The present results show that ApoE plays an important role in latent virus DNA concentrations. In APOE-deficient mice, HSV-1 DNA levels were significantly lower than those in wild-type mice not only in the brain (13-fold) but in every organ analyzed, except for whole blood. Furthermore, the APOE allele possessed by infected mice influenced the efficiency of the establishment (or maintenance) of latent infection: the encephalons of APOE4 animals contained significantly more viral DNA (13-fold) than those of APOE3 mice. Interestingly,
the viral DNA ratios seen for APOE⫹/⫹:APOE⫺/⫺ and
APOE4:APOE3 mice were identical and comparable to the ratios observed in acute infection (7, 8). This agrees with the finding that individuals positive for HSV-1 DNA in the brain and who carry the APOE4 allele are 12 times more likely to develop AD than those possessing only one (or neither) factor (24). The present results regarding the effect of ApoE dosage (presence compared to absence) on latent HSV-1 infection of the brain also agree with the generally accepted dose effect reported for APOE4 in AD (10). In the establishment of latent
infection, the APOE⫹/⫹and APOE4 genotypes were
associ-ated with high HSV-1 DNA concentrations in the nervous
system, while APOE⫺/⫺and APOE3 were associated with low
cerebral loads.
This work is the first to show that latent cerebral HSV-1 infection depends on the APOE profile. This may be important in the onset of AD since the incidence of this disease has been related to the possession of the APOE4 allele and the presence of HSV-1 DNA in the CNS; it is possible, therefore, that these agents are cofactors that promote the disease. This new evi-dence strongly reinforces the idea that HSV-1 is an etiological factor in AD.
ACKNOWLEDGMENTS
This work was supported by a grant from the Obra Social Caja Madrid to the Asociacio´n de Familiares de Enfermos de Alzheimer and by an institutional grant from the Fundacio´n Areces to the Centro de Biologı´a Molecular Severo Ochoa.
We thank F. Mayor for his continuous encouragement and help and L. Carrasco for providing the HSV-1 KOS strain.
REFERENCES
1.Ball, M. J.1986. Herpesvirus in the hippocampus as a cause of Alzheimer’s disease. Arch. Neurol.43:313.
2.Ball, M. J.1982. Limbic predilection in Alzheimer dementia: is reactivated herpesvirus involved? Can. J. Neurol. Sci.9:303–306.
3.Baringer, J. R., and P. Pisani.1994. Herpes simplex virus genomes in human nervous system tissue analyzed by polymerase chain reaction. Ann. Neurol. 36:823–829.
4.Bretsky, P., J. M. Guralnik, L. Launer, M. Albert, and T. E. Seeman.2003. The role of APOE-epsilon4 in longitudinal cognitive decline: MacArthur Studies of Successful Aging. Neurology60:1077–1081.
5.Burgos, J. S., C. Ramirez, F. Guzman-Sanchez, J. M. Alfaro, I. Sastre, and F. Valdivieso.2006. Hematogenous vertical transmission of herpes simplex virus type 1 in mice. J. Virol.80:2823–2831.
6.Burgos, J. S., C. Ramirez, I. Sastre, J. M. Alfaro, and F. Valdivieso.2005. Herpes simplex virus type 1 infection via the bloodstream with apolipopro-tein E dependence in the gonads is influenced by gender. J. Virol.79:1605– 1612.
7.Burgos, J. S., C. Ramirez, I. Sastre, M. J. Bullido, and F. Valdivieso.2003. ApoE4 is more efficient than E3 in brain access by herpes simplex virus type 1. Neuroreport14:1825–1827.
8.Burgos, J. S., C. Ramirez, I. Sastre, M. J. Bullido, and F. Valdivieso.2002. Involvement of apolipoprotein E in the hematogenous route of herpes sim-plex virus type 1 to the central nervous system. J. Virol.76:12394–12398. 9.Burgos, J. S., C. Ramirez, R. Tenorio, I. Sastre, and M. J. Bullido.2002.
Influence of reagents formulation on real-time PCR parameters. Mol. Cell. Probes16:257–260.
10.Corder, E. H., A. M. Saunders, W. J. Strittmatter, D. E. Schmechel, P. C. Gaskell, G. W. Small, A. D. Roses, J. L. Haines, and M. A. Pericak-Vance. 1993. Gene dose of apolipoprotein E type 4 allele and the risk of Alzheimer’s disease in late onset families. Science261:921–923.
11.Deatly, A. M., J. G. Spivack, E. Lavi, D. R. O’Boyle II, and N. W. Fraser. 1988. Latent herpes simplex virus type 1 transcripts in peripheral and central nervous system tissues of mice map to similar regions of the viral genome. J. Virol.62:749–756.
12.DeMattos, R. B., L. K. Curtiss, and D. L. Williams.1998. A minimally lipidated form of cell-derived apolipoprotein E exhibits isoform-specific stimulation of neurite outgrowth in the absence of exogenous lipids or lipoproteins. J. Biol. Chem.273:4206–4212.
13.Farrer, L. A., L. A. Cupples, J. L. Haines, B. Hyman, W. A. Kukull, R.
on November 8, 2019 by guest
http://jvi.asm.org/
Mayeux, R. H. Myers, M. A. Pericak-Vance, N. Risch, C. M. van Duijn, et al. 1997. Effects of age, sex, and ethnicity on the association between apoli-poprotein E genotype and Alzheimer disease. A meta-analysis. JAMA278: 1349–1356.
14.Fraser, N. W., W. C. Lawrence, Z. Wroblewska, D. H. Gilden, and H. Koprowski.1981. Herpes simplex type 1 DNA in human brain tissue. Proc. Natl. Acad. Sci. USA78:6461–6465.
15.Fraser, N. W., and T. Valyi-Nagy.1993. Viral, neuronal and immune factors which may influence herpes simplex virus (HSV) latency and reactivation. Microb. Pathog.15:83–91.
16.Fukumoto, H., M. Tennis, J. J. Locascio, B. T. Hyman, J. H. Growdon, and M. C. Irizarry.2003. Age but not diagnosis is the main predictor of plasma amyloid beta-protein levels. Arch. Neurol.60:958–964.
17.Gerdes, L. U.2003. The common polymorphism of apolipoprotein E: geo-graphical aspects and new pathophysiological relations. Clin. Chem. Lab. Med.41:628–631.
18.Gesser, R. M., T. Valyi-Nagy, S. M. Altschuler, and N. W. Fraser.1994. Oral-oesophageal inoculation of mice with herpes simplex virus type 1 causes latent infection of the vagal sensory ganglia (nodose ganglia). J. Gen. Virol. 75:2379–2386.
19.Hardy, J.2003. Alzheimer’s disease: genetic evidence points to a single pathogenesis. Ann. Neurol.54:143–144.
20.Hill, T. J., H. J. Field, and W. A. Blyth.1975. Acute and recurrent infection with herpes simplex virus in the mouse: a model for studying latency and recurrent disease. J. Gen. Virol.28:341–353.
21.Itabashi, S., H. Arai, T. Matsui, S. Higuchi, and H. Sasaki.1997. Herpes simplex virus and risk of Alzheimer’s disease. Lancet349:1102.
22.Itzhaki, R.2004. Herpes simplex virus type 1, apolipoprotein E and Alzhei-mer’ disease. Herpes11(Suppl. 2):77A–82A.
23.Itzhaki, R. F., C. B. Dobson, and M. A. Wozniak.2004. Herpes simplex virus type 1 and Alzheimer’s disease. Ann. Neurol.55:299–300. (Author’s Reply, 55:300–301.)
24.Itzhaki, R. F., and W. R. Lin.1998. Herpes simplex virus type I in brain and the type 4 allele of the apolipoprotein E gene are a combined risk factor for Alzheimer’s disease. Biochem. Soc. Trans.26:273–277.
25.Itzhaki, R. F., W. R. Lin, D. Shang, G. K. Wilcock, B. Faragher, and G. A. Jamieson.1997. Herpes simplex virus type 1 in brain and risk of Alzheimer’s disease. Lancet349:241–244.
26.Jamieson, G. A., N. J. Maitland, G. K. Wilcock, J. Craske, and R. F. Itzhaki. 1991. Latent herpes simplex virus type 1 in normal and Alzheimer’s disease brains. J. Med. Virol.33:224–227.
27.Jamieson, G. A., N. J. Maitland, G. K. Wilcock, C. M. Yates, and R. F. Itzhaki.1992. Herpes simplex virus type 1 DNA is present in specific regions of brain from aged people with and without senile dementia of the Alzhei-mer type. J. Pathol.167:365–368.
28.Jarvik, G. P., E. M. Wijsman, W. A. Kukull, G. D. Schellenberg, C. Yu, and E. B. Larson.1995. Interactions of apolipoprotein E genotype, total choles-terol level, age, and sex in prediction of Alzheimer’s disease: a case-control study. Neurology45:1092–1096.
29.Johnson, R. P.1998. Viral infections of the nervous system, 2nd ed., p. 133–168. Lippincott Raven, Philadelphia, Pa.
30.Knouff, C., M. E. Hinsdale, H. Mezdour, M. K. Altenburg, M. Watanabe, S. H. Quarfordt, P. M. Sullivan, and N. Maeda.1999. Apo E structure determines VLDL clearance and atherosclerosis risk in mice. J. Clin. Inves-tig.103:1579–1586.
31.Levy-Lahad, E., D. Tsuang, and T. D. Bird.1998. Recent advances in the genetics of Alzheimer’s disease. J. Geriatr. Psychiatry Neurol.11:42–54. 32.Mahley, R. W.1988. Apolipoprotein E: cholesterol transport protein with
expanding role in cell biology. Science240:622–630.
33.Miller, C. S., R. J. Danaher, and R. J. Jacob.1998. Molecular aspects of herpes simplex virus I latency, reactivation, and recurrence. Crit. Rev. Oral Biol. Med.9:541–562.
34.Mori, I., Y. Kimura, H. Naiki, R. Matsubara, T. Takeuchi, T. Yokochi, and Y. Nishiyama.2004. Reactivation of HSV-1 in the brain of patients with familial Alzheimer’s disease. J. Med. Virol.73:605–611.
35.Mufson, E. J., M. D. Ikonomovic, S. D. Styren, S. E. Counts, J. Wuu, S. Leurgans, D. A. Bennett, E. J. Cochran, and S. T. DeKosky.2003. Preser-vation of brain nerve growth factor in mild cognitive impairment and Alz-heimer disease. Arch. Neurol.60:1143–1148.
36.Nahmias, A. J., F. K. Lee, and S. Beckman-Nahmias.1990. Sero-epidemio-logical and -socioSero-epidemio-logical patterns of herpes simplex virus infection in the world. Scand. J. Infect. Dis. Suppl.69:19–36.
37.Nathoo, N., R. Chetty, J. R. van Dellen, and G. H. Barnett.2003. Genetic vulnerability following traumatic brain injury: the role of apolipoprotein E. Mol. Pathol.56:132–136.
38.Pandav, R., H. H. Dodge, S. T. DeKosky, and M. Ganguli.2003. Blood pressure and cognitive impairment in India and the United States: a cross-national epidemiological study. Arch. Neurol.60:1123–1128.
39.Piedrahita, J. A., S. H. Zhang, J. R. Hagaman, P. M. Oliver, and N. Maeda. 1992. Generation of mice carrying a mutant apolipoprotein E gene inacti-vated by gene targeting in embryonic stem cells. Proc. Natl. Acad. Sci. USA 89:4471–4475.
40.Poirier, J., A. Minnich, and J. Davignon.1995. Apolipoprotein E, synaptic plasticity and Alzheimer’s disease. Ann. Med.27:663–670.
41.Roberts, J. S., S. A. LaRusse, H. Katzen, P. J. Whitehouse, M. Barber, S. G. Post, N. Relkin, K. Quaid, R. H. Pietrzak, L. A. Cupples, L. A. Farrer, T. Brown, and R. C. Green.2003. Reasons for seeking genetic susceptibility testing among first-degree relatives of people with Alzheimer disease. Alz-heimer Dis. Assoc. Disord.17:86–93.
42.Rocchi, A., S. Pellegrini, G. Siciliano, and L. Murri.2003. Causative and susceptibility genes for Alzheimer’s disease: a review. Brain Res. Bull.61: 1–24.
43.Roses, A. D.1996. Apolipoprotein E alleles as risk factors in Alzheimer’s disease. Annu. Rev. Med.47:387–400.
44.Rubinsztein, D. C., and D. F. Easton.1999. Apolipoprotein E genetic vari-ation and Alzheimer’s disease. A meta-analysis. Dement. Geriatr. Cogn. Disord.10:199–209.
45.Saunders, A. M., W. J. Strittmatter, D. Schmechel, P. H. George-Hyslop, M. A. Pericak-Vance, S. H. Joo, B. L. Rosi, J. F. Gusella, D. R. Crapper-MacLachlan, M. J. Alberts, et al.1993. Association of apolipoprotein E allele epsilon 4 with late-onset familial and sporadic Alzheimer’s disease. Neurology43:1467–1472.
46.Sawtell, N. M., D. K. Poon, C. S. Tansky, and R. L. Thompson.1998. The latent herpes simplex virus type 1 genome copy number in individual neurons is virus strain specific and correlates with reactivation. J. Virol.72:5343– 5350.
47.Sequiera, L. W., L. C. Jennings, L. H. Carrasco, M. A. Lord, A. Curry, and R. N. Sutton.1979. Detection of herpes-simplex viral genome in brain tissue. Lancetii:609–612.
48.Strittmatter, W. J., and A. D. Roses.1995. Apolipoprotein E and Alzheimer disease. Proc. Natl. Acad. Sci. USA92:4725–4727.
49.Sullivan, P. M., H. Mezdour, Y. Aratani, C. Knouff, J. Najib, R. L. Reddick, S. H. Quarfordt, and N. Maeda.1997. Targeted replacement of the mouse apolipoprotein E gene with the common human APOE3 allele enhances diet-induced hypercholesterolemia and atherosclerosis. J. Biol. Chem.272: 17972–17980.
50.Vega, G. L., M. F. Weiner, A. M. Lipton, K. Von Bergmann, D. Lutjohann, C. Moore, and D. Svetlik.2003. Reduction in levels of 24S-hydroxycholes-terol by statin treatment in patients with Alzheimer disease. Arch. Neurol. 60:510–515.
51.Warren, K. G., S. M. Brown, Z. Wroblewska, D. Gilden, H. Koprowski, and J. Subak-Sharpe.1978. Isolation of latent herpes simplex virus from the superior cervical and vagus ganglions of human beings. N. Engl. J. Med. 298:1068–1069.
52.Weisgraber, K. H., and R. W. Mahley.1996. Human apolipoprotein E: the Alzheimer’s disease connection. FASEB J.10:1485–1494.
53.Whitley, R. J., and B. Roizman.2001. Herpes simplex virus infections. Lancet 357:1513–1518.