JOURNALOFVIROLOGY, Sept. 1977, p. 626-636
Copyright01977 AmericanSocietyforMicrobiology Printed inU.S.A.
Genetic and
Physiological Control of Host Cell Lysis by
Bacteriophage
Lambda
BARRY G. ROLFE ANDJOHN H. CAMPBELL*
Genetics Department,Research SchoolofBiological Sciences,TheAustralian NationalUniversity, Canberra,
A.C.T.2601,Australia, andDepartmentofAnatomy,SchoolofMedicine, University of California,Los
Angeles, California90024*
Receivedforpublication23February 1977
The timing of host cell lysisatthe end ofthe lytic cycleofphageX is under
complex control.The ASproteinstimulateslysis.Anotherphysiological system, the lysis regulator, inhibits lysis from occurring prematurely. The effects of a
seriesofphage and bacterial mutations on these controls aredescribed. They
showthat the Xrex geneplaysa role inregulating lysis under suboptimal growth
conditions. In certain mutant cells, and especially under anaerobic culture
conditions, the rexgene aids in thescheduling of host celllysis. The data also
suggestthat thelysis regulator maycontrol the transition of theASproteinfrom aninactiveto anactivestate.
In a series of papers we are reporting our
studiesonthe control of host cell lysis by bacte-riophage X.When this phage lyrically infects its host, Escherichia coli, itreplicates intracellu-larly for aperiod oftime knownasthe latent period. The cells then lyse at a genetically scheduled time. Controloverthe timingof lysis is complex. An important element is the AS gene product, which is a positive effector of
lysis. Itisthoughttoaidinthe sudden
trans-missionof aphage-specified lytic enzyme (endo-lysin) across the membrane to the cell wall, whereithydrolyzescross-linkages (13, 14, 19). The AS protein ismade throughout the second half of the latent period, butisinactive or inef-fective until the scheduled time forlysis.
Dur-ingthistime it canbeartificially activated by
cyanide. Cells infected with wild-type phage but not with AS mutants lyse promptly when exposed to cyanide later in the latent period (14).
Wehave describedaninhibitory system,the
lysis regulator, which prevents lysis from
oc-curringprematurely (5). The lysis regulator is
athermallysensitive systemandrequires
con-tinuous protein synthesis to block lysis. Thus,
infected cellslyse ahead of schedule if the lysis regulator is disrupted by exposing the cells to elevated temperature(480C)or to
chloramphen-icol. Interestingly, these agents cause slow
lysis eveniftheAS gene is mutationally
defec-tive. Also, AS mutants can lyse a variety of
mutant host strains to some degree. These re-sultssuggestthatthereare twopotential
path-ways for triggering lysis. One requires the AS protein to be in an active state. The other is
independent of the S protein, but both are
in-hibitedby the lysisregulator.
Thebacterialmutationsthatmake cells per-missivehosts forAS- phagespresumably inac-tivate genesthatinfluence thelysis regulator. These mutations alsodecreasethe sensitivityof cellstocolicinK, but otherwise are diverse in
phenotype (16). We have suggested that the
Arex geneisalsoacomponentof thelysis regu-latorysystem (5). The Arex gene isnotable for being cotranscribed with the XcI repressor
gene in a Alysogen when the other phage genes
are repressed (7). Its precise fiction is still unknown, althoughrex gene expressionalters the timing oflysis by phage T1 growing inX lysogens(6). Inthe presentstudywe character-ized therexactivityand timing of hostcelllysis after theinductionofaseriesofmutantA lyso-gens to further understand the controls over
lysis.
MATERIALS AND METHODS
Strains. Thephage strains usedinthisstudyare listed in Table 1, together with the properties by which the various phage mutants were originally selected. All of the A phages carry the cI857 ind-allele. TheAach-4phage wasobtained by spotting about 109 cI857phage particles on a lawn of AN259 cells on LB agar (11) at pH 4.5 and isolating the phage from one of the several dozen resulting plaques. Superficially this mutant displays a nor-mal phenotype on wild-type host cells at neutral pH. Weshalldescribe its mutant properties at low pH elsewhere. TheXclo("clock")mutants wereobtained by selecting for mutants ofXcI857 phage with a shortened latent period (5).
Three strains ofE. coli K-12 were used as hosts for 626
on November 10, 2019 by guest
http://jvi.asm.org/
HOST CELL LYSIS BY PHAGE A 627
TABLE 1. Strains used
Strain Pertinent properties Source/reference
E.coli K-12
tolB Pleiotropic, multicolicin tolerance 1
tolA Pleiotropic,multicolicin tolerance 1
tolP Pleiotropic,multicolicin tolerance 1
cyaA Adenyl cyclase negative (GP1allele) 20
uncA Uncoupled oxidativephosphorylation 2
uncB Uncoupledoxidative phosphorylation 3
unc-405 Uncoupled oxidativephosphorylation 3
rep-5 Defective for certain phage and plasmidchromosome repli- 4
cation
mutU Increased mutation frequency 17
uvrD UVsensitive 17
Wild type Isogenic parent or sister strains for each above bacterial mutant
AN259 argH entA strA 5
QD5003 SuIII M. Howe
233 trpE Su- C.Yanofsky
Phage
XcI857 Thermally inducible Sussman and Jacob (18)
XcI857susS7 Lysisdefective Goldberg and Howe (8)
XcI857 ts9B Lysis defective Harris et al. (10)
XcI857rexQ Nonexclusion of phageT4rII Gussin and Peterson (9)
XcI857 rex5a Nonexclusion of phageT4rII Gussin and Peterson (9)
XcI857clo-2 Premature lysis Campbelland Rolfe (5)
XcI857clo-4 Prematurelysis Campbell and Rolfe (5)
XcI857 clo-9 Premature lysis Campbelland Rolfe (5)
XcI857 ach-4 Forms plaques at pH 4.5 This paper
T4+ Wild type C. Fuerst
T4rII rIIrapid lysis mutant C. Fuerst
aAll A phages also carried the ind- allele.
mostof the experimentsinthisstudy:AN259
argH-entA strAr,QD5003 SuIII+, and233trpE-Su-. The
othermutantbacterialstrains, which allowphage
XsusS7toplate(16),arereferredtobytheirrelevant
mutantgene.Theirprincipal phenotypic character-istics (most ofwhich are pleiotropic) are given in Table 1. Lysogens were constructed in a
conven-tionalmannerand tested for levels of Xrex activity
by comparing the plating efficiencies on them of
phagesT4+andT4rll(5).
Measurement of latent periods. Studies on the timing of host cell lysiswere carried out on
ther-mally induciblelysogens carryingthe
temperature-sensitive XcI857 allele of the repressor gene.
Log-phaseculturesweregrownat30'Cto anappropriate
optical density inLBbroth. Aerobiccultureswere
shaken rapidlyinsidearmflasks inanoscillating
waterbath. Anaerobic cultureswere sealed in cu-vettetubes (1by30cm)andkeptforatleast 1hat
300C to use up all oxygen before induction. The cultureswerethenshiftedto40'C,and theiroptical
densitieswere measured periodically in a Unicam SP600 spectrophotometer. Insomeexperiments po-tassiumcyanideorchloramphenicol (Sigma Chemi-cal Co.) was added to induced cultures to a final concentration of 5 mMor250
jtg/ml,
respectively.Media. The other microbial techniques andmedia used have beendescribed elsewhere (5, 15, 16),
ex-ceptfor theglucose minimalmedium,whichwasM9
(11) supplementedwith 0.5 mMtryptophan and25
mMglucose.
RESULTS
Effect of bacterial mutations on Arexgene
expression.The series ofE. colimutantslisted
in Table 2 have previously been shownto be permissive hosts for AS- phages and
presum-ablyarepartially defectiveintheirlysis
regu-lators(5).Each of thesemutants,togetherwith its isogenic sisteror parent strains, was lyso-genizedwithXcI857rex+phageand tested for its abilitytoplate T4rll phage.Wild-type lysogens completely exclude T4rIIphage growth. Three bacterial mutations, tolA-, tolB-, and tolP-, allowT4rIIphagetoplateontheX lysogensat
the sameefficiencyas onthenonlysogens. The
tol mutations cause a dysfunction ofthe cell
membrane,resulting inmulticolicintolerance, sensitivitytovariousdrugsanddetergents, ab-normal cell shapes,decreasedgrowthyield,and other effects (1; Campbell and Rolfe,
unpub-lisheddata). Six othermutationsdecrease but do not abolish T4rII exclusion. We conclude
VOL. 23, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
628 ROLFE AND CAMPBELL
TABLE 2. ExpressionofXrexactivity in AS-permissive hostsa
Plating efficiency of
Permissivehost phageT4rII Parentallysogens 10-4
cya- (X) 10-4
uncB (X) 10-3
unc-405 (X) 10-3
uncA- (X) 10-3
rep--5 (X) 10-2
mutU- () 10-2
uvrD- () 10-2
tolP- () 1.0
tolA (A) 1.0
tolB- (X) 1.0
aXisXcI857. Alllysogensplatedphage T4+ at an
efficiency of 1.0. "Parental lysogen" refers to the
corresponding isogenic control strain to each mu-tant.In all cases the behavior of these controls was comparable to one another and to our reference strainAN259.
that thegreatmajorityof, butnotall, bacterial
mutationsthatinterfere with the lysis
regula-tor also depress the expression ofrex activity againstT4rllphagegrowth.
Levels ofrex activity of various X phage
mutants.A seriesofXphagemutantssimilarly
weretested forabnormal rex expression when carried asprophagesintwotypesofhostcells (Table 3). In the wild-type host a Xrex+
pro-phage completely prevents T4rII phage from plating, and the two Xrex mutants cause no
exclusion, as expected. The Aclo phages are
intermediateincharacterasthoughthey
spec-ify
reduced levels ofrex activity. The Xach-4 prophage shows no deficiency in T4rII exclu-sion. To test the possibility that this mutantspecifiesanextradegree ofrexactivityinstead ofadeficit,weintroducedit intoatolB- host. A wild-typeAphageisinsufficientto cause exclu-sion inthisbackground. The Xach-4 prophage does excludeand therefore mustexpress a
de-greeofrexactivity above that of thewildtype.
Curiously, the Aclo mutants, which show
re-ducedrex exclusion onwild-typecells, also
ap-pear to havehyper-rexactivity inthetolB host.
Thus, rex gene expression apparently can be
modified by mutation in several directions.
Even mutantsthat appear to havesimilarrex
phenotypes in standard indicator strains may
show demonstrable differences in other cell
backgrounds.
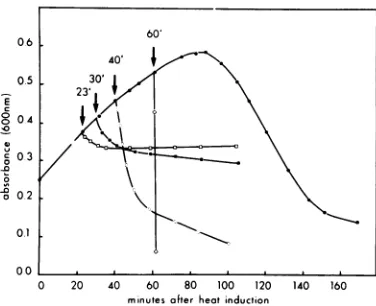
Latent periods of mutant A phages. A
changeinthe durationof thelatentperiod of an
inducedmutantlysogenis asensitive indicator
ofanalteredregulationof hostcelllysis. Figure
1A shows a comparison of the host cell lysis
patternofaArex-lysogenwithaArex+control
lysogen inducedin aerated broth. Cultures of the lysogensweregrown to earlylog
phase
at30°C and then shifted to40°C to thermally
in-duce theprophages. Theopticaldensities of the cultures continued to increase for a
period
of time after induction while phagereplication
proceeded. They then fell precipitously at 41 minasthecellslysedin
synchrony.
Clearlyanintact rex gene is notessential for thenormal
schedulingof hostcelllysis undernormal
aero-bic culture conditions. However, the
inactiva-tion of therex gene candelay lysisunder
var-iousspecializedgrowth conditions, suchas
an-aerobiosis (Fig. 1B) and in glucose minimal medium (Fig. 1C).
Figure 2 shows host celllysis curvesfor an
induced
Xclo-4
lysogen. Herelysisispremature under aerobicconditions anddelayed anaerobi-cally. As is the casefor Xrex mutants,altera-tions in the scheduled time of lysis are not
accompaniedbyasignificant lossinthedegree ofsynchrony of lysis.
Table4summarizesthelengths of latent
pe-riods for the various phagemutantsexamined. Both early anddelayed lysis occuramongthe phagesunder aerobic conditions, whereas all of the alterations observedinthe absenceof
oxy-gen are delays in lysis. In general, anaerobic
conditionsaremoresensitive thanaerobic
con-ditions fordemonstrating changes occurringin
the timingoflysis ofphage mutantswith
al-teredrex expression.
Hostcelllysisby AS- phages:phage AsusS7.
TABLE 3. ExclusionofphageT4rIIby various
mutantAprophagesa
Plating effi- rexalteration of Lysogen ciencyof phage mutant
pro-T4rII phageb
tol+(X) 10-4
tol+ (Xach-4) 10-4
tol+ (Aclo-2) 10-2
-tol+(Aclo-4) 10-2
-tol+(Aclo-9) 10-2
tol+ (ArexQ) 1.0
-tol+(Arex5a) 1.0
-tolB- (X) 1.0
toiB- (Aach-4) 10-4 ++
tolB- (Aclo-2) 10-4 ++
tolB- (Xclo-4) 10-4 ++
tolB
(Xclo-9)
10-4 ++tolB- (ArexQ) 1.0
tolB- (Arex5a) 1.0
aAlllysogens plated phageT4+atanefficiency of
1.0.
b_,
Indicates lesser exclusion by mutant pro-phage than by wild-type propro-phageinthat cellback-ground + + indicatesagreaterdegree of exclusion.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.57.250.64.229.2] [image:3.503.264.454.434.626.2]HOST CELL LYSIS BY PHAGE X 629
0.3
E
0
0 0
a,
u 0.2
-o
-o
a C01
0.1
0.0 minutes after heat induction
0 20 40 60 80 100 120
minutes after heat induction
0 10 20 30 40 50 60 70
minutesafter heat induction
FIG. 1. Host celllysisprofiles of induced Xrex+ and Xrex-lysogens.(A), Aerobic culturesin Lbroth; (B) anaerobiccultures inLbroth; (C)aerobic culturesinglucose-minimalmedium. Symbols: 0,233(XrexQ);
*,233(XcI857).
The XsusS7 phage hasasuppressiblemutation
inthe S gene. Itisunable to form plaqueson
Su- host cells, but will plate on the SullI+
strainQD5003 (8).
Even in thepermissive host,S geneactivity
isnot normal. Figure3 shows acomparison of
the host cell lysis patternsof XsusS7 and XS+
phages in strain QD5003. The XsusS7 phage
has a distinctly shortened latent period.
An-other indication that the S gene product
syn-thesized in QD5003 cells is not normal is its
inabilitytocauselysisat48TC(Fig. 4). Shifting
thetemperature ofwild-type lysogens to48TC
at 23 min after prophage induction does not
delay lysis. In fact, it stimulates lysis (5). In
contrast, shifting the temperature of induced
0.4
0.3
E
0
0
-o
u
o 0.2
-oE
0
-o
a
0.1
0.0
0.3
E 0.2
0 0 10
~0
° 0.1
0
0.0
L
VOL. 23, 1977on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.55.448.55.536.2]0.3L
0 0
0.2
-o
C
0
0.1
oco
0 10 20 30 40 50
0.5 c 0.4
C
0 0
w 0.3
0
-o
° 0.2
0.1
0.0
0 10 20 30 40 50 60 70
minutes afterheatinduction
FIG. 2. Host celllysis profile ofAN259 (Xclo-4) lysogen. (A) Aerobic cultures; (B) anaerobic
cul-tures. Symbols: 0, AN259 (AcI857); 0, AN259
(Aclo-4).
QD5003 (XsusS7) lysogenspreventslysis.
More-over, ifthe inhibited cellsarethenreturnedto
4000,they promptlylyse. Thus, high
tempera-ture converts the suppressed S7 protein to a
reversible inactivestate. Leavingthe cells for
anextendedlength oftimeat48°C resultsina
diminution of the completeness of lysis upon
returnto40°0. Thismayindicateaslow,
nonre-versible denaturation of theS7geneproduced,
or it may merely reflect a slighttemperature
sensitivity ofsomeotherstepinlysis.
Induced QD5003 (XsusS7) lysogens also
re-spondabnormallytochloramphenicol and
cya-nide. Addingchloramphenicol towild-type
ly-sogensat 23 minafterinductioncauses a
syn-chronous lysis 5 to 10 min later. In contrast,
chloramphenicol stimulates only a very
asyn-chronouslysisof theinducedQD5003 (XsusS7) lysogen (Fig.5).Approximately half of the cells lyse earlier than they would ifchloramphenicol hadnotbeen added, butasignificant fractionof
TABLE 4. Latentperiod of various Xphagemutantsa
Latentperiod (miAb Phage strain
Aerobic Anaerobic
A 41 45
Xclo-2 39 43
Xclo-4 34 52
xclo-9 37 47
Xach-4 41 50
XsusS7 300 NTc
Xts9B 125 NT
XrexQ 39 56
Xrex5a 40 48
aAisXcI857, and the hoststrain is either AN259 orAN233.
°Lengthoftimeat40'C foralog-phasecultureof thelysogentofalltoone-halfof its maximaloptical density after thermal induction of the prophage. Measurements areaccurate towithin 2min aerobi-callyand 5minanaerobically.
cNT, Not tested.
0.3
c 0.2
0 0
u
C
-0I
0
9A 0.1
0
0.0
0 10 20 30 40 50
[image:5.503.57.459.68.640.2]minutes after heat induction FIG. 3.Latentperiod of XsusS7 phage in a
per-missive SuIII+ host. Symbols: 0, induced QD5003
(AsusS7)lysogen;0,inducedQD5003 (AcI857) lyso-gen.
A
aerobic induction
/N
*
IN~~~~~~~~~
-.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.266.457.302.615.2]HOST CELL LYSIS BY PHAGE A 631
0.5
0.4
0.3
E
c
0 0 o4
:0.2 \ \ :\ 48
(20').
4002
0
48°(10').
40-0.1
548'(5_)
40400
0.0 I L
0 20 40 60 80 100
minutes after heat induction
FIG. 4. Temperature sensitivity of the
SuIII+-sup-pressedXsusS7 geneproduct. At 23 minafter
induc-tionat40°Ccultures ofQD5003 (XsusS7)lysogens
wereshiftedto48°Cfor theindicatedperiodsoftime before being shifted back to 40°C. Symbols: A, at
48°Cfor 5 minbeforebeing returnedto40°C; 0,at
48°Cfor10minbefore being returnedto40°C; 0,at
48°C for20 min before being returned to40°C; *,
controlleftat48°C;*,control40°Cculture.
the cells actuallyaredelayedinlysis by
expo-sure to it.Chloramphenicolalso is ineffectiveat
stimulatinglysisof these cellsat480C. Cyanidecausesimmediate lysisof
wild-type
lysogens when added at anytime after20min postinduction. It slightly hastens premature lysis of induced QD5003
(XsusS7)
lysogenslateinthelatentperiod (28min orlater),buttends toblock, more than stimulate, lysis whenadded
at 23min (Fig. 6).
Clearly, the SuIII+ suppressor does not
re-store anormalphenotypetotheXsusS7phage.
Itseems unlikely that the remaining defectis
simply a lower quantity of AS gene product synthesized (since lysis is premature and not delayed). Instead, the
suppressed
XsusS7 phage-infected cell seems to be deranged invarious aspects of the temporal control of S protein activity.
Hostcell lysis by AS- phages: phageAts9B. The phage Xts9B has a temperature-sensitive mutation in the S gene and will not form plaques at elevated temperatures (13, 14). When aninduced culture of a Xts9B lysogen is maintained at 400C, the latent period is ex-tended for at least several hours. If the temper-ature isreduced to 100C during this period, the cellslyse immediately.Remarkably, a shift to a low temperature even before the normal sched-ule time for lysis initiates an immediate asyn-chronous lysis (Fig. 7). If the temperature is dropped at 23 min after induction, it takes 17 minfor half of the cells to lyse. Lysis is more synchronousifthe temperature is dropped later in the latent period, and the cells lyse simul-taneously by 41 mm, the normal scheduled time of lysis by phage XcI857. Dropping the temperature ofaculture of aninduced control XcI857lysogendoes not cause lysis.
Adding cyanide to induced wild-type lyso-gensduringthesecond half of the latent period stimulates a lysis to occur prematurely, pre-sumably by activating the S protein ahead of schedule (13). Cyanide can also trigger the lysis of induced Xts9Blysogens even atanelevated, nonpermissive temperature (Fig. 7 and 8).This effectisseenonly ifcyanideisadded after the normally scheduled time for lysis (41 min). Adding cyanide earlier than this time does not havea significant effect, even ifthe
tempera-04
0.3 E
c
0
02
'.2
C
01
00
0 10 20 30 40 50 60 100 110 minutesofter heot induction
FIG. 5. Decreased sensitivity of inducedQD5003
(AsusS7) lysogens to premature lysis by
chloram-phenicol. Symbols: 0,after23 min at40°C, cultures
wereshiftedto48C; A,exposedtochloramphenicol
at40°C; 0, exposedtochloramphenicolat48°C;or
*,leftat40°Cas acontrol. VOL. 23, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.49.243.57.387.2] [image:6.503.254.447.417.601.2]632
0.25
30 0.20
0 0.15
0
-~0.10
0.05
0.0
0 10 20 30 40 50 60
minutes after heat induction
FIG. 6. Stimulation of lysis of induced QD5003 (XsusS7) lysogen by cyanide. A culture ofQD5003 (XsusS7)lysogenwasmaintainedat400C untillysis
wascomplete (a).Parallel cultureswereexposedto5
mM cyanide at 23 min (0) or 27 min (0) after inductionasindicated.
ture is droppedimmediately after itsaddition (Fig.9). Evenincubatingtheinducedlysogens
withcyanideat40'Cfor 30 mindoesnotaffect
the kinetics of lysis when the temperature is
subsequentlylowered (Fig. 9). Thus, theXts9B mutant displays a complex abnormal
pheno-type at a low temperature as well as athigh
temperature.Atalowtemperaturethe induced
lysogens are defective inthe mechanism that
normallydelays lysisuntil the scheduled time.
Also, cyanide willnotstimulate the lytic
proc-essata lowtemperature, although it still will
atahigh temperature.
Effects of certain bacterial mutations on
thephageXlatent period.The seriesof
bacte-rialmutationsthat interfere withXrex
expres-sion were tested for influences on the latent
period ofphage X (Table 5). Themoststriking
effectswere seenwithtwo unc-mutations
(un-coupled oxidation phosphorylation).
Aerobi-cally, the latent period isslightly shortenedby
uncA- and uncB- mutations. Anaerobically,
E
a)
E
0
>
a)
E 25 [
20 L
15
10
5
0
0 20 40 60 80 100 120
minutes after heat induction
FIG. 7. Time course of sensitivity of induced Xts9B lysogens to lysis by a low temperature and
cyanide.At thetimesindicatedafterprophage
induc-tion, cultures ofinduced AN259 (Xts9B) lysogens
were shifted from 40 to 15'C (U) or given 5 mM
potassium cyanide(0), and the length oftime for the
optical densitytofalltoone-half its maximal value is
indicatedbytheordinate.
60' 06
40'
05 23 4 E
0.4 ~0
0.2 0.1 00
0 20 40 60 80 100 120 140 160
[image:7.503.63.257.58.368.2]minutesafterheotinduction
FIG. 8. Lysis of induced Xts9Blysogensby
expo-suretocyanideat400C. Atthe indicated timesafter thermal induction of cultures ofAN259 (Xts9B) lysogens,potassiumcyanidewasaddedtoa concen-trationof5mMand thetemperaturewasmaintained at40TC. Symbols: control maintainedat400C ( 0); cyanideaddedat23 min (0), 30 min (*),40 min (0), and 60 min(c).
lysis isdelayedsomuchthat the latentperiod
cannotbemeaningfullymeasured.Incontrast,
a lack ofoxygen delays the lysis ofwild-type
lysogens by onlyafewminutes.
,a1
0f,
. . * .-8 *
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.262.457.73.293.2] [image:7.503.268.456.378.531.2]HOST CELL LYSIS BY PHAGE X 633
Mostof the other bacterial mutations tested also affect the duration of the latent period. Underaerobic conditions, some promote earlier lysis and others delay lysis. Under anaerobic conditions, only delays in lysis were observed. The rep--5 mutation is unique among those tested in that it affects rexexpression, but does not noticeably alter the latent period of A
0.6 0.5
* 60'
E 0.4
-0
0 0O.3
A. 0.2
-o a
I . .a
0 20 40 60 80
[image:8.503.51.237.77.365.2]minutes after heat induction
FIG. 9. Effect ofcyanideonlatentperiodofXts9B
phageat100C.A cultureofAN259 (Xts9B) lysogen
wasincubatedat400Cfor30 minanddividedinto
fourparts.One partwasshiftedto10C(*);asecond
wasexposedtocyanideandplacedat100C (C>),and
thethirdwasincubated withcyanideat409Cfor30 minand thenshifted to100C (0). The control was
maintainedat40"C (0).
03
20.2
*X
0
0.
phage. Thus, abnormal rex expression is usu-ally, but not always, associated with the abnor-mal scheduling of host cell lysis in mutant bac-teria.
Host cell lysis of tolB- cells by A phage mutants.The mutations in the tolB, -A,and -P genes are the most extreme onesat abolishing rex activity, sensitivity to colicin K, and the requirement for S protein for Aplating.Induced tolB- (X) lysogens also have anodd pattern of lysis. Lysis begins early andcontinues gradu-allyfor 100min.Two of theAmutantsthat are able to express rex activity in tolB- cells
(Xach-4 andXclo-2) partiallyovercomethe tolBeffect onhost cell lysis(Fig. 10).InducedtolB- (Xach-4)lysogensgive anessentially normallysis pro-file, whereas induced totB- (Xclo-2) lysogens give a compound lysis pattern. A slow asyn-chronouslysis begins earlyinthelatentperiod, similar to that occurringin theinduced toEl-(X) lysogens. This lytic process is then inter-rupted by a more synchronizedlysis of the
re-TABLE 5. Latentperiod ofphageXcI857in various mutanthost cells
Latentperiod (min) Lysogen
Aerobic Anaerobic
Wild type (x) 41 45
rep-5 (A) 41 42
mutU (A) 52 73
tolB (X) 65 77
uvrD (A) 54 82
uncA (X) 36 >1,000
uncBh
(_)
37 >1,000aXiskcI857.
0 10 20 30 40 50 60 70 80 90 100 110
minutes after heat induction
FIG. 10. Hostcelllysisprofiles ofvariousinduced tolB-lysogens. Symbols: *,tolB- (Xc1857); U,tolBE
(xclo-2); O,tolB-
(Xach-4);
0,tolBs
(XcI857).VOL. 23, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.503.75.447.170.626.2] [image:8.503.101.413.396.632.2]634 ROLFE AND CAMPBELL
maining cells as occurs in the tolB- (Xach-4)
cultures.
DISCUSSION
Previousphysiologicalstudies have indicated that host cell lysis is controlled by a double regulatory mechanism. The XS gene product is
apositive regulator that promotes the lysis of wild-type cells.Thelysis regulator is the second
componentwhose activity blocks lysis.
Phage-infected cellscanbetriggeredtolyseahead of schedule by blocking protein synthesis or by briefly exposing them to a high temperature. An interpretation of thesedataisthat thelysis regulator is a temperature-labile protein, whose continuous synthesis is necessary for blockingprematurelysis. Accordingly, phageX
should havetwopotentialavenuesfor
trigger-inglysis (activating the S system and blocking the action of the lysis regulator). By using var-ious phage mutants in a tolB- host, we con-firmed that induced lysogens indeed canshow
twotypesof control over lysis. These two
proc-esses aredesignatedlysis I and
lysis
II inTable6. LysisIoccurs ata genetically scheduled time andresultsinasynchronous lysis ofthe cells. It requiresafunctional S gene. Lysis II can occur
in acellat anytime after20 minfrom
induc-tion, is insensitivetothelysisItiming mecha-nism, and causes cells to lyse asynchronously. Presumably lysis II isbroughtaboutby abolish-ing the inhibition of the lysis regulator. This inhibition requires the tolB+ geneproduct and probablyto alesserextentthe normal activities ofthe various other bacterial genes, whose
mu-tational inactivation allows XS-phagemutants toformplaques(5).Lysis IIisindependent ofS. The two patterns of lysis control exhibit a degree ofindependencein thattheycan occur
individually ortogether in aculture of phage-infected cells. For example, wild-type phage lyse wild-type host cells exclusively under S
TABLE 6. Patternsof host celliysisexhibited by various mutantXlysogens
Occurrence of lysis type: Lysogen
I II
toiB- (Xclo-2) + +
tol+(XcI857) +
-tolB- (Xach-4) +
-tolB- (XcI857) - +
tolB- (XsusS7) - +
tol+(XsusS7) -
-tol+(Xts9B) -
-control andlysetolB- cellsexclusively by lysis II. tolB- cells infected byXclo-2 phage actually
are susceptibletoboth patterns oflysis. At 45 min after induction, both lytic processes are
actually occurring simultaneously among the remaining cellsinthe culture(Fig. 10). Despite this evidence ofindependence, thetwocontrol mechanismsareclosely relatedphysiologically. For example, the tolB mutation of the host
drastically affects both control mechanisms, and theach mutation ofXreverses both of these
effectssimultaneously. Also, most of the other
hostmutationsthat enhancelysis IIto alesser
extent also affect thetiming of lysis mediated
by the S system (Table 5). Thus, althoughthe
two systems can be dissected by abnormal
ge-netic orphysiological factors, theyprobablyare
integrated into a single control system in the wild-type-infected cell.
We have suggested on the basis of circum-stantialevidence that theXrexgeneproduct isa
component of the lysis regulator system. The data presented above reinforce this model by bearing out the following three predictions aboutrex geneactivity.
(i) If rex canaffect the timingoflysis, then
one should be able to isolate phage mutants
that lyse early, and some of these mutants
should have altered levels of rex expression. Selecting mutants with ashortened latent
pe-riod has provedtobestraightforward. We have thermallyinducedaculture of a XcI857 lysogen andcollected the free phage particles thatwere
present at 13 min before the normally sched-uled time oflysis (5).
Individual phageswerepickedasplaqueson
sensitive cell lawns and designated Xclo (for "clock" phage). Threeout often phages
exam-ined had an altered rex expression as a pro-phage. Two of these had altered latent periods
in awild-typebacterial host. The third hadan
alteredpattern of host cell lysis of tolB- cells. The other seven phages were normal in both their timing oflysis and rex expression. Pre-sumably, theyweremerely XcI857 phages that passed throughourselection procedures.
(ii) A variety of bacterial mutations allow
AS- mutant phage to form plaques. We have
previously ascribed their permissiveness to an
impaired expression of the lysis regulator (5).
Aspredicted, most of these mutations also af-fect thetimingofhost celllysis byXcI857phage
at least under some conditions and interfere
withthe exclusionofT4rIIphage plating bya
Xrex+prophage (Tables2and4). Thediversity
of these mutations strongly suggests that the
joint impairment ofrex andlysisregulator
ac-tivityismore thanacasualcoincidence.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
HOST CELL LYSIS BY PHAGE x 635
(iii) rex- X mutants have been isolated by
Gussinand Peterson (9) asprophagesunableto
interfere with the replication of T4rII phages. The latent period of one of these mutants, XrexQ, is shortened under aerobic conditions
andlengthened underanaerobic conditions and
inglucose-minimal medium(Fig. 1). The other
possibly shows a marginally delayed host cell
lysis anaerobically (Table 4).
Although there can belittle doubt that the
rex geneisintimately relatedto the lysis
regu-lator, the nature of this relationship remains
obscure. Most mutations, both bacterial and viral, that affect the timing ofhost cell lysis also affectrex activity, but the reciprocal
rela-tion is not as strong.It ispossibletomutatethe
rex gene to a variety of forms without grossly
affecting the length of the latent period. The Xach-4, Xclo-2, and XrexQ phagesall lysetheir hosts at the normal time under usual
condi-tions.
One complexity of the rex mutants is that
they appear to represent more diversity than
simply variationsinthe level ofrexgene
prod-uct or "rex activity." For example, the Xclo
mutationsdecreasethe level of T4rII exclusion
inwild-typeE.coli lysogens, butincrease it in
tolB- lysogens. If the rex gene is capable of mutatingtoqualitativelydifferent alleles,even
ttrex-negative" mutants (selectedascausing no
phageT4rll exclusion) mayhavesome form of
rex activity. In fact, the two Xrex- mutants
studied here,
XrexQ
and Xrex5a, showdiffer-ences in their latent periods. These mutants
presumably are isogenic, although they were
isolated after exposure to nitrosoguanidine, a
chemical thatfrequentlycausesmultiple
muta-tions (9).
The simplest role for rex consistent with these observations so far is that the rex gene
helpstoinsulate thelysisregulator from envi-ronmental or genetic factors that otherwise would disturb the timing oflysis. Under opti-mal growth conditions, Xrex activity is there-forecompletelydispensible.Thisview
rational-izestheinability of Gussin and Peterson(9) to
detect any effect of rex- mutations on the growth cycle of the X phage in theircarefully conductedstudy.
TheXclo mutantsshowthat thelengthof the
latentperiodisgenetically determined andcan
bealteredbymutations.Therefore, the
charac-teristic ofthe wild-type phage mustrepresent
anoptimalvaluesetbyadaptiveevolution. The
primary function ofthe rex gene might be to
ensurethat thisoptimal valueoccurs
irrespec-tive ofoxygen tension. Alternatively or
addi-tionally, a rexfunction may betopromote the
optimal latent periodacross an enlarged range
ofE. coli strainsoccurring in its native anaero-bic environment.
The complexity ofcontrol over host cell lysis
byXphage makesitimpossible at this point to
present more than a plausible model for the
control ofAS protein activity. The lytic
behav-ior of both S+ and S- X strains is compatible
with the following hypothesis. The S protein is
capable ofexisting intwo states, one active and
the otherinactive. During the latterhalfof the
latent period, Sprotein issynthesized and
accu-mulates in the inactive state due to the pres-enceof the lysis regulator. At41min an altera-tionin thephysiology of the cell either
inacti-vatesthelysis regulator ormakes theSprotein
insensitive to the presence of the lysis regula-tor. Poisoning the cell with cyanide can also
convertthe Sprotein totheactive state
prema-turely. The active S protein insults the cell membrane and allows theendolysintodestroy the cell wall.
The S protein of the Xts9B is altered in its
activation properties. A high temperature
causes it tobe more difficultto activate.
Nei-ther thepassageof41 min northeexposure to
cyanide alone will triggerinduced Xts9B
lyso-gens to lyse, althoughthe combination of the
twowilldo so (Fig. 8). A low temperature
fa-vors the active state. The S protein becomes
activeafterashorterspanoftimethannormal,
but no longer demonstrates activationby
cya-nide. Possibly, at alowtemperaturetheS-ts9B protein spontaneously occurs in the activated
state that the wild-type protein assumes in a
cellexposedtocyanide.
TheS-7 proteinsynthesized in anSuIII+ cell has a derangement in its activity similar to
that of the S-ts9B protein. It isreversibly
con-vertedto aninactive statebyanelevated
tem-perature, in which it cannot be activated by
agents that normally trigger the mechanism
(Fig.5). At alowtemperature itsactivityisless
sensitive torestraintby thelysisregulatorand
tostimulationbycyanide and chloramphenicol. These parallels between twoindependentAS
mutants strongly suggest to us alterations in
the conformational states ofan allosteric
pro-tein. Both theS-ts9B and the suppressedS-7
proteinsmight be described aspartially
"desen-sitized" (12) to agents that affect the
equilib-riumbetweenan activeandinactive state. It is
possiblysignificantthat theS7 andts9B
muta-tionsmapcloseto oneanotheratthedistal end
oftheSgene (13).
ACKNOWLEDGMENTS
Wethank PeterFokkerforhistechnicalassistance.
VOL. 23, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
636 CAMPBELL
This study wassupported by Public Health Service grant AI 13089from the National Institute of Allergy and Infec-tious Diseases.
LITERATURE CITED
1. Bernstein, A., B. Rolfe,andK. Onodera. 1972.
Pleio-tropic properties and genetic organization of the tolA,B locus of Escherichia coli K-12, J. Bacteriol. 112:74-83.
2. Buglin,J.D., G. B.Cox,and F.Gibson.1971.Oxidative
phosphorylationinEscherichiacoli. Mutations affect-ingmagnesium ion- or calcium ion-stimulated adeno-sinetriphosphatase.Biochem. J. 124:75-81. 3. Butlin, J.D., G.B.Cox, andF.Gibson. 1973.
Oxida-tivephosphorylationinEscherichia coli K12:the ge-netic and biochemical characterisation of a strain carrying a mutation in the uncB gene. Biochim.
Bio-phys.Acta292:366-375.
4. Calendar,R.,B.Lindquist, G.Sironi,andA.J. Clark. 1969. Characterization ofREP- mutants andtheir interaction withphage P2.Virology40:72-83. 5. Campbell,J.H.,and B. G.Rolfe. 1975. Evidence for a
dualcontrol of the initiation ofhost-celllysiscaused
by phage lambda. Mol. Gen. Genet. 139:1-8. 6. Christensen, J. R., and J. M. Geiman. 1973. Anew
effect of the rex gene ofphageX:prematurelysisafter infectionby phageT1.Virology56:285-290. 7. Dove, W. F. 1968. Thegenetics oflambdoid phages.
Annu.Rev.Genet.2:305-340.
8. Goldberg,A.R.,andM.Howe.1969.New mutations in theScistronofbacteriophage lambdaaffectinghost celllysis. Virology38:200-202.
9. Gussin, G. N., and V. Peterson. 1972. Isolationand properties of rex- mutants ofbacteriophage lambda.
J. Virol.10:760-765.
10. Harris, A. N., D. W. A. Mount, C. R. Fuerst, and L. Siminovitch. 1967. Mutations inbacteriophageX
af-fecting host cell lysis. Virology32:553-569.
11. Miller, J.H. 1972.Experimentsinmoleculargenetics. ColdSpringHarborLaboratory, ColdSpringHarbor,
N.Y.
12. Monod, J., J. P. Changeux, and F. Jacob. 1963. Allo-stericproteins andcellularcontrol systems. J.Mol.
Biol. 6:306-329.
13. Reader, R. W., and L.Siminovitch. 1971. Lysis defec-tivemutants of bacteriophage lambda: genetics and physiology ofS cistron mutants. Virology 43:607-622. 14. Reader, R. W., andL.Siminovitch. 1971. Lysis defec-tive mutants ofbacteriophage lambda: on the role of the Sfunction in lysis. Virology 43:623-637. 15. Rolfe, B., J. Schell,A. Becker, J.Heip,K. Onodera,
and E.Schell-Frederick.1973.Acolicin-tolerant mu-tantofEscherichiacoli with reduced levels of cyclic AMPand a strong bias towardsXlysogeny. Mol.Gen. Genet. 120:1-16.
16. Rolfe, B. G., and J. H. Campbell. 1974. Arelationship betweentolerance to colicin K and the mechanism of phage-induced host cell lysis. Mol. Gen. Genet. 133:293-297.
17. Siegel, E. C. 1973. Ultraviolet-sensitive mutator mutU4 ofEscherichia coli inviablewithpolA.J. Bac-teriol.113:161-166.
18. Sussman, R., and F. Jacob. 1962. Sur unsystemede
repression thermosensible chez le bacteriophage
d'Eschertchia coli. C.R. Acad. Sci. Paris
254:1517-1520.
19. Taylor, A. 1971. Endopeptidase activity of phage A-endolysin. Nature(London) New Biol. 234:144-145. 20. Yokota, T., and J. S. Gots. 1970. Requirement of
adeno-sine 3',5'-cyclicphosphate forflagellaformation in
Escherichia coli and Salmonella typhimurium. J. Bacteriol. 103:513-516.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/