0022-538X/88/062076-07$02.00/0
Copyright © 1988,American Society for Microbiology
Cell Lines
Containing
Varicella-Zoster
Virus
Open
Reading Frame
62 and
Expressing
the
"IE"175
Protein
Complement
ICP4
Mutants
of
Herpes Simplex
Virus
Type
1
JAMES M.
FELSER,1
PAUL R. KINCHINGTON,2 GENEVIEVEINCHAUSPE,1
STEPHEN E. STRAUS,'ANDJEFFREY M. OSTROVEl*
Medical Virology Section, Laboratory ofClinicalInvestigation, NationalInstitute of AllergyandInfectious Diseases,' and the
Unifrnrmed
ServicesUniversity of
the HealthSciences,2
Bethesda,Maryland
20892Received 7 October1987/Accepted22 February 1988
Vero cellswerecotransfectedwithpSV2neo andarecombinantplasmid containingthe varicella-zostervirus (VZV) open reading frame 62 (ORF62). Three neomycin-resistant cell lines were isolated and shown to complement two different ICP4 mutants (tsB21 and d120) ofherpes simplex virus (HSV) type 1 (HSV-1).
VZV-specific RNA could notbe detectedinthese celllines, butfollowinginfection with tsB21,a4.3-kilobase VZV transcript was detected. This RNA increased in quantity when cells were infected in the presence of cycloheximide.AVZV-specific proteinof 175 kilodaltonswasdetected in extracts of all three cell linesfollowing infection with wild-type HSV-1 butnotinuninfectedcells. That VZV RNA andproteinweredetectedonlyin HSV-1-infected cellssuggeststhatacomponent of the HSVvirion activates theexpressionof VZVORF62.The increase in RNA expressionseeninthepresenceofcycloheximideindicates that theproteinencodedbyVZV
ORF62, "IE"175, may beautoregulatory. These data provide further evidence that VZV "IE"175 is the functional analog of the HSV ICP4.
Expression of many herpesvirus genes is coordinately regulatedandsequentially orderedinacascade fashion. The mostextensive analyseshave been done forherpes simplex virus (HSV) type 1 (HSV-1) genes, which are divided into
five temporal groupings (ox,
2,
yl, and "Y2). Five cx orimmediate-early genes of HSV are known. These are
ex-pressed first ininfection and aretranscribed in the absence of de novo protein synthesis (20, 21). The HSV and y
classes of genes are expressed as early and late genes,
respectively.
Less extensiveanalyses ofthetemporalexpressionof the genesofotherherpesviruseshave been conducted. No such studies have been performed with varicella-zoster virus
(VZV), in part because of the difficulty in obtaining high
titers of cell-free virus requiredto synchronously infect the
large numbers of cells needed to study gene expression. Nevertheless, among the products of the 71 open reading frames (ORFs) predicted by theVZV DNAsequence, there are a number ofpolypeptide homologs between HSV and VZV (11). Fromthese, Davison and Scott (11) have
identi-fiedhomologsin VZVfor three HSVotgenes(ICP4, ICP22,
and ICP27) (11). The predicted product of VZV ORF62
sharessignificant amino acid homology with the HSV ICP4. Three regions of the predicted VZV protein, representing approximately650amino acids,haveover50% homologyto
ICP4(10).TheORF62 product is predictedtobeatleast 140
kilodaltons(kDa) in size (10, 11)compared with175 kDa for
the HSV-1 ICP4 (36). Recently, Shiraki and Hyman (29)
detected four immediate-early proteins of VZV. The one
theyestimate to be 185 kDa is closest in sizetothe protein
predicted forORF62. We reported previously that a DNA
segment containing VZV ORF62 complements an ICP4
temperature-sensitive (ts)mutant ofHSV-1, tsB21, thereby
mappingan ICP4-equivalent activity toORF62 (16). In this report, we describe the establishment of
trans-* Corresponding author.
formed cell lines which contain andexpresstheORF62gene
of VZV. These cell linesefficiently complementtwodifferent
ICP4 mutants of HSV-1. Under the conditions at which
ORF62isefficiently expressedin thesecells, we can detect and characterize its transcriptand a 175-kDaprotein prod-uct.
MATERIALS ANDMETHODS
Cells and viruses. Human embryonic fibroblasts (Flow 5000), human fetal foreskin fibroblasts (HFFs), and CV-1 and Vero cellswere propagatedina 1:1 mixtureof minimal essential medium and medium 199 supplemented with 10% heat-inactivated fetal bovine serum, 2mM glutamine, peni-cillin G (100U/ml), andstreptomycin (100 ,ug/ml). E5 cells,
agiftof NealDeLuca,containthegeneencodingthe HSV-1 ICP4 and are able to complement HSV-1 ICP4 deletion
mutants(30). TheKOS strain of HSV-1 andtsB21,an ICP4
temperature-sensitive mutant of HSV-1 (KOS), were
pro-vided by Priscilla A. Schaffer (14). HSV strain d120, also
courtesy of N. DeLuca,isamutantof KOS which contains a4.1-kilobase (kb) deletionin thegeneencoding ICP4 (13). VZV strains Ellenand Scottwere grown in Flow 5000 and HFFcells, respectively, aspreviously described (35).
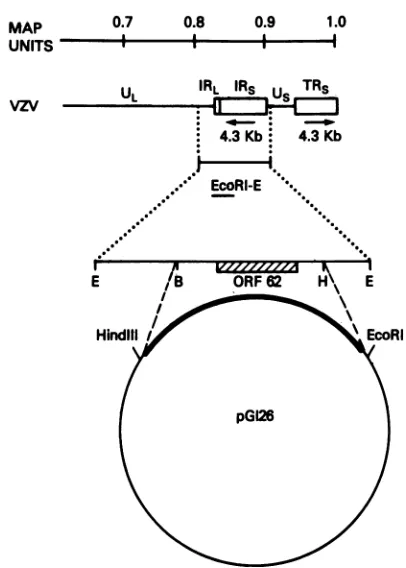
Plasmids. pVZVEcoE contains the EcoRI E fragment (Fig. 1)of VZV strainElleninpBR325. pVZVEcoA contains
the EcoRI A fragmentof VZV strain Ellen in pBR322 (9). pGI26 (Fig. 1)contains the 6.5-kbBglII-HpaI fragment from pVZVEcoA ligated into the SmaI site of pUC9 following blunting of ends with T4 DNA polymerase. Plasmid
pSV2neo contains the bacterial gene encoding G418 resis-tanceunder thetranscriptional controlofthesimianvirus 40
early promoter (32). pSG1, courtesy ofSandra K. Weller, containsthe EcoRIJKfragment, includingthe ICP4geneof
HSV-1 cloned into pBR325 (18).
Nucleicacidisolationandhybridization.CellularDNAwas
isolated and hybridized as described previously (28, 31). Total cellularRNA was isolated and purified by the
guani-2076
on November 10, 2019 by guest
http://jvi.asm.org/
MAP UNITS
vzv
0.7 0.8 0.9
a a a
1.0
UL IRL IRS U TRS
*
I
4.3Kb* 4.3KbE
FIG. 1. The mapposition of ORF62 is shown in the right-hand
portion of the VZVgenome. PlasmidpGI26 contains the indicated
BglII-HpaI6.5-kb fragment cloned into pUC9. Transcript mapping datahaslocalizeda4.3-kb transcripttoORF62asshown previously
(26). Thelocations of restriction sites forenzymesEcoRI (E),BglII
(B), and HpaI (H) arebasedonpublished sequence data (11). UL
andUs, Unique long and shortsegmentsof the virus;IRSandTRs,
internal and terminal repeats of the short segment; IRL, internal repeatof thelongsegment.
diniumthiocyanate-cesium chloride procedure and
hybrid-izedas previously described (7, 17, 26).
Complementationof tsB21byclonedDNAfragments. Com-plementation of tsB21 by protein expressed transiently from transfected DNA fragments was carried out by using a modificationof theprocedure of Sacksetal.(27).CV-1cells
were seeded into 60-mm-diameter dishes (6.25 x
105
cells per dish). The next day, the cells were transfected induplicate with0.5 ,ugofplasmidDNAand 10 jig ofcarrier DNA (salmon testes) in 0.5 ml of HEPES (N-2-hydrox-yethylpiperazine-N'-2-ethanesulfonic acid)-buffered saline. Four hours later, cells were treated with 15% glycerol in HEPES-buffered salinefor 2min.Thecellswereinfected 24 hlaterwithtsB21at39°C atamultiplicityof 2 PFUpercell. After incubation for 20 h at 39°C, the cultures were har-vested and virus titers were determined by the amount of
plaqueon Verocells at34°C.
Viral complementation studies. Virus titrations were per-formed on various cell lines as previously described (16).
Time course complementation studies were carried out by seeding the indicated cells into 60-mm-diameter dishes in
duplicate. The nextday, cells were infected with d120at a
multiplicity of 2 PFUpercell andwere maintainedat37°C.
Cultureswereharvestedatvarious times after infection.The yieldof d120wasdeterminedon E5 cells at37°C.
Biochemical transformation of Vero cells with plasmid
DNA. Vero cells were cotransfected with pSV2neo and
pGI26,andG418-resistant colonieswereselected andgrown
by using a modification of the technique described by
DeLuca etal. (13). Briefly, unlinearized pSV2neo (0.5 ,g)
and pGI26(20
R,g)
werecoprecipitated in the presence of 40,ugof salmon testes DNA in a total volume of 1.0 ml by the calcium phosphate procedure of Graham and van der Eb (19). Theprecipitated DNA was then added to subconfluent monolayers of Vero cellsin60-mm-diameter dishes. After a 4-h incubation at 37°C, the cells were shocked with 15% glycerol andincubated for an additional 2 days at37°C. The cells were thentrypsinized and seeded onto 85-mm-diameter
dishesatadensity of about 5 x 103cells percm2. After cell
attachment, the medium was removed and replaced with
medium containing 1 mg of active G418 per ml. After a
further 5 to 10 days, the G418 concentration was lowered to
600 ,ug/ml. At 12 to 18 days after transfection, individual
G418-resistant colonies wereisolated, amplified, and
subse-quently maintainedin the presence of G418 (600 ,ug/ml).
Western blot (immunoblot) analysis. Rabbit antisera were prepared against a synthetic peptide corresponding to the
C-terminal12amino acidspredictedfrom the sequence data
forVZVORF62(11; P. R. Kinchington, W. Ruyechan, and
J. Hay, manuscriptinpreparation). The peptide was
conju-gated to keyhole limpet hemocyanin and injected into
rab-bits, whereupon antisera were derived. A peptide
corre-spondingtotheC-terminal12aminoacids predictedforVZV
ORF29(22a), which has been identified as the VZV major
DNA-binding protein byhomology to HSV-1 (11), was also
synthesized.
A total of
106
cells were seeded into 60-mm-diameterdishes in duplicate and infected on thefollowing day with
HSV-1strain KOSat37°Cat amultiplicity of2PFU percell.
The infected cultures were harvested 20 h later and the proteins were solubilized in 300
RI
ofbuffercontaining 2%sodiumdodecyl sulfate, 5% 2-mercaptoethanol, 50 mMTris
hydrochloride, and5% glycerol (pH 6.7).
The proteins were separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and electrophoretically
transferredtonitrocelluloseinbuffercontaining
0.25
MTris,0.192 Mglycine, and 20% methanol. Following transfer,the
nitrocellulosewastreated with10% Carnation instant nonfat
milk inbinding buffer (1 mM EDTA, 10 mM Tris
hydrochlo-ride,0.15 MNaCl,0.02%Tween20[pH7.6]) for1h at25°C.
The
blots were then reacted with the anti-peptide seradiluted 1:50 in 1% milk-binding buffer for 2 h at 25°C.
Unbound antibodies were removedby five5-minwashes in
binding buffer, and the blots werereacted with
"251-labeled
staphylococcus protein A (New EnglandNuclearCorp.) at
105dpm/ml in 1% milk-binding buffer for2 hat25°C. Free
protein A was removed by extensive washing in
binding
buffer. Autoradiography was performed on Kodak XAR5
film.
RESULTS
ComplementationoftsB21bycloned DNAfragments.
Com-plementationassayswerecarriedout todeterminethe
ability
ofHSY-1 ICP4or its
putative
VZV counterparttocomple-menttsB21atthe
nonpermissive
temperatureof39°C.
Plas-mid
pSG1,
which contains the ICP4 gene of HSV-1, andpVZVEcoE and pGI26, which contain the VZV
ORF62,
were all found to
complement
tsB21 withcomplementation
indices of40.4, 11.3, and13,
respectively
(Table
1).Theonly
intact VZV ORF contained in pGI26 is
ORF62,
and thisplasmid was therefore usedto establish
stably
transformedcelllines.
Generation of cell lines
containing
ORF62.pGI26
andpSV2neowere cotransfected into Vero cells.G418-resistant
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.80.283.66.350.2]colonies were isolated and screened for their ability to support the replication of tsB21 at 39°C. Three cell lines, termed F19, F113, and F114, efficiently supported the repli-cation oftsB21. Twenty hoursafter infectionwithtsB21ata
multiplicity of 2 PFU percell, cultureswere harvested and
their virus yieldswerecomparedwith thoseobtained
follow-ing infection of VN2 cells at 39°C. VN2 cells are a
G418-resistant line derived from transfection of Vero cells with
pSV2neo alone. Cell lines F19, F113, and F114 yielded, respectively, 26, 39, and 153 times more virus than did
infected VN2 cells. The titers ofthe virus recovered from
cell lines F19, F113, and F114were alsodeterminedat 39°C
onVero cellstodetect recombinantprogeny.Some celllines
cotransformed with the ICP4 gene of HSV-1 have been
showntoyield recombinantprogenyfollowinginfection with
tsB21 (8). Such cell lines contain an HSV-1origin of repli-cation. Recombinantscreeningwasperfortned followingour
experiments, because aVZV replicationorigin iscontained in pGI26 (34). No increase above background titer was
observed, suggesting thatthe efficientproductionof
recom-binantprogeny did notoccur.
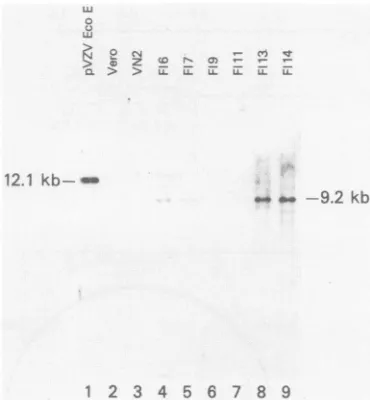
We examinedEcoRI-digested DNAfrom the transformed cell lines andVero cellDNAbySouthern blothybridization analysis by using the gel-purified VZV EcoRI Efragmnentas a probe (Fig. 2). Of four cell lines derived from the initial cotransfection ofpSV2neo and pGI26, the three cell lines
which complemented tsB21 (FI9, F113, and F114) were found to contain VZV DNA sequences. The major band
noted is 9.2 kb, the size of linearized pGI26. Since intact
pGI26 is cut only once with EcoRI in the polylinker se-quence(Fig. 1),thedetectionofasingle majorbandsuggests
integration of the plasmid intothecellsas atandemrepeatof pGI26 at one or more sites. The presence of a number of bands smaller and largerthan 9.2kbmay reflectintegration
of VZV sequences into different areas of the cellular
gen-ome.Theintegrationpatternisuniquefor eachcell line. One cell line (FIll) which did notcomplement tsB21 was nega-tive for VZV sequences. Two other cell lines(F16.and F17)
contained VZV sequences but have not been tested for
abilitytosupporttsB21 replication.VN2cellswerenegative forpGI26sequences, aswere Vero cells.
Cell lines containing ORF62 complement ICP4mutants of
HSV-1. Since initial screening demonstrated that three cell
lines (FI9, F113, andF114) supportthereplication of tsB21,
thecomplementing effectwasexaminedmorequantitatively.
Plaque formation by tsB21at39°Cwasseen onthethree cell
lines containing ORF62 with titers as shown in Table 2.
Plaque formation was also seen on the 14SV-1
[image:3.612.342.527.72.272.2]ICP4-con-taining E5 cell line. Incontrast, tsB21 did notform plaques
TABLE 1. Complementation of tsB21 by transfection withcloned DNA fragmentsa
Plasmid Insert (PFU/ms) Fold
pBR325 4.7 x 102 1.0
pSG1 Contains ICP4gene 1.9 x 104 40.4
ofHSV-1
pVZVEcoE EcoRI Efragment 5.3 x 103 11.3
of VZV
pGl26 Contains ORF62 of 6.1 x 103 13.0
VzV
aCV-1cellsweretransfected with 0.5 ,ug of each ofthe indicated plasmids and 24 hlaterwereinfected with tsB21at39°Catamultiplicityof 2 PFUper
cell.Afterincubation for 20 hat39°C,cultureswereharvested and titerswere
determined.
LL1
0
> 0
N
-12.1 kb--rn
Z
L L11 E LL_
.-*. ---.9.2 kb
l 2 3 4 5 6 7 8 9
FIG. 2. Southernblotanalysis of cellularDNA.
Electrophoreti-cally separated EcoRI digests (10 ,ug) of cellular DNAs were transferred to nitrocellulose and probed with a 32P-labeled gel-purified EcqRI-E.Lane1contains15 pgofpVZVEcoEplus 10jigof VerocellDNA. Lanes 2(Vero),3(VN2),and 7(FIll)werenegative bythisanalysis.VZV sequencesarefound inlanes 4(FI6), 5 (FI7), 8 (FI13), and 9 (FI14). The autoradiograph shown is a 10-day exposure. Asingleband of>9.2kbin sizewas seenwithF19DNA (lane6) with alongerexposure.
efficiently on Veto cells under these nonpermissive
condi-tions.
Asimilarexperimentwascarried out withd120, amutant
of HSV-1 which containsa4.1-kb deletion inthe ICP4 gene (Table 2). At 37°C, d120 produced no plaques on Vero or
VN2cells butyielded manyplaqueson E5, F114, F113, and F19cells, with E5 cells providingthe most efficient support. Time course studies were conducted to demonstrate the relationship between the replication of the HSV-1 ICP4
mutants and time after infection. Cells were infected with
d120 at amultiplicity of2 PFU percell and maintained at
37°C. Ctiltures were harvested at the indicated times, and
titers weredetermined on E5 cells at 37°C (Fig. 3). The E5 cell lineefficiently complemented this virus, exhibiting fold increases(relativetoVN2cells) of2.6 x 104at24h and 2.6
x 105 at 48h, similarto what has been reported earlier (30). FoldincreasesforF114cellswere 458 at 24 h and 2.3 x 104 at48 h.Althoughthedegree of complementationwasless for the VZV-derivedcell lines than thatforE5
cells,
significantTABLE 2. Titer of HSV-1 ICP4mutants on various celllinesa
Cellline tsB21titer (PFU/ml) d120titer (PFU/ml)
Vero 2.0 x 102 <101
E5 1.7 x 108 4.0 x 107
F114 1.0x 108 1.2 x 106
F113 9.8 x 107 4.0x 105
F19 3.6 x 107 2.7 x 104
VN2 NDb <lo,
a Each of the indicated cell lineswas seeded in duplicate into
60-mm-diameter dishesand infected withserialdilutions oftsB21 ord120.Following virusadsorptionfor 1 h, the cells were overlaid with maintenance medium
supplementedwith0.2%humanimmuneserumglobulin (6) and incubated for 7daysat39°Cfor tsB21 or at37°Cford120,after which the cells were fixed and stained with 0.1%crystalviolet.
bND. Not determined.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.317.557.595.671.2] [image:3.612.60.298.600.691.2]imideperml, the only HSV proteins present in the infected cells are the structural proteins of the infecting virions,
- E5
suggesting
that a virionprotein
of HSV stimulates theexpression of VZV ORF62.
Western blots of cellscontainingORF62. Cell lysates from _0 F114 F114, F113, and F19 cells were examined by Western blot 6- Fl13 analyses and probed with antibody raised in rabbits against a
peptide synthesized according to the predicted C-terminal
/1_0
F19 amino acid sequence of ORF62. In the absence of infection,the cell lines contained no detectable VZV polypeptides. However, infection with HSV-1 strain KOS (Fig. 5A) or tsB21(data not shown) led to production of proteins reactive with the ORF62 antibody. The predominant reactive moiety detected in the infected cells was 175 kDa in size. This protein was not seen in KOS-infected or uninfected VN2 cells butwas seenin lysates from VZV-infected HFF cells. When ORF62 antibody was preincubated with the ORF62 synthetic peptide (Fig. 5B), the 175-kDa protein was not soVN2
detected,
indicating
that the Western blotisspecific
forthegeneproduct ofORF62. Wecall this product VZV "IE"175
because of its apparent molecular weight and functional homology to HSV immediate-early gene ICP4. As in the Northern hybridization analyses, we were unable to detect , | , ffi , VZVprotein in the absence of HSV infection, again
suggest-20 30 40 5 6 70 ing that a component of the HSV virion activates the 4nIRD ATFR INFECTION expression ofVZV "IE 175.
Mvvno Pr En111rr-%1I RvUM
FIG. 3. The growth of d120 in cell lines F19, F113, and F114, containingORF62, and E5 cells compared with its growth in VN2 cellsat37°Covertime.
complementation did occur. The VN2 cell line does not
contain VZV sequences and served as the nonpermissive
line. Complementation of tsB21overtime was also studied
with lines F19, F113, and F114. FI14 cells produced the
greatest degree of complementation, exhibiting fold
in-creases(relativetoVN2cells) of 137at24 h, 78at48h,and
292at 72h (datanot shown). These results indicate that the
VZV protein present in these infections is able to
comple-ment two different ICP4mutants of HSV-1.
Progeny viruses recovered from d120-infected E5, VN2, and F114 cells at 24 h were screened for the presence of recombinantprogeny. Undiluted culture materialwasplated
on Verocells at 37°C. Noplaques were observed.
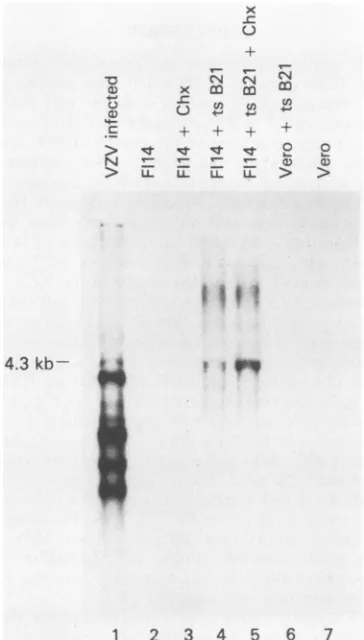
Transcription in cells containing ORF62. Transcripts that
havebeenmappedtotheshortrepeatregionof VZVinclude
a 4.3-kb RNAcorresponding to ORF62 (26). We sought to detect a similar transcript in the transformed cell lines
containing ORF62. Total cellular RNA was isolated from
F114, F113, and F19 cell lines and Northern(RNA) hybrid-ization analyseswere performed by using32P-labeledpGI26
and pVZVEcoA as probes. VZV transcripts were not de-tected when 20,ugof RNAwere probedunderthese condi-tions.However,a4.3-kbtranscriptcould be demonstratedin F114cells following infectionwithtsB21at39°C (Fig. 4).The 4.3-kb transcript was present in increased amounts when infection withtsB21wascarriedoutin thepresenceof 50jig of cycloheximide per ml. VZV-specific RNA was neither detected in uninfected F114 cells treated with cyclohexi-mide, nor wasit seenin uninfected VerocellsorVerocells infected withtsB21.
Although tsB21 doesnotreplicate efficientlyin Vero cells at 39°C, immediate-early proteins are synthesized in these
cells(12).None oftheseimmediate-earlyproteinsis synthe-sized- in the presence of cycloheximide. Therefore, when tsB21 infectionoccurs inthepresenceof 50,ugof
cyclohex--0 C.,
a)
N
x
___ U
-4-CN Ce4
x m m
U
-4-'
LL 11 LL IL
cs
CN
n
4-2
0
I .
4.3
kb-1 2 3 4 5 6 7
FIG. 4. Northern blotanalysesof total cellular RNA. Eachlane contains 2 ,ug of cellular RNA. The blot was hybridized with
32P-labeled pVZVEcoA. Chx indicates that cycloheximide was
addedtothe culture. An18-hautoradiographicexposureisshown.
107
106
1o5
iL
o4
t 10
103
2
101
1oo
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.61.300.65.339.2] [image:4.612.351.533.365.685.2]LI 5LIn )-" J:
>- v -= L
I ii: - t:-. .. L.L t.> E > >
z.!
IC 'Id No
r Z Z
175kDa ... 4U
.0
I r
A
B
FIG. 5. Westernblot anrlysesof celllysates.The celllysatesinpanelsAand BwerereactedwithantibodytoasyntheticORF62peptide. The antibody usedforpanel Awaspreincubatedwith 100 p.gofsyntheticORF29peptideperml,and that used forpanelBwaspreincubated
with100,ug of synthetic ORF62 peptideperml. In bothpanels,thelanes contain thefollowingcelllysates:lane1,VZV-infected HFFcells; lane 2, mock-infected HFFs;lane3,KOS-infected F113cells;lane4,mock-infectedFI13cells;lanes5and6,KOS-infectedandmock-infected F114 cells, respectively; lanes 7 and 8, KOS-infected and mock-infected F19 cells, respectively; lanes 9 and 10, KOS-infected and
mock-infectedVN2cells,respectively.A2-dayexposureis shown. A 175-kDamoietyisseeninlysatesofKOS-infected F19 cellson a7-day
exposure.
DISCUSSION
In this report, we describe the complementation of two
different ICP4 mutants of HSV-1 by the product of VZV
ORF62. Plasmid pGI26, which contains this VZV gene, complements tsB21at39°C.Transformed celllines
contain-ingthisgenealso complementthis mutantat39°C.These cell lines also complement the HSV-1 ICP4 deletion mutant
d120, indicatingthat theproductofORF62cansubstitute for the HSVICP4 whennoICP4moietyispresent. Thedegree
of complementation exhibited by the cell lines was even greater than that seen following transfection of DNA
frag-ments (16) and provides further evidence for locating the VZVequivalent of ICP4to this regionof the VZVgenome.
Although a VZV-specific transcript was not detected in
uninfected cells containing ORF62 (FI14 cells), a 4.3-kb VZV transcript was seen following infection with tsB21. This transcript was also present in cells infected in the presence ofcycloheximide, indicating that an HSV virion protein activates the expression of this VZV gene.
More-over,we areabletodetect VZVproteinonlyintransformed
cells that are jnfected with HSV. This also suggests that a componentof the HSV virionactivatestheexpression of the incorporated VZV gene. Thus, since thisgene product can substitutefor theHSVimmediate-earlygeneICP4 and is 175
kDa, we have called it VZV "IE"175. Furthermore, al-thoughShirakiand Hyman (29) detected fourVZV immedi-ate-early proteins in their studies, our finding ofonly one
major protein moiety is commensurate with the fact that
pGI26 contains onlyone complete ORF.
Themost likely virion protein responsible for the
stimu-latoryeffect is the at-TIF (a-trans-inducing factor, Vmw65)
which has been localizedtothe HSVtegumentand which is knownto induce, in trans, the expression of HSV agenes,
including ICP4 (1, 4). Two upstream DNA elements are
thoughttobeinvolved in theresponseofthe HSV-1 ICP4to
o-TIF(3). One of these is the TAATGARATTC consensus
sequence (R = purine) which, if present, is sufficient for
transcriptional activation(3, 23, 37).A second is aGA-rich
motif, GCGGAAG, which isthought toactivate TAATGA
RATTC-like motifs with apoorconsensus(3).
We analyzed the DNA sequence upstream of VZV
ORF62, assigning the AUG of this ORF a value of +1.
[image:5.612.138.472.73.287.2]Scanning 1,000 bases upstream of the AUG, we noted a
TABLE 3. Regulatory elements upstream from thepredictedVZVORF62
Orien-Regulatoryelement Position" VZVsequence tationb
TATAbox -374to-386 TATATTATATAT +
CCATT box -396 to -400 CCAAT +
-468 to -472 CAAAT -521to-526 GCAATC
GAbox -514to-522 GCGCGAACG +
TAATGARATTC -586to -597 TTATGGGTTAT + -778to-787 TTATGGTTAT + Splbinding sites -568 to-573 GGCGGC
-630to -635 GGCGGG +
-647to-652 GGCGCG
-657to-662 GGCGGG +
-674to-679 GGCGCG
-684to-689 GGCGGG +
-701to-706 GGCGCG
-711to -716 GGCGGG +
-728to -733 GGCGCG
-738to -743 GGCGGG +
-755to-760 GGCGCG
-765to-770 GGCGGG +
"Relative
totheORF62
AUG,which isassignedapositionof +1.bTheorientation (+or-)isgiven withreference to the 5' to3'directionof
theassociated mRNA. R representsapurine.Sources of consensus elements
aregivenintext.
"I".
-L. I-Ir L.
.-. -.L. 6.-.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.314.556.493.694.2]number ofregulatory elements, includingsequences sharing
homology
to both TAATGARATTC and GA-box motifs(Table 3).
AcandidateTATAbox is also shown inTable 3,asare three
possible
CCAATboxes (15). Several putativebinding
sitesforSpl (22, 25) werealsofound.Although the5'terminus of thetranscript associated withORF62 has not
yetbeen
mapped,
it is reasonabletoplace its locationwithinthis
region.
Our analysis therefore indicates that there areregulatory
elements upstream ofORF62 that are similartothose found upstream of the HSV ICP4 gene and which
could
respond
to a-TIF-like activities. Two genes adjacenttothe HSV-1a-TIF(ORFAandORFB) modulate its activity
incotransfectionassays(24). Recent datasuggest that VZV
may encode a related protein, in that its ORF10 shares
significant predicted
amino acid homology with a-TIF (11,24). Furthermore,
VZV ORF11 and ORF12 adjacent toORF10
share amino acid homology and identical orderingalong
thegenome with HSV ORFAandORFB(11, 24).Therehave
recently
been otherreports inwhichcelllinesthat
constitutively
containimmediate-early
genes have beenutilized in studies ofregulation. Boom et al. (2) described
preparation
ofa ratcell linecontainingthemajorimmediate-early
gene of human cytomegalovirus. They, too, wereunable to find a
transcript corresponding
to this gene byNorthern
blotting.
However, treatment of their cells withcycloheximide
for variousamountsof time ledtoexpression
ofmRNAof the
expected
size. Theinterpretation
ofBoomet al.
(2)
was thatcycloheximide
treatment removes thenegative autoregulatory
effect ofthe humancytomegalovirusmajor immediate-early protein
by preventing its synthesis.The dataof
Stenberg
and Stinksi(33) alsoindicate that thisprotein
has anautoregulatory
role. In our studies, atran-script
was not seen in F114 cells in the presence ofcyclo-heximide; however,
infection ofthese cells in the presenceof
cycloheximide
led to an increase in the amount oftran-script compared
with infection in theabsence ofdrug. Oneexplanation
for this is that VZV "IE"175 inhibits its owntranscription
aswell.SmithandSchaffer
(30)
describetheuse of G418-resistantcells
containing
the ICP4 gene of HSV-2 tocomplement
d120.
Additionally,
E5 cells(containing
theICP4ofHSV-1)wereableto
complement host-range
mutantsof HSVtype2,thus
showing
that ICP4products
encodedby
HSV-1 andHSVtype 2are
functionally interchangeable during
produc-tive infection. In
comparison,
ourcell lines which expressVZV "IE"175 arealsoableto
complement
d120andtsB21.It is of note that our three cell lines
complement
thesemutants to different
degrees
and that these resultsparallel
the relative amounts of the 175-kDa
protein produced
oninfectionwithHSV. One
explanation
fortheseresults is thatthe three lines differ in the percentage of cells that contain
ORF62 orinthe percentage of cells competent for infection
with HSV. An alternative
hypothesis
is that allcellscontainORF62 andareinfectioncompetent but their
ability
tomakeVZV
protein
differs.The lattertheory
wouldimply
that VZV"IE"175is
required
instoichiometric,
rather thancatalytic,
amountsfor
replication
to occur.In summary, we offer evidence that the gene for VZV
"IE"175is
responsive
toavirionprotein
ofHSV-1,
presum-ably
a-TIF.Sequence
elementsappropriate
to such aninteractionarepresent upstream of the gene. VZV sequence
data alsoindicate the presenceof
homologs
of HSV-1 a-TIF(ORF10)
and its modulators(ORF11
andORF12)
(11, 24).
This is in contrast to the situation for
pseudorabies virus,
another
a-herpesvirus;
itsmajor
immediate-early
gene isa-TIF
responsive,
and the upstream sequence ofthis genecontains elements of theTAATGARATTCmotif;however, pseudorabies virus itselfhas been reported to not have an
a-TIF homolog (5). We also provide evidence that VZV
"IE"175 inhibits its own transcription, an additional prop-erty that it shares with its HSVanalogICP4.
ACKNOWLEDGMENTS
WethankKenneth D. Croen for helpful suggestions and HollyA. Smith and WilliamC. Reinhold for technicalassistance.
P.R.K.wassupported byaPublicHealthService grantAl 18449 from the NationalInstitute ofAllergy and Infectious Diseases.
LITERATURE CITED
1. Batterson, W., and B. Roizman. 1983. Characterization ofthe herpes simplex virion-associated factor responsible for the induction ofa genes. J.Virol.46:371-377.
2. Boom, R., J. L. Geelen, C. J. Sol, A. K. Raap, R. P. Minnaar, B. P.Klaver, and J. van der Noordaa. 1986. Establishment of a ratcell line inducible forthe expression of human cytomegalo-virus immediate-early gene products by protein synthesis inhi-bition.J. Virol. 58:851-859.
3. Bzik, D. J., and C. M. Preston. 1986. Analysis of DNA se-quenceswhich regulate the transcription of herpes simplex virus immediate early gene 3: DNA sequences required for enhancer-like activity andresponse totrans-activation by a virion poly-peptide. Nucleic Acids Res. 14:929-943.
4. Campbell,M. E., J. W. Palfreyman, and C. M. Preston. 1984. Identification of herpes simplex virus DNA sequences which encodeatrans-acting polypeptide responsibleforstimulationof immediate earlytranscription. J. Mol.Biol. 180:1-19.
5. Campbell,M. E.M., and C. M. Preston. 1987. DNA sequences which regulate the expression of the pseudorabies virus major immediateearlygene. Virology157:307-316.
6. Campione-Piccardo, J., W. E. Rawls, and S. Bacchetti. 1979. Selectiveassayfor herpes simplex virusesexpressing thymidine kinase.J.Virol. 31:281-287.
7. Chirgwin, J. M., A. E.Przybyla, R. J. MacDonald, and W. J. Rutter. 1979. Isolation of biologically active ribonucleic acid fromsources enriched in ribonuclease. Biochemistry 18:5294-5299.
8. Davidson, I., and N. D.Stow. 1985.Expression ofan immediate early polypeptide and activation of a viral origin of DNA replication in cells containingafragment of herpes simplex virus DNA.Virology141:77-88.
9. Davison,A. J., and J. E. Scott. 1983. Molecularcloning of the varicella-zoster virusgenome and derivation of six restriction nucleasemaps. J.Gen. Virol. 64:1811-1814.
10. Davison, A. J., and J. E. Scott. 1985. DNA sequence ofthe majorinverted repeat in thevaricella-zoster virus genome. J. Gen. Virol. 66:207-220.
11. Davison, A. J., and J. E. Scott. 1986. The complete DNA sequenceofvaricella-zoster virus.J.Gen. Virol.67:1759-1816. 12. DeLuca, N. A., M. A. Courtney, and P. A. Schaffer. 1984. Temperature-sensitive mutantsin herpes simplex virus type 1 ICP4permissiveforearlygeneexpression.J. Virol. 52:767-776. 13. DeLuca, N. A., A. M. McCarthy, and P. A. Schaffer. 1985. Isolation and characterization of deletion mutants of herpes simplex virus type 1 in the gene encoding immediate-early regulatory protein ICP4.J. Virol.56:558-570.
14. Dixon, R. A. F., and P. A. Schaffer. 1980. Fine-structure mapping and functionalanalysis oftemperature-sensitive mu-tants in the gene encoding the herpes simplex virus type 1 immediateearlyproteinVP175. J.Virol. 36:189-203.
15. Efstratiadis, A., J.W.Posakony,T.Maniatis,R. M. Lawn,C. O'Connell, R. A. Spritz, J. K. DeRiel, B. G. Forget, S. M. Weissman, J. L. Slightom, A. E. Blechl, 0. Smithies, F. E. Baralle, C. C. Shoulders, and N. J. Proudfoot. 1980. The structureandevolutionof the humanPglobingenefamily.Cell 21:653-668.
16. Felser,J. M.,S. E.Straus,andJ.M. Ostrove. 1987. Varicella-zosterviruscomplements herpes simplexvirustype 1 temper-ature-sensitivemutants.J. Virol.61:225-228.
on November 10, 2019 by guest
http://jvi.asm.org/
17. Glisin, V., R.Crkvenjakov, and C. Byus. 1974. Ribonucleic acid isolated by cesium chloride centrifugation. Biochemistry 13: 2633-2637.
18. Goldin, A. L., R. M. Sandri-Goldin, M. Levine, and J. C. Glorioso. 1981. Cloning of herpes simplex virus type 1 se-quencesrepresenting the whole genome. J. Virol. 38:50-58. 19. Graham, F. L., and A. J. van der Eb. 1973. A newtechnique for
the assayofinfectivity of human adenovirus 5 DNA. Virology 52:456-467.
20. Honess, R. W., and B. Roizman. 1974.Regulationofherpesvirus macromolecularsynthesis. I.Cascaderegulation ofthe synthe-sis ofthreegroupsofviralproteins. J.Virol. 14:8-19. 21. Honess, R. W., and B. Roizman. 1975.Regulationof herpesvirus
macromolecularsynthesis: sequentialtransition ofpolypeptide synthesis requires functional polypeptides. Proc. Natl. Acad. Sci. USA72:1276-1280.
22. Jones,K. A., K. R. Yamamoto, and R. Tjian. 1985. Two distinct transcription factors bind to the HSV thymidine kinase pro-moterin vitro. Cell 42:559-572.
22a.Kinchington, P. R., G. Inchauspe, J. H. Subak-Sharpe, F. Robey, J. Hay, and W. T. Ruyechan. 1988. Identification and characterizationof avaricella-zoster virusDNA-binding protein by using antisera directed against a predicted synthetic oligo-peptide. J.Virol. 62:802-809.
23. Mackem, S., and B. Roizman. 1982. Structural features of the herpes simplex virus a gene 4, 0, and 27 promoter-regulatory sequences which confer at regulation on chimeric thymidine kinasegenes. J. Virol. 44:939-949.
24. McKnight, J. L. C., P. E. Pellett, F. J. Jenkins, and B. Roizman. 1987.Characterization and nucleotide sequence oftwoherpes simplex virus 1 genes whose products modulate at-trans-in-ducingfactor-dependent activation ofagenes.J.Virol. 61:992-1001.
25. McKnight, S. L., R. C. Kingsbury, A. Spence, and M. Smith. 1984.The distaltranscriptionsignals oftheherpes virus tk gene showacommonhexanucleotidecontrol sequence.Cell 37:253-262.
26. Ostrove, J. M., W. Reinhold, C.-M. Fan, S. Zorn, J. Hay, and
S. E.Straus. 1985.Transcription mappingof thevaricella-zoster virus genome. J. Virol. 56:600-606.
27. Sacks,W.R.,C.C.Greene,D. P.Aschman,andP. A.Schaffer. 1985. Herpes simplexvirus type 1 ICP27 isanessential regula-toryprotein.J. Virol. 55:796-805.
28. Sawyer, M.H., J. M.Ostrove, J. M.Felser, and S. E. Straus. 1986. Mapping of the varicella zoster virus deoxypyrimidine kinase gene and preliminary identification of its transcript. Virology 149:1-9.
29. Shiraki, K., and R. W. Hyman. 1987. The immediate early proteinsofvaricella-zoster virus.Virology 156:423-426. 30. Smith, C. A., and P. A. Schaffer. 1987. Mutants defective in
herpes simplex virus type 2 ICP4: isolation and preliminary characterization. J. Virol. 61:1092-1097.
31. Southern, E. M. 1975. Detection ofspecific sequences among DNAfragments separated by gel electrophoresis. J. Mol. Biol. 98:503-517.
32. Southern,P.J., and P. Berg. 1982. Transformation of mamma-lian cells to antibiotic resistance with a bacterial gene under control of theSV40early region promoter. J. Mol.Appl. Genet. 1:327-341.
33. Stenberg, R. M., and M. F. Stinski. 1985.Autoregulationof the humancytomegalovirus major immediate-early gene. J. Virol. 56:676-682.
34. Stow, N. D., and A. J. Davison. 1986. Identification of a
varicella-zostervirusorigin of DNA replication and its activa-tionby herpes simplex virus type 1 gene products. J. Gen. Virol. 67:1617-1623.
35. Straus, S. E., H. S. Aulakh, W. T. Ruyechan, J. Hay, T. A. Casey, G. F. Vande Woude, J. Owens, and H. A. Smith. 1981. Structure ofvaricella-zoster virus DNA. J. Virol. 40:516-525. 36. Watson, R. J., C. M. Preston, and J. B. Clements. 1979.
Separation and characterization ofherpes simplex virus type 1 immediate-early mRNA's. J.Virol.31:42-52.
37. Whitton, J. L., and J. B. Clements. 1984.Replication originsand asequenceinvolved in coordinateinduction ofthe immediate-earlygenefamilyareconserved in anintergenicregion of herpes
simplex virus. Nucleic Acids Res. 12:2061-2079.