0022-538X/92/010405-09$02.00/0
Copyright
C 1992, AmericanSociety
forMicrobiologyA
Glutamine
Residue in the
Membrane-Associating
Domain of the
Bovine
Papillomavirus
Type 1
E5 Oncoprotein
Mediates Its
Binding
to
a
Transmembrane Component
of the
Vacuolar
H+-ATPase
DAVIDJ. GOLDSTEIN,1REINHARD KULKE,2 DANIEL DIMAIO,2 ANDRICHARD
SCHLEGEL'*
Departmentof Pathology, Georgetown University Medical School, 3900 Reservoir Road NW, Washington,D.C. 20007,1
and
Department of
Genetics,
YaleUniversity
Schoolof
Medicine,
NewHaven,
Connecticut065102
Received12September1991/Accepted9October 1991
The44-amino-acidE5oncoprotein is the major transforming protein of bovine papillomavirus type 1. It is a highly hydrophobic polypeptide which dimerizes and localizes to the Golgi apparatus and endoplasmic
reticulum membranes. Recent evidence suggests thatE5modulates thephosphorylationandinternalizationof the epidermal growth factor and colony-stimulating factor 1 receptors and constitutively activates platelet-derived growth factor receptors in C127 and FR3T3cells. Although no direct interaction with these growth factor receptors has yet been identified, the E5 oncoprotein has been shown recently to interact with the hydrophobic 16-kDa component of the vacuolarH+-ATPase(16Kprotein) [D. J.Goldstein,M. E.Finbow, T. Andresson, P. McLean, K. Smith, V. Bubb, and R. Schlegel, Nature (London) 352:347-349, 1991]. In the currentstudy, we have furtheranalyzedtheE5-16K protein complex byfast protein liquidchromatography
and shown that eachE5 dimerappears to bind two 16K proteins. In order to define the specific amino acid residues of E5 whichparticipateinthis binding, mutated E5 epitopefusionproteinswere analyzed fortheir ability tocoprecipitate 16K protein. Transformation-defective mutants containing amino acid substitutions withintheshorthydrophilic carboxyl-terminaldomain retained theabilitytoassociatewiththe 16K protein. However, E5 mutantslacking the glutamine residue in the hydrophobicdomain were markedly inhibited in 16K protein binding. Most interestingly, the placement of a glutamine in several random hydrophobic sequencesfacilitated 16Kprotein binding,defining this residue as apotential bindingsite for the 16K protein component of the proton pump and exemplifying the critical role of hydrophilic amino acids for mediating
specificinteractionsbetween transmembrane proteins.
Papillomaviruses induce the benign proliferation of epithe-lial cellsofmany vertebrate hosts(30). Some
papillomavi-ruses,including those human papillomavirus subtypes which infect genital epithelium, are also associated with lesions
thatmayprogress to cancer(28, 41).Itisbecomingapparent
that
papillomaviruses,
like other-DNA tumor viruses,en-code
transforming
proteins
whichinteractwithcellularpro-teins involved in the control of cell growth (32). For
in-stance, the E6 and E7 proteins of human papillomavirus
types 16 and 18 complex with the p53 (39) and RB (8, 26) proteins, respectively.
Bovine
papillomavirus
type 1(BPV-1) induces the efficient transformation of murine fibroblast cell lines (7, 24) predom-inantly via the activity of its E5gene(6, 18, 31,33). The E5oncoprotein
isahighly
hydrophobic protein
composed of44amino acids that localizes toGolgi apparatus, endoplasmic reticulum, and plasma membranes (3, 4). It iscomposed of
twodistinctdomains,one
being
a30-amino-acid,hydropho-bic,
amino-terminal domain which ispredicted
to traversethe membrane, and the otherbeinga
14-amino-acid,
hydro-philic, carboxyl-terminal
domain which ishighly
conservedamongthe
fibropapillomaviruses (3, 20).
Geneticanalysis
ofthe E5
oncoprotein
hasdefined several amino acids whichare
required
forfocus-forming
ability
within the smallhy-drophilic
domain(20),including
twocysteine
residueswhich mediate homodimer formation. In contrast, conservative amino acid substitutions within thehydrophobic
domain do notsignificantly
alter the transforming ability of E5(20).
*Correspondingauthor.
Although there doesnotappear tobeastrictrequirement for
a
specific hydrophobic
amino acid sequence within this region, the hydrophilic glutamine residue at position 17 appears to becritical forE5 function(20, 21, 23).Several lines of biochemical evidence suggest that
growth
factor activation may be an important component of
E5-mediated cellular transformation. For example, E5
stimu-lates the
phosphorylation
of the epidermal growth factor receptorinaligand-independent mannerand decreases theinternalization of occupied receptors (25). Similarly, the
endogenous
cellular,B-type receptorfortheplatelet-derived
growth factor(PDGF) is
constitutively
activated in C127 and FR3T3 cells stably transformed by the E5 protein (29). On the basis of amino acid sequencesimilarity
between thecarboxyl
termini of the E5protein
and PDGF and the conservation of these same amino acid residues in otherfibropapillomavirus
E5proteins
(20), Pettietal.(29)
specu-lated that activation of the PDGF receptor
might
be medi-atedby adirectinteraction between the E5protein
and thePDGFreceptor.However, direct interaction betweentheE5
protein
andgrowth
factorreceptors has not yetbeen dem-onstrated, and it ispossible
that receptor activation is indirect andrequires
E5 interactions with other cellular proteins.Recently, E5
epitope
fusionproteins
were showntoasso-ciate
specifically
with a 16-kDa cellularprotein
(16K
pro-tein).
Thisprotein
has now been identified as thepore-forming
constituent of vacuolarH+-ATPases
(15)
and isessential for acidification of subcellular compartmentssuch as endosomes,
lysosomes,
clathrin-coated
vesicles,
and Golgi vesicles. Sincegrowth
factorreceptorsareprocessed
405on November 10, 2019 by guest
http://jvi.asm.org/
in the
Golgi
apparatus and are internalized via clathrin-coated vesicles, E5binding
tothe 16K protein within thesecomponents maybean intermediate steptoreceptor
activa-tion and
signal
transduction. Wepreviously
showed that atransformation-defectiveE5molecule
containing
aglycine
atposition
17 failed to bind to the 16K cellular protein,sug-gesting that this
region
of the E5 protein may be abinding site for the 16K protein and that the association may beinvolved in
E5-mediated
transformation(16).
To furthercharacterize the
E5-16K
interaction,anumber ofE5mutants wereexaminedby
coimmunoprecipitation
for theirability
tointeractwith the 16K
protein.
Ourfindings
demonstratethatthe
glutamine
atposition
17 is critical for 16K proteinbinding,
thusdemonstrating
thathydrophilic
amino acidresidues which are
presumably
membrane embedded can mediatespecific
interactions between theox-helical,
trans-membrane domains ofregulatory
proteins.MATERIALS AND METHODS
Cellculture. The cell lines COS-33 (a
gift
from J.Brady)
and CMT4
(13)
were grown in Dulbecco's modifiedEagle
medium
(DMEM)
supplemented
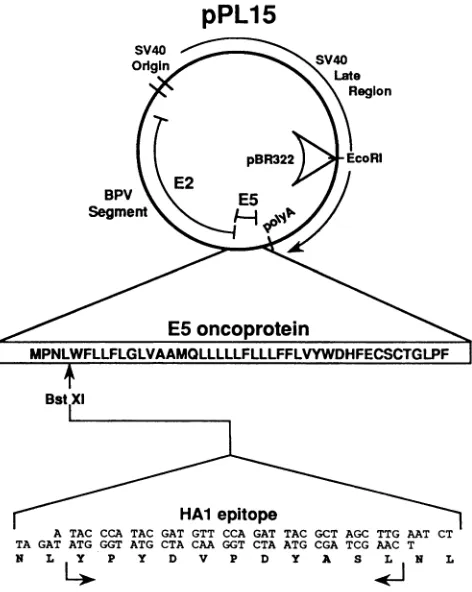
with 10% fetal calfserum.Plasmidconstructions.The
plasmid pPava-1
consistsofthe BstEIl (nucleotide Int]2405)-to-BamHl
(nt4450)
fragment
ofBPV-1,whichcontainstheentireE2-E5open
reading
frames(ORFs)
insertedinplace
of the HindIII (nt5171)-to-B(/I
(nt2770)
large
tumor(T)
antigen-coding
fragment
of simianvirus 40
(SV40)
(35).
Theseviral sequences wereclonedintopBR322at the uniqueEcoRIsite in the SV40 late
region
(nt1782).
Allplasmids
containing mutated E5 ORFs (seeFig.
3B,
4B,
andSB)
werederived from Pava-1(23,
35).
PL15and mutated E5plasmids
used toanalyze
16Kprotein binding
were constructed by inserting the
synthetic
oligonucleotides
corresponding
to the HAl epitope shown inFig.
1 into theunique
BstXI sites within the E5ORFs.
Theseoligonucleo-tides contain
BstXI-compatible
endswhich,
whenligated
intotheBstXlsitewithin theE5gene, resultintheloss of the
enzyme recognition sequence.
DNA transfer and virus
preparations.
Recombinant virus stocks wereprepared
as describedby
Settleman and DiMaio(35).
Briefly,
theplasmid
constructs listed inFig. 3B,
4B. and 5B weredigested
withEcoRI, circularizedby
ligation
ata DNA concentration of 5
pLg/ml,
and used to transfect CMT4 cellsby
the calciumphosphate
method of Grahamand van der Eb
(17).
Following
transfection,
cells were allowed to incubate for5days
at37°C
in DMEM containing 10% fetal calf serum. 1FiM
CdSO4,
and 100 FLMZnCI,.
Primaryvirusstockswere
prepared
by
scraping the cellsinto the medium,repeated
freeze-thawing,
and removal of celldebris
by centrifugation. Amplification
ofvirus stocks was achievedby
infecting
fresh,
inducedCMT4 cells withclari-fied supernatantand
incubating
theinfected cells forupto6 days. This step wasrepeated
toensurehigh-titer
stocks. The final clarified supernatant wasaliquoted
and stored at-70°C.
Virus titers were determinedby
the transactivationmethod ofSettleman and DiMaio
by using
the NL-3D cell line(35).
Typical
virus stocks contained-10'
infectious units per ml. Allsubsequent
infections were done at amultiplicity
of infection of between 10 and 100 infectiousunits per cell.
Fast
protein liquid chromatography (FPLC)
gel filtration. Cell membraneswereprepared
fromPL15-and Pava17GW-infected COS cells
by hypotonic
lysis as describedprevi-ously
(3).
Following
finalcentrifugation,
membrane pelletswere
resuspended
and extracted in 0.5 ml of modifiedpPL15
BstXi
HAl
epitope
A TAC CCA TAC GAT GTT CCA GAT TAC GCT AGC TTG AAT CT
TA GAT ATG GGT ATG CTA CAA GGT CTA ATG CGA TCG AAC T
N L Y P Y D V P D Y A S L N L
FIG. 1. Expression plasmids for wild-type and mutated E5
epitope proteins. pPL15 and mutated E5 epitope plasmids were
constructed by inserting synthetic oligonucleotides encoding the
influenza virus hemagglutinin HAI epitope into the uniqueBstXI
sitewithin the E5 ORF ofplasmid pPava-1 and of pPava-1-derived plasmids containing mutated E5 ORFs. pPava-l expresses the
E2-E5 ORFs of BPV-1 and was used for the construction of
SV40-BPV-1 recombinant viruses as described in Materials and
Methods (35). These viruses wereused to infect COS cells andto produce high-level, transient expression of the E5 protein. The sequences encoded by the E5 ORF and the inserted epitope are
indicated withsingle-letteraminoacid abbreviations.
radioimmunoprecipitation assay(RIPA) buffer(16) contain-ing gel filtration standards (Bio-Rad, Richmond, Calif.). Extracts were separated by gel filtration on a Pharmacia FPLC System (Pharmacia Inc., Uppsala, Sweden), using a Superose 12 HR 10/30 column at a flow rate of0.3 mil/min.
Fifteen 1.0-ml fractions were collected over a molecular
weight range of between 670,000 and 1,350 and were ana-lyzed for the presence of E5 and the 16K protein by immunoprecipitation using the monoclonalantibody 12CA5. Immunoprecipitation assays. Subconfluent COS cell cul-tures in 100-mm plates were infected at a multiplicity of infection of 100 infectious units per cell with recombinant BPV-1-SV40 viruses. Between 48 and 72 h postinfection,
cells were washed with phosphate-buffered saline (PBS),
incubated with methionine-free DMEM for 1.5 h, and
la-belled for 4 h with 750 jCi of a commercial mixture of
3SS-labelled methionine (-90%) and cysteine (-10)
(Trans-label, ICN) in2.5 ml of DMEM at 37°C. Cellswerewashed
twice with PBS and incubated in 1 ml ofa modified RIPA buffer (20 mM MOPS [morpholiinepropanesulfonicacid],150 mM NaCI, 1 mM EDTA, 1% Nonidet P-40, 1%
deoxycho-late, and 0.1% sodium dodecyl sulfate [SDS], pH 7.0) containing 0.1 mM protease inhibitors Nu-p-tosyl-L-lysine
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.318.554.73.369.2]PL1
5
Mol. weight
-*
670 standards18
-158 44 17 2
14
6-3-i
Fraction number -)- 6 7 8 9 10 11 12 13 14 15 16 17 18 19
18-Pava
17G
14-
6--di,
3 -I
FIG. 2. FPLC gel filtrationof wild-type- and mutant-infected COScells.Purified cell membranes from PL15- andPava17G-infectedCOS
cellswereextracted in RIPA buffercontaining molecular weight standardsand separatedon aSuperose 12 columnasdescribedin Materials
andMethods. Collected fractions were immunoprecipitatedby using antibody 12CA5, and precipitated proteins wereelectrophoretically
separatedonSDS-15% polyacrylamide gels and visualized by fluorography. Immunoprecipitatedproteins from fractions6through 19are
shown forboth PL15-(top) andPava17G- (bottom) infectedextracts.Major protein peaksaredesignated withsmallarrowsinfraction 12for
PL15and in fraction14forPava17G. Positionsofgelfiltrationmolecularweightstandards(inthousands)areindicatedat top.Positions of
molecular weight markers (inthousands) used during electrophoresisareindicatedtothe left of the gel. Positions of the16Kand E5epitope
proteinsareindicatedtothe right of the gel.
chloromethyl ketone (TLCK; Sigma) and 0.5 mM phenyl-methylsulfonyl fluoride (Sigma). Followinga30-svortexing,
nuclei were removed by centrifugation in an Eppendorf
microcentrifuge for 4 min. Five hundred microliters of each
extractwasincubated with 3
RI
of ascites fluid 12CA5, whichrecognizes the influenza virus hemagglutinin HAl domain(a
generousgift from I. Wilson) and incubated for 1 hat4°C.
Fifty microliters of protein A-Sepharose CL-4B beads (Pharmacia, Inc.) wasadded, and extractswere allowed to incubate foran additional 30min. Following six washes in
RIPAbuffer, Sepharose beadswereresuspended in 75 ,ul of
sample buffer withorwithout ,-mercaptoethanol, heatedat
100°C for 5 min, and separated by electrophoresis on 15%
SDS-polyacrylamide gels. Gelswerethen fixed withglacial
acetic acid-methanol, treated with Enlightning (New En-gland Nuclear), dried, and exposedtoKodak XAR-5 film for 24 hat -700C.
RESULTS
Construction and expression of wild-type and mutated E5
epitope fusion proteins. To identify cellular proteins that
interact with the BPV-1E5 transforming protein,weinserted
twodifferentepitopesinto theamino-terminal regionof the
E5 protein and expressed the fusion proteins by using a
recombinant SV40-BPV-1virusin COScells(16, 35).Fusion
proteins containinginsertions within this domain ofE5were
shown previously to maintain biological activity and to localize normally tothe Golgi apparatus. Using antibodies
thatrecognize these epitopes,we canefficiently
coimmuno-precipitate the E5 epitopefusionproteinsandanassociated,
16-kDa cellularprotein (16) which has beenrecently
identi-fiedasthe16Kcomponent of the vacuolarH+-ATPase (15). To evaluate the E5-16K protein interaction, the HAl
epitope from the influenza virus hemagglutinin (9) was
inserted into the 5' end of E5 genes, containing various
substitution mutations (20, 23, 36). Oligonucleotides
corre-spondingtothe HAl epitope were inserted into theunique
BstXI site of the pPava-1 plasmid (and its mutant deriva-tives) shown in Fig. 1 (nt 3889). SV40-BPV-1 recombinant virus particles were generated in induced CMT4 cells as
described previously (see Materials and Methods) (35). Virusesexpressingmutantandwild-type E5 fusion proteins
were assayed forexpression and bindingtothe 16Kprotein in COS cells as described previously (see Materials and Methods)(16).
FPLC analysis of the E5-16K protein complex. To further document theE5-16K protein interaction andtogain insights into the nature of the association, metabolically labelled extracts from cells expressing either wild-type HA1-E5 or
mutatedHA1-E5wereanalyzed by gel filtration usingFPLC
asdescribed in Materials and Methods. Recombinant virus
PL15 expresses large amounts of the wild-type HA1-E5 fusion protein in infected COS cells (16). Virus Pava HAl/
17G(previously designated p36W [16]) expressesamutated
HA1-E5 molecule containing a glycine residue in place of
glutamine at position 17. This mutant was shown to be
defective for focus formation, acute morphologic
transfor-mation, and inductionof DNA synthesis (20, 36)and is also
defectiveforbindingtothe 16Kprotein(16). Fractionswere
collected from a molecular weight range between 670,000 and1,500 andwereanalyzedfor thepresenceofE5and the 16K protein by immunoprecipitation with the monoclonal
antibody 12CA5.Theautoradiograms shown in Fig. 2 dem-onstratethat wild-type HA1-E5 and 16K proteins coeluted from the column as a complex with a molecular weight of approximately 44,000 (lane 12).
In contrast, FPLC fractionation of extracts from Pava HA1/17G-infectedcells revealed thepresenceofonlytheE5
4 -16k
* E5
* E5
III
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.131.493.77.284.2]fusion
protein
withvirtually
no detectablecoprecipitated
16K
protein
(Fig. 2).
Inaddition,
whereas thepeak
of wild-type E5 eluted from the column was in fraction12,
mutant E5
protein appeared
in a broadpeak
centered on fraction14,
aposition
relativetostandardscorresponding
to amolecularweight
ofapproximately 12,000.
Itappears thatthe reduced
ability
ofthemutantE5protein
tobind the 16Kprotein
resulted inasignificant
andreproducible
alteration intheelutionpatternofE5.Theapparent shift
corresponds
to a decrease in molecularweight
of about30,000,
a valuewhich could
correspond
to two16Kprotein
molecules. Sincethe extractions and
gradient separations
wereperformed
innonreducing conditions,
it islikely
that the12,000-molecu-lar-weight peak
observed with the mutated E5protein
rep-resents
uncomplexed
E5 dimers(which
run on SDSgels
at14 to 15
kDa).
Thepeak
observed in fraction 12 with thewild-type
E5 represents E5dimers,
eachprobably
com-plexed
withtwo16Kprotein
molecules. Whiletheresolution of FPLC forthesesmallproteins
is insufficienttodefinitively
determine the
stoichiometry
of E5-16Kprotein binding,
previous findings
that monomeric E5protein
can bind the 16Kprotein (16)
areconsistent withthehypothesis
thateach E5 dimercan bindto two 16Kprotein
molecules.Transformation-defective,
carboxyl-terminal mutations inE5donotaffect16K
protein binding.
Wepreviously
demon-stratedthat thetwocysteine
residuesatpositions
37and39,
although
crucial for E5 dimer formation andtransforming
activity,
arenotrequired
for interaction with the16Kprotein
(16).
Thissuggested
that theE5hydrophilic
domainmaynot have a direct role in 16Kprotein binding.
To address thisquestion,
several mutantscontaining
substitutions ofcon-served amino acids withinthe
carboxyl-terminal
domainof E5weretestedfor16Kprotein
association. Included in this series of mutations were amino acid substitutions which abolishE5-mediated cellular transformation and the induc-tion ofacute DNAsynthesis (20, 36).
HAlepitopes
were inserted intotheunique
BstXIsite ofPavavectors withE5 ORFscontaining
varioussubstitutions within this domain.A summary ofthe mutants tested andtheir abilities toinduce transformationandassociate with the16Kprotein
areshown inFig.
3B. Levels of 16Kprotein binding
in this andsubsequent experiments (see Fig.
4 and 5) are expressed relative tothatofthe wild typeas aratio of16Kprotein/E5
protein
as measuredby using
both the Ambis and USB SciScan 5000 automatedscanning
systems.COS cells infected with PL15 and the
carboxy-terminal
substitution mutants were analyzed for expression ofthe
HA1-E5
proteins
and association with the 16K protein bycoprecipitation
with 12CA5 ascites fluid.Theautoradiogram inFig.
3Ademonstratesthat all mutants,with theexception ofPavaHA1/34Q36D41A,
expressed
significant
levelsof E5proteins
and that the 16Kprotein
wascoprecipitated ineachcase. Previous analysis of 34Q36D41A without the
HAl
epitope
demonstratedthatthismutantE5proteinwas stable.Following
overexposure of this autoradiogram, E5 andco-precipitated
16Kproteincouldeasilybe detected from PavaHA1/34Q36D41A-infected
COS cell extracts (data notshown).
Theseresultsdemonstratethat the conserved aminoacids ofthe
carboxyl-terminal
domain of E5, althoughre-quired
fortransforming
activity, may not be involveddi-rectly
in the association with the 16K protein. Threecar-boxyl-terminal
mutants tested appeared to form dimersnormally (Fig.
3A, lanes 8 through 14). MutantsHAl/
30L35S and
HA1/32S,
however,
consistently showed twodimer
populations
thatmigrated
distinctly uponpolyacryl-amide
gel electrophoresis
analysis. This result suggests thatthese mutant molecules may bealigning themselves in two different orientations(parallel and antiparallel) and therefore may not beforming properly alignedE5dimers.
The glutamine residue in the ES transmembrane domain
regulatesbindingtothe 16Kprotein. The observation thatan E5 moleculecontainingaglutamine-to-glycine substitutionat
position 17 failed to induce transformation orbind the 16K
proteinsuggested that this position within E5maycontribute
to 16K
protein binding
and therefore modulate E5-induced transformation. To furtherinvestigate these possibilities, sev-eral substitution mutantscontaining
HAlepitopes
wereas-sayed
for16Kprotein
association(Fig. 4). Pava 17L containsanE5ORF withaleucineat
position
17and,
likemutant17G, failstoinduce E5 transformation ina focus formationassay(23). Pava1ST17Hcontainsathreonineat
position
15 inplace
ofan alanine and a histidine at position 17 in place ofa
glutamine. Unlikemutants17G and
17L,
thismutantinduces efficient E5 transformation (20, 36). Pava 15V21F22Vcon-tains conservative substitutionsatpositions15, 21, and 22 and also
efficiently
transforms cells(20, 36).
COS cells wereinfected with recombinant viruses containing wild-type and mutated versionsof theHAl/ES
protein
andwereassayed by
coprecipitation
ofES andthe 16Kprotein by using
monoclo-nalantibody 12CA5. LikemutantHA1/17G,
mutantHA1/17Lwas severely defective for16Kprotein association
(Fig.
4A,lane2). The16K
protein
wasdetectedonly
when thesamplewas overloadedonthegel(Fig. 4A) orwhenthe
autoradio-gram was
overexposed.
In contrast,transformation-compe-tent mutants HA1/1ST17H and HA1/15V21F22V associate
efficiently
with the 16Kprotein.
These results indicate thattheglutamine residue at
position
17isimportant
for associa-tion withthe 16Kprotein butthatotherpolar aminoacids,
orperhaps charged amino
acids,
cansubstitute forglutamine.
To assesstheability ofmutatedrecombinant E5proteins
toform dimers, immunoprecipitated pelletswereresuspended insam-ple buffer in the absence of ,B-mercaptoethanol. As shown in
the
autoradiogram
inFig.
4B, all mutants in this series appearedtoform dimersnormally.Horwitzetal.(21)reported the characterization ofseveral E5 mutants
containing
substitutions of the hydrophobic middle third of ES with apparently unrelated hydrophobicsequences. In almost all cases, such drasticsequence sub-stitutions severely inhibited ES focus-forming ability. More
recently, Kulkeet al. (23) demonstrated that insertion ofa
glutamine
residueatposition
17 allowed severaltransforma-tion-defective,
hydrophobic substitution mutants to induce stable transformation and stimulate DNA synthesis in anacute assay using C127 cells. To determine whether there
exists acorrelation between the ability ofthese mutants to
induce morphological transformation and association with
the16K
protein,
twomutant ES ORFs contained withinthe Pava vector wereanalyzed for their abilitytoassociate with the 16K protein (sequences shown in Fig.SB).
The first, PavaHR25, is atransformation-defective mutant containingahydrophobic sequence,apparently unrelated to that of the
wildtype,within themiddlehydrophobic domain of
ES
(Fig.5B).
The second, Pava HR25Q, contains the same ESsequence as PavaHR25, withtheexception of a glutamine
residue atposition 17 (23). Insertion of a glutamine at this
position
allows this mutant to induce focus formation and DNAsynthesis(23). To determine whether the generation ofES
transforming
activity with the inserted glutamine wasaccompanied
by alterations in the ability to associate withthe16Kprotein,theHAl epitopeswereinsertedinto the
ES
ORF of both Pava HR25 and Pava HR25Q to generate
recombinant viruses, Pava HR25W and Pava HR25QW.
on November 10, 2019 by guest
http://jvi.asm.org/
A
U)
-J) Cl) U) *) U)
+BME <:
0
C') Lfl Je > a0
C., -t -i 0
Ul) Ul) 0. 2
Eu-)
18-16K->'l
14-E5Mne Monomers
3-1 2 3 4 5 6 7
U)
U,
C.) U) XJ
cn CN
r~- 0 C.) C')
-BME
<:
er
a
tc
uh > as _
N CO) Tr J
Ull) C) EL 2
E5 Dimers
16 K
E5
Monomers
8 9 10 11 12 13 14
R
NH COOH1 ~~~~~~~~~~~~~~44
--- X
trans-30 31 32 33 34 35 36 37 38 39 40 41 formatIon wild type-op-val--tyr--trp--asp--his--phe--glu--cys--ser--cys--thr--gly .++
mutant 32S 33V
16k binding
ser --- --- __
val
--37S --- --- --- --- --- --- --- ser --- -- --- +++
37S39S --- --- --- --- --- --- --- ser --- ser --- ++
34036D41A--- gln --- asp - - - ala +++ +++
30L35S leu -- e-r---sr --- --- - +++
FIG. 3. Coprecipitation of E5-16K protein complex and dimer formation in cells expressing E5 epitope proteins containing amino acid substitutions within the carboxyl-terminal domain. (A) Extracts of COS cells infected with the viruses indicated above the lanes were immunoprecipitated with 12CA5, electrophoretically separated on SDS-15% polyacrylamide gels, and visualized by fluorography. Molecular weights (in thousands) are indicated on the left, and positions of the 16K protein and E5 monomers and dimers are indicated on either side.
3ME, ,-mercaptoethanol. (B) Summaryofmutantscontainingsubstitutions within thecarboxylterminus of E5. The
expanded
sequence shows thepositions of substituted amino acids within the carboxyl-terminal domain. The columns on the right show phenotypes for focus formation andabilitytobind the16Kprotein.Datafor focus formation was obtained from Horwitzetal. (20).Quantitationwasperformed using theAmbis scanner and the USB SciScan 5000 automated scanning system and are expressed relative to that of the wild type as a ratio of 16KproteintoE5protein. + ++,>50%; ++, 20to50%;+,5to19%o; -,<5% (percentagesarerelative to wild-typelevels). Valuesareaveragesof several immunoprecipitation experiments.
Virus preparations were generated in induced CMT4 cells
and used to infect COS cells for expression of the ES
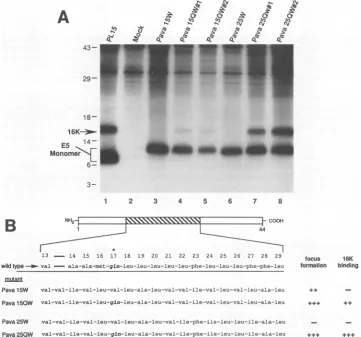
recombinantproteins. Figure SA shows the results of immu-noprecipitation experiments using Pava 25W and two inde-pendent isolates ofPavaHR25QW. Whereasno16Kprotein was detected in precipitations from extracts of Pava HR25W-infected cells (lane 6), significant levels of 16K
protein were coprecipitated from extracts of cells infected with both PavaHR25QW isolates (lanes 7 and 8).
These results imply that the placement ofa hydrophilic
glutamine residue inarandomhydrophobicdomainofE5is
criticalforcell transformation and 16Kprotein association.
Inapparentcontradictiontothesefindingsisthe existence of
a hydrophobic substitution mutant, Pava HR15 (Fig. Sa), whichlacks glutamine andcan still efficientlyinduce focus
formation and DNAsynthesis in C127 cells (21, 23).
Inser-tion ofa glutamine at position 17 (construct PavaHR1SQ) enhancedfocus-forming abilityandacuteinduction of DNA
synthesis
(23). To test whetherthesemutantsassociate withthe 16Kprotein, HAl epitopeswereinsertedto createPava 15WandPavaHR1SQW. Extractsfrom PavaHR15W- and Pava HR1SQW-infected COS cells were analyzed for
E5-16K protein association by
immunoprecipitation
with 12CAS. Figure 5A shows that little E5-associated 16Kpro-tein was detected in Pava 15W-infected extracts but that it was easily detected in extracts ofcells infected with two independent isolates ofPavaHR15QW.
DISCUSSION
The BPV-1 E5
oncoprotein
is the smallestknownprotein
withtransformingactivity forwhich thedetailed
genetic
and biochemicalmanipulations
necessaryforcharacterization ofthe
multiple
steps ofsignal
transductionarepossible.
Acti-vation ofgrowth factor receptors appears to be a central
event in E5-mediated transformation
(25, 29).
Theon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.141.498.76.435.2]A
+BMECMC14 LL
,2
3..
0.
...
-
BME
U.
r_I
C, -J - >
r. r-. u, in
43-
29- 18-16K *
14-E5 -f
Monomers
6-
3-n X
0
0. 0
- E5
Dimers/l 6K
1}- E5
Monomers
1 2 3 4 5 6 7 8 9 10 11
NH2t
1.5 16 17 18 19 20 21 22 23
wildtype -B- ala--met--gln--leu--leu---leu--leu--leu--phe mutant
17G gly __
17L leu --- --- ---
-15T17H thr --- his ++t +++
15V21F22V val phe val --- +++ +++
FIG. 4. Coprecipitationof E5-16Kprotein complexand E5 dimer formationin cellsexpressingE5epitope proteins containingaminoacid substitutions within the middlehydrophobic region. (A)Extracts of COS cells infected with the indicated viruseswereimmunoprecipitated
with12CA5, electrophoretically separatedonSDS-15% polyacrylamide gels,and visualized by fluorography. Proteins eluted fromprotein A-Sepharosewith SDS samplebuffercontaining ,-mercaptoethanol (+,BME)areshownonthe left(lanes1through 6).Toanalyzemutant E5molecules for abilitytoform dimers, proteinswereeluted in the absence ofreducingagent(- ,ME)andareshownin theright panel (lanes
7through 12).Positions ofE5monomers,dimers,and 16Kproteinareindicatedtotherightand leftofthegel.Positions of molecularweight
markers(in thousands)areindicatedtothe left of thegel. (B) Summaryofmutantscontainingsubstitutions within the middlehydrophobic
domain of E5. The baratthetoprepresentsthe E5protein, and thestriped areaindicatesthe substituted region.Theexpandedsequence shows thepositionsof substituted amino acids. The leftmost column contains thenamesof themutantconstructs.Thecentersectionshows the amino acidsubstitution(s)encodedbythecorresponding mutants. The columnsontherightshowphenotypesfor focus formation and
abilitytobindthe 16Kprotein. Datafor focus formationwasobtained from Horwitzetal.(20). +++,>50%; ++,20to50%;+, 5to19%;
-,<5% (percentagesarerelativetowild-type levels).
nism by which E5 activates these receptors, however,
re-mains unclear. Binding of E5tothe 16-kDacomponentof the vacuolar H+-ATPases could be one step involved in the activation of the cellular growth factor signal transduction pathway (15). The results presented in this report further verify the existence oftheE5-16Kprotein complex, suggest that dimeric E5 binds two 16K molecules, and identify an
amino acid in E5 critical for the interaction with the 16K
protein.
FPLC analysis of the E5-16K proteincomplex.Gel filtration by FPLC analysis has providedanalternativeapproachfor
characterizing the nature of the E5-16K protein complex.
Coelutionofwild-type HA1-E5 and16Kproteinssupported
coprecipitation data and provided independent evidence for the in vivo existence of the E5-16K protein complex. In
addition, comparison of elution profiles of wild-type and
mutant E5 molecules provided information regarding the
protein composition of the multimeric complex. The elution patterns ofwild-type E5-16K protein complex were
consis-tentwiththe existence ofacomplex consisting oftwo16K proteins and an E5 dimer. Profiles of eluted E5 protein
expressed frommutant17G-infected cellsdisplayeda
signif-icant and reproducible decrease in apparent molecular weight. The difference observed correspondedtothe size of two16K molecules and reflected the absence of 16Kprotein binding to the E5 dimer. Furthermore, these data suggest that, under the conditions of isolation utilized, no proteins
other than the 16Kproteinarestably boundtotheE5 dimer. However, cell extracts prepared with RIPA buffer do not preserve many protein-protein interactions, and it is not possible to preclude the existence of other E5-associated proteins. Because of the strongly hydrophobicnatureof both E5 and the 16K protein (15), it has been difficult to extract these proteins from cells with nonionic detergents such as
Nonidet P-40 and Triton X-100 (unpublished observations). However, gel filtration ofextracts prepared with the zwitter-ionic detergents CHAPS {3-[(3-cholamidopropyl)-dimethyl-ammonio]-1-propanesulfonate} and CHAPSO
{3-[(3-cholami-R
12
J-COOH
44
trans- 16k formation blnding
_
I
F
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.128.486.73.405.2]A
433-
29-
18-16K--
s
1111 E514-Monome>[ E
6-'9~~ ~ ~ ~~~~~x
ml .m m2
3-1 2 3 4 5 6 7 8
NH2 y > ~~~COON
44
13 - 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 wildtype-- val - ala-ala-met-gln-leu-leu-leu-leu-leu-phe-leu-leu-leu-phe-phe-leu
mutant
Pava 15W val-val-ile-val-leu-val-leu-ala-leu-val-val-ile-val-leu-val-leu-ala-leu
Pava150W val-val-ile-val-leu-gln-leu-ala-leu-val-val-ile-val-leu-val-leu-ala-leu
Pava 25W val-val-ile-val-leu-val-leu-ala-leu-val-ile-phe-ile-leu-leu-ile-ala-leu
Pava 250W val-val-ile-val-le--gln-leu-ala-leu-val-ile-phe-ile-leu-leu-ile-ala-leu
focus 16K formation binding
+4+S ++
[image:7.612.135.496.74.411.2]. . .+. .
FIG. 5. E5 bindingtothe16Kprotein requiresaglutamine residue within themiddle hydrophobic region ofE5.(A)COS cellsinfected with viruses containinghydrophobic substitutions withintheregion between amino acid positions 13and29wereanalyzedforE5-16Kprotein
complexformationbycoprecipitationwithantibody 12CA5. Immunoprecipitated proteinswereseparatedon anSDS-15%acrylamide gel and
visualizedbyfluorography. Positionsof molecular weight markers (in thousands), 16Kprotein, andE5monomers areindicatedtotheleft of thegel. (B) Summaryandaminoacidsequencesofhydrophobic substitutionmutants.Thestriped and expandedarea(amino acidpositions
13through 29)is theregion ofE5 thatwasmutated(23).Theleftmost column showsthenamesof the virusconstructsusedin this study.The center areashowsthe substitutedsequences. The position of the glutamine residue atposition 17is indicated by theasteriskabove the wild-typesequence.The columnsonthe right show phenotypesforfocus formation and 16K protein binding. Data for focus formationwas
obtained from Kulkeetal. (23). + ++, >50%; ++,20to50%; +,5to 9%o; -,<5% (percentagesarerelative towild-type levels).
dopropyl)-dimethylammonio]-2-hydroxy-1-propanesulfonate}
revealed the existence ofaveryhigh-molecular-weight
com-plex(400,000to500,000), suggesting the possibility of other associatedproteins(unpublished observations).
Mapping the requirements for 16K protein binding. By testing the ability ofavariety of E5mutants withsingle or
multiple amino acid substitutionstobindthe16Kprotein,we
identified aspecific amino acid that is essential for binding
the 16Kprotein. The primary determinant for 16Kprotein bindingappearstoreside in thehydrophobic central portion ofthe ES protein. None of the missense mutations in the hydrophilic carboxyl-terminal third of the ES protein
af-fected 16Kprotein binding, includinganumberof mutations
that severely inhibit transformation and/or block
dimeriza-tion. Thus, itappears unlikely that there are many, ifany, specificcontactswith the 16Kproteinin thisportionof the
molecule.However, themutantssurveyedinthisstudyeach
contain onlyone ortwoamino acidalterations, which may not sufficiently destabilize E5-16K protein binding, even
though they interfere withtransformation.
Instrikingcontrast,thereisanabsoluterequirementfora
hydrophilic amino acid in the middle hydrophobic domain forbinding ofthe16Kproteinasassessedby
coimmunopre-cipitationorby FPLC. This residue is glutamine in thecase
ofthe wild type and is an invariant amino acid among all
sequenced fibropapillomavirus ES genes, and it can be
functionally replaced by histidine, another hydrophilic amino acid. In all hydrophobic sequence contexts tested, including random ones with diverse effects on
transforma-tion, there was no detectable 16K protein binding in the
absenceof theglutamine and readilydetectable 16Kprotein binding in itspresence. Thus,itappearsthataglutaminein thehydrophobic region issufficientfor16Kprotein binding, although the effect of its positioning has not been
deter-mined.
E5-16K protein binding and cell transformation. Our
re-sults clearly show that the ability of ES to bind the 16K protein is not sufficient for cell transformation, because many carboxyl-terminal, transformation-defective mutants bind16K.Amongthesevenamino acidsin thisregionshown to be required for cell transformation are two cysteine residues which mediate homodimer formation. A mutant
B
on November 10, 2019 by guest
http://jvi.asm.org/
containing amino acidsubstitutions atbothcysteine residues
failsto form dimers, suggesting that dimerformation
medi-ated through disulfide linkage is necessary for cell
transfor-mation (20). Other transformation-defective mutations within this domaindo notapparently affectdimerformation
or 16K protein, indicating that these amino acids may be
carrying out some otherrequiredfunction, such as
interact-ingwith other proteins ormaintainingacritical
stereochem-ical orientation of the hydrophobic domain.
The ability of mutant HR15 to transform cells indicates
eitherthat E5-16K protein interaction maynotbeabsolutely required for transformation or that the hydrophobic se-quenceof HR15 cansomehowfunctionally substituteforthe
16Kprotein association. Inarelated study, amutant desig-nated HR16Q contains aglutamine within a different
unre-latedhydrophobicsequenceandbinds the 16Kprotein, yetit istransformationdefective, indicating thatnotall hydropho-bic sequencesthat bind the 16K protein can support trans-formation (unpublished results; 23). However, acorrelation
between E5 transformation and 16K protein association is
supported by thefinding that the transforming activity ofa
greatmajority ofmutants, including HR15, is stimulated by the insertion of the glutamine which allows 16K protein binding. In addition, the transformation defect of mutant 17L, which lacks the glutamine and is defective for 16K protein binding, is more severe than that of most other mutants we have tested, including many with mutations in
the carboxyl-terminal hydrophilic domain. Nevertheless,
additional mutants need to be generated in order to clearly
definethe role of16Kprotein binding in E5 transformation.
Disruptionof 16K protein functionas apotentialmechanism
of cellular transformation. The 16-kDa component of the
vacuolar H+-ATPase has been isolated from plasma
mem-brane and endomembrane compartments such as gap
junc-tions (11), lysosomes (19, 34), clathrin-coated vesicles (1,
37), andGolgimembranes (14, 40). Interestingly, disruption of the gene encoding the 16-kDa component of the yeast vacuolar proton pump resulted in the failure to process
newly synthesized carboxypeptidase Y, indicating that the
16K protein is required for acidification and formation of
trans-Golgi vesicles (27). Considering the intracellular
loca-tionsand activityof the 16Kprotein, there areseveral ways
inwhich the bindingofE5tothis pore-formingproteincould
conceivably affect growth factorreceptors. For example, a
local pH change induced by the E5-16K protein interaction may affect certain aspects of receptor processing, such as
ligand internalization and vesicular transport. Since the E5
protein is known to accumulate in the Golgi apparatus (4,
16), it is possible that it exerts its transforming effects on
growth factorreceptors atthis site. E5 alteration of
proton-translocating ability in the Golgi network may affect the
normal movement ofreceptors through this apparatus,
per-haps resulting inreceptor clustering and subsequent
activa-tion. Several lines of evidence exist which indicate that incompletely processed, Golgi-localized PDGF receptors
can be activated internally before reaching the cell surface (2, 12, 22, 29). E5 is alsopresent in the plasma membrane
andmayalsoexertaneffectatthis site(3). Martinetal. (25)
have shownthatE5 expressionis associated with agreater
number of epidermal growth factor receptors on the cell
surface, secondary to an inhibition ofreceptor
down-regu-lation. Analogous to what may be occuring in the Golgi
network, E5bindingtothe16K protein intheprotonchannel mayinhibitendosome acidificationand/or endosome
forma-tionand therebyprevent receptordown-regulation.
The finding that the 16K protein may be acomponentof
gap junctions offers an additional
regulatory
site for E5action (11). Inhibition of normal cell-cell communication via disruption of gapjunction function occurs
commonly
during
cellular transformation, and the gap
junctional
protein
con-nexin is the targetfor the viraloncoprotein
src (38). Identi-fication of the physiologic responses toE5-16K
protein
association should help to understand the mechanism of E5-mediated transformation as well as the cellular controls of signal transduction.
Finally, the role of hydrophilic, intramembrane amino acids in mediating the interactions of transmembrane pro-teins such as
E5
and the 16Kprotein may berepresentative
of a more generalbiologic phenomenon. Forexample,recent evidence suggeststhat
hydrophilic
residues within thetrans-membrane domain of the a chain of the T-cell receptor induce interaction with the
CD3-8
chain through an acidic residue in its transmembrane domain (5). It remains to bedetermined whether an analogous polar or charged amino
acid inatransmembrane domain ofthe 16Kprotein induces association with the
E5
protein or whether theE5
proteinassociates with additional cellularproteins via such interac-tions.
ACKNOWLEDGMENTS
This research was supported in part by grant R01CA53371-01 from the National Cancer Institute. R.K. was supported by a fellowship from the German Cancer Research Center.
REFERENCES
1. Arai, H., M. Berne, and M. Forgac. 1987. Inhibition of the
coated vesicle proton pump and labeling of a 17,000-Dalton polypeptide by N,N'-dicyclohexylcarbodiimide. J. Biol. Chem.
262:11006-11011.
2. Bejcek, B. E., D. Y. Li, and T. F. Deuel. 1989. Transformation by v-sis occurs by an
internal
autoactivation mechanism. Sci-ence245:1496-1499.3. Burkhardt, A., D.
DiMaio,
and R.Schlegel.
1987. Genetic and biochemical definition of the bovine papillomavirus E5 trans-forming protein. EMBO J. 6:2381-2385.4. Burkhardt, A., M.
Wilningham,
C. Gay, K.-T. Jeang, and R. Schlegel. 1989. The E5 oncoprotein of bovine papillomavirus is oriented asymmetrically in Golgi and plasma membranes. Virol-ogy 170:2381-2385.5. Cosson,P.,S.P. Lankford, J. S. Bonifacino, and R. D. Klausner. 1991. Membrane protein association by potential intramem-brane charge pairs. Nature (London) 351:414-416.
6. DiMaio, D., D.Guralski, and J. T. Schiller. 1986. Translation of open reading frame
E5
of bovine papillomavirus is required for its transforming activity. Proc.Natl.
Acad. Sci. USA 83:1797-1801.7. Dvoretzky, I., R. Shober, S. K. Chattopadhyay, and D. R. Lowy. 1980. A quantitative in vitro assay for bovine papilloma virus. Virology 103:369-375.
8. Dyson, N., P. M. Howley, K.
Munger,
and E. Harlow. 1989. The human papillomavirus-16 E7 oncoprotein is able to bind to the retinoblastoma gene product. Science 243:934-937.9. Field, J., J.-I. Nikawa, D. Broek, B. MacDonald, L. Rodgers, I. A. Wilson, R. A. Lerner, and M.
Wigler.
1988. Purification of a RAS-responsive adenylyl cyclase complex from Saccharomy-ces cerevisiae by use of an epitope addition method. Mol. Cell. Biol. 8:2159-2165.10. Finbow, M. E., T. E. J. Buultgens, N. J. Lane, J. Shuttleworth, and J. D. Pitts. 1984. Isolation and characterisation of arthropod gap junctions. EMBO J. 3:2271-2278.
11. Finbow, M. E., P. J. Jackson, J. N. Keen, L. Meagher, P. Thompson, and J. B. C. Findlay. Structure of a
16kD
integral membrane protein that has identity to the putative proton channel of the vacuolarH+-ATPase.
Biochem J., in press. 12. Fleming,T. P., T. Matsui, C. J. Moiloy, K. C. Robbins, and S. A.Aaronson. 1989. Autocrine mechanism for v-sis transformation
on November 10, 2019 by guest
http://jvi.asm.org/
requires cell surface localization of internally activated growth factor receptors. Proc. Natl. Acad. Sci. USA 86:8063-8067. 13. Gerald, R. D., and Y. Gluzman. 1985. New host cell system for
regulated simian virus 40 DNA replication. Mol. Cell. Biol. 5:3231-3240.
14. Glickman, J., K. Croen, S. Kelly, and Q.Al-awqati. 1983. Golgi membranes contain an electrogenic H'-pump in parallel to a chloride conductance. J. Cell Biol. 97:1303-1308.
15. Goldstein, D. J., M. E. Finbow, T. Andresson, P. McLean, K. Smith, V. Bubb, and R. Schlegel. 1991. The bovine papilloma-virus E5 oncoprotein binds to the 16 kilodalton component of vacuolar H+-ATPases. Nature(London) 352:347-349. 16. Goldstein, D. J., and R. Schlegel. 1990. TheE5 oncoprotein of
bovinepapillomavirusbindstoa 16 kd cellular protein. EMBO J. 9:137-146.
17. Graham, F. L., and A. J. Van der Eb. 1973. A new technique for the assay of human adenovirus 5 DNA. Virology 52:456467. 18. Groff, D., and W. D. Lancaster. 1986.Genetic analysis of the 3'
earlyregiontransformation and replication functions of bovine papillomavirus type 1. Virology 150:221-230.
19. Harikumar, P., and J. P. Reeves. 1983. The lysosomal proton pumpis'electrogenic. J. Biol.Chem.258:10403-10410. 20. Horwitz, B. H., A.Burkhardt,R. Schlegel, and D. DiMaio. 1988.
44-Amino acidES transforming protein of bovine
papillomavi-rusrequires ahydrophobic core and specific carboxyl-terminal aminoacids. Mol. Cell. Biol. 8:4071-4078.
21. Horwitz, B. H., D. L. Weinstat, and D. DiMaio. 1989. Stable cellulartransformationinduced by a 16-amino-acid segment of the bovine papillomavirus E5 protein linked to a random se-quenceofhydrophobic amino acids. J. Virol. 63:4515-4519. 22. Keating, M. T., andL. T.Williams. 1988. Autocrine stimulation
ofintracellular PDGF receptors in v-sis-transformed cells. Sci-ence239:914-916.
23. Kulke, R.,B. H.Horwitz, T.Zibeilo,and D.DiMaio. 1992. The central hydrophobic domain of the bovine papillomavirus E5
transforming protein can be functionally replaced by many hydrophobic amino acid sequences containingaglutamine. J. Virol. 66:505-511.
24. Lowy, D. R., I. Dvoretzky, M. F. Law, L. Engel, andP. M.
Howley. 1980. Invitrotumorigenic transformation byadefined subgenomicfragment of bovine papilloma virus. Virology103:
369-375.
25. Martin, P., W. C. Vass, J. T. Schiller, D. R.Lowy, and T.J.
Velu. 1989. The bovinepapillomavirus E5transforming protein can stimulate the transforming activity of EGF and CSF-1 receptors.Cell 59:21-32.
26. Munger,K.,B. A.Werness, N. Dyson,W.C.Phelps,and P. M.
Howley.1989. Complexformation of humanpapillomavirus E7
proteins with the retinoblastomatumorsuppressor gene prod-uct.EMBO J. 8:4099-4105.
27. Nelson, H., and N. Nelson. 1990. Disruption of genes encoding subunits of yeast vacuolarH+-ATPase causes conditional le-thality. Proc.Natl. Acad. Sci. USA 87:3503-3507.
28. Orth, G. 1987.Epidermodysplasia verruciformis, p. 199-243. In
N. P. Salzman and P. M. Howley (ed.), The papovaviridae. Plenum Press, New York.
29. Petti, L., L. Nilson, and D. DiMaio. 1991. Activation of the platelet-derived growth factor receptor by the bovine papillo-mavirus E5transforming protein. EMBO J. 10:845-855. 30. Pfister, H.1983. Biology andbiochemistry ofpapillomaviruses.
Rev. Physiol. Biochem. Pharmacol. 99:111-181.
31. Schiller, J. T., W. C. Vass, K. H. Vousden, and D. R. Lowy. 1986. E5 open readingframe of bovine papillomavirus type 1
encodesatransforming gene. J. Virol. 57:1-6.
32. Schlegel, R. 1990. Papillomaviruses and human cancer. Semin. Virol. 1:297-306.
33. Schiegel, R., M. Wade-Glass, M. S. Rabson, and Y.-C. Yang. 1986. The E5transforming gene of bovine papillomavirus
en-codes asmall, hydrophobic polypeptide. Science 233:464-467. 34. Schneider, D. 1981. ATP-dependent acidification of intact and
disrupted lysomes: evidence foran ATP-drivenpump. J. Biol. Chem. 256:3858-3864.
35. Settleman, J., and D. DiMaio. 1988. Efficienttransactivation and morphological transformation by bovine papillomavirus genes expressed from a bovinepapillomavirus/simian virus40
recom-binant virus. Proc.Natl. Acad. Sci. USA 85:9007-9011. 36. Settleman, J., A. Fazeli, J. Malicki, B. H. Horwitz, and D.
DiMaio. 1989.Genetic evidence that acutemorphological trans-formation, induction of cellular DNA synthesis, and focus formation are mediated by a single activity of the bovine papillomavirus E5 protein. Mol. Cell. Biol. 9:5563-5572. 37. Sun, S.-Z., X.-S. Xie, and D. K. Stone. 1987. Isolation and
reconstitution of thedicyclohexylcarbodiimide-sensitive proton pore of the clathrin-coated vesicle protein translocating com-plex. J. Biol. Chem. 262:14790-14794.
38. Swenson, K. I., H. Piwnica-Worms, H. McNamee, and D. L. Paul. 1990.Tyrosine phosphorylation of the gap junction protein connexin43 is required for the pp60vsc-induced inhibition of communication. CellReg. 1:989-1002.
39. Werness,B.A.,A.J. Levine,and P. M.Howley. 1990. Associ-ation of human papillomavirus types 16 and 18 E6 proteins associate withp53. Science 248:76-79.
40. Zhang, F.,and D. L.Schneider.1983. Thebioenergetics of Golgi apparatusfunction: evidence foranATP-driven proton pump. Biochem.Biophys. Res. Commun. 114:620-625.
41. zurHausen,H.,andA. Schneider. 1987. The role of papilloma-viruses in human anogenital cancer, p. 245-263. In N. P.
Salzman and P. M. Howley (ed.), The papoviridae. Plenum Press, New York.