Copyright
q
1997, American Society for Microbiology
Exon 1 Leader Sequences Downstream of U5 Are Important
for Efficient Human Immunodeficiency Virus Type 1
Gene Expression
CHRISTIAN LENZ, ANDREAS SCHEID,
ANDHEINER SCHAAL*
Center of Biological and Medical Research and Institut fu¨r Medizinische Mikrobiologie und Virologie,
Heinrich-Heine-Universita¨t, D-40225 Du¨sseldorf, Germany
Received 26 August 1996/Accepted 31 December 1996
In previous studies, little attention has been paid to maintaining the native HIV-1 leader sequence in
reporter constructs analyzing the human immunodeficiency virus type 1 (HIV-1) promoter activity. To
inves-tigate a possible influence of the leader sequence on HIV-1-driven gene expression in the presence as well as
in the absence of Tat, an expression vector was designed for transcripts consisting of the native HIV-1
tat
1.4
mRNA leader followed by the open reading frame for the bacterial chloramphenicol acetyltransferase (CAT).
Deletion mutants with mutations within the leader sequence downstream of U5 (lsdU5) were constructed, as
well as a mutant containing a mutation with a reverse orientation of this region. Quantification of CAT protein
in HeLa-T4
1cells transiently transfected with wild-type and mutant leader constructs showed that the exon
1-derived lsdU5 region has an influence on basal as well as Tat-induced protein expression. The dramatic
decrease in the level of CAT protein upon deletion of lsdU5 was paralleled by a drop in the steady-state level
of CAT mRNA. Deletion of the exon 1-derived lsdU5 region also decreased the expression of mRNAs containing
authentic HIV-1 sequences instead of CAT. The effect observed with the reporter constructs was not due to the
loss of binding sites for nuclear factors, as could be shown with DBF1 and Sp1 mutant constructs. Nuclear
run-on transcription assays showed that the presence or absence of lsdU5 did not influence the rate of
transcription. This indicates that the exon 1 lsdU5 element functions at the posttranscriptional level in the
processing, nucleocytoplasmic export, or stabilization of HIV-1 transcripts.
After the integration of human immunodeficiency virus type
1 (HIV-1) into the host cell genome, the expression of viral
genes is mediated by the cellular transcription machinery. In
addition, viral gene expression depends on the virally encoded
Tat protein, because a defect in Tat leads to a
replication-defective phenotype characterized by low-level synthesis of all
viral proteins (12, 17). The HIV-1 promoter is thought to be
regulated by attenuation (19, 22). According to this model,
transcription initiation in the absence of Tat results in the
accumulation of short transcripts (2, 16, 20, 26, 27, 43, 50)
generated by RNA polymerase II complexes with low intrinsic
processivity. Tat binding to the
trans
-activation responsive
re-gion (TAR) modifies the processivity of the transcription
com-plex, thus increasing the elongation rate, which in turn may
result in a greater promoter clearance, thereby indirectly
in-creasing the initiation rate (19).
Since Tat is not a virion-associated protein, it is present in
newly infected cells only after it is synthesized from the
inte-grated provirus. Therefore, basal HIV-1 promoter activity in
the absence of Tat should at least enable the synthesis of a
level of Tat sufficient to set up the positive feedback loop of
Tat
trans
-activation. In addition to the nonprocessive
transcrip-tion complex that is responsive to Tat, a second type of
tran-scription complex is proposed to assemble at the HIV-1 long
terminal repeat (LTR) promoter (28). This complex is
proces-sive, does not respond to Tat, and may be induced by cellular
transcription factors such as NF-
k
B and Sp1 (19). The
tran-scriptional activity of these complexes may be responsible for
raising the synthesis of viral mRNAs to a level that is sufficient
for the generation of effective amounts of Tat.
Most of the studies analyzing the HIV-1 promoter activity
used chimeric constructs in which the HIV-1 5
9
LTR was
ligated upstream of a reporter gene. In view of the high
moter activity in the presence of Tat, the basal HIV-1
pro-moter activity is assumed to be weak, but direct comparisons
between the basal activity of the HIV-1 promoter and other
viral or cellular promoters are inconsistent. This may be
par-tially due to the deduction of promoter activity from the
mea-surement of the activity of reporter gene products (10, 33, 46,
47).
Reporter gene constructs used in the analysis of Tat
trans
-activation contain the U3 region and sequences from the R
region including TAR. TAR differs from other transcriptional
regulatory elements in that it functions at the level of RNA, a
phenomenon that has thus far been observed only in
lentivi-ruses (11). Downstream of the transcription initiation site are
also located DNA regulatory elements that have been shown to
influence the transcription of various viral and cellular genes
(13, 15, 18, 21, 31, 49). Thus, Tax
1-independent transcription
from the human T-cell lymphotropic virus type 1 promoter
depends on the 45-bp downstream regulatory element 1 (DRE
1) located between nucleotides 202 and 246 downstream of the
transcription initiation site (21). Deletion of DRE 1 resulted in
a 10-fold decrease in basal chloramphenicol acetyltransferase
(CAT) reporter gene expression (21). Since DNA sequences
downstream of the transcription initiation site are
simulta-neously represented as RNA, they may contain
cis
-acting
reg-ulatory elements that function at the posttranscriptional level.
Sequences from nucleotides
2
3 to
1
77 of the herpes simplex
virus type 1 5
9
noncoding leader were able to increase the
stability of a hybrid CAT mRNA at least twofold in infected
* Corresponding author. Mailing address: Institut fu¨r Medizinische
Mikrobiologie und Virologie, Geb. 22.21, Universita¨tsstr. 1, D-40225
Du¨sseldorf, Germany. Phone: 12393. Fax:
(49-211)-81-12227. E-mail: [email protected].
2757
on November 9, 2019 by guest
http://jvi.asm.org/
cells (7). Moreover, the leader was responsive to
transcrip-tional activation by virus-specified transcription factors (7).
The present search for downstream regulatory elements in
HIV-1 was prompted by the findings of such elements in other
viruses and cellular genes and by previous findings that the
reporter genes used in the analysis of the HIV-1 promoter
activity were not precisely inserted into the position of an
HIV-1 open reading frame (ORF). As a consequence, the
reporter constructs contained only partial HIV-1 leader
se-quences and/or leader sese-quences derived from the reporter
gene. We therefore substituted the
tat
ORF in an authentic
tat
1.4 cDNA with that of CAT devoid of the bacterial leader (Fig.
1). The
tat
1.4 context was chosen because
tat
mRNAs belong
to the multiply spliced class of viral mRNAs that appear at the
very early stage of HIV-1 gene expression to generate the
trans
-activator Tat (23, 24). The exon 1-derived 106-bp leader
sequence downstream of the U5 region (lsdU5) that is present
in all HIV-1 transcripts was identified to be important for
mRNA and protein expression, irrespective of whether Tat
was present. A similar influence of exon 1 lsdU5 on
Tat-independent basal mRNA steady-state levels was observed
with transcripts in which the CAT reporter was again replaced
by authentic viral sequences containing point mutations that
inactivate the
tat
translation initiation codon. Measurement of
transcription rates showed that lsdU5 does not affect basal
HIV-1 promoter activity and indicated that exon 1 lsdU5
in-fluences posttranscriptional events, e.g., RNA processing,
ex-port, or stability.
MATERIALS AND METHODS
Oligonucleotides.Oligonucleotides were synthesized and purified as previ-ously described (39). Restriction sites used for cloning are underlined. The oligonucleotides are as follows: 302, 59-ACGGAATTCCGGATGAGC; 376, 59 -TGCTATGTCGACACCCAATTCAGTCGCCTCCCCTC; 377, 59-CTGAAGC GCGCACGGCAAGAGGCGAGGGGAGGCGACTG; 423, 59-GGGTGTCG ACATAGCAGAATAGGCGTTACTCGACAGAGGAGAGCAAGAAATGG AGAAAAAAAT; 426, 59-CGCGCAATTGGGTG; 427, 59-TCGACACCCAAT TG; 440, 59-AGTGGCGCCGCTCTCCTCTGTCGAGTA; 441, 59-AGTGGCG CCAAGAAATGGAGAAAAAAAT; 442, 59-ATTTTTTTCTCCATTTCTTGC GCCCGAACAGGGA; 447, 59-GAAATGGAGAAAAAAAT; 565, 59-GAAGC GCGCACGGCAAGAGGCGAGGGGAGGCGACTGCAAGAAATGGAGA AAAAAAT; 571, 59-AGTGGCGCCCGAACAGGGACTTGAAAGCGAAAG GGAAACCAGAGGAGATCTCTC; 572, 59-AGTGGCGCCCGAACAGGGA CTTGCCCGCGCCCGGGACACCAGAGGAGATCTCTC; 573, 59-GAAGCG CGCACGGCAAGATTCGATGTGATTCGACTGAATTGGGTG; and 574, 59-GAAGCGCGCACGGCAAGATTCGATGTGATTCGACTGCAAGAAATG. PCR.PCR amplification of DNA fragments used in cloning procedures was carried out with 2.5 U ofTaqDNA polymerase (Perkin-Elmer) and standard PCR reagents (38) for 30 cycles (30 s at 948C, 1 min at 508C, 1 min at 728C). Products were extracted with phenol and chloroform-isoamyl alcohol, ethanol precipitated, and resuspended in Tris-EDTA (TE) for further enzymatic diges-tion.
Recombinant plasmids.HIV sequences were derived from plasmid pNLA1 (48), which is a cDNA derivative of pNL4-3 (1). The nucleotide numbering system used is that of the parent sequence HIVNL43 (34). The CAT gene used for construction of the parent vector LTR1.4tatCAT was derived from pJS133
(45) as anEcoRI-XbaI fragment containing the CAT gene plus flanking regions. TheBspEI-MscI fragment within the CAT gene was substituted with the corre-sponding fragment of pDS56/RBSII 6xHis (kindly provided by D. Stu¨ber, Hoff-mann-La Roche, Basel, Switzerland), a CAT expression vector carrying a silent mutation in the internalEcoRI site (GAATTC to GAATTT). The resulting fragment was cloned into pGEM-Zf3(2) (Promega, Madison, Wis.), in which the multiple cloning site 39 toXbaI had been deleted, yielding EcoCATDE. For construction of LTR1.4tatCAT (Fig. 1), the following fragments were arranged between theEcoRV andBamHI sites of pSP73 (Promega) by using several subclones: theEcoRV (nt2343)-BssHII (nt1256) fragment of pNLA1; the HIV-1 sequencesBssHII-SalI (nt15330) obtained as double-stranded DNA by PCR with the overlapping oligonucleotides 376 and 377; theSalI-BspEI frag-ment, i.e., the junction of thetatmRNA leader and the CAT ORF generated by PCR with oligonucleotides 423 and 302; the BspEI-XbaI fragment of EcoCATDE; and theXbaI-BamHI fragment of pSVT7 (6) carrying the simian virus 40 (SV40) polyadenylation signal and the smalltintron sequence.
(i) Plasmid26/n.To reverse the orientation of lsdU5, two separate PCRs were run with primer pairs 440-442 and 447-302, generating two products with a 17-bp overlap. The products were extended by PCR and then used to replace the 400-bpNarI-BspEI fragment of LTR1.4tatCAT.
(ii) PlasmidDN/26.To delete the complete lsdU5, the 400-bp NarI (nt
1181)-BspEI fragment of LTR1.4tatCAT was replaced by a synthetic insert generated by PCR with primers 441 and 302.
(iii) PlasmidDsj1.4/26.The exon 4-derivedtatleader sequence was deleted by replacing the 326-bpBssHII-BspEI fragment of the parent vector with a synthetic insert generated by PCR with primers 565 and 302.
(iv) PlasmidDB/sj1.4.To delete 27 bp spanning from theBssHII site to splice junction 1.4, the 41-bpBssHII-SalI fragment of the parent vector was replaced by aBssHII-SalI linker consisting of annealed (20 min at 378C) oligonucleotides 426 and 427.
(v) PlasmidDn/B.PlasmidDn/B was constructed by cutting LTR1.4tatCAT with NarI andBssHII, joining nonhomologous ends with Xenopus laevisegg extract as previously described (40), and selecting a construct with a regenerated
BssHII site.
Point mutations in the DBF1 and/or the Sp1 binding sites were introduced by PCR into the parent vector and constructDsj1.4/26, using the primer combina-tion 572-302 for mutacombina-tion of the DBF1 site and the combinacombina-tions 573-302 and 574-302 for mutation of the Sp1 sites in LTR1.4tatCAT andDsj1.4/26, respec-tively. Double mutants were generated by replacing theKasI (nt1181)-BssHII fragments of the Sp1 single mutants with the PCR product generated with primer pair 572-302 recut withBssHII. The DBF1 site of isolate NL4-3 was replaced with that of isolate LAI by using PCR products generated with primer pair 571-302.
For the construction of LTR1.4Etat2, the 562-bpSalI-KpnI (nt15888) frag-ment of LTR1.4E was replaced with the corresponding insert of SpNLA1tat2in which aStuI site had been inserted into the first and second codons of thetat
ORF. For the construction of LTR1.4E from LTR-E/X-pA (40), the 529-bp
EcoRV-EcoRI (nt15288) fragment was replaced with the 1,460-bpEco
RV-EcoRI fragment of pNLA1, and this was followed by exchange of the 905-bp
BssHII-SalI fragment with a 41-bp double-stranded DNA generated by PCR with oligonucleotides 376 and 377. For plasmid LTR1.4Etat2Dn/B, the following fragments were ligated: theAflII (nt162)-KpnI vector fragment of LTR-E/X-pA, the AflII-SalI insert of LTRtatCATDn/B, and the SalI-KpnI insert of SpNLA1tat2.
All plasmid sequences whose construction involved synthetic oligonucleotides or PCR products were verified by automated DNA sequencing.
Cell culture and transfection.HeLa-T41cells (29) were propagated and transfected as described previously (39) with minor modifications. For electro-poration, 33106viable cells (trypan blue exclusion) in 200ml of detach medium (Eagle’s minimum essential medium supplemented with 10 mM sodium bicar-bonate and 25 mM HEPES) were mixed with CsCl gradient-purified plasmid DNA. The amount of DNA within cotransfection experiments was kept constant by adding pSVT7 as mock DNA. The transfection volume was adjusted to 350ml with electroporation medium (detach medium supplemented with 10% fetal calf serum). The cells were exposed to a single pulse with a setting of 550 V/cm and 960mF (Bio-Rad gene pulser with capacitance extender). The cells were kept at room temperature for 5 min before being transferred to culture flasks containing Dulbecco’s modified Eagle’s medium supplemented with 10% fetal calf serum and 2.5mg of amphotericin B per ml. When the cells were analyzed 48 h after transfection, the medium was changed 24 h after transfection.
CAT ELISA.Cells were lysed 48 h after transfection with 13reporter lysis buffer (Promega). Aliquots of the cell extracts were used for determination of the protein concentration (Bio-Rad). For monitoring transfection efficiency, the cells were cotransfected with 5mg of pGL3-control (Promega), a eucaryotic expres-sion vector carrying the firefly luciferase gene. Cell extracts were analyzed by the luciferase assay system (Promega) with a DCR-1 luminometer (DIGENE, Belts-ville, Md.). The cell extracts were then shock-frozen in liquid nitrogen and stored at2708C until thawed for quantification of CAT protein by a CAT enzyme-linked immunosorbent assay (ELISA) (Boehringer Mannheim).
Northern blot analysis.Total RNA was isolated 30 h after transfection by the single-step method (8), using RNA-clean (AGS, Heidelberg, Germany). For Northern blot analysis, the poly(A)1RNA of 75 to 110mg total RNA was FIG. 1. Schematic representation of the expression vector LTR1.4tatCAT
containing the HIV-1 59LTR, the cDNA of the authentic 1.4tatmRNA leader, the bacterial CAT ORF and the SV40 polyadenylation signal. ThetatORF (open arrow) was substituted with that of CAT. It is preceded by the lsdU5 containing the splice junction between exons 1 and 4 (sj1.4). Nucleotide26 (26) defines the upstream border of the translational start context. For construction of the lsdU5 series of constructs, the restriction sitesEcoRV (EV),NarI (N), andBssHII (B) were used (see Materials and Methods).
on November 9, 2019 by guest
http://jvi.asm.org/
isolated with Dynabeads oligo(dT)25(Dynal, Oslo, Norway), subjected to elec-trophoresis on a 1.5% agarose–formaldehyde gel, and blotted onto a positively charged nylon membrane (Boehringer Mannheim). After UV cross-linking, the membrane was hybridized with digoxigenin- (DIG) or 32P-labeled antisense RNA probes in H-SDS-buffer (50% formamide, 53SSC [13SSC is 0.15 M NaCl plus 0.015 M sodium citrate], 50 mM sodium phosphate, 0.1%N -lauroylsar-cosine, 7% sodium dodecyl sulfate [SDS], 2% blocking reagent, 50mg of yeast RNA per liter) at 688C. To monitor transfection efficiency and RNA loading, the membrane was hybridized with a DIG- or32P-labeled antisense RNA probe specific for exon 5 of human growth hormone (hGH) mRNA, which was syn-thesized from 5mg of the cotransfected plasmid pXGH5 (44). Hybridization signals were visualized by using an anti-DIG alkaline phosphatase conjugate (50 mU per ml; Boehringer Mannheim) and chemiluminescence (250mM CDP-Star; Tropix) or with a PhosphorImaging plate scanner (FUJIFILM BAS-1500). The chemiluminescent signals were quantified with a CSC camera (Cybertech, Berlin, Germany) and Raytest software TINA 2.08 (Straubenhardt, Germany), which was also used for quantifying radioactive signals. Additionally, both chemilumi-nescent and radiolabeled blots were exposed to specific films (ECL film and Hyperfilm MP; Amersham) and signals were scanned by laser densitometry (Molecular Dynamics). In all three quantification methods, the CAT mRNA signal intensities were normalized to that of the corresponding hGH signals.
Nuclear run-on transcription assay.Nuclei were isolated by Dounce homog-enization, using a protocol described by Bender (5), except that cells and nuclei were pelleted at 4003gand 2003g, respectively. For RNA synthesis, approx-imately 107nuclei in 200ml were mixed with an equal volume of 50 mM Tris-HCl (pH 7.5)–200 mM KCl–12 mM MgCl2–5 mM dithiothreitol–20 U of RNase inhibitor (Boehringer Mannheim)–1 mM each ATP, CTP, and GTP–100mCi of [a-32P]UTP (3,000 Ci/mmol; Amersham). After a 20-min incubation at 278C and an additional 15-min chase with 1ml of unlabeled 100 mM UTP, the RNA was extracted by consecutive 20-min incubations at 378C with DNase I (50 U; Boehr-inger Mannheim) and proteinase K (80mg; Boehringer Mannheim), followed by phenol-chloroform extraction and ammonium acetate-ethanol precipitation. Un-incorporated nucleotides were removed with Microcon-30 concentrators (Ami-con). Aliquots with equal radioactivity were hybridized with 5mg of immobilized CAT antisense or sense RNA in H-SDS-buffer at 508C. The hybridization of hGH mRNA, derived from the cotransfected plasmid pXGH5, served to nor-malize the CAT signals. The unlabeled CAT and hGH transcripts were gener-ated by in vitro transcription (Megascript Kit; Ambion) and immobilized onto positively charged nylon membranes (Boehringer Mannheim) with a slot blot
apparatus (Minifold II SRC 072/0, Schleicher & Schu¨ll). Hybridization signals were quantified with a PhosphorImaging Plate Scanner (FUJIFILM BAS-1500) and Raytest software TINA 2.08.
RESULTS
The exon 1 leader sequence downstream of U5 influences
basal, Tat-independent gene expression.
To analyze the role
of the
tat
leader sequence in basal, Tat-independent gene
expression, we constructed the eukaryotic expression vector
LTR1.4tatCAT, containing the HIV-1 5
9
LTR, the cDNA of
the authentic 1.4
tat
mRNA leader, the bacterial CAT ORF,
and the SV40 polyadenylation signal (see Materials and
Meth-ods) (Fig. 1). Mutations were introduced into this parent
vec-tor to analyze the leader sequence downstream of U5, lsdU5
(Fig. 2). In the following, the lsdU5 reaches from the
Nar
I site
(N) at the 3
9
end of U5 (nt
1
181) to a position 6 nt upstream
of the Tat translation initiation codon. The preservation of
these 6 nt and the fortuitous identity of the first two codons for
Tat and CAT leave the translation initiation signal (25) of the
transcripts unchanged. Construct
2
6/n contains the DNA
se-quence corresponding to lsdU5 in the opposite orientation,
with loss of the reversed
Nar
I restriction site (n). In
D
N/
2
6,
lsdU5 is deleted. In the other vectors, it is partially deleted,
using as dividing points the
BssH
II site (B) within the exon 1
sequence and the splice junction between exons 1 and 4 (sj1.4).
In
D
sj1.4/
2
6, the exon 4-specific leader sequence is deleted, in
D
B/sj1.4, the 3
9
27 nt of exon 1 (nt
1
262 to
1
288) is deleted,
and in
D
n/B, the 5
9
73 nt of lsdU5 (nt
1
183 to
1
255) is deleted.
Transfection of the parent
tat
leader construct (LTR1.4tat
CAT) into HeLa-T4
1cells resulted in efficient CAT
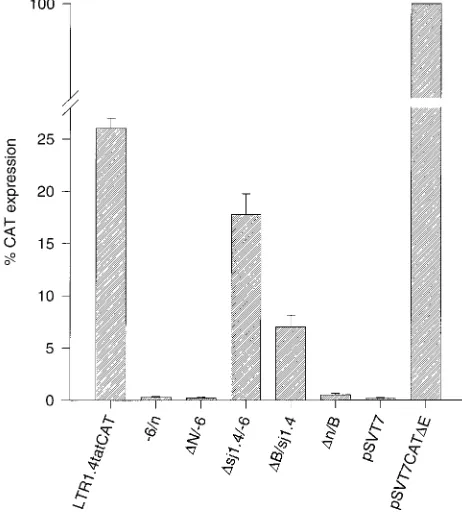
[image:3.612.71.283.67.373.2]expres-sion (Fig. 3). Even in the absence of Tat, the rate of CAT
synthesis was 26% of the rate obtained with the reference
FIG. 2. Schematic representation of the lsdU5 series of constructs. All des-ignations were as described in the legend to Fig. 1.
FIG. 3. Basal, Tat-independent CAT protein expression in HeLa-T41cells transfected with constructs of the lsdU5 series. HeLa-T41cells were transfected with 10mg of plasmid DNA, and CAT protein was quantified by an ELISA 48 h after transfection. CAT expression of pSVT7CATDE-transfected cells was set at 100%. Results are normalized to equal transfection efficiency and expressed as the mean of at least five independent transfections6standard error of the mean.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.319.550.420.678.2]vector pSVT7CAT
D
E, in which CAT expression is driven by
the SV40 early promoter. Deletion of the entire lsdU5 (
D
N/
2
6) reduced CAT expression to background levels. Among the
constructs with partial deletions of lsdU5, the most
pro-nounced reduction in CAT expression (52-fold) was observed
with construct
D
n/B, indicating that the lack of the exon 1
sequence of lsdU5 is mainly responsible for the loss of CAT
expression in construct
D
N/
2
6. Deletion of the 3
9
27 nt of exon
1 (
D
B/sj1.4) resulted in a moderate (3.7-fold) reduction of
CAT expression, whereas no significant decrease was observed
with the exon 4 deletion mutant
D
sj1.4/
2
6. Inversion of lsdU5
reduced CAT expression to background levels.
To differentiate between the possible causes for the
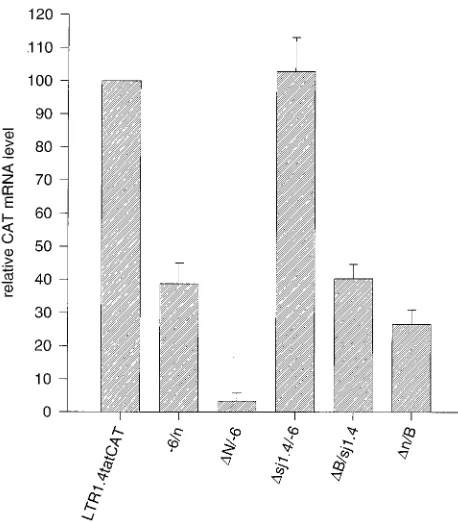
ob-served differences in CAT expression, as a first step Northern
blot analyses were done with the poly(A)
1RNA of HeLa-T4
1cells transfected with the lsdU5 series of constructs and a
CAT-specific probe (Fig. 4). The mRNA was quantified by
chemiluminescence or
32P-phosphorimaging in conjunction
with laser densitometry of exposed films (Fig. 5). The CAT
mRNA levels largely paralleled CAT protein expression. Thus,
the biggest drop in CAT protein expression with deletion of
lsdU5 (
D
N/
2
6) corresponded to a greater than 30-fold drop in
CAT mRNA; deletion of exon 4-derived leader sequences had
no significant effect on signal intensity; and deletion of the 3
9
and 5
9
regions of the exon 1-derived lsdU5 resulted in a two- to
fourfold decrease of CAT mRNA expression, respectively.
These findings with the lsdU5 deletion vectors support the
conclusion that exon 1-derived leader sequences are important
for the high Tat-independent activity of the parent vector
LTR1.4tatCAT.
Surprisingly, the mutant with the reverse-oriented leader
(
2
6/n) yielded significant amounts of mRNA, i.e., 39% of that
of the parent vector, even though CAT protein expression was
not detectable. The low level of protein expression conceivably
could be due to the presence of a fortuitous AUG start codon
upstream of the CAT AUG. This point was not further
pur-sued.
The lsdU5 dependence of mRNA levels is independent of Tat
trans
-activation of the LTR.
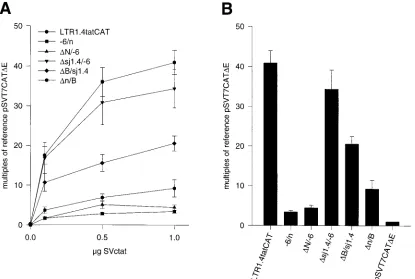
Constructs of the lsdU5 series
were cotransfected with increasing amounts (0.1, 0.5, and 1
m
g)
of the
tat
expression vector SVc
tat
, and CAT protein was
quantified after 48 h. The stimulation of CAT expression was
dependent on the quantity of cotransfected SVc
tat
,
approach-ing saturation levels with the largest quantity of SVc
tat
, i.e., 1.0
m
g (Fig. 6A). For constructs that had shown measurable
basal CAT expression, i.e., LTR1.4tatCAT,
D
sj1.4/
2
6, and
D
B/sj1.4 (cf. Fig. 3), maximal
trans
-activation was calculated
to be 157-, 194-, and 292-fold. Relative to the parent vector
LTR1.4tatCAT, the effect of deletions within the lsdU5 was
the same in the presence of maximum Tat stimulation as in
the absence of Tat (cf. Fig. 6B and Fig. 3). Thus, deletion of
the exon 4-derived
tat
leader (
D
sj1.4/
2
6) caused no
signif-icant change in CAT expression, whereas deletions of the 5
9
region of the lsdU5 (
D
n/B) and the short 3
9
end of exon 1
(
D
B/sj1.4) reduced CAT expression by 4.6- and 2-fold,
re-spectively. These findings indicate that the mechanism by
which lsdU5 sequences influence CAT expression is
inde-pendent from the Tat/TAR-mediated mechanism of
tran-scriptional activation.
Exon 1-derived lsdU5 mediates Tat-independent mRNA
ex-pression in the context of viral sequences.
It could be argued
that the high protein and mRNA levels observed with the
intact lsdU5 construct were due to the replacement of
[image:4.612.85.269.69.306.2]authen-FIG. 4. Northern blot analysis of mRNA isolated from HeLa-T41cells trans-fected with constructs of the lsdU5 series. Poly(A)1RNA isolated from HeLa-T41cells transfected with 10mg of plasmid DNA was subjected to formaldehyde-agarose gel electrophoresis and blotted onto a nylon membrane as described in Materials and Methods. (A) Blot probed with a DIG-labeled CAT-specific an-tisense RNA. (B) Reprobing of the same blot for hGH mRNA derived from the cotransfected plasmid pXGH5. The DIG-labeled probes were visualized by using an anti-DIG alkaline phosphatase conjugate and chemiluminescence. Lanes: 1, LTR1.4tatCAT; 2,26/n; 3,DN/26; 4,Dsj1.4/26; 5,DB/sj1.4; 6,Dn/B; 7, pSP73.
FIG. 5. Steady-state CAT mRNA levels in HeLa-T41cells transfected with constructs of the lsdU5 series. Northern blot analyses were quantified by chemi-luminescence or32P-phosphorimaging in conjunction with laser densitometry. hGH mRNA was used as a standard to normalize the CAT hybridization signals. CAT mRNA expression of LTR1.4tatCAT was set at 100%. Results are ex-pressed as the mean of five quantifications6standard error of the mean.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.320.549.413.674.2]tic viral sequences by the reporter ORF. To analyze
wheth-er lsdU5 also functions in the context of HIV-1 sequences,
HeLa-T4
1cells were transfected with expression vectors
LTR1.4E
tat
2and LTR1.4E
tat
2D
n/B. Translation of Tat from
the mRNAs derived from these plasmids is obviated by point
mutations in the translational start codon. To avoid
interfer-ence with transcription initiating at the 3
9
LTR (4), the SV40
polyadenylation signal used in the CAT constructs was
main-tained. As with the corresponding CAT transcripts, expression
of mRNAs containing the wild-type leader was high (Fig. 7,
lane 1) and almost undetectable with the exon 1 lsdU5 deletion
D
n/B (lane 2). In case of the wild-type leader construct, a
spliced 1.5-kb transcript and an unspliced 4-kb mRNA were
detected, indicating that functional Rev is translated from the
1.5-kb transcript. Furthermore, the unspliced mRNA was
translated into the viral Env glycoproteins, resulting in the
formation of syncytia (data not shown). Neither an unspliced
mRNA nor syncytia were observed with the lsdU5 deletion
mutant LTR1.4E
tat
2D
n/B. These results also confirm the
reg-ulatory function of exon 1 lsdU5 in the context of authentic
viral sequences.
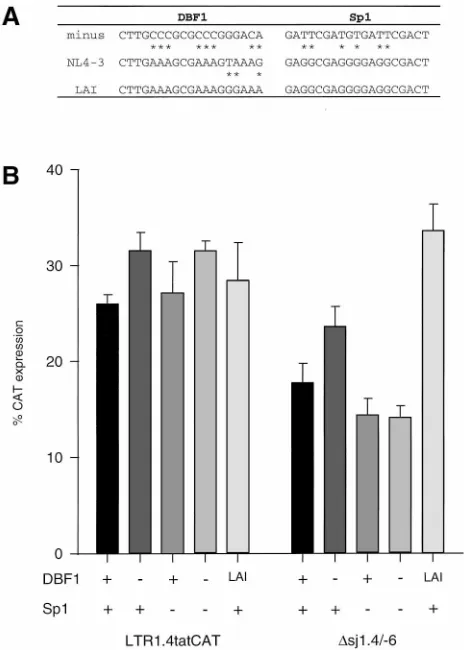
Reduction of CAT expression in lsdU5 deletion mutants is
not due to the loss of binding sites for nuclear factors within
lsdU5.
Within the exon 1-derived part of lsdU5, two binding
sites are localized for the constitutive nuclear factor Sp1 and
one is localized for the recently described downstream
bind-ing factor 1 DBF1 (15). These bindbind-ing sites have been
pro-posed to be involved in the regulation of the basal HIV-1
promoter activity. We therefore asked whether the loss of
these elements could be responsible for the lowered CAT
expression in the lsdU5 deletion mutants. Point mutations
which have been shown to abolish binding of both nuclear
factors (15) were introduced into the NL4-3-derived binding
sites of LTR1.4tatCAT and the exon 4-leader deletion mutant
D
sj1.4/
2
6 (Fig. 8A). Neither of the point mutations or their
combination significantly affected CAT protein expression,
ir-respective of the presence of the exon 4-derived
tat
leader
sequence (Fig. 8B). Thus, the decrease of CAT expression in
lsdU5 deletion mutants shown in Fig. 3 is not due to the loss of
Sp1 and/or DBF1 binding sites. The influence of Sp1 and
DBF1 binding sites on CAT expression under the present
experimental conditions is in contrast to the findings of el
Kharroubi and Verdin (15). While the HIV-1
LAI-derived Sp1
sites used by these authors are identical to the HIV-1
NL4-3-derived Sp1 sites analyzed in the present study, the DBF1 sites
from these isolates differ in three nucleotides (Fig. 8A). We
therefore substituted the DBF1 sites in LTR1.4tatCAT and in
D
sj1.4/
2
6 with that derived from HIV-1
LAI. An approximately
twofold stimulation of CAT expression was observed with the
exon 4 deletion mutant
D
sj1.4/
2
6 but not with the parent
vector LTR1.4tatCAT.
The lsdU5 does not influence the transcription rate.
The
correspondence between CAT expression and mRNA
steady-state levels expressed from the lsdU5 series of constructs could
be explained by two different mechanisms. lsdU5 could
func-tion as an element that is important for basal transcripfunc-tion or
as an RNA element that posttranscriptionally regulates RNA
polyadenylation, nucleocytoplasmic export, or stability. To
dis-tinguish among these possibilities, nuclear run-on transcription
assays were performed with nuclei isolated from cells
trans-fected with the lsdU5 constructs and the hGH expression
[image:5.612.101.516.70.349.2]vec-tor pXGH5. Incorporation of [
a
-
32P]UTP into preinitiated
FIG. 6. CAT protein expression in HeLa-T41cells transfected with constructs of the lsdU5 series and the Tat expression vector SVctat. (A) Cotransfection with increasing amounts (0.1, 0.5, and 1mg) of SVctat. (B) Cotransfection with 1mg of SVctat. CAT protein was quantified and results are normalized as described in the legend to Fig. 3. CAT expression of pSVT7CATDE-transfected cells was set at 1. Results are expressed as the mean of at least three independent transfections6
standard error of the mean.
on November 9, 2019 by guest
http://jvi.asm.org/
transcripts was determined by hybridization with CAT and
hGH antisense RNA immobilized onto nylon membranes. The
CAT hybridization signals were quantified and normalized to
the corresponding hGH signals. The observed differences
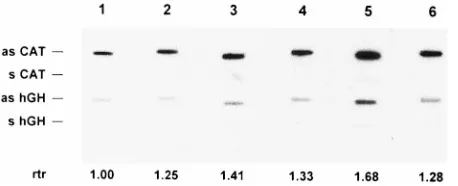
be-tween the relative transcription rates of nuclei containing
wild-type or mutant lsdU5 plasmids were less than twofold (Fig. 9).
This finding excludes the possibility that the observed influence
of exon 1 lsdU5 sequences on the steady-state mRNA level is
due to their action as transcriptional enhancers. Conversely, it
supports the conclusion that exon 1-derived lsdU5 sequences
in the RNAs function in processing, nuclear export, or
stabi-lization of the transcripts.
DISCUSSION
In the absence of the
trans
-activator Tat, the HIV-1
pro-moter is thought to possess a low basal activity. The
demon-stration of stimulatory elements downstream of the
transcrip-tion initiatranscrip-tion site in other systems led us to ask whether
sequences downstream of TAR in HIV-1 that influence the
basal promoter activity could be identified. For this reason, the
CAT reporter plasmid LTR1.4tatCAT, containing the coding
region for the entire authentic HIV-1
tat
1.4 mRNA leader
sequence downstream of the HIV-1 U3 region, was
con-structed. The reporter gene was devoid of bacterial leader
sequences and inserted precisely into the native
tat
ORF.
Sur-prisingly, CAT protein expression with this parent construct
was rather efficient, i.e., 26% of that of the reference construct
in which the CAT gene is under the control of the SV40 early
promoter. When lsdU5 was completely deleted, no CAT
pro-tein could be detected. The basal CAT expression of both
constructs was paralleled by mRNA steady-state levels.
For further localization of elements responsible for the
ef-ficient expression with the wild-type leader, the lsdU5 element
was partially deleted. The high basal reporter expression
ob-served with LTR1.4tatCAT could have been due to the
gen-eration of an artificial DNA enhancer by joining exon 1 and
exon 4 in the cDNA. The exon 4 deletion mutant
D
sj1.4/
2
6
revealed that the element responsible for efficient expression
does not overlap the splice junction. The partial deletion
mu-tants allowed the identification of an exon 1-derived 106-bp
element located between U5 and the major splice donor as
being responsible for high basal CAT expression.
Since an enhancer can function in either orientation relative
to the promoter, the leader mutant
2
6/n was constructed to
analyze the orientation dependence of the lsdU5 element.
Re-versing the orientation of the lsdU5 element reduced the
mRNA steady-state level, but not to the baseline level
ob-served with
D
N/
2
6, indicating a partial orientation
depen-dence of the lsdU5 element.
[image:6.612.319.551.69.394.2]The influence of lsdU5 on the mRNA steady-state level was
confirmed in transcripts with authentic HIV-1 sequences
down-stream of the leader sequence. A drastic decrease in mRNA
expression was again observed when most of the exon 1 portion
of lsdU5 was deleted. It should be emphasized that the high
level of mRNA containing the wild-type leader was generated
in the absence of Tat and that, furthermore, functional viral
FIG. 7. Northern blot analysis of mRNA isolated from HeLa-T41cells trans-fected with constructs LTR1.4Etat2 and LTR1.4Etat2Dn/B. Poly(A)1RNA isolated from HeLa-T41cells transfected with 20 mg of plasmid DNA was subjected to formaldehyde-agarose gel electrophoresis and blotted onto a nylon membrane as described in Materials and Methods. (A) The blot was probed simultaneously with DIG-labeled antisense RNAs corresponding to env se-quences from nt 8648 to 8887 and to exon 5 sese-quences in hGH mRNA. Arrows indicate the unspliced (us) and spliced (s) mRNAs. (B) Longer exposure of the unspliced transcript signal. The DIG-labeled probes were visualized as described in legend to Fig. 4. Lanes: 1, LTR1.4Etat2; 2, LTR1.4Etat2Dn/B; 3, pSP73.
FIG. 8. Effect of mutations of the DBF1 and Sp1 binding sites on the basal CAT expression in HeLa-T41cells transfected with LTR1.4tatCAT and deletion mutantDsj1.4/26. (A) Nucleotide sequences of the DBF1 and Sp1 binding sites of HIV-1 isolates NL4-3 and LAI and of mutated binding sites that have been shown to lack binding activity (15). (B) Basal, Tat-independent CAT protein expression in HeLa-T41cells transfected with 10mg of plasmids containing the DBF1 site of virus isolate NL4-3 (1) or LAI (LAI) and the Sp1 sites of NL4-3 (1), which is identical to that of LAI. Derivatives of these plasmids contained the mutated binding sites (2) as indicated in panel A. CAT protein was quantified and results are normalized and expressed as described in the legend to Fig. 3, except that only three independent experiments were evaluated.
on November 9, 2019 by guest
http://jvi.asm.org/
proteins were translated from these transcripts. Since Tat is
essential for viral replication (12, 17), it can be postulated that
the observed influence of lsdU5 may be superimposed in an
HIV-1-infected cell with counteracting regulatory effects
me-diated by Nef or by elements in the viral 3
9
LTR that are absent
from the analyzed transcripts.
Measurement of transcription rates by nuclear run-on
showed no significant difference between the transcriptional
activities of the wild-type and mutant lsdU5 constructs,
indi-cating that in this system, lsdU5 does not function as a DNA or
RNA element to enhance basal promoter activity.
Addition-ally, the results argue that the HIV-1 promoter is fairly active
in HeLa-T4
1cells and able to synthesize full-length
tran-scripts, even in the absence of Tat.
Sequences within the exon 1 portion of the lsdU5 element
have been reported to contain binding sites for the nuclear
factors DBF1 and Sp1 and to be important for basal HIV-1
promoter activity (15). In that study, mutation of the DBF1 site
alone or in conjunction with a mutation of the Sp1 sites
re-sulted in a twofold and fourfold decrease in CAT activity,
respectively. Since in the present study, deletion of all three
binding sites (
D
N/
2
6) did not reduce transcription and point
mutations in the binding sites did not affect gene expression, an
influence of these downstream elements on promoter activity
can be excluded in our system. In an attempt to resolve the
apparent discrepancy between these two sets of results, we
exchanged the DBF1 site in our constructs, which is derived
from isolate NL4-3, with that of isolate LAI (15). Interestingly,
this exchange resulted in an approximately twofold increase in
CAT protein expression only in mutant
D
sj1.4/
2
6, which is
lacking exon 4 leader sequences, as are the constructs used by
el Kharroubi and Verdin (15). In a recent report, point
muta-tions of individual binding sites in transient transfecmuta-tions did
not significantly affect promoter activity (14). This could be in
agreement with our results if the DBF1 site analyzed in that
study had been derived from isolate NL4-3.
The role of lsdU5 as an element that posttranscriptionally
influences the mRNA steady-state level is in good agreement
with the observation that lsdU5 acts independently of Tat
trans
-activation of the HIV-1 promoter. The basal CAT
ex-pression pattern of the lsdU5 series of plasmids persisted when
Tat protein was supplied in
trans
. It is unlikely that the
differ-ent CAT levels, which in the presence of Tat amount to 1 order
of magnitude, actually reflect different
trans
-activation
mech-anisms among wild-type and mutated
tat
leaders, because every
plasmid of the lsdU5 series contains an identical intact TAR
element and mutations are far downstream of TAR. The
ob-served pattern of CAT expression among the constructs is
more likely to be a consequence of posttranscriptional events
influenced by exon 1 lsdU5 and unaffected by the presence of
Tat.
The cytoplasmic expression of a subset of HIV mRNAs
encoding structural as well as some accessory proteins is
reg-ulated at the posttranscriptional level.
cis
-acting inhibitory
se-quences (INS, IR, or CRS) contained in the
gag
,
pol
,
env
, and
vif
coding regions (9, 30, 32, 35–37, 41, 42) were shown to trap
these mRNAs in the nucleus (30), to decrease mRNA stability
(42), or to prevent translation (9). lsdU5 differs from all of
these sequences in that it positively affects the mRNA
steady-state level. Furthermore, it should be noted that in contrast to
the inhibitory sequences, exon 1 lsdU5 is present in all HIV-1
transcripts.
Among the posttranscriptional events that might be
influ-enced by exon 1 lsdU5, i.e., RNA processing, transport, or
stability, the last deserves serious attention because of recent
findings by Agy et al. (3), who demonstrated that both a
cel-lular mRNA and an RNA containing the first 111 nt of the
HIV-1 leader sequence upstream of the CAT gene are more
rapidly degraded in HIV-1-infected cells than in uninfected or
simian immunodeficiency virus-infected cells. In contrast, an
RNA containing the entire HIV-1 leader sequence and
extend-ing through nt 684 of the
gag
coding region was more stable
than the cellular
actin
mRNA (3). In view of these results, it
could be envisioned that due to its presence in all HIV-1
transcripts, the exon 1-derived lsdU5 may confer selective
re-sistance to virus-induced degradation of mRNAs.
The results presented here identified the exon-1 lsdU5
ele-ment, present in all viral transcripts, as being important for
posttranscriptional events in HIV-1 gene expression. The role
of lsdU5 in viral RNA processing, transport, and stability is
currently under investigation. Furthermore, it would be of
in-terest to determine whether alternative leader sequences,
re-sulting from incomplete splicing of the small noncoding exons
2 and 3, may also influence HIV-1 gene expression at the
posttranscriptional level.
ACKNOWLEDGMENTS
We thank J. M. Smith and D. Stu¨ber for providing plasmids pJS133
and pDS56/RBSII 6xHis, respectively. We thank Martina Austrup and
Maria Thieme for excellent technical assistance.
This work was supported by the Bundesminister fu¨r Forschung und
Technologie (FKZ II-023-86) and by grants from the
Kinder-AIDS-Hilfe Deutschland e.V. and from the Heinz-Ansmann-Stiftung.
REFERENCES
1.Adachi, A., H. E. Gendelman, S. Koenig, T. Folks, R. Willey, A. Rabson, and M. A. Martin.1986. Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. J. Virol.59:284–291.
2.Adams, M., L. Sharmeen, J. Kimpton, J. M. Romeo, J. V. Garcia, B. M. Peterlin, M. Groudine, and M. Emerman.1994. Cellular latency in human immunodeficiency virus-infected individuals with high CD4 levels can be detected by the presence of promoter-proximal transcripts. Proc. Natl. Acad. Sci. USA91:3862–3866.
3.Agy, M. B., C. H. Sherbert, and M. G. Katze.1996. Development of an in vitro mRNA degradation assay utilizing extracts from HIV-1- and SIV-infected cells. Virology217:158–166.
4.Ashe, M. P., P. Griffin, W. James, and N. J. Proudfoot.1995. Poly(A) site selection in the HIV-1 provirus: inhibition of promoter-proximal polyade-nylation by the downstream major splice donor site. Genes Dev.9:3008– 3025.
[image:7.612.66.295.71.164.2]5.Bender, T. P.1990. Identification of newly transcribed RNA, p. 4.10.5– 4.10.10.InF. M. Ausubel, R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl (ed.), Current protocols in molecular biology. Greene Publishing Associates and Wiley Interscience, New York, N.Y.
FIG. 9. Transcription rates in nuclei isolated from cells transfected with the lsdU5 plasmids as determined by nuclear run-on transcription. The incorpora-tion of [a-32P]UTP into elongated CAT transcripts was determined by hybrid-ization to 5mg of CAT antisense RNA (as CAT) immobilized onto nylon membranes. Hybridization of hGH mRNA derived from the cotransfected plas-mid pXGH5 to 5mg of hGH antisense RNA (as hGH) was used to normalize the CAT signals. CAT (s CAT) and hGH (s hGH) sense RNA served as controls. The relative transcription rates (rtr) were obtained by32P-phosphorimaging and are corrected for the uridine content of the CAT transcripts. Lanes: 1, LTR1.4tatCAT; 2,26/n; 3,DN/26; 4,Dsj1.4/26; 5,DB/sj1.4; 6,Dn/B.
on November 9, 2019 by guest
http://jvi.asm.org/
6.Bird, P., M. J. Gething, and J. Sambrook.1987. Translocation in yeast and mammalian cells: not all signal sequences are functionally equivalent. J. Cell Biol.105:2905–2914.
7.Blair, E. D., C. C. Blair, and E. K. Wagner.1987. Herpes simplex virus virion stimulatory protein mRNA leader contains sequence elements which in-crease both virus-induced transcription and mRNA stability. J. Virol.61: 2499–2508.
8.Chomczynski, P., and N. Sacchi.1987. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Bio-chem.162:156–159.
9.Cochrane, A. W., K. S. Jones, S. Beidas, P. J. Dillon, A. M. Skalka, and C. A. Rosen. 1991. Identification and characterization of intragenic sequences which repress human immunodeficiency virus structural gene expression. J. Virol.65:5305–5313.
10. Cullen, B. R.1986.trans-activation of human immunodeficiency virus occurs via a bimodal mechanism. Cell46:973–982.
11. Cullen, B. R.1993. Does HIV-1 Tat induce a change in viral initiation rights? Cell73:417–420.
12. Dayton, A. I., J. G. Sodroski, C. A. Rosen, W. C. Goh, and W. A. Haseltine. 1986. Thetrans-activator gene of the human T cell lymphotropic virus type III is required for replication. Cell44:941–947.
13. Derse, D., and J. W. Casey.1986. Two elements in the bovine leukemia virus long terminal repeat that regulate gene expression. Science231:1437–1440. 14. el Kharroubi, A., and M. A. Martin.1996.cis-acting sequences located downstream of the human immunodeficiency virus type 1 promoter affect its chromatin structure and transcriptional activity. Mol. Cell. Biol.16:2958– 2966.
15. el Kharroubi, A., and E. Verdin.1994. Protein-DNA interactions within DNase I-hypersensitive sites located downstream of the HIV-1 promoter. J. Biol. Chem.269:19916–19924.
16. Feinberg, M. B., D. Baltimore, and A. D. Frankel.1991. The role of Tat in the human immunodeficiency virus life cycle indicates a primary effect on transcriptional elongation. Proc. Natl. Acad. Sci. USA88:4045–4049. 17. Fisher, A. G., M. B. Feinberg, S. F. Josephs, M. E. Harper, L. M. Marselle,
G. Reyes, M. A. Gonda, A. Aldovini, C. Debouk, R. C. Gallo, et al.1986. The
trans-activator gene of HTLV-III is essential for virus replication. Nature 320:367–371.
18. Hultmark, D., R. Klemenz, and W. J. Gehring.1986. Translational and transcriptional control elements in the untranslated leader of the heat-shock genehsp22. Cell44:429–438.
19. Jones, K. A., and B. M. Peterlin.1994. Control of RNA initiation and elongation at the HIV-1 promoter. Annu. Rev. Biochem.63:717–743. 20. Kao, S. Y., A. F. Calman, P. A. Luciw, and B. M. Peterlin.1987.
Anti-termination of transcription within the long terminal repeat of HIV-1 bytat
gene product. Nature330:489–493.
21. Kashanchi, F., J. F. Duvall, P. F. Lindholm, M. F. Radonovich, and J. N. Brady.1993. Sequences downstream of the RNA initiation site regulate human T-cell lymphotropic virus type I basal gene expression. J. Virol. 67:2894–2902.
22. Kessler, M., and M. B. Mathews.1992. Premature termination and process-ing of human immunodeficiency virus type 1-promoted transcripts. J. Virol. 66:4488–4496.
23. Kim, S. Y., R. Byrn, J. Groopman, and D. Baltimore.1989. Temporal aspects of DNA and RNA synthesis during human immunodeficiency virus infection: evidence for differential gene expression. J. Virol.63:3708–3713. 24. Klotman, M. E., S. Kim, A. Buchbinder, A. DeRossi, D. Baltimore, and F.
Wong Staal.1991. Kinetics of expression of multiply spliced RNA in early human immunodeficiency virus type 1 infection of lymphocytes and mono-cytes. Proc. Natl. Acad. Sci. USA88:5011–5015. (Erratum,89:1148, 1992.) 25. Kozak, M.1987. An analysis of 59-noncoding sequences from 699 vertebrate
messenger RNAs. Nucleic Acids Res.15:8125–8148.
26. Laspia, M. F., A. P. Rice, and M. B. Mathews.1989. HIV-1 Tat protein increases transcriptional initiation and stabilizes elongation. Cell59:283–292. 27. Laspia, M. F., A. P. Rice, and M. B. Mathews.1990. Synergy between HIV-1 Tat and adenovirus E1A is principally due to stabilization of transcriptional elongation. Genes Dev.4:2397–2408.
28. Lu, X., T. M. Welsh, and B. M. Peterlin.1993. The human immunodeficiency virus type 1 long terminal repeat specifies two different transcription com-plexes, only one of which is regulated by Tat. J. Virol.67:1752–1760. 29. Maddon, P. J., A. G. Dalgleish, J. S. McDougal, P. R. Clapham, R. A. Weiss,
and R. Axel.1986. The T4 gene encodes the AIDS virus receptor and is
expressed in the immune system and the brain. Cell47:333–348. 30. Maldarelli, F., M. A. Martin, and K. Strebel.1991. Identification of
post-transcriptionally active inhibitory sequences in human immunodeficiency virus type 1 RNA: novel level of gene regulation. J. Virol.65:5732–5743. 31. Mansour, S. L., T. Grodzicker, and R. Tjian.1986. Downstream sequences
affect transcription initiation from the adenovirus major late promoter. Mol. Cell. Biol.6:2684–2694.
32. Mikaelian, I., M. Krieg, M. J. Gait, and J. Karn.1996. Interactions of INS (CRS) elements and the splicing machinery regulate the production of Rev-responsive mRNAs. J. Mol. Biol.257:246–264.
33. Muesing, M. A., D. H. Smith, and D. J. Capon.1987. Regulation of mRNA accumulation by a human immunodeficiency virustrans-activator protein. Cell48:691–701.
34. Myers, G., B. Korber, B. H. Hahn, K. T. Jeang, J. W. Mellors, F. E. Mc-Cutchan, L. E. Henderson, and G. N. Pavlakis.1995. Human retroviruses and AIDS. A compilation and analysis of nucleic acid and amino acid sequences. Theoretical Biology and Biophysics Group, Los Alamos National Laboratory, Los Alamos, N.M.
35. Nasioulas, G., A. S. Zolotukhin, C. Tabernero, L. Solomin, C. P. Cunning-ham, G. N. Pavlakis, and B. K. Felber.1994. Elements distinct from human immunodeficiency virus type 1 splice sites are responsible for the Rev de-pendence ofenvmRNA. J. Virol.68:2986–2993.
36. Reddy, T. R., G. Kraus, M. Suhasini, M. C. Leavitt, and F. Wong Staal.1995. Identification and mapping of inhibitory sequences in the human immuno-deficiency virus type 2vifgene. J. Virol.69:5167–5170.
37. Rosen, C. A., E. Terwilliger, A. Dayton, J. G. Sodroski, and W. A. Haseltine. 1988. Intrageniccis-actingartgene-responsive sequences of the human im-munodeficiency virus. Proc. Natl. Acad. Sci. USA85:2071–2075. 38. Saiki, R. K., D. H. Gelfand, S. Stoffel, S. J. Scharf, R. Higuchi, G. T. Horn,
K. B. Mullis, and H. A. Erlich.1988. Primer-directed enzymatic amplifica-tion of DNA with a thermostable DNA polymerase. Science239:487–491. 39. Schaal, H., M. Klein, P. Gehrmann, O. Adams, and A. Scheid.1995.
Re-quirement of N-terminal amino acid residues of gp41 for human immuno-deficiency virus type 1-mediated cell fusion. J. Virol.69:3308–3314. 40. Schaal, H., P. Pfeiffer, M. Klein, P. Gehrmann, and A. Scheid.1993. Use of
DNA end joining activity of a Xenopus laevis egg extract for construction of deletions and expression vectors for HIV-1 Tat and Rev proteins. Gene 124:275–280.
41. Schwartz, S., M. Campbell, G. Nasioulas, J. Harrison, B. K. Felber, and G. N. Pavlakis.1992. Mutational inactivation of an inhibitory sequence in human immunodeficiency virus type 1 results in Rev-independentgag ex-pression. J. Virol.66:7176–7182.
42. Schwartz, S., B. K. Felber, and G. N. Pavlakis.1992. Distinct RNA se-quences in thegagregion of human immunodeficiency virus type 1 decrease RNA stability and inhibit expression in the absence of Rev protein. J. Virol. 66:150–159.
43. Selby, M. J., E. S. Bain, P. A. Luciw, and B. M. Peterlin.1989. Structure, sequence, and position of the stem-loop in tar determine transcriptional elongation by tat through the HIV-1 long terminal repeat. Genes Dev. 3:547–558.
44. Selden, R. F., K. B. Howie, M. E. Rowe, H. M. Goodman, and D. D. Moore. 1986. Human growth hormone as a reporter gene in regulation studies employing transient gene expression. Mol. Cell. Biol.6:3173–3179. 45. Shiau, A., and J. M. Smith.1988. Improved cat gene cassette for promoter
analysis and genetic constructions. Gene67:295–299.
46. Sodroski, J., R. Patarca, C. Rosen, F. Wong Staal, and W. Haseltine.1985. Location of the trans-activating region on the genome of human T-cell lymphotropic virus type III. Science229:74–77.
47. Sodroski, J., C. Rosen, F. Wong Staal, S. Z. Salahuddin, M. Popovic, S. Arya, R. C. Gallo, and W. A. Haseltine.1985.trans-acting transcriptional regula-tion of human T-cell leukemia virus type III long terminal repeat. Science 227:171–173.
48.Strebel, K., D. Daugherty, K. Clouse, D. Cohen, T. Folks, and M. A. Martin. 1987. The HIV ’A’ (sor) gene product is essential for virus infectivity. Nature 328:728–730.
49.Theill, L. E., O. Wiborg, and J. Vuust.1987. Cell-specific expression of the human gastrin gene: evidence for a control element located downstream of the TATA box. Mol. Cell. Biol.7:4329–4336.
50.Toohey, M. G., and K. A. Jones.1989. In vitro formation of short RNA polymerase II transcripts that terminate within the HIV-1 and HIV-2 pro-moter-proximal downstream regions. Genes Dev.3:265–282.