JOURNAL OF VIROLOGY,Sept.1979, p. 608-620 Vol. 31, No.3
0022-538X/79/09-0608/13$02.00
Structural
Analysis
of Precursor
and Product
Forms
of
Type-Common
Envelope Glycoprotein
D
(CP-1 Antigen)
of
Herpes
Simplex
Virus
Type
1
ROSELYN J. EISENBERG,I* CASSANDRAHYDREAN-STERN,'ANDGARY H.COHEN2
DepartmentofPathobiology, Schoolof VeterinaryMedicine,'andDepartment ofMicrobiologyand Centerfor
Oral HealthResearch,SchoolofDentalMedicine,2 University ofPennsylvania, Philadelphia,Pennsylvania
19104
Received forpublication16February 1979
The type-common CP-1 antigen of herpes simplex virus type 1 (HSV-1) is associated in theinfected cell withtwocomponents, a 52,000-molecular-weight glycoprotein (gp52 orpD) and a 59,000-molecular-weight glycoprotein (gp59or D). Thelarger form (D) is also foundinthe virion envelope. It waspostulated thatpDisa precursorof D. Wefound that pD sharedmethionine and arginine tryptic peptides with D isolated from infected cell extracts. D isolated from infectedextracts hadthe sametryptic methioninepeptide profileasD isolated from the virionenvelope. Thus,processing of pDtoD doesnotinvolve anymajor alterationsinpolypeptidestructure.Furthermore,Ddidnotsharetryptic methi-oninepeptides with the other majorglycoproteins of HSV-1. Using [2-3H]mannose as a specific glycoprotein label, we found that pD, which is a basic protein (isoelectric point =8.0) contained a 1,800-molecular-weight oligomannosyl core moiety andwasprocessed by further glycosylationandsialyationtoa moreacidic andheterogeneous molecule D, which hasamolecularweightofatleast59,000. Herpes simplex virus type 1
(HSV-1)
glyco-proteins are present in infected cells in both a precursorandproduct form (2,7,19). The major glycosylated proteinsarederived from four an-tigenically distinct polypeptides (19). The ma-tureglycoproteins havebeendesignated A, B,C, and D, and theprecursors havebeendesignated pA, pB,pC, and pD by Spear (19). This termi-nology was accepted provisionally at the 1978 HerpesWorkshopatCambridge, England. Proc-essing ofthese glycoproteins from a lower-mo-lecular-weight precursor to the higher-molecu-lar-weight product doesnotrequire protein syn-thesis (19), and the modification of each mole-cule apparently involves several glycosylation steps(2, 19).
The virionenvelope contains several glycopro-teins (2, 7, 19), one component atapproximately 59,000 daltons, and a,,30,000-molecular-weight complex (gpl30) containing two or three com-ponents. Previously,wedescribed an HSV-spe-cificantigen, CP-1 (2, 3), purified from HSV-1-infected cells and having the properties of a glycoprotein. This glycoprotein stimulated the productionof high titers of type-common neu-tralizingantibody. Our recent study (2) showed that the type-common CP-1 antigenic activity wasassociated in infectedcellswithtwo molec-ular weight components, a
52,000-molecular-weight glycoprotein (gp52) anda 59,000-molec-ular-weight glycoprotein (gp59). We presented evidence that gp52 is theprecursorofgp59,the latter being foundin the infected cellextract as well as in the purified virion. We believethat theprecursor,gp52,correspondstopDand that the product, gp59, corresponds to D. In this paper, weshall adopt the Cambridge terminol-ogy and referto the two forms of CP-1 as pD andD.
The present study was undertaken to docu-mentthesynthesis and processing of the HSV type-commonglycoproteinD. Theevidence pre-sented here demonstrates that pD shares me-thionine and arginine tryptic peptides with D. The firstdetectable precursor, pD, contains an 1,800-molecular-weightoligomannosylcore moi-ety and is basic in nature (isoelectric point = 8.0). Furthermore, pD is processed by further glycosylation and sialyation to a more acidic and heterogeneous molecule, D, which has a molec-ularweightof at least 59,000. Finally, D does not appear to share any methionine peptides with the gpl3O complex extracted from the virion.
MATERIALS AND
METHODS
Ceilcultures.Conditions for the growth and
main-tenanceof KB cells and baby hamster kidney cells havebeen previously described(3).
608
on November 10, 2019 by guest
http://jvi.asm.org/
STRUCTURE OF GLYCOPROTEIN D (CP-1) OF HSV-1 609
Virus preparation and titration. The procedures used for the preparation of virus stocks of HSV-1 (strainHF), as well as the plaque assay, were described previously (3). For infection, unless otherwise noted, an input multiplicity of 20 PFU of HSV per cell was employed.
Virus purification. HSV-1 was purified by the method of Spear and Roizman (20). For radioactive labeling of virus 10 ,uCi of [3S]methionine (specific activity, >200 Ci/mmol) per ml, 30 uCi of [methyl-3H]methionine (specific activity,9Ci/mmol) perml, or10,uCi of[2-3H]mannose (specific activity, 18Ci/
mmol) per ml was added at5 hpostinfection (p.i.). Cellswereharvested for viruspurificationat18hp.i. Tosolubilize the HSVenvelope,purified virionswere
suspended in0.02 MTris buffer (pH 7.5) containing
0.05 M NaCl and 0.5% Nonidet P-40 (NP40) and incubated at37°C for45min.TPCK (L-1-tosylamide-2-phenethyl chloromethyl ketone) and TLCK (N-a-p-tosyl-L-lysine chloromethyl ketone hydrochloride) were added, each at a concentration of0.1 mM, to
inhibit proteolytic activity. Nucleocapsids were
re-movedby centrifugationat 100,000 x g for1 h.The 100,000-x-g pellet was reextracted with NP-40 and recentrifuged. The supernatants from both extractions werepooled and frozenat-70°C.
Pulse-chase experiments. Pulse-chase
experi-ments were performed on KB cell cultures (60-mm plates containing label in 1.5 ml of Hanks salts) essen-tially by thesamemethodasdescribedpreviously (2). The following labels and total amounts were used: [35S]methionine (specific activity, 503 Ci/mmol), 200 ,uCi;[3H]methionine (specific activity, 9 Ci/mmol), 1.0 mCi; [U-_4C]arginine (specific activity, 340 mCi/
mmol),125ytCi;[3H]arginine (specific activity,22Ci/
mmol),1mCi;[2-3H]mannose (specific activity,18Ci/
mmol),1.0mCi; and[U-'4C]glucosamine (specific
ac-tivity,238mCi/mmol), 100PiCi.The pulse period for mannoseand glucosamine labeling was 30 min, and that for methionine andargininewas 15min.After the pulse and chase, the cells were lysed as described previously (2), except that TPCK and TLCK (0.1 mM)
were added to all solutions and all glassware was
sterilizedtominimizeproteolysis.
Immunological procedures.The antisera used in thisstudy have been described previously (2, 3). Anti-CP-1 serum wasemployed as amonoprecipitin anti-body against pD and D components. Anti-ENV-1 se-rum (previously designated anti-HSV-ENV serum
[2]) prepared against the NP40 extract of the virion envelope ofHSV-1was employedtoprecipitate the envelope glycoproteins.
Immune precipitations were carried out as previ-ously described (2), except that: (i)0.1mMTPCK and
0.1 mMTLCKwereaddedto themixtures, and (ii)
themixtureswere maintained at4°Cfor 2days.
ElectrophoresisonSDS-PAGE. Sodiumdodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was carried out in slabs of 10% acrylamide cross-linked with 0.26%N,N'-methylenebisacrylamide
(BIS) as describedpreviously (2) or inslabs of 10%
acrylamide cross-linked with0.4%
N-N'-diallyltartar-diamide (DATD) essentially as described by Spear
(19). Afterelectrophoresis, thegelswerestainedwith Coomassiebrilliant blue as describedpreviously (2).
For autoradiography, the gels were dried on filter paper and placed in contact with Kodak X-Omat R (XR-5) film. For fluorography, thegels were impreg-nated with 2,5-diphenyloxazole (1) placed in contact
with Kodak X-Omat R film and stored at -70°C. Protein standards ranging from 15,000 to 130,000 dal-tons were run on eachgel (2). Two-dimensional gel electrophoresis was performed by the technique of O'Farrell (15).
Preparation of samples for tryptic peptide analysis.Proteins wereeluted fromBIS cross-linked gels and prepared for trypsinization by theprocedure ofVogtetal. (23). DATD cross-linkedgel sliceswere
dissolvedin 2ml of 2%periodic acid per fourtofive slicesby the procedure of Gibson (6). Both methods gaveidentical tryptic methionine peptide profiles for themajor capsid protein ofHSV-1(M. Ponce de Leon, G. H.Cohen, andR.J.Eisenberg,unpublished data). For DATDgels, bovineserumalbuminwasaddedas aproteincarrier (1 mg per fourtosixgel slices), and the proteinswereprecipitated with25%trichloroacetic acidat4°C for12to 18h.Precipitateswerecollected bycentrifugation at 100,000xg for2 h(W. Gibson, personalcommunication), dissolved in 0.1 NNaOH,
andprecipitated twicemorewithtrichloroacetic acid. Trypsinization andion-exchange chromatographyon
Chromobeads P (Technicon) were carried out as de-scribed by Vogt et al. (23). Briefly, the pellet was oxidized for1 h at0°C with 0.1 ml of freshly made performic acid (0.1 ml of H202+1.9mlof formic acid incubated for 1 h at25°C). A 1-ml amount of water was added to stop the reaction, and the sample was lyophilized. The dried protein was dissolved in 0.5 ml of0.05 M ammonium carbonate and lyophilized a second time. Theprotein wasdissolved in 0.2 ml of 0.05Mammonium carbonatecontaining TPCK-tryp-sin(Worthington Biochemicals Corp.) in a 10:1ratio of bovineserumalbumintotrypsin andincubated for
1 hat 37°C. Asecond equalamount of trypsin was added, and the reactionwasallowedtoproceed for 1 hmore.The trypsinized protein waslyophilized, sus-pended in buffer A (280 ml of acetic acid, 4 ml of pyridine,and 716ml of water,pH 2.45), and storedat
-200C.
Cation-exchange chromatography on a jacketed Chromobeads P column (1by21cm)wascarriedout
essentiallyby the procedure of Vogtetal. (23). The column temperaturewasmaintainedat50°C. Peptides
wereeluted in 2.2-ml fractions, withapH gradient of
2.45 to 3.8. Three mixing chambers were used to
generate thisgradient. Mixing chambers1and2each contained180ml ofbufferA, and chamber3contained
60ml of bufferAand120ml of buffer B (143 ml of acetic acid, 161ml of pyridine, and696ml of water, pH 4.9). At theend ofthegradient elution (pH 3.8),
the column was washed with buffer B.
Preparationofpronasedigests for Bio-Gel P6 chromatography.Cytoplasmicextracts were
precip-itated with 20% trichloroacetic acid at40C.The pre-cipitates werecentrifuged at 10,000 rpm for30min, suspended in a minimalvolumeof0.1 N NaOH,and
dialyzed against severalchangesofdistilledwaterfor 16h at4°C.Thedialysateswerebroughtto a
concen-tration of 0.1 MTris,pH8.0,containing1mM CaCl2
(T-C buffer) (13).Pronase(10mg/ml,autodigestedin
VOL. 31, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
610 EISENBERG, HYDREAN-STERN, AND COHEN T-C buffer for2h at37°C) was addedtotheprotein sample in three portions over a period of 72 h to
achieve afinal concentration of1 mg of pronaseper mg of protein. Toluene was addedtoretard bacterial growth, andincubation was carriedout at37°C.
Pro-nasedigestswerecentrifugedat10,000xgfor30min,
and thesupernatants werefrozenat-20°C. The fol-lowing procedure was employed to extract proteins fromBIS-linked polyacrylamidegels. The appropriate bandswereexcisedfrom the driedgel and dissolvedin
5 ml of T-Cbuffer containing0.5mgof pronase. The gel pieceswereincubated inthissolution for3days, andadditional pronase (0.5 mgat atime) wasadded each day. Theextract wascentrifugedat 10,000 xg, and thesupernatantwasfrozenat-20°C(R.Klemenz,
personal communication).
Toremove sialic acid, samples wereadjustedto a
pH of1.5to 1.6 withH2SO4 (final concentration,0.05
M) andheatedintubes fitted withTeflon-lined caps
at 90°C for 1 h (5). The samples wereimmediately chilled, then adjustedtopH7.0with 1MTris.
Gel filtration. The pronase-digested glycopeptides weresubjectedtogelfiltrationon aBio-Gel P6 column (1.8by 75 cm;200 to 400mesh; Bio-RadLaboratories) equilibrated with0.1Mammoniumacetatecontaining
0.01%sodium azide. The flowrateof thecolumnwas
16 ml/h. Fractions were collected and assayed for radioactivity. Bovineserumalbuminwasused to de-fine the void volume (V0). The
['4C]glucosamine-la-beledglycopeptide markers (A=4,300; B=3,600;C = 1,800[25]), graciously supplied by Clayton Buck of the Wistar Institute, Philadelphia, Pa.,were usedto
calibrate thecolumn.Atleastonemarker wasincluded when [3H]mannose-labeled glycopeptides were chro-matographed (Fig. 6through 8). The total volume of thecolumn(Vi)wasdeterminedby using[3H]mannose
or [3S]methionine. The position of sialic acid was
determined by gel filtration of ['4C]sialic acid (all isotopes were obtained from New England Nuclear Corp.).
RESULTS
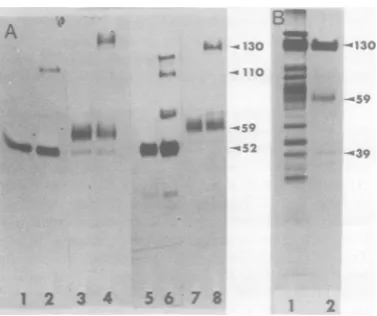
SDS-PAGE ofHSVglycoproteins. Figure 1A shows the labeling pattern of [3H]methio-nine-labeled proteins (tracks 1 through 4) and [3S]methionine-labeled proteins (tracks 5 through 8) immunoprecipitated from cytoplas-mic extracts after a 15-minpulse at 5 h p.i. or
aftera15-minpulseat 5hp.i.followedbya5-h
chase. Two antiserawereused:anti-CP-1 serum,
amonoprecipitinserumdirected against pD, and anti-ENV-1 serum, prepared against NP40
ex-tracts ofvirion envelope from HSV type 1 (2). Tracks 1 and5show that pD was immunopre-cipitated by anti-CP-1 serum from the pulse-labeled cytoplasmic extract. Tracks 3 and 7 show that the same serum precipitated D from the cytoplasmic extract of 5-h chase. Anti-ENV-1
serum precipitated pD from pulse-labeled
ex-tracts(tracks2and 6) aswellasadditional bands
at 110,000 molecular weight (110K) and 130K. This serum precipitated D from the chase
A
t ;l.w
*--*0 II- I
7::-O
6-4 M.; 4 _
td .~,
S_- _.%o
W&.W
1 2 3 4 5 6 7 8 S.
FIG. 1. SDS-PAGE analysis. (A)Fluorogram ofa
10% DATD cross-linkedpolyacrylamide gel of
im-muneprecipitates obtained from lysates of HSV-1-infected KB cells. Track 1, [3H]methionine pulse, anti-CP-I serum; track 2,[3H]methioninepulse, anti-ENV-1 serum; track 3,[3H]methionine chase, anti-CP-1 serum; track 4, [3H]methionine chase, anti-ENV-1 serum; track 5, [35S]methioninepulse, anti-CP-1 serum; track 6, [35S]methionine pulse, anti-ENV-1 serum; track 7,[35S]methioninechase, anti-CP-1 serum; track 8, [35S]methionine chase, anti-ENV-1 serum. (B) Autoradiogramof electrophoreti-cally separated (BIS-linked gels)polypeptides ob-tainedfrom purified virus; track 1, purified virions, labeled with[35S]methionine; track 2, [35S]methio-nine-labeled molecules extracted by NP40from virus particles.
(tracks4 and8) and also precipitatedbandsat
85K and 130K. The results of this experiment
are essentially the same as we previously
re-ported (1). Figure 1B shows thelabelingpattern ofpolypeptides extractedwith NP40from puri-fied [35S]methionine-labeled virions. As previ-ously shown, the glycoproteins extracted from the virion have the same molecularweights in SDS-PAGE as the corresponding mature gly-coproteins immunoprecipitated from infected cellextractsafterthe chase.Thesamepatterns
were obtained when arginine was employedas the aminoacid label.
Tryptic peptide analysis of pD and D. To obtain sufficient material for tryptic peptide analysis,multiplesamplesof each of the immu-noprecipitates shown in Fig. 1Aweresubjected
toSDS-PAGE.The bands correspondingtopD and Dwereeluted,trypsinized, and co-chromat-ographed on a Chromobeads P column. The radioactive elution profiles obtained for pD (52K) and D (59K)werequite similar (Fig. 2A). Forboth proteins, two major methionine pep-tideswereresolved,oneelutingfrom thecolumn withtheflow-throughand thesecondeluting at pH 3.33 ± 0.04 (mean of six determinations).
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.507.276.465.78.236.2]STRUCTURE OF GLYCOPROTEIN D (CP-1) OF HSV-1 611
This experiment indicates that pD and D are
structurallyrelated. The tryptic fingerprint
pro-file of methionine-labeled D extracted from
pu-rified virionswasidenticaltothatof methionine-labeled D obtained from the chase (Fig. 2B). These studies show that processing of pDtothe
matureformof D found in the virion doesnot
involve any detectable change in methionine
peptides.
The fingerprint profile for the methionine-la-beledpeptides of D is unusual intwo respects.
First, the flow-through represents atleast
two-thirds of the radioactivelabel. Second, the pat-ternisremarkably simple, in that onlyonemajor
peptide isactuallyresolved by being retainedon
the column. We considered the possibility that theflow-throughmaycontain apeptide(s)
pos-sessing trypsin-insensitive sites. In the light of
reports that carbohydrate and especially sialic acidresidues block trypsin action (9),we
consid-ered the possibility that the flow-through may
contain a peptide(s) possessing sites that are
900
500 300 250
z o 200
I-i 150
r 100 a 0.5
0
z z
w
In
a.
u
B
blocked totrypsin action. However, removal of sialic acid from D by mild acid hydrolysis before trypsinization had noeffecton the methionine
profile (datanotshown). Alternatively, as
sug-gested by H. Diggelmann (personal communi-cation), carbohydrate-containing peptides may notbind toChromobeads P. If the methionine label in the flow-throughrepresentsalarge
frac-tion of the totalpeptides, then it is possible that pD and D contain some peptides which are
different.
For thisreason,wecompared the tryptic
fin-gerprint profiles ofarginine-labeled pD and D. Both molecules share all arginine peptides (Fig. 3). Moreover, alarge numberofpeptides were
resolved, indicating that the trypsinization
pro-cedure was effective and that pD and D both
containalarge number of arginine residues.
We conclude that the conversion of pDtoD doesnotentailanymajor alteration in the
pep-tideportion of the molecule and that glycopro-tein D from the chaseisapparently identicalto
S~~~~
I300 200
25 z
0
g00
-w 75 a
50 a
I.
25
) z
0 X IaIAJ
IL
1.)
20 30 40 50 60 70 90 00 110 120 130 140 I50 60
FRACTION NUMBER
ISO 9
FIG. 2. Tryptic fingerprint analysis ofthetype-common(CP-1) glycoprotein ofHSV.(A) pD ( )isolated by immunoprecipitationand SDS-PAGEfrom [35S]methionine pulse-labeledextracts(15 minat 5 hp.i)was
co-chromatographedon acolumn ofChromobeads P with[3H]methionine-labeled D(---) isolated ina
similarfashion fromthe chase(15-minpulseat 5 hp.i., followed bya5-hchase).
0 20 30 40 50 60 70 80 90 00 110 120 30 140 150 ITO ISO
FRACTION NUMBER WAS
t
---.
VOL. 31, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.507.60.436.305.611.2]612 EISENBERG, HYDREAN-STERN, AND COHEN
0 10 20 30 40 50 60 70 80 90 100 110 120 130
FRACTION NUMBER
140 150 160 170 180 190 200
FIG. 3. Trypticfingerprint analysis ofthetype-common(CP-1) glycoprotein ofHSV.pD ( ) isolatedby immunoprecipitationandSDS-PAGEfrom[14C]argininepulse-labeledextracts(15minat5hp.i)was
co-chromatographedon acolumnofChromobeads P with[3H]arginine-labeledD(---) isolatedinasimilar fashion fromthe chase (15-minpulseat5 hp.i., followed by a5-h chase). Freearginine elutesatpH3.63
(fraction 142).
glycoprotein D found in the envelope of the vron.
Is D derived from one ofthe
glycopro-teins in thegp13Ocomplex?Itispossible that
D is derived fromoneof theglycoproteins that
make up the gpl30 complex analogous to the situation for Rauscher leukemia virus(10, 11).If thiswerethecase,Dshould contain asubset of
tryptic peptides found in the gp130 complex. Theprofile for gpl30extractedfrom the virion with NP40 (Fig.4A) consistsofa flow-through
andthree majormethioninepeptideselutingat
pH3.11, 3.23, and3.74. Anidenticalprofilewas
obtained for the gpl30 complex isolated from the chase by immunoprecipitation. Figure 4B
comparesthe elutionprofileof D withthegp130 complex.Inthisexperiment,bothproteinswere
extractedfrom thevirion. The threemethionine peptidesofthegpl30 complexelutedatpH3.12, 3.20, and 3.76, whereasthe methionine peptide
of D eluted at pH 3.36. It is clear from this
experiment that D does not share methionine peptideswith the gpl30 complex. It should be notedthatthereareunresolved
methionine-con-taining peptidesthatappearinthe column
flow-throughfor eachprotein.Although it is possible that some of the unresolved peptides may be
sharedbyDandthegpl30complex, theelution profileoftheresolvedpeptidesindicates thatD isnotderived fromgpl30.
Incorporation of
[2-3Hlmannose
into pD and D. We employed [2-3H]mannose as thecarbohydrate label to study the glycosylation
steps involved inthe conversion of pD into D. This precursor has been reported tobe
incor-porated into macromolecules as mannose or
othercarbohydrates (17). Whenuninfected cells
were pulse-labeled with mannose, a variety of proteins were labeled to a small extent. When
infected cells werepulse-labeled with mannose
(Fig. 5,track1), mostof thelabelwas
incorpo-rated into sixbands. Four of thesebandsat52K, 65K, 110K, and 130K correspond inmolecular weight to [35S]methionine-labeled proteins im-munoprecipitated by anti-ENV-1 serum (Fig.
1A, track 6). In addition, some mannose label
was incorporated into a band that comigrates
withthe154Kcapsid protein, and about 20% of the totallabelwasassociated withadiffuse band at about 17K. The fluorogram in Fig. 5 was
overexposed to enhance the fainter bands and doesnot accurately reflect the amountoflabel in each band. When thegelwascutinto2-mm
slices,dissolved in H202, and assayed for radio-activity, the labelwasdistributedasfollows: 3%
in154K, 18% in 130K, 20% in 110K, 11% in65K, 25% in52K, and 15% in 17K. The band at 17K
was also seen in pulse-labeled uninfected cells
and is apparently notvirus specific. The other five bands did not correspond to mannose-la-beled bands in uninfectedcells. When infected
cellswerepulsed with [2-3H]mannose and then
chased(Fig.5, track 2), bandsappeared at52K, 59K, and 130K, indicating thatprocessing was
occurring. However, the presence of the 52K
band in the chase indicated thatnot all ofthe glycoprotein Dwas processed. All of the
man-nose-labeled viral glycoprotein precursors and
products were immunoprecipitable by
anti-ENV-1 serum (data notshown). However, the mannose-labeled bands at 17K and 154K were
notimmunoprecipitable.
Gel filtrationofoligomannosyl residues
on Bio-Gel P6: processing of pD into D.
)o
BO
..L
0!
/WR
gV_
'k
600
560
520 480 440 400 360 320 280 240 200 160 120 80 40
IL
z z
3
LI
J. VIROL.
3c 2e 2E 24
w 22
z Z 2C
0 cr is
4 IE
u 14
i
12 2 10
a. u 8
E 4 2
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.507.65.472.50.227.2]STRUCTURE OF GLYCOPROTEIN D (CP-1) OF HSV-1 613
A
---13030KKVirusChose35S3vmet.met. 160 6oFRACTION NUMBER
59K Virus 35SMet. -130K Virus HMet.
10 20 30 40 50 60 70 80 90 l00 110 120 130 40 150 160 170 190 190
* FRACTON NUMBER
FIG. 4. Trypticfingerprint analysis of HSV glycoproteins. (A) gpl3O complex(---) isolatedby immuno-precipitationand SDS-PAGEfrom[3H]methionine-labeledchasewasco-chromatographedon acolumnof
Chromobeads P with[35S]methionine-labeledgp13O ( ) extractedfrom purifiedvirions with NP40. (B) [3S]methionine-labeled D( ) extractedfrom purifiedvirions withNP40wasco-chromatographed with
[3H]methionine-labeled gpl3O complex(----)extractedfrom purifiedvirions withNP40.
Figure 6shows the elution profiles of
pronase-digestedglycopeptidesobtained fromcells
pulse-labeled with [2-3H]mannose. The cytoplasmic
extractof both uninfectedand infected KBcells
(Fig.6A andB)containedarelatively
symmet-rical peak of oligomannosyl residues with an
estimated molecular weight of 1,800. This size corresponds tothat of thecore oligosaccharide
which is known to be attached to cellular and viralglycoproteins byadolichol-mediated
mech-anism (24). To examine theoligosaccharidesof
pD directly, thebandcorrespondingtopDwas
excised from the SDSgel,eluted in thepresence ofpronase,andappliedtoaBio-Gel P6 column.
pDcontainedthesamesizeoligomannosyl core
moiety as wasfound in the total pulse-labeled
extract(Fig.60). Itappearsthat all of the
pulse-labeled glycoproteins contain the same
oligo-mannosylcore.Totestfor thepresenceof sialic
acid, pronase-treated pDwassubjectedtomild
acidhydrolysis.This treatment hadnoeffecton
theelutionprofileoftheoligomannosylresidues
isolatedfrompD(Fig. 6D), indicatingthat sialic acidwasnotpresent.
To follow processing of the carbohydrate, a
similar set ofexperimentswasperformed using
the [2-3H]mannose-labeled cytoplasmic extract obtained from thechase. The totalextract(Fig. 7A) contained the 1,800-molecular-weight gly-copeptide seen in the pulse, but in addition it
contained aheterogeneoussetofglycopeptides
thatelutedinpositionscorrespondingtolarger
molecules. Approximately 33% of the label eluted ina3,600-molecular-weightfraction and
approximately 10% eluted withorjustafter the voidvolumeand hadamolecularweightof4,300 or greater. Mild acid treatment of the chase
resulted inadramatic alteration of the elution
profile (Fig. 7B). A significant amount of the
larger glycopeptidematerialseeninFig. 7Awas
a. 0
B
500 240
210
0
VOL. 31, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.507.71.412.54.394.2]614 EISENBERG, HYDREAN-STERN, AND COHEN
.
__
-
.uS.:
<.Xs
*::. e
_. ...
_zEF'''
130
-< 110( .I*
'I5l
1I--
52
1
2
FIG. 5. Pulse-chase experiment with
[2-3Hlman-nose asaprecursor.Fluorogram ofa 10%
polyacryl-amidegel(BIS linked)of cytoplasmicextractsfrom
HSV-1-infectedKBcells. Track 1: cellswere
pulse-labeledfor30minat5hafter infection. Track2:cells
werepulse-labeled for30minat5h after infection andthen chased with nonradioactive medium for5 h. The film was overexposed to enhance the faint bands.
shifted tolower molecular weights. The glyco-peptidesobtaineddirectlyfromDhadasimilar
elution profile tothose seen inthe total chase
(Fig.7A).However,asomewhatgreaterfraction
ofthe label (27%) was present in molecules of greaterthan 3,600molecularweight.Treatment
of Dwith mild acid (Fig. 7D) lowered the
mo-lecularweight ofthelargerglycopeptides. The
elution profile ofmost ofthe label (83%) was
shifted to abroad peakwithanaverage
molec-ular weight greater than 1,800. These results
suggest that most of the higher-molecular-weightoligomannosyl residues found in D
con-tain sialic acid. We conclude thatprocessing of
pD
to D involves chain elongation,including
extensive
sialyation.
Figure 8A shows the elution profile of the oligomannosyl residues obtained by pronase digestionof the NP40extractofpurified virions. Itcanbeseenthat theelutionprofile is similar
tothatseenfor thechase.However, most of the [3H]mannose label was associated with mole-cules of 3,600 molecular weight or greater. A smallfraction of thecounts (15%)wasfound in the 1,800-molecular-weight region. Thus, there appear to be at least three different
mannose-labeled oligosaccharides in the virionenvelope. Furthermore, these results suggest that virion glycoproteins areheterogeneouswithrespectto
carbohydrate content. Treatment with mild acid had precisely the same effect on the virion gly-copeptides as it had on glycopeptides from D (cf. Fig. 7D and8B). These results indicate that D found in the chase is probably the final gly-cosylated form of the glycoprotein found in the virion.
Weconclude that there appear to be at least
twoposttranslationalprocessing steps in produc-tion of the matureglycoprotein D. The first step involves the addition of an 1,800-molecular-weight mannose-containing oligosaccharide to
thecorepeptidetoformpD. The secondstep(s), which involves addition offurthercarbohydrate residuesincluding sialic acid tothe 1,800-molec-ular-weight oligosaccharide, probably accounts forthe slowermobility of D in SDS-PAGE.
Incorporation of
["4C]glucosamine
into HSVglycoproteins.HSV-infected KB cells do notincorporatesignificant amounts of ['4C]glu-cosamine label intotrichloroacetic acid-precipi-table materialduring a30-min pulse. However, we found that label added during the 30-min pulse was incorporated into acid-precipitable material during the 5-h chase period. Honess and Roizman (7) observed that therateof entry oflabeledglucosamineintotheacid-solublepool of bothinfected anduninfected cells was much greater than its rate ofwithdrawal. Moreover, the fact thatmostof the solublepoolwas UDP-N-acetyl glucosamine suggested that this larger pool would not exchange with the medium to permitaneffective chase. Thus,wefound that label added to HSV-infected cells during the pulse was present long enough to be incorpo-rated eventually into glycoproteins during the "chase" period. Figure 9A shows the elution profile ofglycopeptides obtained from the pro-nase-digested["C]glucosamine-labeled
"chase." Approximately23% of theglucosaminelabelwasin the voidvolume, andmostof theremaining labeled molecules had a molecularweight
cor-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.507.104.228.73.423.2]STRUCTURE OF GLYCOPROTEIN D (CP-1) OF HSV-1 615
4300 1800
I*f 3600 Vt
C~~~~~~~~~~~~~~~~i i
!t
,. I I
0 60 80 100 120 140 160 ISO 200
FRACTION NUMBER
4300 1g00
- Vo1 3600
B i I
Vt
0 60 60 100 120 140 160 10O 200
FRACTION NUMBER
n
z
4
100 120 140So O0 FRACTION NUMBER
FIG. 6. Gelfiltration chromatographyonBio-Gel P6 ofpronasedigestsobtained from HSV-1 uninfected
and infectedKB cellspulse-labeled with [2-3HJmannose. Glycopeptideswere obtained from: (A) thetotal
cytoplasmic extract of uninfected KB cells; (B) the total cytoplasmic extract of HSV-infected cells; (C)
glycoproteinpDexcisedfrom apolyacrylamidegel;(D)glycoproteinpD fromthegel treated with0.05MH2SO4
at90°C for h.
respondingto the 3,600-molecular-weight peak labeled with [2-3H]mannose. Mild acid treat-ment(Fig. 9B) converted approximately 36% of theglucosaminelabeltoaposition
correspond-ingtoauthentic sialicacid,and45%ofthelabel then eluted at amolecular weight of
approxi-mately 3,000.Littleifanylabelwasobserved in
the1,800-dalton peak beforeorafter acid treat-ment.
Itshould be notedthat Honess and Roizman (7) reported that 30to40% of the glucosamine labelpresentinpurifiedvirionswasintheforn
of sialic acid. Ourresults showed thatasimilar
amount of radioactive glucosamine (36%) was
convertedtosialicacid in infected cells. Does theconversion ofpDtoDinvolvea
changein electricalchargeas weli as
mo-lecular weight? The results presented so far
suggestthat theprocessedformof D issialyated and theunprocessedform isnot.Thus,the
pre-cursor and product should differ in charge as
wellasmolecularweight.Totestthisprediction,
wesubjected the
[tS]methionine-labeled
pulseand chaseextracts totwo-dimensionalgel elec-trophoresis. Although several changes were
noted in thepositionof labeledpolypeptidesof thepulseandchase, thecomplexityof the
pat-ternsmade itimpossible topinpoint where pD and Dwerelocated. Wethereforeemployedour
antisera asimmunological probes to select the
pDand D componentsoutof thiscomplex
mix-ture ofproteins. Anti-CP-i serum
immunopre-cipitated pD (Fig.1OA; cf.Fig. 1A,track5)which
hadanisoelectric point (pI) of8.0.Inaddition,
there was a trailing edge ofprotein extending into the lowerpHrange.This antiserum
immu-noprecipitated D as aheterogeneous molecule
of 59Kto64K, havinganaveragepIof 6.4(Fig. lOB;cf.Fig. 1A,track7).It should be noted that
the higher-molecular-weight material had a
700
600o
0 2
z
300
200F
100
FRACTIONNUMBER
z
900 800 700
600 500 400 300 200 100
VOL. 31,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.507.94.415.63.397.2]616 EISENBERG, HYDREAN-STERN, AND COHEN
2
2
0
z
2
2
4n
2
2
o 60 s0 100 120 140 o0 a0 200 FRACTION NUMBER
un
0 z I
2
0 60 60 m0 120 140 a
FRACTION NUIMBER
FIG. 7. Gel filtrationchromatography on Bio-Gel P6ofpronasedigestsobtainedfrom HSV-infectedcells
pulse-labeledwith[2-3H]mannose for30min and chased with unlabeledmediumfor5h.Glycopeptideswere
obtainedfrom:(A)the totalcytoplasmicextractof infected cells; (B)the totalcytoplasmicextracttreated with 0.05 MH2SO4at90°C forIh; (C) glycoproteinDexcisedfromapolyacrylamide gel;and(D) glycoproteinD
fromthegel treated with 0.05 MH2SO4 at 90°C for 1 h.
greaternegative charge,possibly duetothe fact that it was more heavily
sialyated.
Figure 10C showstheresults oftwo-dimensional electropho-resis of[3S]methionine-labeled polypeptides ex-tracted withNP40 frompurified
virions.Several labeled components were seen, including: (i) a 59K to 64K molecule, pI 6.4 (D); (ii) a 130K component with an average pI of 5.0; and (iii) acomponent(s)
with apIofapproximately6.0anda molecular weight of approximately 70K to 90K.Thus it appears that theprocessedform of D found in the virion has the same pI and molecularsize asitdoes in the chase. We con-cludethat the conversion of pD to D involves a changeinelectrical chargeaswellasmolecular weight, and that both ofthese alterations are due to theaddition ofcarbohydrate,
especially
sialicacid,totheprotein.
DISCUSSION
The results presented in this study firmly establish a structural relationship between the twoforms of theCP-1antigen, pD andD.
Tryp-tic
fingerprint profiles
ofmethionine- andargi-nine-labeledproteinsshow thatbothprecursor and product forms share these peptides. The processedform ofCP-1 (D) found in the cyto-plasm has thesamemethioninepeptidesasthat foundinpurifiedvirions.Moreover, the tryptic fingerprintprofile of Ddiffersfrom thatofthe gpl30 complex. Thus, it wouldappearthatD is notabreakdownproduct ofa larger-molecular-weightglycoprotein. The presentdata confirm and extendpreviousobservations(2, 19)thatpD and D are antigenically related to each other andareunrelatedtothe 130Kglycoproteins of HSV.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.507.115.423.68.394.2]STRUCTURE OF GLYCOPROTEIN D (CP-1) OF HSV-1 617
1800 S.A. Vt
A
0 60 80 100 120 140 60 180 200
FRACTION NUMBER
B
4300 1800 \Ob 3600 i
f $
400[
350
w 0 z
z
I
C-,
300 250S 200
lSO
100
50
0 60 80 100 120 140 160 FRACTION NUMBER
FIG. 8. Gelfiltration chromatographyon Bio-GelP6ofpronasedigestsobtainedfrom
[2-3H]mannose-labeledpurifiedHSV virions.Glycopeptideswereobtainedfrom: (A)theNP40extractofpurified virions;and
(B)theNP40extractofpurifiedvirions treated with 0.05 MH2SO4at90'C forI h.
Mannoselabeled in the 2-tritium position
ap-pearstobeanexcellentchoiceas acarbohydrate precursorsince: (i) it is rapidly incorporated into
oligosaccharides in theform ofmannoseorother
sugars (17); (ii) it is rapidly incorporated into
viral glycoproteins; (iii) it is incorporated into the initialcoreoligosaccharide allowing the
ini-tialstepsofprocessingtobestudied; and (iv) it is not converted into sialic acid (Fig. 7B) (26). Using [2-3H]mannose as the carbohydrate
probe,wehave shownthat processing of pDto
D aswell asthe processing of the other
glyco-proteins of HSV involves alterations of the
car-bohydrate portion of the molecule.By analyzing
pronase digests of pD and D, we were ableto
follow the flow ofmannoselabel froman
1,800-molecular-weight moiety added to the protein within 5 minofsynthesis (unpublished data)to
larger, more heterogeneous carbohydrate
com-ponents. A similar flow was found for the
un-fractionated mixture of glycoproteins in the pulse and chase,suggesting that the other HSV glycoproteins may be processed in a fashion
similar toD. Theseresultsagree withthose of Honess and Roizman(7),whostudiedthe
proc-essing of oligosaccharides by using glucosamine
asthelabel inamixtureofHSVglycoproteins.
However, beforewe canconcludethat all HSV
glycoproteinsare processedsimilar toD,it will
benecessarytoexamine theprocessing ofeach
of the other glycoproteins individually. Spear (19), for example, reported that the processing of Adiffers from that of the otherglycoproteins and in factdoesnotchangeinmolecularweight. That processing of the oligosaccharide portion of D involves extensivesialyationwasshownin
severalways. First, mild acidhydrolysis of the
larger mannose-labeled glycopeptidesconverted
themtolower-molecular-weight forms.Second, when glucosamine-labeled glycopeptides were
subjected to this treatment, 36% of the label
eluted fromBio-Gel P6atthesamepositionas
sialic acid.
Thisfinding predicted thatprocessing ofpD
to D involved not only a change in molecular
weight but alsoasignificantalteration in
molec-ularcharge. Two-dimensional gel electrophore-sis studies indicatethatpDisabasic molecule
withaplof8.0, whereas the processed form, D,
ismoreacidic (averagepl=6.4).In thisregard,
it is worth noting that Thompson et al. (22) reported that the surface charge of
HSV-in-fectedcells differs from that ofuninfectedcells
as well astheenvelopeofpurifiedvirions.They foundthat,whereasuninfectedcontrolcells
fo-cused ataplof4.48,infected cells became less
acidic, focusing at a pI of 4.58. The pI ofthe
purifiedvirionwasstillhigher (5.0to5.8).From
400
w 0
z z
< 300
I
L- 200
0
S.A. Vt
2
180 200
VOL. 31, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.507.67.441.73.316.2]618 EISENBERG, HYDREAN-STERN, AND COHEN
0 60 80 100 120 140 160 180 200 FRACTION NUMBER
In
w
0
z
z
4
II 0)
a.,
100 120 140 160 FRACTION NUMBER
FIG. 9. GelfiltrationchromatographyonBio-GelP6ofpronasedigestsobtainedfromHSV-infectedcells. Thecellswerepulse-labeled for30minwith[2-3HJmannose (narrowline)or[4C]glucosamine (heavy line),
then chasedfor5 h. Glycopeptideswereobtainedfrom: (A) cytoplasmic extract;and(B) totalcytoplasmic extracttreated with 0.05 MH2SO4 for1hat90PC.
thesedataaswellasdirectanalysisofthe
num-ber ofamino groups, they concluded that the
plasmamembrane ofHSV-infectedcells,aswell asthe virionenvelope,weremodifiedbyaddition
ofbasic aminogroups,possiblypresentin virion glycoproteins.Our isoelectricfocusingdata
sug-gestthatglycoproteinD hasasignificant
num-ber of basic amino groups, and therefore this
glycoproteinmaycontributesubstantiallytothe electrokinetic properties of the plasma
mem-brane andvirion envelope. Itis evident that a
detailedanalysisof the amino acidcomposition of D isrequired.However,basedonthe data in
Fig. 3,we canatleastspeculateabout the
min-imum numberofarginineresidues inD. Assum-ingthat the peakwith the smallest number of
counts(Fig.3,fractions 102through 107) repre-sents one arginine peptide (12), we estimated
that there are approximately 30 arginine
resi-dues in the molecule.
Assuming
that the coreprotein (minus carbohydrate) has amolecular
weightof 50,000(L.Pizer, G.H.Cohen,andR. J. Eisenberg, unpublished data) and contains 416amino acids eachwithanaveragemolecular
weightof120, thenapproximately 7%ofthe D proteinisarginine. Themajor drawbackofthis estimation is thatpeakheightsarenotalways reliable.
Our results suggest thatprocessingofthe
oli-gosaccharide(s) of Doccursinafashionsimilar
tothatreportedfor the Gglycoproteinof vesic-ular stomatitis virus(8, 21)aswellasotherviral
and cell glycoproteins (21, 24). In all cases of eucaryotic glycoprotein synthesis studies to date, asparagine-linked oligosaccharides are
formed first byenblocaddition ofa"simple"or
mannose-richcorecontaining N-acetyl
glucosa-mine andmannose viaadolichol intermediate
(24). Thisisfollowedbyaddition of othersugars,
including galactose, N-acetyl glucosamine,
fu-cose,and sialicacidtoforn acomplexstructure.
Insome casesit has been shown that, before
addition ofthese other sugars, some mannose
residuesareremoved (21). We havenoevidence so far to suggest that this step occurs in the
conversionofpDto D.Studies of this possible intermediate processingstep, involvingamore
detailed kineticanalysis of thepulseand chase,
areunderway.
As noted before, D isolated from the
cyto-plasm isstructurallyandantigenicallysimilarto
Disolated fromthevirus. However,one differ-enceisevident.Dasisolated from thecytoplam
contains mannose label in a
low-molecular-weightform (less than500;seeFig. 7C), whereas noneof themannoselabel in the virion is ina
low-molecular-weightcarbohydrate (Fig.8).The
natureofthis material isnotknown presently. Thelossof thismaterialmight reflectafurther
processingstepin the maturation ofthe virion w
U, n
zo 4
I
a-to
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.507.65.458.73.308.2]STRUCTURE OF GLYCOPROTEIN D (CP-1) OF HSV-1 619
lecular weight of the oligosaccharides from that of theglycosylated peptide (4, 14). If this addi-tiverelationship is valid for protein D of HSV, thenthere are no more than two to three aspar-agine-linked oligosaccharide chains present on thismolecule.
ACKNOWLEDGMENTS
Thisinvestigation was supported by Public Health Service grantDE-02623 from the National Institute of Dental Re-search.R.J.E. was supported in part by aspecial grant from theUniversity of Pennsylvania.
We thank WilliamLawrence and Wesley Wilcox for their help in preparation of this manuscript,Heidi Diggelmann for allowing us toread her manuscript before publication, and ClaytonBuck for supplying us with marker glycopeptides. We also wishtothank Walter Taylor formasteringthetechnique oftwo-dimensional gel electrophoresis and running the sam-ples, and we thank Madeline Cohen for her excellent technical assistance.
C
I
oI4.9s
-1309
'
'''-:^.&.:
.5
9KFIG. 10. Two-dimensional gel electrophoresis of
HSV-Iglycoproteins. The immuneprecipitates in A and Bwereobtainedbyusinganti-CP-1 serum. (A)
Immuneprecipitates obtained fr-om extracts
pulse-labeled with[3'S]methioninefor15minat5hafter infection (see Fig. IA, track5). (B)Immune
precipi-tates obtained from extracts pulse-labeled with
[3'SJmethioninefor15minat5hafter infection,then chased with unlabekedmediumfor5h(seeFig. IA,
track7).(C)NP40extractof['S]methionine-labeled
purifiedvirions(seeFig. JB,track2). envelope.
It is noteworthy that the glycopeptides iso-lated from D containsomeof the 1,800-molecu-lar-weightormannose-richcoreoligosaccharide.
Itisunlikelythat band D excised fromthe gel
wascontaminated withpD,because thedistance between pD and D was sufficiently great to
insure that Dwasisolated free ofa
contanminat-ing precursor. Thus, it is possible that D may
containmore thanoneasparagine-linked
oligo-saccharide. Previous experiments with
tunica-mycinsuggestedthat theunglycosylatedform of D has a molecular weight in SDS-PAGE of
approximnately 50,000 (Pizer etal., unpublished data).In thecaseof otherviruses,themolecular
weight of tunicamycin-tr',~ated
carbohydrate-free polypeptides corresp6nds closely to the
weight predicted bysubtraction of the total
mo-LITERATURE CITED
1. Bonner, W. M., and R. A. Laskey.1974.A film detection method for tritium labeledproteinsandnucleic acidsin polyacrylamide gels. Eur. J. Biochem. 46:83-88. 2. Cohen, G. H., M. Katze, C. Hydrean-Stern, and R. J.
Eisenberg.1978.Type-commonCP-1antigenofherpes simplex virus is associated with a
59,000-molecular-weight envelopeglycoprotein. J. Virol. 27:172-181. 3. Cohen, G. H.,M.PoncedeLeon,and C. Nichols. 1972.
Isolation ofa herpes simplex virus specific fraction
whichstimulates the production ofneutralizing anti-body.J. Virol.10:1021-1030.
4. Collins,J.K.,andC. A.Knight.1978. Purificationof theinfluenzahemagglutinin glycoprotein and charac-terization of itscarbohydratecomponents.J. Virol.26: 457467.
5. Downs, F.,and W.Pigman.1976.Qualitativeand
quan-titativedetermination of sialicacids,p.233-240. In R. L.Whistler and J. N. BeMiller(ed.),Methods in
car-bohydrate chemistry.Academic PressInc.,NewYork. 6. Gibson,W. 1974.Polyomavirusproteins.Adescription ofthe structuralproteinsof the virion basedon
poly-acrylamide gel electrophoresis and peptide analysis. Virology62:319-336.
7. Honess,R.W.,and B.Roizman.1975. Proteinsspecified by herpes simplex virus. XIII. Glycosylation of viral polypeptides.J.Virol.16:1308-1326.
8. Hunt,LA.,J. R.Etchison,and D. F. Summers. 1978. Oligosaccharidechainsaretrimmedduring synthesisof
theenvelope glycoproteinof vesicular stomatitusvirus. Proc. Natl.Acad. Sci. U.S.A. 75:754-758.
9. Klemenz,R.,and H.Diggelmann.1978. Thegeneration of the two envelope glycoproteins of RousSarcoma Virusfroma commonprecursorpolypeptide. Virology
85:63-74.
10. Krantz,M. J., M.Strand, and J. T. August. 1977. Biochemicalandimmunological characterizationofthe major envelope glycoprotein gp69/71anddegradation fragmentsfrom Rauscher leukemia virus. J. Virol. 22: 804-815.
11. Marquardt, H., R. V. Gilden,and S.Oroszlan. 1977. Envelope glycoproteinsof Rauscher murine leukemia virus: isolation and chemical characterization. Bio-chemistry16:710-717.
12. Milman, G., S. W. Krauss, and A. S. Olsen. 1977. Tryptic peptide analysisofnormalandmutantforms of hypoxanthine phosphoribosyl-transferase from HeLa cells. Proc.Natl.Acad. Sci.U.S.A. 74:926-930. 13.Nakamura,K.,and R. W.Compans.1977. Thecellular
site of sulfation of influenza virusglycoproteins. Virol-A
*.:
PoI
P.'..s2
B
t::
Op.
""':'.'
- 59K VOL. 31, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.507.67.204.70.369.2]620 EISENBERG, HYDREAN-STERN, AND COHEN
ogy79:381-392.
14. Nakamura,K., and R. W.Compans.1978.
Glycopep-tidecomponentsofinfluenzavirusglycoproteins.
Virol-ogy86:432-442.
15. O'Farrell, P.1975.High resolution two-dimensionalgel
electrophoresis ofproteins. J. Biol. Chem. 250:4007-4021.
16. Ponce de Leon, M.,H.Hessle,andG. H. Cohen. 1973. Separationofherpes simplex virus-induced antigens by concanavalinAaffinitychromatography.J. Virol. 12: 766-774.
17. Schwarz, R. T., M. F. G. Schmidt, U. Anwer, and H.-D. Klenk. 1977. Carbohydrates ofinfluenzavirus. I.
Glycopeptides derived from viral glycoproteins after labelingwithradioactivesugars. J. Virol. 23:217-226.
18. Shively, J. E., M. J. Kessler, and C. W. Todd. 1978. Aminoterminalsequencesof the major trypticpeptides
obtained from carcinoembryonic antigen by digestion with trypsin in the presence ofTriton X-100. Cancer Res. 38:2199-2208.
19. Spear,P.G.1976.Membrane proteins specified by herpes simplex viruses. I. Identification offour glycoprotein
precursorsandtheir productsintype-l-infected cells. J.Virol.17:991-1008.
20. Spear,P.G., and B. Roizman.1972.Proteins specified by herpes simplex virus.V.Purification and structural
proteinsoftheherpesvirion.J.Virol. 9:143-159. 21. Tabas, I.,S.Schlesinger,andS.Kornfeld.1978.
Proc-essingofhighmannoseoligosaccharidestoform
com-plextypeoligassacharideonthenewlysynthesized
pol-ypeptidesofthevesicular stomatidis virus Gprotein andtheIgG heavychain. J. Biol. Chem. 253:716-722. 22.Thompson,C.J.,J. J.Docherty,R.C.Boltz,R.A. Gaines,and P. Todd. 1978.Electrokinetic alteration of the surface ofherpes simplexvirusinfectedcells.J. Gen. Virol.39:449-461.
23. Vogt,V.M.,R.Eisenman,and H.Diggelmann.1975. Generation of avianmyeloblastosisvirus structural
pro-teinsby proteolytic cleavageofaprecursorpolypeptide.
J. Mol. Biol. 96:471-493.
24. Waechter,C.J.,and W. J.Lennarz.1976.The role of polyprenol-linked sugars in glycoprotein synthesis. Annu. Rev.Biochem. 45:95-112.
25. Warren, L., J. P. Fuhrer, C. A. Buck, and E. F.
Walborg.1974.Membraneglycoproteinsinnormaland virustransformedcells, p.1-26.InJ.Schultz andR. E. Block(ed.),Miami WinterSymposium,vol.8.Academic PressInc.,New York.
26. Yurchenco,P.D.,C.Ceccarini,and P. H.Atkinson. 1978.Labelingcomplex carbohydratesof animal cells withmonosaccharides. MethodsEnzymol.50:175-204.
J. VIROL.


![FIG. 4.precipitation[3H]methionine-labeled[3S]methionine-labeledChromobeads Tryptic fingerprint analysis of HSV glycoproteins](https://thumb-us.123doks.com/thumbv2/123dok_us/1507616.103440/6.507.71.412.54.394/precipitation-methionine-methionine-labeledchromobeads-tryptic-fingerprint-analysis-glycoproteins.webp)


![FIG. 7.pulse-labeledfromobtained0.05 Gel filtration chromatography on Bio-Gel P6 ofpronase digests obtained from HSV-infected cells with [2-3H]mannose for 30 min and chased with unlabeled medium for 5 h](https://thumb-us.123doks.com/thumbv2/123dok_us/1507616.103440/9.507.115.423.68.394/labeledfromobtained-filtration-chromatography-ofpronase-obtained-infected-mannose-unlabeled.webp)
![FIG. 8.labeled(B) Gel filtration chromatography on Bio-Gel P6 of pronase digests obtained from [2-3H]mannose-purified HSV virions](https://thumb-us.123doks.com/thumbv2/123dok_us/1507616.103440/10.507.67.441.73.316/labeled-filtration-chromatography-digests-obtained-mannose-purified-virions.webp)


