0022-538X/80/05-0521/11$02.00/0
Structural
Analysis
of the
Capsid
Polypeptides
of
Herpes
Simplex
Virus
Types
1
and 2
GARY H. COHEN,`* MANUELPONCE DE
LEON,'
HEIDI DIGGELMANN,2 WILLIAM C.LAWRENCE,3 STEVEN K. VERNON,4ANDROSELYN J.EISENBERG3
DepartmentofMicrobiology and Center for Oral Research, SchoolofDental
Medicine,'
andDepartmentof Pathobiology,SchoolofVeterinaryMedicine,3 University ofPennsylvania,Philadelphia, Pennsylvania19104;SwissInstituteforExperimental CancerResearch,CH-1066Epalinges,Switzerland2; and
DepartmentofBiological ProductDevelopment, WyethLaboratoriesInc.,Philadelphia,
Pennsylvania 191014
Capsidsofherpessimplexvirus(HSV)types1and2containsevenpolypeptides
ranging inmolecularweightfrom 154,000 to12,000 (termed NC-1
through
NC-7in orderofdescending molecular weight). Antibodies
prepared
toHSV-1capsid
polypeptides isolated fromsodiumdodecyl
sulfate-polyacrylamide gels
reacted inan immunofluorescence assay against HSV-i-infected KB cells. Three of the
antibodies (anti-NC-i, anti-NC-2, and anti-NC-3,4) also reacted with
HSV-2-infected cells. Tryptic peptide analysis showed that each of the HSV-1 capsid
polypeptideshad auniquemethioninepeptideprofile, andnoneappearedtobe
derived from themajor capsidpolypeptide. Comparativepeptide analysisof
HSV-1andHSV-2 showed thatonepolypeptide (NC-7, 12,000molecularweight) had
anidentical methioninepeptideprofileandaverysimilararginine
peptide
profilein both virus types. The arginine peptide profile of NC-7 of HSV-1 was very
different from thearginine profileof KBhistoneH4.
Although
therewerecertainintertypic similarities in the methionine peptide profiles of the other capsid
components, especially in NC-1 (the major capsid protein), there was no case
where thetrypticpeptideswereidentical inthetwovirus types.
Gibson and Roizman (14, 15) analyzed the
proteincomposition ofherpessimplexvirustype
1(HSV-1)andHSV-2nucleocapsids.Twomajor
intranuclear formsweredescribed: (i) an Aform
lacking DNA and containing one minor and
threemajorstructuralpolypeptides;and(ii) a B
formcontainingallofthepolypeptides foundin
the Acapsidaswellas two additional
polypep-tides. Recentlywe presented a preliminary
re-port (G. H. Cohen, W. C.Lawrence, M. Ponce
deLeon, S.Vernon, andH. Diggelmann, Abstr.
Annu. Meet. Am. Soc. Microbiol. 1978, S42, p.
219) confirming these results (14, 15) and
de-scribing an additional 12,000-molecular-weight
polypeptide (NC-7). Heilman et al. (16) also
found a protein pattern for capsids similar to
thatreported by Gibson and Roizman (14, 15)
and confirmedourreportthat HSV capsids
con-taina 12,000-molecular-weight polypeptide.
Although HSV-1 differs significantly from
HSV-2 in a number ofbiological, biochemical,
and biophysical characteristics (29), the two
forms also exhibit similarities. DNA-DNA hy-bridization studies indicate that HSV-1 and
HSV-2 exhibit approximately 50% homology in
their DNA with 85% matchingofbase pairs of
thehomologousregions (18). 1 and
HSV-2 capsid polypeptides have very similar
molec-ularweights,and thepossibility exists that they
have thesame orsimilar amino acid sequences.
However, the major capsid protein of HSV-1
(154,000 molecularweight) hasa slightly faster
electrophoretic mobilitythan thecorresponding
protein found in HSV-2capsids (155,000
molec-ularweight) (3, 8). This differencewas utilized
todetermine the location of the genecodingfor
the (NC-1) polypeptide in intertypic
recombi-nants (20, 22). Even though the major capsid
polypeptides of HSV-1 and HSV-2 are
distin-guishableonthe basis ofelectrophoretic
mobil-ity, theyarerelatedimmunologically (8, 16, 26).
Inaddition, Heilman etal. (16) showed that a
secondcapsidpolypeptide,p40 (equaltoNC-3),
found in the heavyor B capsids of HSV-1 and
HSV-2,hadboth cross-reactive and type-specific
antigenic determinants. Except for reports of
amino acidcomposition(9) and ofdisulfide
link-agesin HSV-2 polypeptides (38), little is known
about the chemicalstructureof theHSV capsid
oritspolypeptides.
The purpose of the present studies was to
characterize the capsid polypeptides ofHSV-1
and HSV-2. We will present evidence to show
that all ofthe HSV-1 capsid polypeptides
iso-lated from sodiumdodecyl sulfate
(SDS)-poly-acrylamide gelsareimmunogenic andstimulate
521
on November 10, 2019 by guest
http://jvi.asm.org/
COHEN ET AL.
production of antibodies thatreactwith
HSV-1-infected cells. Three of these also react with
HSV-2-infected cells. Tryptic peptide analysis
showed that each of the HSV-1capsid
polypep-tides containedauniquesetofmethionine
pep-tides, andnoneappearedtobe derived fromthe
major (NC-1) capsid polypeptide. Comparative
analysis showed that two polypeptides, NC-1
andNC-7, had similar tryptic peptide profiles in
HSV-1 and HSV-2. The other capsid
compo-nentsof HSV-1 showed fewer structural
similar-ities with the corresponding components of
HSV-2.
MATERIALS AND METHODS
Cell cultures. Conditions for thegrowth and
main-tenance of KB cells and baby hamster kidney cells have beendescribedpreviously (7).
Viruspreparationand titration. The procedures used for the preparation of virus stocks of HSV-1 (strain HF) and HSV-2 (Savage strain) have been
describedpreviously (5, 7).
Preparation ofinfected KB cells for
nucleocap-sidpurification.KBcellswerepropagatedinplastic
rollerbottles (490cm2) essentiallyassuggested by C. Shipman, Jr. (personal communication). The roller
bottleswerepretreated withEagle minimal essential mediumsupplementedwith 10% calfserumfor1hat
36°C, and thenwereseededwith 2.5 x 107 KB cells. Monolayers wereobtained within 72 handwere in-fected with0.5 PFU of HSV-1or 10PFU ofHSV-2
percell.At 5 hpostinfection,the mediumwasreplaced
with 50ml ofEagle minimal essential medium
con-taining 1/10 of the usualamountof methionine and5
,uCi of [35S]methionine (specific activity, >400 Ci/
mmol),or 5,uCiofL-[methyl-3H]methionine (specific
activity, 100Ci/mmol),or1,uCiof['4C]arginine
(spe-cificactivity,270mCi/mmol),or 5uCiof[3H]arginine
(specific activity, 18 Ci/mmol) per ml. All
radioiso-topes were purchased from New England Nuclear
Corp. The cells were incubated for 24 h at 34C,
harvestedbycentrifugation,andwashed with0.01 M
phosphate buffer (pH7.2).
Purification ofnucleocapsids. The method
em-ployedwas acomposite of four established procedures (9, 14, 23, 28). Briefly, nuclei were isolated from
in-fectedcells(14)andsuspendedinphosphate bufferto a density of106 nuclei per ml. The suspension was
homogenized and sonicated three times for30 s.
So-diumdeoxycholatewasaddedtoafinalconcentration of0.5%, and thesuspensionwasincubated for 1 hat 36°Cwithconstantagitation.DNase (100
jig/ml)
and RNase (120pg/ml) wereaddedto thepreparationin the presence of1 mM MgCl2andincubatedat36°C for1 h. MgCl2 (final concentration,0.02M)wasthen addedtoprecipitatethedeoxycholate.Theprecipitate was removed by centrifugation at 10,000 x gfor 20 min, and the supernatantfluidwas pelletedthrough 36%sucroseinanSW27rotor at90,000xgfor90min. The pellet was suspended in 5 ml of 40% (wt/vol) CsCl,sonicatedbriefly,overlaid witha12to30% linearCsClgradient (12 ml),andcentrifugedinan SW27.1 rotor at40,000 x gfor 17 h. Thecapsids of HSV-1
werevisible as two bands in the center of the gradient.
HSV-2capsids were not resolved into two populations,
butappearedas abroad band. When required, further
purificationof the heavy (buoyant density = 1.2890 g/
cm3)orlight(buoyant density = 1.2750g/cm3) HSV-1capsidswas achieved by a second sedimentation step
throughalinear22to 40% CsCl gradient for 90minat 90,000 x g. Capsidsstored in 38% CsCl at 4°C were
stable for several weeks. The purity of the
prepara-tions was determined in three ways: (i) by electron
microscopy (35, 36); (ii) by SDS-polyacrylamide gel
electrophoresis (PAGE) analysis; and (iii) by mixing
experiments in which [35S]methionine-labeled
unin-fected nucleiwereaddedtounlabeled purified capsids
before asecond round ofpurification. The purified
capsid preparation containedless than 5% of the label
of the mixed starting preparation, and SDS-PAGE
analysis revealed no radioactive bands comigrating
with capsid polypeptides.
Histones were extracted from uninfected KB cell nucleiaccordingtothe procedure of Bonneretal.(1).
SDS-PAGEanalysis.SDS-PAGEwascarriedout inslabs of 10%acrylamide cross-linked with 0.4% N,N-diallyltartardiamide or on slabs of 18% acrylamide cross-linked with 0.26%N,N-methylenebisacrylamide asdescribedpreviously(6, 10). Afterelectrophoresis, the gelswere stained with Coomassie brilliant blue, driedonfilterpaper,andplacedincontactwith Kodak X-Omat R (XR-5) film. For fluorography, the gels
wereimpregnated with2,5-diphenyloxazole(2) before drying, andexposedtoX-rayfilm at-70°C. Protein standardsranging from 15,000to130,000daltonswere run oneachgel (6).
Preparation of samples for tryptic peptide
analysis. Polypeptideswereeluted from N,N-meth-ylenebisacrylamide cross-linkedgelsandprepared for trypsinization according totheprocedure ofVogtet
al. (37). N,N-Diallyltartardiamide cross-linked gel slicesweredissolvedin 2%periodic acid, accordingto
theprocedure of Gibson (13), andpreparedfor
tryp-sinizationasdescribedpreviously (10).Bothmethods
gaveidenticaltryptic methionine peptideprofiles for themajor capsid polypeptides. Trypsinization and ion-exchangechromatographyonChromobeadsP (Tech-nicon) were carried out accordingto theprocedure described by Vogt (37) and modified as described previously (10).
Immunological procedures.(i) Preparationof antisera.Antiseratoeachofthecapsid polypeptides
of HSV-1 were prepared in the following manner.
Capsid polypeptides prepared from unlabeled cells
were located in unstained gels by bracketing with labeled polypeptides. After electrophoresis, the gels
were washed with water, covered with plasticwrap, andplacedincontactwithX-rayfilmat4'C.Thegel
slicescontainingthepolypeptidesweremaceratedand mixed with Freundcomplete adjuvant.Approximately
538pgofcapsid proteinwassubjectedtoSDS-PAGE.
Rabbits received fourweeklyintramuscularinjections followed bya 10-day rest period. Each rabbit then receivedasubcutaneousinjectionandwasbled9days later. We estimate that the total amount ofprotein used for immunizationwas asfollows: NC-1, 324 ug;
NC-2,54pg; NC-3,4,64pg;NC-5, 59pg;NC-7, 37pg.
ImmunoglobulinGwaspreparedfrom each antiserum
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
STRUCTURE OF HSV-1 AND -2 CAPSID POLYPEPTIDES
asdescribedbyMontgomeryetal.(21).
(ii) Immunofluorescence. A modification of the indirect procedure describedbyReed etal. (27)was
used. Briefly,monolayersof KB cellsweregrownon
Lab-Tek slides(MilesLaboratories,Inc.)infected with HSV-1orHSV-2 andfixed with3.7%formaldehydeat 14 hpostinfection. The slideswerewashedwith
phos-phate-bufferedsaline,dehydratedfor7minat-20°C with acetone, and washed with phosphate-buffered saline. Thechamberswereoverlaid for30minat36°C
with50IL ofphosphate-bufferedsalinecontainingthe appropriate antibody. The slides werewashed three times with phosphate-buffered saline and incubated
with a mixture of fluorescein
isothiocyanate-conju-gatedgoat anti-rabbitimmunoglobulinG and rhoda-mine-conjugated bovine serum albumin. The latter
componentallowedus to visualize cells that didnot
specificallyreactwithantibody.
RESULTS
Analysisof HSV-1 and HSV-2capsids by
SDS-PAGE. Figure 1 shows a fluorogram of
electrophoretically separated 1 and
HSV-2 capsid polypeptides labeled with radioactive
methionine. HSV-1 capsids (track 1) consisted
of seven polypeptide bands with molecular
weights of 154,000 (154K) (NC-1), 50K (NC-2),
40K (NC-3),38K(NC-4),33K5),26K
(NC-6), and 12K(NC-7).HSV-2capsids (track2)also
consistedofsevenbands,and with three
excep-tions their molecularweightswereroughly
com-parable tothe corresponding polypeptidesseen
inHSV-1. These have also beendesignated
NC-1throughNC-7in orderofdecreasingmolecular
weight. Theexceptions were NC-1andNC-3,4.
Figure 1 shows that NC-3 of HSV-1 had the
same molecularweight (40K) as NC-4 of
HSV-2. NC-1 of HSV-2wasslightly largerthanNC-1
ofHSV-1, althoughthedifferenceisnotevident
inFig. 1.NC-6 in HSV-1 doesnotshow up well
in Fig. 1. This componentwas present in such
lowquantities that itwas not studied any
fur-ther.
Ourpresentstudies employed isopycnic CsCl
centrifugation for capsid purification. A
light-capsid population usually represented 90% and
aheavy-capsid population comprised10% of the
total recoverable radioactivity. In agreement
with previous studies (14-16), we found that
thesetwopopulations differed notonly in
den-sitybut intwoother respects:(i) electron
micro-scopic studies showed that, whereas heavy
cap-sids appeared to be impermeable to negative
stain, 80% of the light capsids usually were
permeable; (ii) SDS-PAGE analysis indicated
that polypeptides NC-3 andNC-4 were
under-represented inlight capsids. Inourhands,
HSV-2capsids could not be resolved into two
popu-lations.
To investigate whether the capsid
polypep-NCI
NC 2
NC3
C.
5-NC4
NC 5
-- 154,155
_. 50
-430
-.4 33
NC 6
NC 7
.. _--mm_m -412
1 2
FIG. 1. SDS-PAGE analysis of purified HSV-1
andHSV-2capsids.Fluorogramofa 10% N,N-dial-lyltartardiamide gel. Track 1,
[3H]methionine-la-beled HSV-1 capsids; track 2,
[35S]methionine-la-beledHSV-2 capsids. Molecular weights (x103) on theright.
tides ofHSV-1 and HSV-2 exhibited
imnmuno-logical cross-reactivity, antibodies were
pre-pared to individual gel bands of HSV-1 and
utilized inanimmunofluorescent assay.
Immunological studies of HSV-1 and
HSV-2capsids.Figure2shows
examples
of thetype ofimmunofluorescence observedwhen
an-tibody prepared to each of the HSV-1 capsid
polypeptides was reacted with HSV-1-infected
KBcells.Antibody preparedagainst 1,
NC-2, andNC-3,4 (the twocloselymigrating bands
were evaluated as a mixture) exhibited strong
nuclear reactions. Antibody against the
lower-molecular-weight capsid polypeptides (NC-5
and NC-7) showed much weakerbut still
posi-tivereactions.
Table 1 summarizes our observations of the
relative intensity and location offluorescence
seen when these antibodies were reacted with
HSV-1- and HSV-2-infected cells. Anti-NC-1,
-NC-2,and -NC-3,4 reacted against cells infected
witheither virus type. In each case the
homol-ogousreaction was stronger than the
heterolo-gous reaction. These results agree with those
reported for NC-1 (8, 16, 26) and NC-3 (16).
Anti-NC-5 and anti-NC-7 reacted weakly with HSV-1-infected cells and showed no detectable cross-reactivity with HSV-2-infected cells at the antibody levels employed (Table 1). Because the
correlation of immunological cross-reactivity
with structural similarity is difficult to predict
(17,20, 23),wenext examinedthe polypeptides VOL.34,1980
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.293.414.69.261.2]ofHSV-Iand HSV-2by tryptic peptide analysis.
Trypticpeptide analysis ofHSV-1 capsid
polypeptides. HSV-1 capsid polypeptides
la-beledwith either[35S]methionineor
[3H]methi-FIG. 2. Immunofluorescence analysis ofHSV- 1-in-fectedKB cells using antibodies tothe capsid
pro-teins. The concentrations of antibodies used in this
assayaregiven in Table1. (A) Anti-NC-1; (B) anti-NC-2; (C) anti-NC-3,4; (D) anti-NC-5; (E) anti-NC-7; (F)normalrabbitantibody, 1.3 mg/ml.
TABLE 1. Summary ofimmunofluorescenceanalysis
of HSV-1- andHSV-2-infected cells using antibodies
toHSV-1 capsid proteins
Relative immuno-Antiserum e i fluorescence
inten-prprdt ells infected Concnofan- Siy
prepared
tod
withHSV tibody'(mg/ sitybHSV-locapsi
typeml1)
Cyto-component: Nucleus plasm
NC-1 1 0.06 +++ ++
2 0.6 +++ +
NC-2 1 0.06 +±+ _
2 1.2 ++ _
NC-3,4c 1 0.18 +++ +
2 1.8 ++
-NC-5 1 0.68 ++
-2 1.7 -
-NC-7 1 0.44 +
2 1.6 -
I-"The concentration of antibody (immunoglobulin
G) utilized against HSV-1-infected cells is the lowest concentration yielding maximum fluorescence. For HSV-2, where the reaction waspositive,the concen-trationofantibodyis the lowest amountyielding max-imum fluorescence. When the reaction wasnegative
(NC-5 and NC-7 versus HSV-2), the concentration shown inthe tablewasthehighest tested.
bStrongest reaction indicated by +++; negative
reaction indicatedby-.
' This antiserum waspreparedagainstamixtureof
NC-3and NC-4.
oninewereseparated bySDS-PAGE. Each
com-ponentwasextracted fromthe gel, trypsinized,
and cochromatographed on acolumn of
Chro-mobeads P with thetrypticpeptidesof themajor
capsid polypeptide NC-1. Figure 3 shows that
thetrypticdigestofNC-1couldbe resolvedinto
approximately 14methionine peptides in
addi-tion totheflow-through fraction. Free
methio-nineelutedatfraction 38,aposition thatdidnot
correspondtoanyofthemajormethionine
pep-tides ofNC-1.ThetrypticpeptidesofNC-2(Fig.
4A) were resolved into a complex pattern that
was quite different from that of NC-1. Two
methionine peptides (at fractions 130 and 155)
clearly overlapped, suggesting that these two
peptides mightbestructurally similar. However,
it isalsopossible that the overlapping peaksin
this complex pattern represented structurally
differentpeptideswith similarelutionproperties
(12). Thetrypticpeptide
profile
ofNC-3,4 (Fig.4B) was different from that ofNC-1 or NC-2.
The
profile
ofNC-5wasstrikingly
different fromthat of any of the othercapsid polypeptidesand
wasnotable in that:(i)all ofthepeptides eluted
at acidic pH values, and (ii) one peptide
ex-hibitedsomevariabilityinelution. Thearrowin
Fig. 4Cindicatesthepositionof thispeak,which
was seen prominentlyinsome experiments and
was aminorpeakinotherexperiments.Finally,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.248.444.84.255.2]z
z
o 200
I
I-NC
I vs METHIONINE80
EU
0
70 X 60 <
0
.c
50 _E
llJ
40 Z
z 0
30 I
20 2
10 e 2
L
6
0 20 30 40 50 60 70 80 90 100 o 120 130 140 150 160 170 180 F R A C T O N N U M B E RFIG. 3. Tryptic peptide analysis of methionine-labeledNC-Iobtained from HSV-1 capsids. NC-I labeled
with[3H]methioninewasco-chromatographedon acolumn of Chromobeads P with[35S]methionine.Inthis
and eachof the following figures,the arrowrepresents the start of a basic wash of the column.
the tryptic peptide pattern forNC-7 (Fig. 4D)
was rather simple, consisting of one resolved
methionine peptide which eluted toward the
basic end of the pH gradient. This peptide did
not correspond to that of free methionine (cf.
Fig. 3), nor did it appear to correspond to a
methionine peptide ofany of the other capsid
polypeptides.ThusNC-7doesnotappeartobe
abreakdownproduct ofalarger capsid protein.
Thisseries ofexperiments demonstratedthat
eachof theHSV-1capsidpolypeptides appeared
to contain a characteristic and unique set of
tryptic peptides. Inone case (NC-2) therewere
twopeptidesthatmightcontain sequences
sim-ilar to those inNC-1.However,noneof the other
capsid polypeptides contained tryptic peptides
that coeluted with those of NC-1, and none
appearedtobe similartoanyothercapsid
poly-peptide.
Comparison of the tryptic methionine
peptides
obtained from HSV-1 and HSV-2capsid polypeptides.Intheseexperimentsthe
tryptic peptides of each HSV-1 capsid
poly-peptidewerecochromatographedwith the
tryp-tic peptides of the corresponding polypeptide
from HSV-2. If the high degree of intertypic
DNA homology (18) were reflected by similar
amino acid sequences in thecapsidpolypeptides,
we would expect to see overlaps (complete or
partial) inthepeptideprofiles.Figure 5A shows
that the overall elution profiles for NC-1 of
HSV-1 and HSV-2, though not identical,
ap-peared quitesimilar,and severalpeaksappeared
tooverlap. The second casewhere there was a
strong similarity is shown in Fig. 5B. The
me-thionine
profile
ofNC-7 from HSV-1wasessen-tially the same as that of NC-7 from HSV-2.
However,thesimplicityof the methionine
pep-tide
profile
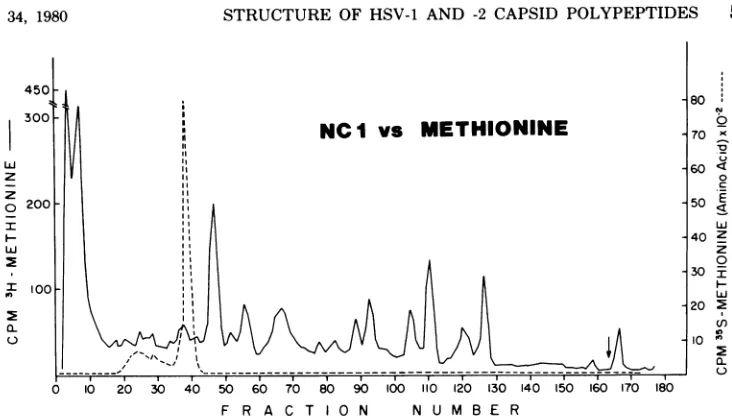
forNC-7made it difficulttoconcludethat this protein is identical in the two virus
types. Figure 6shows that the
arginine peptide
profile
forNC-7was verysimilar butnotiden-tical in HSV-1 and HSV-2. The data in Fig. 5
and6suggestthat there may beahigh degreeof
intertypic homology ofamino acid sequence in
thesetwopolypeptides.
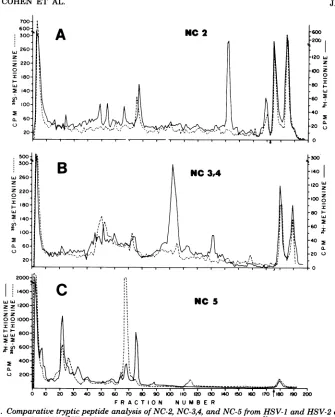
Figure 7 shows the elution profiles for three
capsid polypeptidesofHSV-1 and HSV-2 that
exhibited fewer overlapping peptides. The
re-solved methionine peptides of NC-2 isolated
from HSV-1 and HSV-2 (Fig. 7A) were clearly
different,eventhoughthepolypeptideshad the
sameapparentmolecularweight by SDS-PAGE
and cross-reacted immunologically.
Parentheti-cally,theNC-2sofHSV-1 andHSV-2 contain a
significant amount of radioactive label which
elutes at a basic pH. The methionine peptides
of the NC-3,4 region (Fig. 7B) of HSV-1 and
HSV-2were different, despiteintertypic
immu-nological cross-reactivity. However, in one
re-gion ofthe elution profile (fractions 40 to 80),
the patterns looked similar, and it is possible
that the common antigenic site(s) might be
found in thisregion. Figure 7C shows that
NC-5 of HSV-1 had two methionine peptides that
coeluted withpeptidesof NC-5 of HSV-2. In this
case, the possibility for structural
similarity
shown by peptide analysis was not consistent
with the negative immunological data. In
sum-mary,Fig.7demonstrates that there are
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.71.437.47.256.2]z
0
ni
e1
n-z
z
0
n e. ri
CL
L'i
z z 0
x
ui
2
(f) I
2
(L
w
z o
tL
u 3 20 30 40 50 60 7o 80 90 00 110 20 3041i400 60i70 180 190
F R A C TI 0 N N U M BE R
FIG. 4. Tryptic peptide analysisofmethionine-labeled HSV-1capsidpolypeptides. Trypticpeptides of NC-1 ( ) labeled with either[35S]methionineor[3H]methioninewerecochromatographedon aChromobeads P column with[3H]methionine- or[35S]methionine-labeledpeptides(----)of:(A)NC-2, (B)NC-3,4, (C)NC-5,
or(D) NC-7. In eachcasetheleft-handordinate representsNC-1. Thearrowin (C) (fraction 58)marksthe
position ofthe variablepeptide.
tantstructural differences betweenpolypeptides 7s of HSV-1 and HSV-2 had identical
methio-with similar molecularweights,andFig.5,6, and nine tryptic peptides andvery similar arginine
7demonstrate thatstructural similaritiesordif- peptide profiles. Each of these components
ferences maynotbe consistent with
immunolog-
might be a host protein or a highly conservedical data. viral component.
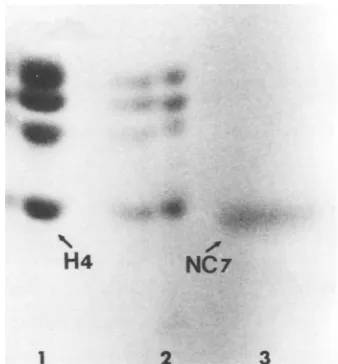
Figure
8 shows that NC-7 ofIs NC-7 the host cell histone H4? The NC- HSV-1migratedinSDS-PAGE
gels
withamo-J. VIROL.
z 0
I7
LL
Z
I
PI
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.125.384.53.527.2]o o 20 30 40 50 90 120 30 40 ISO 60 170 T180 190 200
F R A C TI0 N N U M B E R
FIG. 5. Comparativetryptic peptide analysis ofNC-I and NC- 7 isolatedfromHSV-I andHSV-2capsids. Cochromatography of: (A)[3H]methionine-labeledNC-IfromHSV-I( )with[35S]methionine-labeled
NC-1from HSV-2 (---); (B) [35S]methionine-labeledNC-7fromHSV-1 ( )with[35S]methionine-labeledNC- 7
from HSV-2 (---).
600F
0 10 20 30 40 50 60 70 80 90 100 110 120 130 140 150 160 170 FRACTION NUMBER
FIG. 6. Comparativetryptic peptide analysis ofNC-7from HSV-1 and HSV-2 capsids. Cochromatography of[4C]arginine-labeledpeptides fromHSV-2( )with[3H]arginine-labeledpeptides from HSV-1 (---).
LLJWLA zz
zz
0z z0
LLJ wL
o o
in
to
I.,
LAJ
z z
z z
0 0 o
oT
II
I(0)
450 _
300 _
Z Z
ZZ
3
3t
L) I
a-u 15
:~~~~~~~~~~~~~~~1
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.68.438.54.393.2] [image:7.503.69.437.460.628.2]528 COHEN ET AL.
a-w
z z
0
tL l0~
zI
z z
z z 001,
I x
I
ww
tLA
a-A
NC 2'r
Duj
C)LZz
0
I
I
0.
IO
I
z
Z
)z
I -o~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~o
I Q.
C.,
NC S
20 30 40 50 60 70 80 90 00 110 120 30 140 I50 60 17rt 90 200
[image:8.503.90.425.53.470.2]F R ACT 0N NU MB E R
FIG. 7. Comparative tryptic peptide analysis of NC-2, NC-3,4,andNC-5 from HSV-1 andHSV-2capsids.
Cochromatography of (A) ['Himethionine-labeledNC-2from HSV-1 ( )with[35S]methionine-labeled NC-2fromHSV-2(---); (B)['H]methionine-labeledNC-3,4 from HSV-1 ( )with["S]methionine-labeled
NC-3,4fromHSV-2(---); (C) ['Himethionine-labeledNC-5from HSV-1( )with['5S]methionine-labeledNC-5
fromHSV-2(---).
bilitysimilartothat of H4histone ofuninfected
KBcells. TocompareNC-7with H4wedecided toexamine thearginine tryptic peptidesofthese
two polypeptides. KB cells were labeled with
[3H]arginine for 18 h, a nuclear fraction was
prepared,and thehistone fraction wasisolated
(1). HSV-1 capsids were prepared from
[14C]-arginine-labeled KB cells. These preparations
were subjected to SDS-PAGE, and the bands
corresponding to KB-H4 and NC-7 were
ex-tracted from the gel, oxidized, trypsinized, and
cochromatographed onChromobeads P. Figure
9 shows that thetryptic peptide profilesfor
KB-H4and NC-7aredifferent.Fromthisexperiment
we conclude thatNC-7 isnotthe KB host his-toneH4.
DISCUSSION
This study was designed to examine
immu-nologicaland structuralpropertiesof thecapsid
polypeptides of HSV-1 and HSV-2. We found
that each of the HSV-1 capsidpolypeptides
ex-tracted from SDSgelswas capableofinducing
antibody which reacted with HSV-1-infected
cells.Inaddition, antibodytothe HSV-1capsid
polypeptides NC-1, NC-2, and NC-3,4 also
re-NC
3,4
J. VIROL.
B
on November 10, 2019 by guest
http://jvi.asm.org/
acted with HSV-2-infected cells. Similar results
have beenreported previously for NC-1 (8, 16)
and NC-3 (16). Since the antibodies were
pre-pared to SDS-denatured proteins, it ispossible
thatsites notnormallyavailable toprovokean
immuneresponsewereexposedand became
im-munogenic. Thus, as wasshown in the case of
simian virus 40 andpolyoma virus (32) as well
asforadeno-associatedvirions (17),theserawe
prepared may turn out to have greater
cross-reactivity than sera prepared against
undena-tured polypeptides. The immunological
cross-reactivity that we observed reflects intertypic
structural similarities, but the extent of these
similarities is difficult to assess. It is
possible
thatstructuralhomologybetween
polypeptides
H4
1
NC
NC7
[image:9.503.65.234.245.427.2]2 3
FIG. 8. SDS-PAGE analysis of NC-7 from HSV-1 and KB cell histones. Fluorogram of an 18o
N,N-methylenebisacrylamide-linked gel. Only the lower-molecular-weight region of the gel is shown. Track 1,
[3H]arginine-labeled histones extracted from
unin-fected KB cells; track 2,same astrack1, half as much protein; track 3, ["4C]arginine-labeled NC-7 from HSV-1.
may actually be great even though the sites
responsible for immunological cross-reactivity
are hidden in the native protein. For
example,
a-fetoprotein
andserumalbumin haveamarkedstructural similarity but do not cross-react
un-less antibody prepared against unfolded
poly-peptide chainsisutilized (30).
Trypticpeptideanalysiswasutilizedto
estab-lish two points: (i) whethereach ofthe HSV-1
capsid polypeptidesiscomposedof auniqueset
ofmethionine-labeled peptides; and (ii) the
de-gree of structural relatedness between
corre-sponding HSV-1 and HSV-2 capsid
polypep-tides. Ourdata indicatethat thecapsid
compo-nentsobservedonSDS-PAGEare notgenerated
as a result ofproteolytic cleavage of the major
capsid polypeptide (NC-1),andeach appears to
consist ofa unique setof methionine peptides.
Inoneinstance(NC-1 and NC-2 of HSV-1, Fig.
4A), severalmethionine peptides areshared. It
is important to bear in mind that because the
methionine peptide patterns are complex, it is
possible that there was random overlapping of
structurally different peptides (12). Further
studies will be required to determine whether
these peptides are truly shared among viral
structural components. The degree of
related-ness between the corresponding HSV-1 and
HSV-2 capsid polypeptides is more
difficult
toassess. It is clear that in all cases except one
(NC-7), the intertypic methionine peptide
pro-files were notidentical inspite of the factthat
the proteins have similar apparent molecular
weight values. In thecase ofNC-7,
small
differ-ences in the arginine peptide profileswere
de-tected. Thus, we can concludethat there is no
complete intertypic conservation of amino acid
sequence inproteinsthatpresumably have
sim-ilar structural functions.
One explanation for the differences in the
tryptic peptide patternsfor HSV-1 and HSV-2
capsidpolypeptidesis thatthey are due to
intra-typic variability. Such variability hasbeen
dem-onstratedfor certainHSV-1virionpolypeptides
2'
, 21
Z 5 II
0
c
I
a-0
-4 '-
-250
200w.
z
150 Z
0.
S02
CL
0 to 20 30 40 50 60 70 80 90 100 110 120 130 140 150 160 170 480
FRACTION NUMBER WASH
FIG. 9. Comparative tryptic peptide analysis ofNC- 7fromHSV-1 and KB cell histone H4. Cochromatog-raphy of['4C]arginine-labeledpeptides fromNC- 7( ) with[3H]arginine-labeledpeptides from H4 (---). VOL. 34, 1980
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.503.75.438.518.627.2](24) but not for all components (25). Since we
examinedonlyonestrain of HSV-1 andHSV-2,
we cannotconclude thatnointratypicvariability
exists.Some of the differencesweobservedmay
be due to this phenomenon rather than to
inter-typic differences. Assuming, however, that the
differences we observed in the tryptic peptide
profiles
are due tointertypic rather thanintra-typic variability, tryptic peptide analysis could
beutilizedto verify the parental origin of
poly-peptides in HSV-1 xHSV-2 recombinants. It is
interesting that the
intertypic
forms of NC-1appear to contain a significant number of
pep-tidesin common.Furthermore,thearginineand
methionine peptides of NC-7 are closely
con-served between thetwo virustypes. We
specu-late that some of thesepeptidescontain highly
conserved aminoacid sequences.
HSV-specific polypeptides smaller than 25K
havebeen described (19, 28), but little is known
about their structure or functional properties.
Low-molecular-weight polypeptides isolated
from other viruses such as the
papovaviruses
and adenovirus
apparently
areimportant
com-ponents inthe condensation of viral DNA.
Pa-povaviruses,
forexample,
utilize host histonesfor this purpose (31), whereas adenovirus core
proteins arebasicand
presumably
function inasimilar manner (11). We have shown here
by
trypticpeptide analysisthatNC-7 isnottheKB
H4histone. Y.Roth (submittedfor
publication)
haspresented datathat NC-7couldbe extracted
with acid from infectedcell
nuclei,
chromatinofHSV-1-infected
cells,
orpurified
HSV-1capsids.
Low-molecular-weight basic
proteins
haveal-ready been reportedto be present in
herpesvi-ruses (4, 34), and it is
possible
that NC-7 is abasic
capsid protein
involvedintheeventslead-ing tothepackaging of HSV DNA.
ACKNOWLEDGMENTS
Thisinvestigationwassupported byPublic Health Service grantDE-02623 from the National Institute ofDental Re-searchandbygrantno.3.706-0.76SRfrom theSwissNational Science Foundation. Partofthisworkwas donebyG.H.C. duringascholarlyleaveatthe Swiss Institutefor Experimen-tal CancerResearch, Lausanne,Switzerland.
We thankWesleyWilcoxforhelpinpreparation of this manuscript, Madeline CohenandCassandraHydrean-Stern for theirexcellent technicalassistance,Roman Klemenz for manyhelpfuldiscussions,andYvonneRothfor hergenerous helpwith the histoneportionofthispaper.
LITERATURE CITED
1. Bonner, J., G. R. Chalkley, M. Dahmus, D. Fam-brough, F.Fujimura,R.-C. C.Huang,J. Huber-man, R.Jensen,K.Marushige,H.Ohlenbusch,B. Olivera,andJ.Widholm. 1968.Isolation and char-acterization ofchromosomalnucleoproteins. Methods Enzymol. 12B:3-65.
2. Bonner, W.M.,and R. A.Laskey.1974.Afilm detection methodfortritium-labeledproteinsand nucleicacidsin
polyacrylamide gels.Eur. J. Biochem. 46:83-88. 3. Cassai,E.N.,M.Sarmiento,and P. G.Spear. 1975.
Comparisonof the virion proteins specified by herpes simplexvirustypes1and2.J. Virol.16:1327-1331. 4. Chantler,J.K.,and W.S.Stevely.1973.Virus-induced
proteinsinpseudorabies-infectedcells.I. Acid-extract-ableproteinsof thenucleus. J. Virol. 11:815-822. 5. Cohen, G.H.,M.N.Factor,and M. Poncede Leon.
1974.Inhibition ofherpes simplexvirus type2
replica-tionbythymidine.J.Virol. 14:20-25.
6. Cohen,G.H.,M.Katze,C.Hydrean-Stern,and R. J. Eisenberg.1978.Type-commonCP-1antigenofherpes simplex virus is associated with a 59,000-molecular-weightenvelope glycoprotein.J.Virol.27:172-181. 7. Cohen,G.H.,M.PoncedeLeon,and C. Nichols.1972.
Isolation ofa herpes simplex virus-specific antigenic fraction whichstimulates theproductionofneutralizing antibody.J.Virol. 10:1021-1030.
8. Courtney,R.J.,and K.L. Powell.1975.Immunological
and biochemical characterization ofpolypeptides in-ducedby herpessimplexvirustypes1and2,p.63-73. InG.de-The,M.A.Epstein,and H.ZurHausen(ed.), OncogenesisandherpesvirusesII.InternationalAgency
forResearchonCancer, Lyon.
9. Dreesman,G.R.,J. R.Suriano,S. R.Swartz,and R. M. McCombs. 1972. Characterization ofthe herpes
virion. I. Purification and amino acidcompositionof
nucleocapsids. Virology50:528-534.
10. Eisenberg,R.J.,C.Hydrean-Stern,and G. H. Cohen. 1979.Structuralanalysisof precursor andproductforms of type-commonenvelope glycoproteinD (CP-1 anti-gen) ofherpes simplexvirustype 1.J. Virol. 31:608-620.
11. Everitt, E.,and L.Philipson.1974.Structural
polypep-tides of adenoviruses. XI. Purification of three low molecularweightvirionproteinsof adenovirustype2 and theirsynthesis during productiveinfection. Virol-ogy62:253-269.
12. Fey,G.,J. B.Lewis,T.Grodzicker,and A. Bothwell. 1979.Characterization ofafusedproteinspecifiedby
theadenovirustype 2-simianvirus40hybrid Ad2+NDl
dp2.J. Virol.30:201-217.
13. Gibson,W. 1974.Polyomavirusproteins.Adescription
of thestructuralproteinsofthe virion basedon
poly-acrylamide gel electrophoresis and peptide analysis.
Virology62:319-336.
14.Gibson, W., and B. Roizman.1972.Proteinsspecified by herpes simplex virus. VIII. Characterization and
compositionofmultiplecapsidforms ofsubtypes1and 2.J.Virol.10:1044-1052.
15. Gibson,W.,and B. Roizman. 1974. Proteinsspecified
byherpessimplexvirus. X.Stainingandradiolabeling
propertiesof Bcapsidand virionproteinsin
polyacryl-amidegels.J.Virol. 13:155-165.
16. Heilman,C.J., Jr.,M.Zweig,J.R.Stephenson,and B. Hampar. 1979. Isolation ofanucleocapsid
poly-peptideofherpessimplexvirustypes1and2possessing
immunologicallytype-specificandcross-reactive deter-minants. J. Virol. 29:34-42.
17. Johnson,F. B.,N. R.Blacklow,andM. D.Hoggan. 1972. Immunological reactivity of antisera prepared
against the sodium dodecyl sulfate-treated structural
polypeptidesofadeno-associatedvirus.J. Virol. 9:1017-1026.
18. Kieff,E.D.,B. Hoyer,S.L.Backenheimer, andB. Roizman.1972.Geneticrelatedness oftype1andtype 2herpessimplexviruses.J. Virol. 9:738-745. 19. Marsden, H. S., I. K. Crombie, and J. H.
Subak-Sharpe. 1976.Controlofprotein synthesisin
herpes-virus-infected cells:analysisofthepolypeptidesinduced bywildtypeand sixteentemperature-sensitivemutants
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
of HSV strain17. J. Gen.Virol.31:347-372.
20. Marsden, H. S., N. D. Stow, V. G. Preston, M. C. Timbury, andN. M.Wilkie.1978.Physical mapping ofherpes simplex virus-induced polypeptides. J. Virol. 28:624-642.
21. Montgomery, P. C., K. J. Dorrington, and J. H. Rockey.1969.Equine anti-hapten antibody.The
mo-lecular weightsof thesubunits of equine immunoglob-ulins. Biochemistry8:1247-1258.
22. Morse,L.S.,L.Pereira,B.Roizman,and P. A. Schaf-fer. 1978. Anatomy ofherpes simplex virus (HSV) DNA.X. Mapping ofviralgenesbyanalysis of polypep-tidesandfunctionsspecified byHSV-1xHSV-2
recom-binants.J.Virol. 26:389-410.
23. Olshevsky, U.,and Y.Becker. 1970. Herpes simplex virus structural proteins. Virology 40:948-960. 24. Pereira, L.,E.Cassai,R.W.Honess,B.Roizman,M.
Terni, andA. Nahmias. 1976.Variabilityinthe
struc-tural polypeptides ofherpes simplex virus 1 strains: potential applicationinmolecularepidemiology. Infect. Immun.13:211-220.
25. Powell,K.L.,R.Merkovic,and R. J.Courtney.1977. Comparative analysisofpolypeptides induced bytype 1andtype2strains ofherpessimplexvirus.
Intervirol-ogy8:18-29.
26. Powell,K.L.,and D. H. Watson. 1975. Somestructural antigens of herpes simplexvirustype1.J. Gen. Virol. 29:167-178.
27. Reed,C.L.,G. H.Cohen,and F.Rapp.1975.Detection of a virus-specific antigen on the surface of herpes simplex virus-transformed cells. J. Virol. 15:668-670. 28. Robinson,D.J.,and D. H. Watson. 1971.Structural proteinsofherpes simplexvirus. J.Gen.Virol. 10:163-171.
29. Roizman,B. 1978. Theherpesviruses,p.769-848.InD.
P. Nayak (ed.), Themolecular biology of animal viruses, vol.2.Marcel Dekker Inc., New York.
30. Ruoslahti, E., and E. Engvall. 1976. Immunological crossreaction between alpha-fetoproteinandalbumin. Proc. Natl.Acad.Sci. U.S.A. 73:4641-4644.
31. Sambrook,J. 1978. The molecularbiologyof the
papo-vaviruses,p.589-672.In D. P. Nayak (ed.), The
molec-ularbiologyofanimal viruses,vol. 2. Marcel Dekker Inc.,NewYork.
32. Shah,K.V.,H.L.Ozer,H.N.Ghazey, andT.J.Kelly, Jr.1977. Common structuralantigen of papovaviruses ofthe simian virus40-polyoma subgroup. J. Virol.21: 179-186.
33. Spear,P.G.,and B. Roizman. 1972.Proteinsspecified by herpes simplex virus. V. Purification and structural polypeptides of the herpesvirion. J. Virol.9:143-159. 34. Stevens, J. G., G. J. Kado-Boll, and C. B. Haven.
1969. Changes in nuclear basic polypeptides during pseudorabiesvirus infection.J. Virol.3:490-497. 35. Vernon, S.K., W. C. Lawrence, and G. H. Cohen.
1974.Morphologicalcomponentsofherpesvirus. I. In-tercapsomeric fibrils and the geometryof the capsid. Intervirology 4:237-248.
36. Vernon, S. K., W. C. Lawrence, G. H. Cohen,M. Durso, and B. A. Rubin. 1976.Morphological
com-ponentsof herpesvirus. II. Preservationof virus during negative staining procedures. J. Gen. Virol.31:183-191. 37. Vogt,V.M.,R.Eisenman, and H. Diggelmann.1975. Generation of avian myeloblastosis virus structural polypeptides by proteolytic cleavage of a precursor
polypeptide.J.Mol. Biol.96:471-493.
38. Zweig,M.,C. J.Heilman,Jr.,and B.Hampar.1979. Identification ofdisulfide-linked protein complexes in thenucleocapsidsofherpessimplex virustype2.
Virol-ogy94:442-450.



![FIG. 6.of [4C]arginine-labeledpeptides Comparative tryptic peptide analysis of NC-7 from HSV-1 and HSV-2 capsids](https://thumb-us.123doks.com/thumbv2/123dok_us/1496527.102345/7.503.68.438.54.393/fig-arginine-labeledpeptides-comparative-tryptic-peptide-analysis-capsids.webp)