JOURNAL OF VIROLOGY, Feb.1994, p. 787-796 Vol. 68, No. 2 0022-538X/94/$04.00+0
Copyright C 1994, American Society for Microbiology
The Genomic Instability Associated with Integrated Simian
Virus 40 DNA Is Dependent on the Origin of Replication and
Early Control Region
DEBORAHJ. HUNTER*AND ELIZABETH G. GURNEY Departmentof Biology, University of Utah, Salt Lake City, Utah 84112
Received 16 April 1993/Accepted 26 October 1993
DNArearrangements in the form of deletions and duplications are found within and near integrated simian virus 40 (SV40) DNA in nonpermissive cell lines. We have found that rearrangements also occurfrequently
with integratedpSV2neo plasmid DNA. pSV2neo contains the entireSV40 control region, including the origin
ofreplication, both promoters, and theenhancer sequences. Linearized plasmid DNA was electroporated into Xl, anSV40-transformed mouse cell line that expresses SV40 large T antigen (T Ag) and shows
very
frequent rearrangements atthe SV40 locus, and intoLMtk-, a spontaneously transformed mouse cell line that contains no SV40 DNA. Stability was analyzed by subcloning G-418-resistant clones and examining specific DNA fragments foralterations in size. Five independentXl clones containing pSV2neo DNA were unstable at both the neo locus and the T Ag locus. By contrast, four Xl clones containing mutants of pSV2neo with smalldeletions in the SV40 core origin and three Xl clones containing a different neo plasmid lacking SV40 sequences were stable at theneo locus, although they were still unstable at theTAg locus. Surprisingly, five
independent LMtk- clones containingpSV2neo DNA were unstable at the neo locus. LMtk- clones containing origin deletionmutants were more stable but were not as stable as theXl clones containing the same plasmid
DNA.We concludethat the SV40 origin of replication and early control region are sufficient viral components
for thegenomicinstabilityat sitesofSV40 integration and that SV40TAg is notrequired.
Simian virus 40 (SV40) DNA integrated into the host genome of nonpermissive cells shows ongoing genomic
insta-bilitycharacterized by both duplications and deletions of SV40 and host flanking DNA (4, 24, 27, 42, 43, 57, 64). In the overreplication model, based on the onion skin model (6, 58) andfrequently invoked to explain the instability, SV40 largeT
antigen (TAg) would bind to the SV40 core origin and initiate unscheduled DNA replication, leading to overreplication of theorigin andflankingsequences. The rearrangementswould then be the result of recombination between the amplified
DNA strands. Alternatively, the viral DNA may be excised from the chromosome, replicate in the form of an extrachro-mosomal element, and reintegrate into the host DNA at the
same or a newchromosomal site (11, 15, 52). However, SV40 extrachromosomal elements havenotbeenisolated frommost
nonpermissive cell linescontaining integrated SV40 DNA (3, 4, 27,57).
Inthe cell line XI,isolatedas arearranged subclone of the BALB/3T3 cell line SVT2, the SV40DNA ishighly unstable, with new rearrangements detected in almost every subclone
(24).The rearrangementsareobservedmorefrequently inthe
DNAsequencesclosest to the SV40origin of replication(2,3). Theenhancedinstability ofXl cells,ascompared with that of SVT2 andother celllines withintegratedviralDNA,provides
an opportunityto readilyexamine thegenomicinstability.
In this study we have examined the dependence of the genomic instability of SV40 DNA in Xl cells and another
nonpermissive cell line on theSV40originofreplication and SV40 T Ag. We first determined the stability of the SV40
wild-type origin of replication and auxiliary sequences froma
reporterplasmid atseveral sites ofintegration inXl cells and
*Correspondingauthor. Mailingaddress: Departmentof
Microbi-ology and ImmunMicrobi-ology, University of Michigan, Ann Arbor, MI 48109-6020. Phone: (313)936-0876. Fax:(313) 764-3562.
compared this with the stability ofareporter plasmid witha
mutated SV40originregion. Thiswasaccomplished by
trans-formingXl cells withpSV2neo,which hasthewild-typeSV40
origin ofreplication, or with pSV2oriA plasmids,which have small deletions in the SV40 core origin, and monitoring for restriction fragment length polymorphismsby usingSouthern blots. Second, we tested the role of T Ag in inducing
rear-rangements byanalyzing pSV2neo in LMtk- cells,which do
notexpressTAg. Ourresults indicate that the presence of the SV40 core origin ofDNAreplication along with the flanking
auxiliary regions and early promoter is sufficient to induce genomic instability.
MATERIALSANDMETHODS
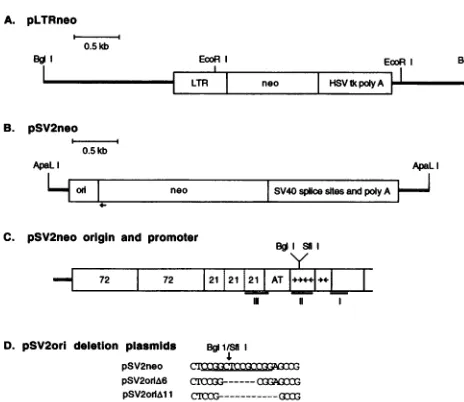
Plasmids used in cloning studies. The plasmid pLTRneo (Fig. 1A) contains the long terminal repeats (LTRs) of the aviansarcomavirus, the Tn5neogene,theherpes simplexvirus thymidine kinase polyadenylation sequences, and pBR322
vectorsequences. pLTRneo wasconstructed frompRMH140 (29) by deleting the SV40 DNA originally found 3' to the herpes simplex virus thymidine kinase polyadenylation
se-quences.
pSV2neo (63)wasobtained from the American Type
Cul-tureCollection(ATCC37149).Asillustrated inFig.1B andC,
pSV2neo contains the 342-bp PvuII-to-HindIII SV40 DNA
region consisting of theorigin ofDNAreplication, earlyand
late promoters, andenhancer sequences (SV40nucleotides 1
to270 and5171 to5243)andasecondSV40fragmentwith the
early polyadenylation signal and splice sites. Both 5' and 3' untranslated TnS neo sequences, which are not found in pLTRneo, arepresent inpSV2neo, accountingforthe differ-encein sizeof theneo regionfound in thetwoplasmids.
Thereplicationactivityof theSV40originin
pSV2neo
was inhibitedbymakingsmall deletions inTAgbinding
domainII,787
on November 9, 2019 by guest
http://jvi.asm.org/
0 1 kb
1....1
b ori
;f/,s//X t Ag
[image:2.612.65.297.69.270.2]I
, ,f TAgFIG. 2. Map ofintegrated Xl SV40 DNA. The integratedSV40 APaLI DNAinXlcellsis indicatedby thebars,and theflankingmouseDNA J is indicated by the line. The DNA regions corresponding to the completecoding regionsfor largeand smallTAg areindicatedby the hatched bars under the restriction map. ori marks thesite of theSV40 originofreplication, enhancers, andearlypromoter.TheHindIII(H) andBglI (b) sites present in theintegrated SV40 DNAandflanking mouse sequences are marked(24).
D. pSV2ori deletion plasmids BgISf1i pSV2neo
pSV2orIA6 C1X3G---QOflG pSV2MolM1 CrOG---(3
FIG. 1. Plasmidmaps.(A) The 4.7-kb pLTRneo plasmidlinearized attheunique BglI site. The RoussarcomavirusLTRs, TnSneogene,
andherpes simplex virusthymidine kinase (HSV tk) polyadenylation signalare indicated. ThepBR322vectorsequences areillustratedby theheavy lines. (B) The 3.97-kb ApaLI fragment of pSV2neo, illus-trating the SV40 control region(ori), theTn5neogene,theSV40 early splice sites and polyadenylation region,andpBR322vectorsequences.
The location of the primer used in sequencing the pSV2oriAplasmids is marked by the arrow under the neo gene. The pBR322 vector
sequences areillustratedbytheheavy lines. (C) Enlargedview ofthe SV40 control region.The 72-bpenhancers,the 21-bprepeats, andT Ag bindingsites III and I constitute theSV40 earlypromoterregion. TheSV40coreorigin consists of the AT-rich region,TAg binding site II (box with the BglI site andSfiI sites), and the early palindrome (imperfect inverted repeat)region. T Ag binding sitesareunderlined andlabeledI, II,andIII.(D)The DNAsequenceof theSfiI-BglI site
in theSV40 origin of replicationinpSV2neo.TheSfilrecognitionsite
(CCGGN5CCGG) is underlined, and the SfiI-BglI cleavage site marked. The nucleotides deletedin pSV2oriA6 and pSV2oriAll are
indicated.
usingthe Si nucleasedeletion method ofGluzmanet al.(22).
Deletions in the origin were generated first by linearizing
pSV2neowith
Sfil,
whichwasfollowedby S1nucleaseattackatthe SfiI-BglI site (Fig. 1C). After Si nuclease digestion and
religationtogivecircularmolecules,plasmids with deletionsin theorigin region (pSV2oriA) wereidentifiedby screeningfor the loss of the BglI site in the SV40 origin. Twenty-two plasmids screened by BglI restriction mapping retained the BglI cleavage site in the pBR322vectorbutnot in theSV40 origin. ThepSV2oriAcandidateswere tested forSV40 origin
replication activity in Cosl cells, as described below. Two
replication-defective plasmids, pSV2oriA6 and pSV2oriAl11,
were sequenced acrosstheoriginwith Sequenase 2.1 (United
States Biochemical, Cleveland, Ohio) by using the modified protocol ofSchuurmanand Keulen (62). The oligonucleotide primerusedforsequencing hybridizedwithin theneogeneand is represented by the arrow in Fig. 1B. A 6-bp deletion spanningthe
Sfi1
sitewasfound inpSV2oriA6 (Fig. 1D), whilepSV2oriAl11 had an 11-bp deletion. In addition, a second deletion ofapproximately 250 bp ispresentin the 5'
untrans-latedregion ofneoin pSV2oriA6.
The lacZexpressionvectorpM4-lacZ, usedtooptimize cell electroporation conditions, was kindly supplied by Chuxia
DengandMario Capecchi (Department ofHumanGenetics, University ofUtah, Salt Lake City).
Plasmid DNAwasprepared byalkalinelysisandpurified by
polyethylene glycol precipitation (59). The electroporation conditions usedtotransform Escherichiacoli MC1061F' with
supercoiled plasmids, usingtheBio-Rad Gene Pulser Appara-tus,were 2.45 kV,400Ql, and 25 ,uF. Competent MC1061 F'
cellswereprepared by the method ofDoweretal. (18).
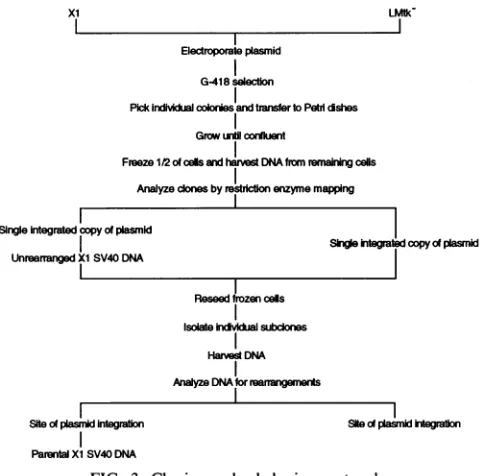
Cellgrowth conditions and clone selection. Xl cells, illus-trated inFig. 2, containoneintegratedcopyofSV40 consisting
of the SV40 early coding region, early promoter, enhancers,
core origin, andasmall duplication,which containsa partial,
scrambledsection of theearlycontrolregionand TAgexon 1
(3, 24). Xl cells were cultured as previously described (24).
The geneticin (G-418; GIBCO) concentration used to select Xl clones containing integrated copies of the reporter plas-midswasdetermined by assayingXlcells for G-418 sensitivity.
Nospontaneousneomycin-resistant Xl cellswereobserved in
2 x 107cellsat 400,ug of G-418 perml.
The clones Xl pSV2neo2O (neo20) and Xl pSV2neol7 (neol7) were selected with 600 ,ug of G-418 per ml in
monolayer culture. The clone Xl pLTRneoA26 (LTRA26)
and the remaining Xl clones were selected with 400 pug of G-418 perml in 0.35% agarose. Subclones ofneo20, neol7,
and LTRA26 were grown in 0.35% agarose either with or
without G-418,asindicated.Individual subcloneswerepicked
from the agarose and seeded in monolayer cultures with or
without 300 ,ug of G-418 per ml. All other Xl clones were
subclonedasmonolayercolonies in the absence ofG-418.The
protocol forisolating individual clones and subclones is
out-lined inFig.3.
Thetransformedmousefibroblastcell line LMtk- (34)was
culturedat37°Cin Dulbecco'smodified Eagleminimal
essen-tial medium supplemented with 1 mM sodium pyruvate and 5% calfserum(DV-5). The G-418 toxicity curvefor
LMtk-cells was determined, and clones containing integrated
plas-midswere selected in400 pugofG-418perml. LMtk- clones weregrown as monolayersin the presenceof G-418.
LMtk-subcloneswere isolatedasmonolayercolonies in the absence
ofselection.
Cosl cells (21)were grownat 37°Cin Dulbecco's modified
Eagle minimal essential medium supplemented with 1 mM sodium pyruvateand 10% fetal calfserum(DV-10).
Eukaryotic-cell electroporation. Xl cellswere transformed with linearized plasmids by using the Bio-Rad Gene Pulser Apparatus.The initial electroporation conditions forXl cells
wereselectedby determining the 50% survivalrateof the cells, followingelectroporationeither inphosphate-bufferedsucrose
(PB sucrose) (272mMsucrose,7 mMNaPO4 [pH 7.4],1 mM
MgCl2) or in Dulbecco's phosphate-buffered saline without
Ca2'
andMg2+ (PBS) (140mMNaCl,2.7 mM KCl,8.1 mMNa2HPO4, 1.5 mM KH2PO4). Xl cells were trypsinized,
A. pLTRneo
0.5kb
Bo EcoR
II
T«
B. pSV2neo
0.5kb
EooR Bg[ 1 neo HSV Epoly.AI
ApaL
L~
b H H bH H H
I I - II I I
I1
neo0 SV40splicesltesandpolyAAC. pSV2neo origin and promoter
B soI y
- 72 72 I21.21.21 AT .+.. .
I 11 11
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.326.555.72.134.2]SV40 CONTROL REGION CONFERS GENOMIC INSTABILITY 789
SingleIntegratedcopy ofplasmld
UnreamangedXISV40 DNA
Siteof pasdintegration
ParentalXiSV40 DNA xi
S e jtegr copyf plasnid
[image:3.612.58.299.72.310.2]SiteO plaow ntegratio
FIG. 3. Cloning and subcloning protocol.
washed in eitherPB sucrose or PBS, and resuspended in the appropriatebufferto1.25 x 107cellsperml.Aliquotsof cells
(0.8 ml) were transferred to 0.4-cm Gene Pulser cuvettes (Bio-Rad),and 50-Velectroporation incrementswereapplied
to sequentialcuvettes. The cellswere incubatedonice for 10
minafterelectroporationandplatedasmonolayersat37°Cfor 24to48 h priorto determination of thepercentcell survival.
The electroporation conditions for cells resuspended in PBS
wereoptimized in atransient expressionassayusing the lacZ expression vector pM4-lacZ and screening for blue colonies
(60).
Xl clones with integrated plasmid DNA were obtained
following the electroporation of S to 10 jig of linearized
plasmid DNA dissolved in PB sucrose into 107 cells. The
electroporation conditions for cells suspended in PB sucrose were420 V and 25 jiF withapulse decayrangeof 12.8to 13.3
ms.When the cellswere suspended in PBS, the Gene Pulser
settingswere500 V and 25 jiF withapulse decayrangeof 0.4
to0.6ms.Forty-eighthours afterelectroporation,thesurviving
cellswerecloned either inagaroseor asmonolayerculturesin DV-5 containing 400 jig of G-418 per ml, unless indicated otherwise.
Theelectroporation conditions for LMtk- cellswere
deter-minedasdescribed above forX1cells. G-418-resistant LMtk
-cloneswere selectedfollowingtheelectroporation ofS jig of
linearized plasmid into 107cellssuspended in 0.8 ml of PBS. The Gene Pulsersettingswere950 V and 25 ,uFwithapulse
decaytime of 0.4 ms.Forty-eighthours afterelectroporation,
thecellswereclonedasmonolayerculturesinDV-5 containing
400,ugof G-418perml.
Electroporations were conducted with linearized plasmid
DNAto increase theefficiencyof transformation (19)and to
control the site ofintegrationwithin theplasmidmolecule(19, 25, 48). pSV2neoandpSV2oriA plasmidswerelinearized with
ApaLI,whichcleaves within thepBR322vectorsequenceson
both sides of the SV40 sequences (Fig. 1B), priorto
electro-poration. Originally, pLTRneowascleaved withEcoRI,which cleaves once within the pBR322 vector sequences and once
within the LTR (Fig. 1A). Although G-418-resistant clones
LMU- suchasXl LTRneoA26(LTRA26)wereobtained with EcoRI-cleaved pLTRneo,cuttingwithin the LTR may decreaseneo
expression. Subsequentelectroporations wereconductedwith
BglI-cleavedpLTRneo (BglI cleaves oncewithin the pBR322
vector sequences [Fig. 1A]), and the clones Xl LTRneoD6
(LTRD6)and Xl LTRneoD8(LTRD8)wereisolated. Theelectroporationconditions used totransfect Cosl cells
at4°Cin0.2-cm Bio-Radcuvettes were450V,400fl,and 3 ,iF
with a pulse decay time of 0.8 ms. The plasmidswere
resus-pended in PB sucrose, and 2 ,ug of supercoiled DNA was introducedbyelectroporationinto 1.5 x 106Cosi cells in 0.1 mlofPB sucrose.The cellswereincubatedat room
tempera-turefor 10 minpriortobeingreseeded in DV-10.
Cosl replication assay. The pSV2oriA candidates were
tested forSV40origin replication activityin Coslcells,which
constitutivelyexpressSV40TAg.Theplasmidstobe assayed
were electroporated into Cosl cells, and 72 h later cellular DNAwasextracted(see below).TheDNA wasdigestedwith EcoRI andDpnI,Southernblotted, andprobedwithpBR322
to identify replication products(53).
Harvesting ofgenomicDNA.Chromosomal DNA was har-vested from monolayer cell cultures as described previously (24). DNAconcentrationsweredeterminedby measuringthe
optical density of the sample at 260 nm or by fluorometric assays (23).
Southern blots. Southern blot
analysis
wasperformed
aspreviously described (24) with minor modifications.
Briefly,
DNA wasdigestedwith restriction enzymesat aconcentration of 5U/,ugofDNAfor 3to18 hatthe
appropriate
temperature. The DNA separated in agarosegels
was blotted ontonylon
(HyBond;Amersham)ornitrocellulose
(Schleicher
&Schuell)
membranespriortohybridizationin 5x SSC
(1
x SSC is 0.15M NaCl plus 0.015 M sodium
citrate)-50
mM sodium phos-phate (pH6.7)-5
mM sodiumpyrophosphate-0.1%
sodiumdodecyl sulfate-200
jig
of thesynthetic heparin
substitutepolyanetholesulfonic
acid(Calbiochem)
per ml. Nick-trans-latedSV40DNA,plasmid DNA,orPCRfragmentswereusedas probes,asindicated.
Statistical analysis. To test for
significant
differences in rearrangement frequencies with each set ofclones,
contin-gency tableanalysis
(SAS
JMP,1989;
SAS InstituteInc.,Cary,
N.C.) was applied. Onlythe clones which were subcloned asmonolayer cultures were used in the statistical
analysis.
The three clones whichweresubcloned in agar(neo20, neol7,
andLTRA26)
were notincluded in thestatisticalanalysis
because of the selective pressureon theTAgcoding region.
RESULTS
Genomicrearrangementsoccur atthesiteofintegrationof
thereporterplasmidpSV2neo.Xl cells havea
high
frequency
of rearrangement at the SV40
integration site;
therefore,
wewere interested in
determining
whether a reporterplasmid
integratedatasecond site in Xl cells would also exhibitahigh
rearrangementfrequency.
TheApaLI
fragment
ofpSV2neo
wasintroduced intoXl cellsby
electroporation,
asoutlined inFig. 3 and described in Materials and Methods. Cloneswere selected with G-418 and
analyzed
forsingle-copy pSV2neo
insertion and
unrearranged parental
Xl SV40DNAby
restric-tion enzymemapping.
Clones that met these criteria weresubcloned, and the individual subclones were
analyzed
for restrictionfragment length
polymorphisms,
indicativeof chro-mosomal rearrangements. Restriction enzymemapping
indi-cated thatpSV2neo
wasintegrated
at adifferent chromosomal location in each clone chosen for furtheranalysis
(results
notshown).
Electroporate plasmid
G-418selection
Pickindividk cokobresandtranstertoPetddshes GrowuLn cornuet
Freeze 1/2 of ceis and harvest DNA fromremainingclls
Analyze cones by restrictonenzymemapping
Reseedfrozencels
Isolehdvlkhasubdones Harvest DNA
Analyze DNA torrearangemeris VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
0
CM'
0
MI2 5 7 11 13 A154)
7.0kb
-4.5kb
-4.1kb
-0
101112131415C
2.3kb
4 64IvII 1
SV40
probe
H H H HHH
1.1 0.75 117 052 4.5kb
1.17kb
1.1kb 1.3kb
0.75kb
pLTRneo probe
H
pSV2neo ori
0.55 3.42kb
FIG. 4. Xl pSV2neo2O subclone analysis. (A) Southern blot with 2.5 p.g of HindIII-digested genomic DNA per lane isolated from G-418-resistant neo20 (lanemarkedneo20)and neo20 subclones grown in the presence of G-418(numberedlanes).Theblotwashybridizedwith
anSV40probe,whichhybridizestoparentalXlSV40DNAandSV40sequences inpSV2neo.The XlSV40 Hindlllrestriction map is drawn below theautoradiogram.Thesizes of theparentalXlSV40 HindIIIbandsarelistedtothe rightof the blot. TheApaLI pSV2neo fragmentin neo20 isindicatedbyanarrow.The1.1-kbHindlllXlSV40bandismissinginneo20,andnovelSV40bandsarepresent.DNAlinear size standardswere runin lane M.(B)Southern blotasinpanelAbutprobedwithpLTRneo,whichhybridizestonec andpBR322sequencespresentin theApaLI pSV2neofragment(Fig.1A).Thegenomic instabilityof theplasmidDNAin this cellline is illustratedbythenovel bandspresentinmostof the subclonescomparedwith thesingle pSV2neoband (marked byarrow)inneo20.H,HindIIIsites.
Inthefirstexperiment,Xl pSV2neocloneswereselected in
monolayercultures in the presence of 400 pLgof G-418 per ml. Twoclones,neo20 andneo17,werethen subcloned in agarose with and without G-418 selection.Figure 4shows aSouthern blot of genomic DNA isolated from neo20 (lane labeled
neo20) and subclones of neo20 (numbered lanes), digested
with Hindlll, and probed with SV40 DNA (Fig. 4A) or
pLTRneo (Fig. 4B). Bands which hybridize with the SV40
probebutnotwith thepLTRneoproberepresenttheXlSV40
parental DNA. The Xl SV40 1.1-kb HindIll fragment was
missing and additional SV40 restriction fragments were presentin neo20, indicatingthat the Xl SV40parental DNA had rearranged. Only one integrated copy of pSV2neo was present (arrows in Fig. 4). Both features of neo20 were
confirmed with other blotsusingdifferent restriction enzymes
(data notshown).
Restrictionfragment lengthpolymorphismswerepresentin the neo gene in most neo20 subclones cultured with G-418 (Table 1;Fig.4Bshows 15 of these subclones), indicatingthat
rearrangements occurred at the site ofpSV2neo integration.
Fifteen of the neo20 subclones had rearrangements in the
parentalXlSV40DNA(Table1;Fig.4A[10subclonesarenot
shown]). In some of the subclones, rearrangements were
observed in the Xl SV40DNAbutnot inthe pSV2neoDNA
TABLE 1. SummaryofrearrangementsinXlclones Rearrangements'
XIclone Noselection G-418selection
xi xi
SV40 neo SV40 neo
pSV2neo2O(neo20) 20/21b 18/21b 15/25b 20/25b
pSV2neol7 (neo17) 4/15b 4/15"b 6/13"b 813b
pSV2neoAl
(neoAl) 14/14 2/14pSV2neoA5 (neoA5) 11/12 1/12
pSV2neoB5 (neoB5) 5/10 4/10
LTRneoA26(LTRA26) 5/10" 0/10b 119b 0/9"
LTRneoD6 (LTRD6) 6/9 0/9
LTRneoD8 (LTRD8) 6/9 0/9
pSV2oriA6B4 (oriA6B4) 7/14 0/13
pSV2oriA6Cl
(oriA6C1) 1/10 0/10pSV2oriA6C3
(oriA6C3)
12/14 0/14pSV2oriAll
(oriAllAI) 3/13 0/13a Number of subclones withrearrangements/totalnumber of subclones.
bSubcloned in agar.
A
7.0kb.-
4.5kb.-
3.7kb-xi
m
4.5kb
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.102.513.74.384.2]SV40 CONTROL REGION CONFERS GENOMIC INSTABILITY 791
(Fig. 4A, lane 9), while in other subclones, rearrangements wereobservedatthesite of pSV2neointegrationbutnotin the Xl SV40 DNA (notshown).
The stability of the integrated reporter plasmid in the absence of selection was determined by subcloning neo20 without G-418. As summarized in Table 1, mostof the neo20 subclonesisolated without drugselection had rearrangements
atboth the pSV2neo and the Xl SV40 integrationsites. Rearrangements were also observedatthesites of pSV2neo and XI SV40 integration in agarose subclones of neol7, as
summarized in Table 1. Restriction enzyme mapping of the parental Xl SV40DNAandpSV2neo in neol7 indicated that theparentalXl SV40 DNAwasunrearrangedbutatleasttwo
copies of pSV2neo were integrated in tandem (results not
shown). Eightof 13G-418-selectedsubclones of neol7 showed rearrangementsat the site ofpSV2neo integration, while 6of the 13 subclones had rearrangements at the Xl SV40 site of integration. RearrangementsatthepSV2neo and the Xl SV40 DNAsiteswerealsoobserved in the absence ofdrugselection
(Table 1). The frequency of rearrangements at the Xl SV40
and thepSV2neo integration sitesvaried from clonetoclone. neo20 had averyhigh rearrangement rate associated with the
single,integratedcopy ofpSV2neo,whileonly4of 15 agarose subclones of neol7 showed rearrangements at the site of integration with two tandem copies ofpSV2neo.
In the second experiment, G-418-resistant Xl pSV2neo
clones were selected in agarose rather than in
monolayer
culture,tominimize thenumber of clones with rearrangements in theparentalXlSV40 DNA(24).ThreeXlpSV2neoclones,
neoAl, neoA5,andneoB5,wereidentifiedthatmetthecriteria ofhavingone integratedcopyofpSV2neo andunchanged
Xl SV40 DNA. The cloneswere subclonedasmonolayer
cultures in the absence of G-418.Rearrangementswereobservedatthe sites of pSV2neo integration and at the Xl SV40 DNAintegration site in subclones ofneoAl, neoA5,and neoB5, as summarized in Table 1. The fraction of neoAl, neoA5, and neoB5 subcloneswith rearrangementsattheparentalXlSV40
DNAsitewassignificantlyhigher (P<0.001) than thefraction with rearrangementsatthethreeindependentpSV2neo inser-tion sites.
Anumber of conclusionscanbe drawn from the
analysis
ofintegrated pSV2neo in Xl cells. First, rearrangements were observedat thesite of
integration
ofa reporterplasmid
with the entire SV40 controlregion,
including
the SV40origin
ofreplication, earlyand late promoter sequences, and enhancers.
Second, these rearrangementswere
independent
of selection with G-418. Third, the rearrangements were observedinde-pendent of the site of chromosomal
integration. Finally,
thefrequency of rearrangement
appeared
to vary from clone to clone.Aplasmidthatdoes not haveSV40DNAis stable inXlcells.
To demonstrate that the
instability
wasnot a commonfeature ofplasmidintegration
inXlcells,
weanalyzed
thestability
ofatransfected plasmid
entirely lacking
SV40sequences. ThreeXI clones transfected with pLTRneo, clones
LTRA26,
LTRD8, and LTRD6, were isolated in agarose cultures andexamined forrearrangementsatthe siteofthe XlSV40DNA and at the pLTRneo
integration
site. Restriction enzymemappingindicated thatonecopy ofpLTRneo was
integrated
at anindependentsite in each of the three clones and that theparental SV40DNAwas
unrearranged
(results
notshown).
Rearrangementswere notobserved atthe
pLTRneo
siteofintegrationinnineG-418-resistantLTRA26subclones isolated in agar, andonly oneof the nine subclones had a
rearrange-ment attheparental XlSV40 site
(Table
1).
Rearrangementswerealsonotobservedatthesite ofpLTRneo
integration
in 10E. coli
00
0 0
8 >
7.0kb- 0.
4.5kb- *
4.1kb- * -
3.7kb-2.3kb- I
I
5.7kb
-Cosl
000~ ~ ~
~ ~ ~
C4)4C')C4)U
0.0 D.
7.0kb-4.5kb 4.1kb- * 3.7kb
2.7kh-
-2.3kb-
-0
I-
I
1t
1-E. col
0
2
Xcze
_x
4)2 4)2 4)2
\0 \0
Cl Co C; CL 0.0. 0. 0.
5.7kb-@I 14
6
FIG. 5. Coslreplicationassay.Southern blotsprobedwith
pBR322
areshown. Lanes in the panelslabeledCoslcontained4 jig ofCosl DNAdigestedwithEcoRI(R)andDpnI (D).TheCosl cellshad been mock transfected or transfected with pSV2neo, pLTRneo, or the
pSV2oriA
plasmids, asindicatedabove thelanes. Lanesinthepanels
labeledE.colicontained 200 ng ofplasmidDNAisolated fromE.coli
anddigestedwith
DpnI
(D)and/orEcoRI,asindicated.LTRA26subclones isolated in the absence of G-418
(Table
1);
however,
5 of the 10 subclones had rearrangements at theSV40
integration
site.LTRD6and LTRD8 subcloneswereisolatedas
monolayer
cultures in the absence of G-418 selection.Rearrangements
were not observed at thepLTRneo
integration
site in nine subclones of LTRD6 and in nine subclones of LTRD8. Rear-rangements wereobserved, however,
at the site ofXlSV40
DNA
integration
insixofthe nine subclonesofbothLTRD6 andLTRD8,
asreported
inTable 1.The resultsofanalysis
of thepLTRneo
clonesindicate thatintegration
ofplasmid
DNAinXlcells is notsufficienttoconfer
genomic
instability
atthe site ofintegration.
Plasmidswithsmalldeletionsin thecore
origin
arestable inXlcells. Todeterminethe
relationship
between the rearrange-mentsobserved atthe siteofpSV2neo
integration
in Xl cells and the presenceof theSV40
origin
ofreplication
inpSV2neo,
we made small deletionsin the
SV40
replication origin.
Two deletionplasmids,
pSV2oriA6
andpSV2oriAll,
which have deletions of 6 and 11bp,
respectively,
at theBglI
site inTAg
binding
site II(Fig.
1C andD),
were tested forreplication
activity
inCosl
cells(Fig. 5).
Plasmid DNA isolated fromE.coli
shows thetypical
dammethylation
patternofsensitivity
toDpnI
digestion
(Fig. 5, panels
labeled E.coli).
This patternwould be retained if the
plasmid
DNA did notreplicate
inCoslcells.
However,
plasmid
DNAreplicated
inCosl
cells is insensitive toDpnI
digestion.
AllCosl
DNAsamples
weredigested
with both EcoRI andDpnI
so thatreplicated
mole-cules,
which are not cutby
DpnI,
would be linearized. The EcoRIplasmid
DNAbandsareshown inFig.
5 in theE.coli
lanes marked R.A strong
replication signal
was observedwithpSV2neo
inCosl cells
(Fig. 5).
Noreplication
wasdetected withpLTRneo,
which doesnotcontain theSV40
origin
ofreplication.
Afaint bandcorresponding
to thepSV2oriA&6
EcoRIbandpredicted
for thereplicated
plasmid
DNA was detected inCosl
cells VOL.68, 1994on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.316.554.74.274.2]M
7.0kb- I-oriA6
4.5kb--w 0
3.7kb- * I.. d- *.g. a*
1.2kb
-1.1kb
-SV40 probe
b b
Xi
F--3.0kb
b
3.8kb
Xo ~~MM
A*, 0 -, *,-*:. 0.* *
+4 -4.5kb * -4.0kb
* -2.7kb
neoprobe
pSV2neol
A
14
kb-7.0kb
-
4.5kb-
3.7kb-B
<MX
z
MM
6! _
. ,V.,
:~~~~
v
FIG. 6. XI pSV2oriA6C3 subclone analysis. (A) SV40-probed Southern blot containing 2.5 pLg ofBglI-digestedgenomic DNA per lane harvested from oriA6C3 (labeled lane) and oriA6C3 subclones (unlabeled lanes). The bandrepresenting BglI pSV2oriA6 integrated DNAis labeled oriA6. The lanewith XI DNA is indicated, and the XI SV40BglIrestriction map is drawn below. DNA linear size standards were run inthe laneslabeled M. b,BglIsites. (B) Blot as inpanel A but probed with a neo-specific PCR fragment that hybridized only to
pSV2oriA6bands.AsinglepSV2oriA6bandis observed in the oriA6C3 lane and in all the subclone lanes. The band observed at the bottom of the first subclone lane isanartifact not observed in other blots.
(Fig. 5). This replicated band, however, wasbarelydetectable compared with pSV2neo replication inCosl cells.Replication products were not detected in genomic DNA isolated from CosI cells transfected withpSV2oriA1 1 (Fig. 5), indicating that the deletion disrupted the replication function of the origin.
The ApaLI fragment of pSV2oriA6 or pSV2oriA 1I was
introduced intoXI cells by electroporation, and
G-418-resis-tantcloneswere isolated in agarose. Four clones meetingthe criteria ofparental XI SV40DNAandone integrated copy of the reporterplasmidweresubcloned inmonolayer cultures in the absence ofselection. Restriction enzyme mappingduring
theanalysis for single-copy integration ofpSV2oriA indicated that the plasmid DNA was integrated at a different chromo-somal location in each clone.
Analysis of three pSV2oriA6 clones and one pSVoriAl1 clone, oriAllAl, revealed that integrated plasmid DNAwith small deletions in the SV40coreoriginremained stable inXI
cells, whereas rearrangements still occurred atthe site of the
parental XI SV40 DNA (Table 1).An example ofrestriction
analysisof14subclones of oriA6C3 is shown inFig.6.Genomic
DNA digested with BglI, which does not cleave within the
pSV2oriA6 ApaLI fragment, gave an identical band for oriA6C3 and all of its subclones when hybridized with a
neo-specific probe (Fig. 6B). The same invariant band was
observed with the SV40 probe (Fig. 6A), although 12 of the subclones showed a new pattern ofSV40 bands representing
rearrangementsthat occurred atthe Xl SV40DNA site(Fig. 6Aand Table 1). Similar resultswere obtained with orizA6B4 and oriA6Cl and with oriAllAl (Table 1). These results suggestthatspecific sequences presentwithin the SV40origin
ofreplication affectgenomicstability.
TAg isnotrequiredforinduction of rearrangementsatthe
site of pSV2neo integration in LMtk- cells. It has been difficulttodetermine the role ofTAg ingenerating
rearrange-mentsinintegrated SV40DNAbecause the phenomenonwas
found in SV40-transformed rodent cells that continuously
expressTAg(4, 24, 27, 28, 42,43, 57,64). Investigationof the
co
FIG. 7. LMtk pSV2neo andLMtk pSV2oriA6 subclone
analy-sis. (A) Southern blot containing 2.5 jigof EcoRI digested genomic
DNAperlane, harvested from LNC3 and LNC3 subclones(unlabeled lanes). The blot was hybridized with nick-translated SV40, which
hybridizestotheSV40DNAflankingbothsides of the Tn5neogene. The lane labeledMcontained linear size standards.(B)Southern blot
containing 2.5 jig ofHindlll-Bgll-digested genomic DNA per lane, harvested from LorizX6A3 and LoriA6A3 subclones. The blot was hybridizedwith nick-translatedpSV2neo. (C)Southernblotcontaining
2.5 jig of EcoRI-digested genomic DNA per lane, harvested from LoriA6B6 and LoriA6B36 subclones. The blot was hybridized with nick-translatedpSV2neo.Thesubclone DNAinonelanewaslost,but another blot demonstrated that theplasmidDNAinthis subclone had
notrearranged.
genomic instability of pSV2neo integrated into a cell line
lackingtheTAggenepresentedanopportunitytodirectlytest
the roleofT Ag.
TheApaLIfragment ofpSV2neowasintroducedby
electro-poration into mouse LMtk cells, a cell line completely lacking SV4O DNA. G-418-selected clones were analyzed for
single-copy integration of pSV2neo by restriction mapping.
Five LMtk- pSV2neo clones with one integrated copy of
pSV2neowerechosenfor furtheranalysis. Restriction enzyme
mapping indicated that pSV2neowasintegrated ata different site in each of the five clones (resultsnot shown).The LMtk
-pSV2neo clones were subcloned in monolayercultures in the
absence of G-418. DNA harvested from the subclones was
digested with restriction enzymes, Southern blotted, and
hy-bridizedwith SV40- orneo-specific probes.
Rearrangementswere observed at the pSV2neo sites in all five LMtk- clones despite the absence ofTAg. A Southern blotof EcoRI-cleavedgenomicDNAfrom LNC3 and from subclones ofLNC3 hybridizedwithSV40DNAisshown inFig.
7A.Two of the 11I subclones have newbands. Theresultswere
identical when the blot was hybridized with a neo-specific
probe instead ofSV40 (data not shown),confirming that the bandswerederived from theApaLI pSV2neo fragment. Bands
were not observed with genomic DNA from LMtk cells
hybridized with either probe (data not shown).
Rearrange-A
>(Mx o
OV) "to
B
>CDcn-i
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.578.346.482.66.316.2] [image:6.578.36.275.67.226.2]SV40 CONTROL REGION CONFERS GENOMIC INSTABILITY 793 TABLE 2. SummaryofrearrangementsinLMtk- clones
neo
LMtk-clone rearrangementsa
(noselection)
pSV2neoA3 (LNA3)... 3/11
pSV2neoA6(LNA6)... 6/17
pSV2neoB6(LNB6)... ... 6/17
pSV2neoC1(LNC1)... 4/13
pSV2neoC3 (LNC3)... 2/11
pSV2oriA6A3 (LoriA6A3)... 0/16
pSV2oriA6B6 (LoriA6B6)... 1/16
pSV2oriA6C5 (LoriA&6C5)... 3/15
pSV2oriAllA4 (LoriAlIA4)... 0/13
a Number of subclones withplasmid rearrangements/total numberof
sub-clones.
ments were observed at the sites of pSV2neo integration in LNA3, LNA6, LNB6, and LNC1 (Table 2). Each set of subclones was analyzed by using at least two restriction
en-zymes toverify that new bands were due to rearrangement and
not to incomplete restriction enzyme digestion (data not
shown).
The genomic instability observed at the site of the integra-tion of a reporter plasmid containing the SV40 origin of replication did not require T Ag in LMtk- cells. These results indicate that the rearrangements must beinduced by theDNA
alone or by some factor present in LMtk- cells.
Deletions in the SV40 core origin increase the genomic
stability of integrated plasmid DNA in LMtk- cells. To determine whether the rearrangements were dependent on a functional origin of replication, pSV2oriA6 and pSV2oriA11
were introduced into LMtk- cells by electroporation, and G-418-resistantcolonies were selected and analyzed for single-site insertions. Four clones, LoriA6A3,
LoriA6B6, LoriA6C5,
and
LoriA1lA4,
with asingle pSV2oriA plasmid integrated atadifferent site were chosen for subcloning.
Rearrangements were not observed in any of the 16 sub-clones ofLoriA6A3 (Fig. 7B) orin anyof the 13 subclones of
LoriAlllA4
(Table 2). Rearrangements were observed, how-ever,in 3 of the 15 subclonesofLoriA6C5 (Table 2) andin 1of the 16 subclones of LoriA6B6 (Fig. 7C). Each set of subclones was analyzed by using at least two restriction
en-zymestoverify that thenewbandsweredue to rearrangement andnot to incomplete restriction enzyme digestion (data not
shown). Acontingency table analysis indicated that the fre-quencyofpSV2oriAsubcloneswith rearrangements in LMtk-cellswas significantly lower than that ofpSV2neo subclones with rearrangements (P < 0.01). Small deletions within the
core origin ofreplication that diminished replication activity
increased the genomic stability of the integrated plasmid
DNA;however, the deletions didnottotally inhibit
rearrange-ments. These results, combined with the results presented
above for theXl cells,indicatethat thewild-typeSV40origin regioninducedgenomic instabilityat thesite of chromosomal
integration. Small deletions in thecore origin disrupted this function.
DISCUSSION
The instability of integrated SV40 DNA in nonpermissive cells has been observed in many different cell lines(4, 24, 27, 28, 42, 43, 57, 64). In each of thesestudies, SV40 TAgwas
expressedinthe cells and thepredominantmodels invoked the replication functions ofTAg and the SV40origin of
replica-tion. However, Breitman et al. (7) previously reported rear-rangementsatthesiteofintegrated pSV2gptin the absence of
TAg in Chinese hamster ovary
(CHO) cells,
and Heartleinet al. (26) reported that complexplasmids containing
the SV40regionwereunstable in the absence ofTAg.Theresults ofour
study prove that the SV40 control
region
isrequired
for the observedgenomic instabilitybut that TAg isnotrequired.Genomic instability is correlated with the presence of the SV40origin ofreplicationinXl cells.To
analyze
the role of the SV40 origin of replication in the genomic instability ofintegrated SV40 DNA in
nonpermissive
celllines, pSV2neo
wasintegratedinto the SV40-transformedcell line Xl.Inthe five independent pSV2neo clones analyzed, rearrangements
wereobservedat each site ofplasmid integration.
Rearrange-ments were not observed at three independent pLTRneo
integration
sites in Xlcells,
indicating
thatintegration
offoreign DNA into the cellular genome is not sufficient for
generating rearrangements. Small deletions within the SV40
core origin in pSV2neo decreased the rearrangement
fre-quency, indicating that sequences present in the PvuII-to-Hindlll SV40
origin fragment
ofpSV2neo
arethe maximum sequences required to confergenomic instability
to theinte-gratedDNA.
Theinstabilityof the Xl SV40DNAwasmaintainedinthe Xl pSV2neo clones, Xl
pSV2oriA clones,
andXlpLTRneo
clones,indicating
thatindependent
events wereoccurring
at thetwoseparateintegrationsites.The chromosomal site of theXl SV40 DNA, in the absence of
integrated
viral DNA, is stable(2); therefore,
the SV40 DNA is notintegrated
at apreexistingchromosomalrecombination hot spot. In
addition,
sevencellular geneswerestablein Xl
cells, indicating
that theinstabilityassociated with the site ofSV40DNA
integration
isnotdueto ahighrearrangementratethroughoutthe genome
(24).
TAgisnotessentialforthegenomicinstabilityofintegrated
SV40DNA.The presenceof theSV40control
region, including
the
origin, enhancer,
and promoter sequences,wassufficientto causegenomic instability
atthe site ofintegration
ofpSV2neo
in five
independent
LMtk- clones in the absence of T Ag.There is no statistical difference
(P
>0.05)
between the fractionof LMtk-pSV2neo
subclones with rearrangementsat theneo locus in the absence ofTAgand the total fraction of XlpSV2neosubclones with rearrangementsattheneolocus in the presenceofTAg,indicating
that the presence ofTAg
does notincreasethegenomic instability
ofpSV2neo.
Small deletions in theSV40
origin
ofreplication
decreased the rearrangementfrequency
in LMtk- cells.Rearrangements
wereobserved with
pSV2oriA
plasmids
at asignificantly
lowerfrequency
(P
<0.01)
than observed withpSV2neo.
Rearrange-ments at the site of
integration
offoreign
DNAcontaining
mutations in the SV40 core
origin
thatdisrupt
the viralreplication activity
have beenpreviously reported
(13, 45-47).
The
integration
offoreign
DNAinto LMtk-cells, however,
is notsufficientto induce rearrangements(38, 69).
Theincreased fraction of LMtk-
pSV2oriA
subclones with rearrangements at the neo locuscompared
with the XlpSV2oriA
subclones may beareflection ofa morepermissive
state for recombination in LMtk- cells. Geneamplification
rates vary among different cells lines
(54),
and a similar variation in permissiveness for DNA rearrangements may exist.Although
thepossibility
thatintegration
occurred in recombination hot spots cannot be excluded for any of the reporterplasmids
thatexhibited rearrangements inthisstudy,
it isunlikely
thatpSV2neo
integrates
into hot spots morefrequently
than eitherpSV2oriA
orpLTRneo.
Flanking DNA may influence the
frequency
of rearrange-VOL.68, 1994on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.61.301.87.209.2]ments. The fraction of subclones with rearrangements at the site of the parental Xl SV40 DNA varied between clones from 100% of the subclones with rearrangements (neoAl) to only 10% of the subclones with rearrangements (oriA6C1), al-though the number of subclones examined(14 and 10, respec-tively, in the examples cited) was too small to establish the statistical significance of the variation. Three factors might affect the observed frequency of rearrangement. First, the number of subclones with rearrangements was determined conservatively because faint minor bands were excluded. Sec-ond, duplications in the mouse DNA and part of the SV40 earlyregion,moused(TG)repeats thatare potential Z-DNA
tracts, and other repetitive mouse DNA sequences flank the SV40integration site in Xl cells (2, 24) and may contributeto
the different recombination rates. Small changes in these regions might nothavebeendetectedby the restriction enzyme analysis used in this study.Third, the cellular environments of the clones may not be identical. Gross chromosomal aberra-tions,including chromatid breaks, dicentric chromosomes, and translocations, which are frequently observed in transformed cell lines (34, 54, 56, 65), may exist and might affect the mechanisms involved in generatingthe rearrangements.
The fraction ofXl pSV2neo subclones with rearrangements
attheXISV40sitewasstatisticallyhigher(P<0.001)then the fraction of subclones with rearrangements at the site of pSV2neo integration. The SV40 origin of replication and auxiliary sequencesarepresent in both theparental Xl SV40
DNA and pSV2neo, and we assume that both origins are exposed to the sameTAgpopulation inXl cells. However, the different DNA sequences flanking the integration sites may influence the frequency of rearrangements (9, 10, 35, 39, 40, 47, 49, 52, 55, 72, 74, 76, 77, 78) observedatthe Xl SV40 site and thepSV2neo sites. ThepSV2neointegrationsites havenot
been characterized.
Models to explain the observed instability associatedwith
the SV40 origin. Many of the models proposed to explain
rearrangements associated with integrated SV40 DNA have beenbasedonthe viral DNAreplicationfunctions ofTAg and the SV40 origin ofreplication(15, 24, 28, 33). The permissive-nessof cellstosupport viralDNAreplicationis associated with theability of the polymeraseot/primasetointeractdirectlywith
T Ag (16, 17, 44). In cells that are nonpermissive for viral replication, T Ag-based replication models require, at some
level, that this inhibition of viralreplicationbe circumvented. Previous reports (7, 26-28, 66)andour results reported here indicate thata TAg-basedreplication model is not requiredto
explainthegenomic instabilityofintegratedSV40DNAin cell linesnonpermissive for viral replication. However, integration of theSV40origin ofreplicationintochromatin maycreate or
unmask acryptic function of the SV40origin, independent of
T Ag, that could activate recombination or repair enzymes, resulting in the observed rearrangements. The activation of a cryptic eukaryotic replication origin by a chromosomal
rear-rangement has been reported(37).
Different models basedonDNA structurerather than origin activity may also explain the instability associated with the SV40 origin region. Secondary structures can arrest DNA replication, leading to genomic instability (1, 14; reviewed in reference61). The rearrangements, therefore, may be induced byDNAsecondary structurespresent in theSV40 core origin and auxiliary regions, including nuclease-hypersensitive sites presentin the21-bp repeats, the 72-bp enhancers (20, 31, 32), andthemajor late promoter (81, 82). Furthermore, the SV40
DNAcontrolregion is richinpalindromic sequences with the potential of forming cruciforms (5, 51, 67), which have been
implicated as recombination hot spots (30, 36, 75, 79, 80).
CruciformstructuresinSV40DNAhavenotbeen detectedin
vivo(80);however,integrationof theSV40 controlregioninto chromatin may induce or allow cruciform formation. Se-quenceswith thepotential offormingZ-DNA, whicharealso implicated in recombination (8,70, 71, 73,74),are presentin the SV40earlypromoter(50).
MaintainingthepalindromeatTAgbindingsite IImaybe
important for permitting DNA rearrangements.
Rearrange-ments were observed at the site ofintegration of an
origin-defective plasmid with a deletion that disrupts the perfect palindrome in T Ag binding site 11(45) and at the site of
integration of a replication-defective construct with a 4-bp
insertion (and a 1-bptransition) in T Agbindingsite II that
only partially disrupted the palindrome (12, 13). We also observed rearrangements at the site of integration of
pSV2oriA6 and pSV2oriA11 in LMtk- cells. Our
results,
however, indicate that the presence of small deletions inTAg binding site II in
pSV2oriA6
and pSV2oriA11 decreased the frequency of rearrangement. The permissiveness of the cell line for DNA rearrangements, in conjunction with residualsecondarystructure, may
explain
the rearrangementsobservedat the
pSV2oriA6
sites in LMtk- cells.Alternatively,
theexpressionofTAg in Xl cells may interfere with recombina-tion in the Xl pSV2oriA clones by blocking hot spots for recombination (73). The roles of the early
palindrome
and other secondary structures associated with the SV40 controlregionwere notinvestigatedin thisstudy.
Transcriptionhasbeen showntoincrease theaccessibilityof
DNA to recombination machineryornuclease attack
(41, 48,
68).InpSV2neo,pSV2oriA6,
andpSV2oriAl11,
neoexpression
is under the control of the SV40 early promoter. It is not known what effect deletions in the core origin have on the
secondarychromatinstructureof
pSV2oriA6
andpSV2oriA
11during transcription. At this time, we cannot
distinguish
be-tween
possible
contributionsof thetranscription
components(SV40
early promoter andenhancers)
and thereplication
components
(SV40
coreorigin andauxiliary
regions)
ininduc-ingthe rearrangements.
ACKNOWLEDGMENTS
We thank Margaret Woolf, Edward Rock, Mindy Larsen, Irene Chen, and Kerstin Beijer for excellent technical assistance, David Berrigan for help with the statistical analysis, Kirk Thomas for pRMH140,and Lauri Fraser for the LMtk- cells. Our research has benefited from discussions with Ted Gurney, Leena Gandhi, Rich Davis, JohnRoth,and Dana Carroll.
This workwas supportedbyAmerican CancerSocietygrant MV-485. The synthesis of oligonucleotide primers was subsidized by National CancerInstitutegrant 5P30 CA42014.
REFERENCES
1. Baran,N.,A. Lapidot,and H. Manor. 1991. Formation ofDNA triplexes accounts for arrests of DNA synthesis at d(TC)n and d(GA)ntracts.Proc.Natl. Acad.Sci.USA88:507-511.
2. Beagley, C. T. 1989. Spontaneous rearrangement ofintegrated simian virus 40 DNA inmousecells:analysisbygenomic cloning inbacteriophages. M.S.thesis.UniversityofUtah,Salt LakeCity. 3. Beagley, C.T.,T. Gurney, Jr.,and E. G. Gurney (Universityof
Utah).1993.Unpublishedresults.
4. Bender, M. A., and W. W. Brockman. 1981. Rearrangement of integrated viral DNA sequences in mouse cells transformed by simian virus 40. J. Virol.38:872-879.
5. Bergsma,D.J.,D.M.Olive,S. H.Hartzell,andK.N. Subrama-nian.1982.Territorial limits and functional anatomy of the simian virus 40replication origin.Proc.Natl. Acad. Sci. USA 79:381-385. 6. Botchan, M., W. C. Topp, and J. Sambrook 1978. Studies on simian virus 40 excision from cellular chromosomes. ColdSpring HarborSymp.Quant.Biol. 43:709-719.
on November 9, 2019 by guest
http://jvi.asm.org/
SV40 CONTROL REGION CONFERS GENOMIC INSTABILITY 795 7. Breitman, M. L., L.-C. Tsui, M. Buchwald, and L. Siminovitch.
1982. Introduction and recovery of a selectable bacterial gene from the genomeofmammalian cells. Mol. Cell. Biol.2:966-976. 8. Bullock, P., J. Miller, and M. Botchan. 1986. Effects of
poly[d(pGpT)* d(pApC)] and poly[d(pCpG)-d(pCpG)] repeats
on homologous recombination in somatic cells. Mol. Cell. Biol. 6:3948-3953.
9. Butner, K. A., and C. W. Lo. 1986. High frequency DNA rear-rangements associated with mousecentromeric satellite DNA. J. Mol. Biol.187:547-556.
10. Caddle,M.S.,R H.Lussier, and N. H. Heintz. 1990. Intramolec-ular DNAtriplexes,bentDNAand DNA unwinding elements in the initiation region of an amplified dihydrofolate reductase replicon. J. Mol. Biol.211:19-33.
11. Carroll, S. M., P. Gaudray, M. L. De Rose, J. F. Emery, J. L. Meinkoth,E.Nakkim,M.Subler, D. D. Von Hof, and G. M. Wahl. 1987. Characterization of an episome produced in hamster cells thatamplifyatransfected CAD gene at high frequency:functional evidence for a mammalian replication origin. Mol. Cell. Biol. 7:1740-1750.
12. Chen, S.,D. S.Grass,G.Blanck, N.Hoganson, J.L.Manley,and R. E. Pollack 1983. A functional simian virus 40 origin of replicationisrequired forthegeneration of a super T antigenwith
amolecularweight of 100,000in transformed mouse cells. J. Virol. 48:492-502.
13. Chen, S.,andR.Pollack 1986. DNArearrangement and the role ofviral origin in SV40-transformed mouse cells. Cancer Cells 4:381-386.
14. D'Ambrosio, E., and A. V. Furano. 1987. DNA synthesis arrest sitesattherightterminus of ratlong interspersed repeated (LINE or L1Rn) DNA family members. Nucleic Acids Res. 15:3155-3175.
15. Daya-Grosjean, L.,andR. Monier. 1978.Presence of a freeviral DNA in simianvirus40-transformed nonproducer cells. J. Virol. 27:307-312.
16. Dornreiter, I., L. F. Erdile,I. U. Gilbert, D. von Wiflkler, T. J. Kelly, and E. Fanning. 1992. Interaction of DNA polymerase a-primasewith cellularreplicationprotein A andSV40Tantigen. EMBOJ. 11:769-776.
17. Dornreiter, I.,A.Hoss, A. K. Arthur, and E. Fanning. 1990.SV40 T antigen binds directly to the large subunit of purified DNA polymerase alpha.EMBO J.9.3329-3336.
18. Dower, W. J., J. F. Miller, and C. W. Ragsdale. 1988. High efficiencytransformationof E.coliby high voltageelectroporation. Nucleic Acids Res. 16:6127-6145.
19. Folger, K. R.,E. A.Wong, G. Wahl, and M. R, Capecchi. 1982. Patterns ofintegration ofDNAmicroinjected into cultured mam-malian cells: evidence for homologous recombination between injected plasmidDNAmolecules. Mol. Cell. Biol. 2:1372-1387. 20. Gerard, R.D., B. A. Montelone, C. F. Walter, J. W. Innis, and
W. A.Scott. 1985.Roleofspecificsimian virus 40 sequences in the nuclease-sensitive structure in viral chromatin. Mol. Cell. Biol. 5:52-58.
21. Gluzman, Y. 1981. SV40-transformed simian cells support the replicationofearlySV40mutants.Cell23:175-182.
22. Gluzman,Y., J. Frisque,andJ.Sambrook. 1980.Origin-defective
mutantsofSV40.ColdSpringHarbor Symp. Quart. Biol. 44:293-300.
23. Gurney, T., Jr.,and E.G. Gurney. 1984. DABA fluorescence assay for submicrogram amounts of DNA, p. 5-11. In J. M. Walker
(ed.), Methods in molecularbiology, vol. 2. Nucleic Acids. Hu-manaPress, Clifton,N.J.
24. Gurney, T., Jr.,andE. G.Gurney. 1989.Spontaneous rearrange-ment of integrated simian virus 40 DNA in nine transformed rodent cell lines. J. Virol. 63:165-174.
25. Hanahan, D., D. Lane,L. Lipsich, M. Wigler, and M. Botchan. 1980. Characteristics of an SV40-plasmid recombinant and its
movement into and out of the genome of a murine cell. Cell
21:127-139.
26. Heartlein,M.W., J.H.M.Knoll, and S. A.Latt. 1988. Chromo-some instabilityassociated with human alphoid DNAtransfected into theChinesehamster genome. Mol. Cell. Biol.8:3611-3618. 27. Hiscott,J., D.Murphy, and V. Defendi. 1980. Amplification and
rearrangement of integrated SV40 DNA sequences accompany theselection ofanchorage-independent transformedmouse cells. Cell22:535-543.
28. Hiscott, J. B., D. Murphy, and V. Defendi. 1981. Instability of integrated viral DNA in mouse cells transformed by simian virus 40. Proc. Natl. Acad. Sci. USA 78:1736-1740.
29. Hudziak, R. M., F. A. Laski, U. L. RajBhandary,P. A.Sharp, and M. R Capecchi. 1982. Establishment of mammalian cell lines containing multiplenonsensemutations and functionalsuppressor tRNA genes. Cell 31:137-146.
30. Hyrien,O.,M. Debatisse,G. Buttin,and B. R de Saint Vincent. 1987. A hotspot for novel amplification joints in a mosaic of Alu-like repeats and palidromic A + T-rich DNA. EMBO J. 6:2401-2408.
31. Innis, J. W.,andW.A.Scott. 1984. DNAreplicationand chroma-tin structure of simian virus 40 insertion mutants. Mol. Cell. Biol. 4:1499-1507.
32. Jongstra, J., T. L. Reudelhuber, P. Oudet, C. Benoist, C.-B.Chae, J.-M. Jeltsch, D. J.Mathis, and P. Chambon. 1984. Induction of altered chromatin structures by simian virus 40 enhancer and promoter elements. Nature (London)307:708-714.
33. Kaufman, R. J., andP. A.Sharp.1982.Amplification and expres-sion of sequences cotransfected with a modular dihydrofolate reductase complementary gene. J. Mol. Biol. 159:601-621. 34. Kit, S., D. R. Dubbs, L. J. Piekarski, and T. C. Hsu. 1963.Deletion
of thymidine kinase activity from L cells resistant to bromode-oxyuridine. Exp. Cell Res. 31:297-312.
35. Klobutcher, L. A., and D. M. Prescott. 1986. The special case of the hypotrichs, p. 111-154. In J. G. Gall (ed.), The molecular biology of ciliated protozoa. Academic Press, New York. 36. Leach, D. R. F., and F. W. Stahl. 1983. Viability ofA phages
carrying a perfect palindrome in the absence of recombination nucleases. Nature (London) 305:448-451.
37. Leu, T.-H., and J. L. Hamlin. 1992. Activation of a mammalian origin of replication by chromosomal rearrangement. Mol. Cell. Biol. 12:2804-2812.
38. Lo,C. W. 1983. Transformationbyiontophoretic microinjectionof DNA: multiple integrations without tandem insertions. Mol. Cell. Biol. 3:1803-1814.
39. McArthur, J. G., L. K. Beitel, J. W. Chamberlain, and C. P. Stanners. 1991. Elements which stimulate gene amplification in mammalian cells: role of recombinogenic sequences/structures and transcriptional activation. Nucleic Acids Res. 19:2477-2484. 40. McArthur,J. G., and C. P. Stanners. 1991. A genetic element that
increases the frequency of gene amplification. J. Biol. Chem. 266:6000-605.
41. Mink, M., A. N. Basak, and H. Kuntzel. 1990. Restoration of the yeast LEU2 gene by transcriptionally controlled recombination between tandem repeats. Mol. Gen. Genet. 233:107-113. 42. Mougneau, E., F. Birg, M. Rassoulzadegan, and F. Cuzin. 1980.
Integration sites and sequence arrangement ofSV40DNA in a homogeneous series of transformed rat fibroblast lines. Cell 22:917-927.
43. Mounts, P., and T. J.Kelly,Jr. 1984. Rearrangements of host and viral DNA in mouse cells transformed by simian virus 40. J. Mol. Biol. 177:431-460.
44. Murakami, Y., C. R. Wobbe, L. Weissbach, F. B. Dean, and J. Hurwitz. 1986. Role of DNA polymerase a and DNA primase in simian virus 40 DNA replication in vitro. Proc. Natl. Acad. Sci. USA 83:2869-2873.
45. Murnane, J. P. 1986. Inducible gene expression by DNA
rear-rangements in human cells. Mol. Cell. Biol. 6:549-558.
46. Murnane, J. P. 1990.Influence of cellular sequences oninstability of plasmid integration sites in human cells. Somatic Cell Mol. Genet. 16:195-209.
47. Murnane, J. P., and M. J. Yezzi. 1988. Association of high rateof recombination with amplification of dominant selectable genein human cells. Somatic Cell Mol. Genet. 14:273-286.
48. Murnane, J. P., M. J. Yezzi, and B. R. Young. 1990. Recombina-tion events during integraRecombina-tion of transfected DNA into normal human cells. Nucleic Acids Res. 18:2733-2738.
49. Murnane, J. P., and B. R. Young. 1989. Nucleotide sequence VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
analysis of novel junctions near an unstable integrated plasmid in human cells. Gene84:201-205.
50. Nordheim, A., and A. Rich. 1983. Negatively supercoiled simian virus 40 DNA contains Z-DNA segments within transcriptional enhancer sequences. Nature (London) 303:674-678.
51. Panayotatos, N., and R. D. Wells. 1981. Cruciform structures in supercoiledDNA. Nature (London) 289:466-470.
52. Passananti, C., B. Davies, M. Ford, and M. Fried. 1987. Structure of an inverted duplication formed as a first step in a gene amplification event: implications for a model of gene amplifica-tion. EMBO J. 6:1697-1703.
53. Peden, K. W. C., J. M. Pipas, S. Pearson-White, and D. Nathans. 1980. Isolation of mutants of an animal virus in bacteria. Science 209:1392-1396.
54. Perry,M.E., M. Commane, and G. R.Stark.1992.Simian virus 40 large tumor antigen alone or two cooperating oncogenes convert REF52 cells to a state permissive for gene amplification. Proc. Natl.Acad.Sci. USA 89:8112-8116.
55. Ponticelli, A. S., and G. R. Smith. 1992. Chromosomal context dependenceofaeukaryotic recombinationalhot spot. Proc. Natl. Acad.Sci. USA 89:227-231.
56. Ray, F. A., D. S. Peabody, J. L. Cooper, L. S. Cram, and P. M. Kraemer. 1990. SV40 Tantigen alone driveskaryotypeinstability thatprecedes neoplastic transformation of humandiploid fibro-blasts. J.Cell. Biochem. 42:13-31.
57. Sager, R., A. Anisowicz, andN. Howell.1981.Genomic rearrange-mentsin a mousecellline containingintegrated SV40 DNA. Cell 23:41-50.
58. Sambrook, J., M. Botchan, P. Gallimore, B.Ozanne, U. Petters-son,J.Williams, and P. A.Sharp. 1975. Viral DNA sequences in cells transformed by simian virus 40, adenovirus type 2 and adenovirus type 5. ColdSpringHarborSymp. Quant.Biol. 39:615-632.
59. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning:alaboratorymanual, 2nded. ColdSpringHarbor Labo-ratoryPress, ColdSpring Harbor, N.Y.
60. Sanes, J. R., J.L.R.Rubenstein, andJ. F. Nicolas. 1986. Useof
arecombinantretrovirustostudypost-implantationcelllineagein mouseembryos. EMBO J. 5:3133-3142.
61. Schimke, R. T., S. W. Sherwood,A. B.Hill, and R. N. Johnston. 1986. Overreplication andrecombination of DNA in higher
eu-karyotes: potential consequences and biological implications. Proc.Natl. Acad. Sci. USA 83:2157-2161.
62. Schuurman, R., and W.Keulen.1991. Modifiedprotocolfor DNA sequenceanalysisusing Sequenase2.0.BioTechniques 10:185. 63. Southern,P.J.,and P.Berg. 1982.Transformation of mammalian
cellstoantibiotic resistance withabacterial gene under control of theSV40early regionpromoter. J.Mol.Appl.Genet. 1:327-341. 64. Steinberg, B., R. Pollack, W. Topp, and M. Botchan. 1978. Isolation and characterization of T antigen-negative revertants from a line of transformed ratcellscontaining one copy of the SV40genome. Cell 13:19-32.
65. Stewart, N., and S. Bacchetti. 1991.Expression ofSV40 largeT
antigen,butnotsmall tantigen, isrequired for theinduction of chromosomal aberrations in transformed human cells. Virology 180:49-57.
66. St-Onge, L.,L. Bouchard,andM. Bastin. 1993.High-frequency
recombination mediated by polyomavirus large T antigen defec-tive in replication. J. Virol. 67:1788-1795.
67. Tenen, D.G., L. L. Haines, and D. M. Livingston. 1982. Binding of ananalog of the simian virus 40 T antigen to wild-type and mutant viralreplication origins. J. Mol. Biol. 157:473-492.
68. Thomas, B. J., and R. Rothstein. 1989. Elevated recombination rates intranscriptionally active DNA. Cell 56:619-630.
69. Thomas, K. R., K. R. Folger, and M. R. Capecchi. 1986. High frequency targeting of genes to specific sites in the mammalian genome.Cell 44:419-428.
70. Treco, D.,and N. Arnheim. 1986. The evolutionarily conserved repetitive sequence d(TG AC), promotes reciprocal exchange and generates unusual recombinant tetrads during yeastmeiosis. Mol. Cell.Biol. 6:3934-3947.
71. Treco, D., B. Thomas, and N. Arnheim. 1985. Recombination hot spots in the human ,-globingene cluster:meioticrecombination of human DNAfragmentsinSaccharomyces cerevisiae. Mol.Cell. Biol. 5:2029-2038.
72. Wahl, G. M., B. R. de SaintVincent, and M. L. DeRose. 1984. Effect of chromosomal position on amplification of transfected genesin animal cells. Nature(London) 307:516-520.
73. Wahls, W. P., and P. D. Moore. 1990.Homologousrecombination enhancement conferredby the Z-DNAmotifd(TG)30isabrogated bysimian virus 40 TantigenbindingtoadjacentDNA sequences. Mol. Cell. Biol. 10:794-800.
74. Wahls,W.P., L. J. Wallace, and P. D. Moore.1990.Hypervariable minisatellite DNA is ahotspot for homologousrecombinationin human cells. Cell 60:95-103.
75. Ward, P., and K. I.Berns. 1991. Invitro rescueofanintegrated hybrid adeno-associated virus/simian virus 40 genome. J. Mol. Biol. 218:791-804.
76. Wegner, M., S. Schwender, E. Dinkl, and F. Grummt. 1990. Interactionofaprotein withapalindromicsequencefrom murine rDNAincreases the occurrenceofamplification-dependent trans-formation in mouse cells. J.Biol. Chem. 265:13925-13932. 77. Wegner, M., S.Schwender, E. Dinkl, and F. Grummt. 1990. An
amplification-promoting sequence from mouse genomic DNA: interaction with a trans-acting factor that alsoaffectsgene expres-sion. DNA Cell Biol. 9:311-321.
78. Wegner,M., G. Zastrow, A. Klavinius, S. Schwender, F.Muller,H. Luksza, J. Hoppe, J. Wienberg, and F. Grummt. 1989. Cis-acting sequences from mouse rDNA promote plasmid DNA amplifica-tionandpersistenceinmousecells:implicationof HMG-1 in their function. Nucleic Acids Res. 17:9909-9932.
79. West, S. C., and A. Korner. 1985. Cleavage of crucifrom DNA structuresbyanactivityfromSaccharomycescerevisiae. Proc.Natl. Acad. Sci. USA 82:6445-6449.
80. Zannis-Hadjopoulos, M., L. Frappier, M.Khoury,and G. B. Price. 1988. Effect of anti-cruciform DNA monoclonal antibodies on DNAreplication. EMBO J. 6:1837-1844.
81. Zhang, L., and J.D.Gralla. 1989. In situnucleoproteinstructure attheSV40majorlate promoter: melted andwrappedDNAflank thestartsite. Genes Dev. 3:1814-1822.
82. Zhang,L., and J.D.Gralla.1990. Insitunucleoprotein structure
involving origin-proximal SV40 DNA control elements. Nucleic Acids Res. 18:1797-1803.