0022-538X/86/010138-07$02.00/0
Copyright ©D 1986, American Society for Microbiology
Functional
Interactions
of the
Simian Virus 40 Core Origin of
Replication with Flanking Regulatory Sequences
ANGELO L.DELUCIA, SUMITRA DEB, KATHRYN PARTIN, AND PETERTEGTMEYER*Department of Microbiology, State University ofNew York, Stony Brook, New York 11794 Received 28May 1985/Accepted 24September 1985
Weconstructeda matched setof plasmidstoinvestigate theinteractionsofessentialcoresequences ofthe simian virus40 replication origin with flanking regulatory sequences. Deletions of either T-antigen-binding
regionIorthe21-base-pairrepeatedpromoterelementsreduced replicationto50to70%of wild-type levels. The simultaneous deletionofbothregions decreased replicationtoless than5% of wild-type levels. Thus, the double deletion greatly amplified the defects of the single deletions. We conclude that region I and the 21-base-pairrepeatshave related rather than independent functions in DNAsynthesis.Insertionofasynthetic regionI or theadenovirus 2 major late promoter atthe late side ofisolated core sequences in place of the 21-base-pair repeats failed to restore replication. In contrast, insertion of a single 72-base-pair enhancer elementstimulated replication of thecoreorigin morethan fivefold. Thus, threedistinctregulatory elements appeartofacilitatecoreDNAreplication by related mechanisms. Flankingsequenceshave onlyasmall direct
effect on T-antigen binding to naked core DNA. Possible mechanisms of action include the regulation of
transcriptionorofchromatin structure.
Simian virus 40(SV40) isan attractive modelfor eucary-otic DNA replication (5, 28). A single viral protein, T antigen, binds to a unique origin of replication to initiate each round of viral DNA synthesis. Intracellular viral nucleoprotein closely resembles cellular chromatin, and evehtsintheinitiation andelongationof viralDNAsynthesis mimiceventsin thereplicationof cellular chromosomes (15). The recent establishment ofcell-free systems for complete cycles of viral DNA replication provides a means for the identification of cellular replication proteins (20, 21, 25). Hence, acompletemutational analysisof the SV40originof replication promisesabetterunderstandingofprotein-DNA interactions and events that occur within the origin.
Previousstudies haveestablishedthe approximate bound-aries ofthe smallest origin that would support detectable levelsof replication. We refer to these sequences as
the
core origin.Deletion analysesateither endoftheorigin locatethe limitsof the core between nucleotides 5208 and 5222 at the earlyend and nucleotides26 and 30at the lateend (1, 7, 8, 10, 16, 17, 26). DNA regions flankingboth endsofthe core origin increase the efficiency of DNA replication. These ancillary regions consist of the high-affinity T-antigen-binding region I at the early end of the core and the 21-base-pair (bp)repeatedpromoterelementsatthe late end of the core. Estimates of DNAreplication
after single deletionsof either ancillary region varywidely. Theyrange from20to70%
of wild-typelevels after deletion ofregionI (8, 17) and from10 to100%ofwild-type levels after deletion of promoter elements (16, 19). These variations probably reflect differences both in the contextofsurrounding DNA and in assay conditions.We constructed a matched set ofplasmids to investigate further the interaction of essential core sequences with flanking regulatory sequences that facilitate core function. We compared the effects of single and double deletions of ancillary regions on replication efficiency under standard conditionsat avarietyof times after transfection. Although singledeletions ofregionIorthe21-bp repeatsdelayedand
*Correspondingauthor.
reduced DNA synthesis, maximal levels ofreplicationwere greaterthan50%of wild-type levels. In contrast,the simul-taneous deletionof
both
regions drasticallyreduced but did noteliminatecorefunction.Thus,well-defined sequencesat eitherend ofthe core origin have related rather than inde-pendent functions in facilitating DNA replication. Further-more, replacement of the 21-bp repeats by a single 72-bp repeatof theSV40 enhancer restored replication efficiency. Finally, ourresults establish the autonomy ofa small core originin thecompleteabsence ofadjacent SV40sequences.MATERIALS AND METHODS
Plasmid constructions. The common pOR vehicle was derived from pML2 (22) by deletion ofthe small fragment fromEcoRI to AccI after conversion of theAccI site to a HindlIl site. This segment served asan acceptorfor SV40 origin restriction fragments. The pOR4 andpOR3
plasmids
contain SV40 sequences 5171 to 160 between the SV40 HindIll and BstNIsites andanEcoRI linkerattheBstNIend. PlasmidspOR2andpOR1includeSV40 sequences 5171to39 between the SV40 HindlIl and NcoI sites and a 23-bp polylinker(5'-GTCGACCGGATCCCCGGGAATTC-3')
withSall, BamHI, SmaI, XmaI, and EcoRI sites. Plasmids pOR4andpOR2werederivedfromSV40wild-typestrain 776 (28), andplasmids pOR3andpOR1 were
derived
fromSV40 mutantcs-1097(8), which hasadeletion of sequences 5178to 5208.Four variants of pOR1 were also
constructed.
MutantpORl-SVI
has a 19-bpsynthetic T-antigen-binding
region
I correspondingto SV40 nucleotides 5186-5209(24a) inserted at the BamHI site of the polylinker. Plasmid pOR1-Ad2 contains the major late promoter within adenovirus 2 se-quences 5815 and 6035 between XhoI and EcoRI linkers insertedattheSall andEcoRI sites of thepolylinker.Mutant pORl-TATA hasan AA to CG substitution at SV40 nucle-otides 17 and 18 ofthe TATA box. It was constructedby
replacing theBglI-NcoI
fragment of pOR1 with thecorre-spondingrestrictionfragmentofmutantCW44ML obtained from B. K. Wasylyketal. (29).Plasmid
pORl-Enh
contains138
on November 10, 2019 by guest
http://jvi.asm.org/
Core
11,
TATA
1I11
21
bp
Enhancer
pOR4
5171
5235
N3
Bgl
I N,II
pOR3
BgI
I
A~~~
A A
II
BglI
pOR1
Hind
Nco EcoR
Nco Eco R
EARLY
PAL I
NDROMELATE
5171
5243/1
37AAGCTTTCTCACTACTTCTGGAATAGC:TCAGAGGCCGAGGCG6CCTCGGCCTCTGCATAAATAMAAAAATTAGTCAGCCATGG
TTCGAA
GTGATGAAGACCTTATCGAGTCTCCGGCTCCCCGGAGCCGGAGACGTATTTAII11111TAATCAGTCGGTACC
HindIlI Nco
TTGCAAAAGCCTAGGCCTCCAAAAAACT
AACGTTTTCGGATCCGGAGGITITIII
5178
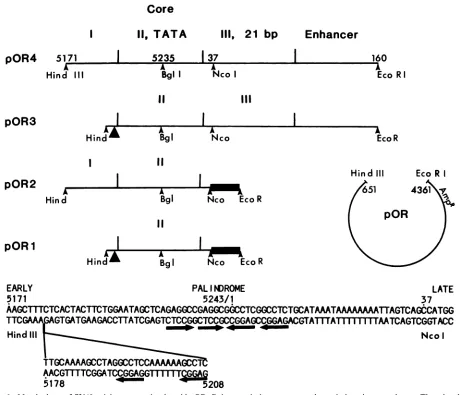
208FIG. 1. Matchedsetof SV40originmutantsinplasmid pOR. Relevantviralsequences and restriction sitesareshown.The trianglesin
pOR3andpOR1indicateaninternal deletion ofSV40sequences5178to5208. Theheavybars inpOR2andpOR1representa23-bp polylinker.
The arrowheads below the sequences locate recognition pentanucleotides for T antigen. Constructions were confirmed by dideoxy
-sequencing.
SV40 enhancer sequences 107 to 182 inserted between the BamHI and EcoRI sitesof thepOR1 polylinker.
DNAreplication.The assayfor DNAreplicationinCOS-1 cells has beenpreviously described (3).Transfection of 0.5 p,gofplasmidDNA per75-cm2 flask of cells falls within the
limits ofalinear dose-response curvefor replication under ourconditions. Transfected cells were maintained
continu-ouslyat 36°C. To insureequivalent transfection efficiencies witheachplasmidpreparation,DNAswerepurified twice by
equilibrium centrifugation in CsCl gradients containing ethidiumbromide. The input DNAs were quantitated both
by measuring the optical density at 260 nM and by densitometry of form I DNA separated in 1% agarose gels
and stained with ethidium bromide.
DNase footprinting. The DNase I footprinting assay has beendescribed in detailpreviously (4). In thepresentstudy, theEcoRI-HindIII originfragmentwaslabeledonthe 3' end of theearlystrandatthe EcoRI site. Tantigenwasboundto
DNA in 100 Rl of 20 mM piperazine-N,N'-bis(2-ethanesulfonic acid(PIPES) [pH7]-0.1 mMEDTA-50to100 mMNaCl. Afterbinding for 60 minat 0°C orfor 10min at
37°C, 5
RI
of DNase I in 20 mM MgCl2-10 mM CaCI2wasadded for 5 minatthe sametemperature asthat used inthe binding reaction. The DNase I concentrationsat0 and37°C
were 0.125 and0.0005 U perreaction, respectively.
RESULTS
Deletion of flanking regions from the core origin. We constructed amatchedsetof fourplasmidstoquantitatethe effects ofadjacent regulatorysequences on theefficiencyof
coreorigin replication (Fig. 1).Thewild-typeSV40 originin pOR4 contains all known origin components and includes T-antigen-binding region I, the core origin that overlaps T-antigen-binding region II,three 21-bprepeatsoftheearly promoterthatoverlapT-antigen-binding regionIII,andpart of the SV40enhancer. Previous studies have shown that the SV40originisindependentofenhancerfunction(1, 10, 19)in
the presence of the 21-bp sequences. Clone pOR3 has a
deletion of T-antigen-binding region I; clone pOR2 has a
deletion of theearlypromoterregion;andclonepOR1hasa
deletion of both regions.
We measured the replication efficiency ofthe clones by transfection of cloned DNA into COS-1 cells that constitu-tively expressed SV40 T antigen. After incubationat 36°C,
A
Hind III Ico I
160
III
Hind
A
Eco R I
pOR2
A
Nco
II
Hin d
A
Eco R
Bgl
HindIII Eco R I
I
I
I
I
I
I
I
i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.82.542.71.466.2]weextracted DNA atdaily intervals. The DNA was cut with MboI to distinguish methylated input DNAfrom unmethyl-ated progeny DNA and analyzed by gel electrophoresis, blotting, and hybridizationwithradiolabeledpBR322DNA. We used MboI fragments that were common to all of the clones to quantitate the amount of progeny DNA by densitometry ofautoradiograms.The residualinputDNAin the autoradiograms served as an internal control for the amount of plasmid DNA added to each
sample.
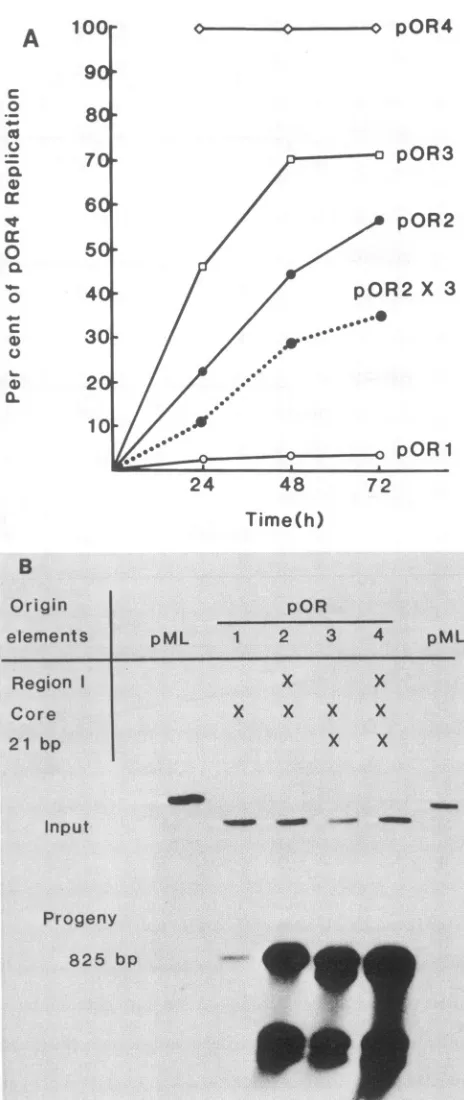
The polylinkers present in pOR1 and pOR2 had no effect on replication (data not shown). Yields at later times after infection are not shown because levels of progeny DNA decreased, presumablyas aresult of cell death and detach-ment caused byplasmid DNA replication.All deletion clones replicated more slowly and to lower levels than did wild-type pOR4. Fig. 2A summarizes the courseofDNA replication duringthe 3 days after transfec-tion, and Fig. 2B shows an example ofthegel blot
analysis
ofprogenyDNAyields72 h after transfection.Thedelay
in replication may account for the variations in estimates of levels of DNA replication reported in previous studies for similar deletions. At maximal levels ofreplication, origins
with deletions in T-antigen-binding region I replicated ap-proximately 70%aswell aswild-typeorigins.
Thisefficiency
ofreplication is similartothatofauthentic whole viralDNA with theidentical deletion (8). Deletion ofthe21-bp repeats reduced replication to 50 to60%
ofwild-type
levels. In contrast tothesingledeletions,
the simultaneous deletion of region Iand the 21-bp repeatscauseda moredrastic inhibi-tionofDNAreplication,toless than 5% ofwild-type
levels. Thus, thedoubledeletion reducedDNAreplicationtolevels that were at least sevenfold lower than theproduct
of the percentages ofwild-typereplication
forthe twosingle
dele-tions(shownbythedotted line inFig. 2A).Weobtainedthe sameresults with threedifferentbatchesofinput
DNA.This unexpectedfinding
indicates that the two regions either shareredundant functionsorcooperatein thesamefunction. Inboth cases, nonhomologoussequencesadjacent
toeither end of the coreorigin
would have at least one related function in theinitiation ofDNAreplication.
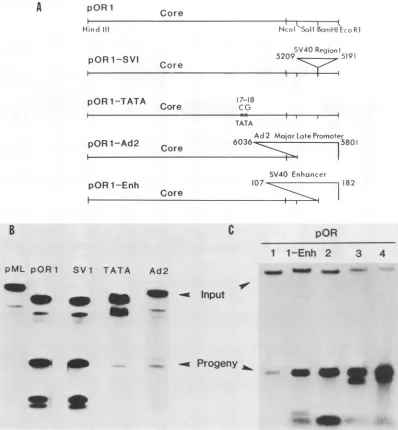
Rearrangement of regions flanking the core origin. To investigatethe role ofT-antigen-binding
region
I in replica-tion, we inserted a 19-bpsynthetic region
I with BamHIoverhangs
into the BamHI site of thepolylinker
ofpOR1
(Fig. 3A). The synthetic segmentcorresponds
to SV40 sequences 5191 to 5209, which contain tworecognition
pentanucleotides and a vital
7-bp
spacer(Fig. 1).
These signals are sufficient to direct thehigh-affinity
binding
ofTantigen
(24a). Theinsertionplaced
region
I50bp
tothe late side ofthecenterofthecoreorigin
rather than 34bp
tothe early side, at its usualposition.
Relative to the core, the polarity ofthesyntheticbinding
sequenceswasthesame as thatofthenaturalbinding
sequencesinwild-type pOR4 (Fig.
3B). The rearrangement failed to increase DNAreplication
abovepOR1 levels.Thus,
region
Imustbelocated eitheron theearlyside of the originor at aprecise
distancefrom the core tofacilitatethe initiation ofDNAsynthesis.
Substitution ofaforeignpromoter forthe21-bprepeats. To examine the role of promoter functions in
replication,
we substituted sequences within andadjacent tothe coreorigin
(Fig. 3A). Substitution of CG for AA sequences at nucleo-tides 17 to 18 of the TATA box reduced thereplication
of pOR1 to approximately 10% of core levels(Fig.
3B). Wasylyketal.(29)showed that this mutationcauses approx-imately a 20-fold reduction in transcription starting down-streamfrom TATA sequences, with nosignificant change in1d
A
'
9
C 0
-4-0
7
0
oS
o
4
C e
0)
a
0.
B
Origin
elements
_________ M4. ll
-Region
Core
21
bp
24 48 72
Time(h)
pOR
pML
1 2 3 4 pMLx x
x x x x
x x
Input
Progeny
825 bp
FIG. 2. Replication of
wild-type
and mutantorigins
in COS-1 cells. At various times aftertransfection, DNAwasextracted, cut withMbol, separatedbyagarosegelelectrophoresis,transferredtoGeneScreen,andhybridizedwithnick-translatedpBR322DNA.(A) Quantitative summary of levels of accumulated progeny DNA at various times aftertransfection. The dotted line shows the level of
replicationexpectedforthedoubledeletion of
pOR2
andpOR3
ifthe twodeletionsaffectedindependentfunctions.Itis theproduct
ofthe percentages ofwild-typereplicationfor thetwosingle
deletions.(B) Autoradiogram (4-h exposure)ofinputand progeny DNAsextracted 3daysafter transfection.Inputplasmids areidentifiedatthetop of thegel.Xindicates the presence of theorigin
elements.on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.323.555.65.615.2]Hind III
Core
Nco -SoIIBon,Hl Ec o R SV40Region
5209 5191
Core
pORl1-TATA
17-18Core
CG
TATA TATA
pORl-Ad2
pOR1-Enh
H
-Core
Ad 2 Major LatePromoter
6036 , - 5801
SV40 Enhancer
107 182
Core
C
pOR
1
1-Enh 2
SV 1 TATA Ad2 In1 j ~ ino mw,
_
,
Inputoa
-s
Os_
Progeny [image:4.612.108.506.76.506.2]SR
I.
FIG. 3. Replicationof thecoreorigininartificial sequencecontexts.DNAwasextracted from COS-1 cells 72 h aftertransfection, and
assays were performed asdescribed in the legend to Fig. 2. (A) Summary of the origin constructions tested. (B) Autoradiogram (24-h exposure)ofinputandprogeny DNAs; input plasmidsare identifiedatthetopof thegel. (C)Autoradiogram (1-h exposure) ofinputand
progenyDNAs;input plasmidsareidentifiedatthetopof thegel. SeeFig. 1 and 3A foradescriptionof theinput plasmids. the amount of RNA initiating from upstream sites in vivo,
andreduces DNAreplicationabout fivefold in COS-1 cells. Because nucleotides 17 to 18 are components of both the TATA sequence and the core origin, the cause of the decrease in replication is ambiguous. The mutation could affectoriginfunction eitherbyaltering transcription orbya
mechanismindependent oftranscription. Nevertheless, it is significant that all promoter elements can be deleted or
mutated without completely abolishing DNA synthesis. If transcription is essential for the activation of replication, then transcripts originating from foreign sequences in the plasmidmaybecapableofreplacingnaturalpromoter func-tion in the replication process. To test this possibility, we
inserted thestrongadenovirus 2majorpromoterbetween the EcoRI and SalI sites of the polylinker (Fig. 3A). In this construction, the TATA sequences of the adenovirus
pro-moterare located40 bp upstream from the natural TATA
box in the coreorigin. The substituted promoter inhibited replication5-to 10-foldascomparedwiththat ofpOR1 (Fig. 3B).Wedo notknow how the adenovirussequencesinhibit theSV40 origin. Nevertheless,theinhibitionemphasizesthe needtousematchedplasmids tocompare mutant origins.
Substitution of a 72-bp repeat for the 21-bp repeats. Al-though the SV40 enhancer elements are not required for replication in the presence ofthe 21-bprepeats (1, 10, 19), the enhancers of the relatedpolyomavirus have been impli-cated in DNAreplication (6, 23).Totestthefunction ofthe SV40 enhancer in the absence of the 21-bp repeats, we
inserted it between the BamHI and EcoRI sites of thepORi polylinker(Fig. 3A).Theinsertionplacesthe enhancer inits usualorientation 13bptothelate side of the NcoI site of the
core origin. Thus,the 72-bp element islocated at the same
site as the SV40 T-antigen-binding region I rearrangement discussed previously but 13 bp farther upstream from the
A
pOR 1pOR 1-SVI
B
pML pOR1
3
4
on November 10, 2019 by guest
http://jvi.asm.org/
00C,
100 mM NaCipOR 1 pOR2
37°0C,100mM NaCI
pOR1 pOR2
370C, 50mM NaCI
pORI pOR2
0 25 50 0 25 50
5mm~m
_ __M
a 4a
O 25 5Q O 55
*?
or,.s
:," .;.
amW:.'.. : . am
an so
*.._~~~~~~~f..
mwmwa.OM
_ Q
4-.
.L
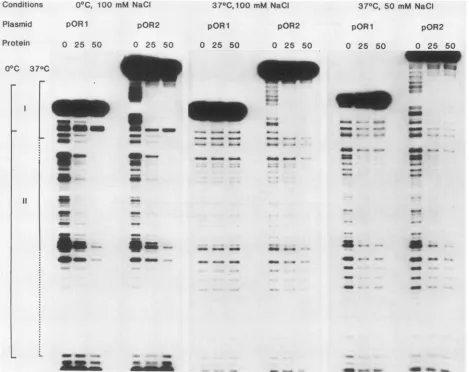
FIG. 4. Binding ofTantigentocoreoriginDNAinthepresence(pOR2)andabsence(pOR1) ofregionI.Increasingamounts(nanograms)
ofprotein (T-antigen)wereaddedtoafixedamountof DNA (0.1 ng)from eachmutantunderavariety oftemperatureandionic conditions,
asindicatedatthetopofthefigure. Afterbindingateither0or37°C,theappropriateamountof DNase Iwasaddedtoeach sampleatthe
same temperature. The reaction was stopped, andend-labeled cleavage products were analyzed in denaturing gels. The boundaries of DNase-protected regions I and1I aresummarizedattheleftof thefigure.Thelate limitof regionII shownatthebottom ofthefigurewas
difficulttodefine byusingthefootprinting assay becauseofa paucity ofDNase I sitesin thatregion. Itwasestablished with afragment
survivalassay inthepresenceofexcessDNaseI (27).
usual position of the 21-bp elements. The 72-bp enhancer sequence stimulated replication 5-fold abovecorelevels, or
approximately halfaswellasthe21-bprepeatsinpOR3 (Fig. 3C). The successful interchange of 21- and72-bp functions suggests that they have arelated role inreplication.
Effectsofflanking regionsonT-antigen bindingtothecore
origin. We also examined the effects ofadjacent sequences
onthebinding ofTantigentoregionIIwithinthecoreorigin.
Although bindingtoregionIIisindependentofregionsIand III atlowtemperatures(17, 27),recent findingssuggestthat region II binding interactions may be altered at 37°C (9). Figure4showsDNase footprintsofT-antigen bindingtothe origin under a variety of conditions. In contrast to the findingsofDixonandNathans (9),Tantigenboundtoregion Iwithagreateraffinity thanto regionII at37°C. Wedonot knowthereasonforthis difference. At 0°C, bindingtoregion lI wasequivalentinthepresence(pOR2)orabsence(pOR1)
of region I. At 37°C, the presence of region I enhanced
bindingtothecoretoa smallextent inboth 50 and 100mM NaCl. Interestingly, temperature determined the extent of
DNase I protection by Tantigen. At 37°C, region Iwidened, and region II became morefocused than at lower
tempera-tures. T-antigen bindingto regionIII had no effecton core binding underany conditionsused (datanot shown). Thus, the slight enhancementofcore binding by region I at 37°C may explain the slight decrease in DNA replication in the absence of region I, but it does not explain the drastic impairment after the deletion of both regionI and the 21-bp repeats.
DISCUSSION
Deletions beyond limits on either side of the T-antigen-binding palindromeof the SV40 originabolish DNA replica-tionaltogether (10, 24).We define these essentialsequences
asthe coreorigin. In contrast,complete deletions ofeither T-antigen-binding regionIorthe21-bprepeatsreduce butdo not eliminate replication. Thus, these regions flanking the earlyand late ends of thecoreoriginhaveafacilitating rather than an obligatory function. If the two regions performed independent functions inreplication, theirsimultaneous de-Conditions
Plasmid
Protein 0 25 50
OOC 370C
r-0 25 50
I1
..- -- --..o 4m...m Am---w
-.:- qg.".A
4mmmo ammm OEM*
M-4wom
4m 0"6400
40M
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.82.551.72.444.2]letionwould result inalevel ofreplication equivalenttothe
product
of the percentages ofwild-type replication for the two individual deletions. Because the level resulting from thecombined defect is far less than this level, we conclude that theflanking regions
have at least one related function. Furthermore, theability
of the 72-bp enhancer element to replace the 21-bp repeats in replication suggests thatit also shares arelatedfunction withregion
Iand the21-bprepeats. The three distinct regulatoryregions
that facilitate coreorigin
function areimportant
componentsof the SV40 pro-moter. The 21- and 72-bp repeats bind positive regulatoryproteins,
whereas region I binds the T-antigenrepressor(4,9, 13,
14, 17, 24, 24a, 30). Together, theregions
regulatethe level of transcription and determine the initiation sites for early mRNA (10, 12-14, 30). These findings suggest apossible
role fortranscription
in the initiation of DNAreplication.
However,this idea is difficulttoreconcile witha numberoffindings
atpresent. The insertion of the adenovi-rus 2major
late promoter upstream from theorigin
inhibits DNAreplication
rather than facilitates it. Furthermore, DNAreplication
continues, although at lower levels, after deletion or mutation of all known elements of the early promoter-operatorregion, including
the TATA sequences within the core origin. Finally, Li and Kelly showed that oL-amanitin does not inhibit SV40 DNAsynthesis
signifi-cantly
in vitro (20).Thus,
thequantity
oftranscription
does not correlate well with the levels ofreplication.
Neverthe-less,
it remainspossible
that low levels oftranscription
are sufficient forreplication.
Afurtherpossible requirement
for the correct initiation oftranscription might explain
the role of region I inreplication
as well as the inhibition ofreplica-tion
by
aforeign
promoter.SV40
T-antigen-binding region
I, the 21-bp repeats, and the 72-bp repeat share a second common feature. Eachregion
bindsregulatory
proteins
that may facilitate events within the coreorigin
(4, 13, 30). The facilitation could reflect adirect interaction between proteins. Forexample,
Tantigen
bindstoregion
Iinaposition
that may allowcontact withTantigen
boundtothecoreorigin. Indeed,
the presence ofregion
I increasedslightly
thebinding
ofTantigen
tothecore at
37°C
in vitro. Because Tantigen
coversonly
the centerpalindrome
of theorigin
at37°C,
thebinding
of T antigen toregion
Iorof thepositive regulatory protein Spl
to the
21-bp
repeats (13) could alsodirectly
facilitate thebinding
of cellular proteins to theadjacent
coreregion.
Alternatively,
the tightbinding
ofproteins
oneither sideof theorigin
may preventtheformation of nucleosomes within the core DNA (2, 11, 16, 18) and indirectly increase the access of viral orcellularproteins
tothe open coreregionof viral chromatin. Ifproteins
bound to flanking regions doorganize
chromatin to allowreplication,
then the location and nature of boundproteins
must be critical because substitution ofenhancer,
but notregion I, sequencesfor the21-bp
repeats restoresreplication
function.Although generally
considered part ofanoncodingregu-latory region,
the SV40 origin contains an open reading frame for 23 amino acids between nucleotides 5191 and 16(28, 29).
Itisnotknown whether thispotential peptideexistsor would have any function in replication. Mutant pOR1 would removethe21- and 72-bprepeats ofthepromoter for the
peptide
and wouldalsochangeitsreadingframe through the deletion of region I sequences. Thus, both deletions would reduce the function ofa single gene product.The SV40 regulatory sequences are organized for com-pactness and
efficiency.
The early and late promotersover-lap
each otheras wellasthe originofreplication. A numberof viral and cellularregulatory proteinsrecognizesequences in region I, the core origin, the 21-bp repeats, and the enhancers (4, 9, 13, 14, 17, 24, 24a, 30). These proteins exclude nucleosomesdirectly and perhaps at a distance (2, 11, 16). We show here that three distinct regions separated by over 70 bp cooperate ina commonfunction that facilitates replication. The intricacies of this integrated control region present achallengeandanopportunity for theanalysis of the interactions among eucaryotic control mechanisms. The isolationofasmall,autonomous coreoriginsuggeststhat the SV40 controlregioncanbe dissected into individual compo-nents for a detailed genetic analysis.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grants CA-18808.CA-28146, and CA-09121 from the National Institutes of Health. We thank R. Dixon and D. Nathans for SV40 mutant CS-1097,B. Wasylykand P. ChambonformutantCW44ML, andJ. Logan and T. Shenk for the plasmid containing the adenovirus 2 major late promoter.
ADDENDUM IN PROOF
J. J. Li, K . Peden, R. Dixon, and T. J. Kelly also have shown that, in the absence of the 21-bp repeats, the SV40 enhancers stimulate DNAreplication in vivo butnot invitro (personal communication).
LITERATURECITED
1. Bergsma, D. J., D. M. Olive, S. W. Hartzell, and K. N. Subramanian.1982. Territorial limits and functional anatomy of the simian virus 40 replication origin. Proc. Natl. Acad. Sci. USA 79:381-385.
2. Cereghini, S., and M. Yaniv. 1984. Assembly of transfected DNAinto chromatin: structuralchanges in the origin-promoter-enhancerregion uponreplication. EMBO J. 3:1243-1253. 3. Cohen,G.L.,P.J.Wright,A. L.DeLucia, B. A. Lewton,M.E.
Anderson, and P. Tegtmeyer. 1984. Criticalspatial requirement within theorigin of simian virus 40 DNA replication. J. Virol. 51:91-96.
4. DeLucia,A.L.,B. A.Lewton,R.Tjian, and P. Tegtmeyer. 1983. Topography of simian virus 40 A protein-DNA complexes: arrangement ofpentanucleotideinteraction sites at the origin of replication.J. Virol. 46:143-150.
5. DePamphilis,M.L.,andP. M. Wasserman.1982. Organization andreplicationof papovavirusDNA,p. 37-114.InA.S. Kaplan (ed.), Organization and replicationof viral DNA. CRC Press, Inc., Boca Raton. Fla.
6. DeVilliers, J., W. Schaffner, S. Lupton, C. Tyndall, and R.
Kamen. 1985. Enhancerfunction is essential forpolyomavirus DNAreplication. Nature(London)312:242-246.
7. DiMiao, D., and D. Nathans. 1980. Cold-sensitive regulatory
mutantsof simian virus 40. J. Mol. Biol. 140:129-142. 8. DiMaio,D.,and D. Nathans.1982.Regulatorymutantsof simian
virus 40.Effectof mutations at a T antigen binding site on DNA replication and expression of viral genes. J. Mol. Biol. 156:531-548.
9. Dixon,R.A.,and D.Nathans.1985. Purificationof simian virus 40largeTantigen by immunoaffinity chromatography.J. Virol. 53:1001-1004.
10. Fromm, M., and P. Berg. 1982. Deletion mapping of DNA regions required for SV40 early region promoter function in
vivo.J. Mol. Appl. Genet. 1:457-481.
11. Gerard,R. D.,B.A. Montelone, C. F. Walter, J. W. Innis, and
W. A. Scott.1985. Roleofspecificsimian virus 40 sequences in the nuclease-sensitive structure in viral chromatin. Mol. Cell. Biol. 5:52-58.
12. Ghosh, P. K., and P. Lebowitz. 1981. Simian virus 40 early
mRNA's containmultiple5'termini upstreamanddownstream froma Hogness-Goldbergsequence: ashift in 5' terminiduring
on November 10, 2019 by guest
http://jvi.asm.org/
the lytic cycle is mediated by large T antigen. J. Virol. 40:224-240.
13. Gidoni, D., W. S. Dynan, and R. Tjian. 1984. Multiple specific contacts between a mammalian transcription factor and its cognate promoters. Nature(London) 312:409-413.
14. Hansen, U., D. G. Tenen, D. M. Livingston, and P. A. Sharp. 1981. Tantigen repression ofSV40early transcriptionfrom two promoters. Cell27:603-612.
15. Hay, R. T., and M. L. DePamphilis. 1982. Initiation ofSV40 DNA replication in vivo: location and structure of5' ends of DNAsynthesized inthe oriregion. Cell28:767-779.
16. Innis, J. W., and W. A. Scott. 1984. DNA replication and chromatin structure of simian virus40insertionmutants. Mol. Cell. Biol.4:1499-1507.
17. Jones, K. A., R. M. Myers, and R. Tjian. 1984. Mutational analysis of simian virus40largeTantigen binding sites. EMBO J. 3:3247-3255.
18. Jongstra, J., T. L. Reudelhuber, P. Ouden, C. Benoist, C.-B. Chae, J.-M. Jeltsch, B. J. Mathis, and P. Chambon. 1984. Induction ofaltered chromatin structures by simian virus 40 enhancer and promoter elements. Nature (London) 307:708-714.
19. Learned, R. M., R. M. Myers, and R. Tjian. 1981.Replication in monkey cells of plasmid DNA containing the minimal SV40 origin, p. 555-566. In D. S. Ray and C. F. Fox (ed.), The initiation ofDNAreplication. Academic Press, Inc.,NewYork. 20. Li, J.-J., and T. J. Kelly.1984.Simian virus40 DNAreplication
in vitro. Proc. Natl. Acad. Sci. USA81:6973-6977.
21. Li, J.-J.,andT. J. Kelly. 1985. SV40DNAreplication in vitro: specificity of initiation andevidence for bidirectional replica-tion. Mol. Cell. Biol.5:1238-1246.
22. Lusky, M., and M. Botchan. 1981.Inhibition of SV40 replication
in simiancells byspecificpBR322 sequences. Nature(London) 293:79-81.
23. Muller, W. J., C. R. Mueller, A.-M. Mes, and J. A. 1lassell. 1983. Polyomavirus origin forDNAreplication comprises mul-tiple geneticelements. J.Virol.47:586-599!
24. Myers, R. M., and R. Tjian. 1980.Constructionand analysisof simian virus40origins defective in tumor antigen binding and DNA replication. Proc. Natl. Acad. Sci. USA 77:6491-6495. 24a.Ryder, K., E. Vakaiopoulou, R. Mertz, I. Mastrangelo, P.
Hough, P. Tegtmeyer, and E. Fanning. 1985. Seventeen base pairs ofregionIencodeanoveltripartite binding signal for SV40 Tantigen. Cell 42:539-548.
25. Stillman, B. W., and Y. Gluzman. 1985. Replication and supercoiling of SV40 DNA in cell-free extracts from human cells. Mol. Cell. Biol. 5:2051-2060.
26. Subramanian, K. N., and T. Shenk. 1978. Definition of the boundaries of theoriginof DNAreplication in simian virus 40. NucleicAcidsRes. 5:3635-3642.
27. Tegtmeyer, P., B. Andersen, S. B.Shaw, and V. G. Wilson. 1981. Alternative interactions of the SV40 A protein with DNA. Virology 115:75-87.
28. Tooze, J. (ed.). 1980.DNA tumorviruses,molecularbiologyof tumor viruses, part 2. Cold Spring Harbor Laboratory, Cold Spring Harbor,N.Y.
29. Wasylyk, B. K., C.Wasylyk,H.Matthes, M. Wintzerith, and P. Chambon. 1983. Transcription from the SV40early-early and late-lateoverlappingpromotersinthe absence ofDNA replica-tion. EMBOJ. 2:1605-1611.
30. Wildeman, A.G., P.Sassone-Corsi, T.Grundstrom,M. Zenke, and P.Chambon. 1984.Stimulation ofinvitrotranscription from the SV40 early promoterby the enhancer involves a specific trans-acting factor. EMBO J. 3:3129-3133.