0022-538X/91/031228-08$02.00/0
CopyrightC) 1991, AmericanSociety forMicrobiology
Differential Induction of Structural Changes
in the Simian Virus
40
Origin of Replication by T
Antigen
JAMES A. BOROWIEC,t FRANK B. DEAN,AND JERARDHURWITZ* Program in Molecular Biology andVirology, MemorialSloan-KetteringCancerCenter,
New
York,
New York 10021Received 20September 1990/Accepted 20 November 1990
TheATP-dependent binding of the simian virus40(SV40)largetumorantigen (T antigen)to theSV40 origin ofreplication(on)results in thestructural distortionoftwo critical elements withinflanking regionsofonand the untwisting of the DNA helix. We examined the effect ofchangesintemperature, ATPconcentration, and other reaction parametersonthegeneration of these DNA structuralchanges.Wefound that induction ofthe twolocalized structural transitionswere highly anddifferentially sensitiveto reactionconditions. Significant distortion ofthe early palindromeelement, shown previouslyto resultfromDNAmelting, requiredlowlevels ofATP(10 to 30,uM) but temperaturesabove25°C.Distortion of the AT tract occurred at lowtemperatures (5°C) but required relativelyhigh concentrations ofATP(>300 ,LM). Thus,T antigencaninducestructural changes withinonecriticalelement ofonwithoutgeneratingsignificant structural distortionwithinthesecond element. The response ofori untwisting to reaction conditions generally increased in parallel with or fell intermediate between the inductions of localized structural transitions. We suggest that on untwisting and localized structural distortions are interdependent consequences ofT-antigen binding to oni. These results suggestamodel for the structural events occurring during the initial steps ofSV40 DNA replication.
The initiation ofDNAreplication requiresgross changes
in theduplex DNA structurecomprisingtheorigin of repli-cation. The DNAmustbedenaturedtoallowtheenzymatic machinery access to the bases in order to synthesize the complementary strand. The nature of the protein-DNA interactions thatcan generate sufficient force to melt DNA
sequences withinanorigin remainspoorly understood. We
are studying the initiation ofDNA replication using simian
virus 40(SV40) with the goal of understanding the molecular events required to achieve large-scale denaturation ofan originofreplication.
SV40iswidely usedasaeukaryoticmodelsysteminvitro to examine the mechanistic steps constituting the initiation of DNA replication (1, 6, 23). The SV40 core origin of replication (ori) is contained within a fairly small region (approximately 65 bp),andregionscritical for DNA replica-tionhavebeendefined bydetailed genetic investigation (10, 12). The initiation of DNA replication requires sequence-specific binding to oriof the SV40 large tumor antigen (T antigen), the only virus-encoded protein required for repli-cation of the SV40 chromosome. Efficient binding of T antigento oriat elevated temperatures (e.g., 37°C) requires ATPandresults inacomplex thatprotectsthe completecore
ori sequencefrom DNase I digestion (2, 9, 11). This
ATP-dependent complex containsupto12monomersof T antigen arrangedinastructurewithadistinctbilobedappearance (9, 17). The addition of single-stranded DNA-binding protein (SSB; e.g., human SSB or Escherichia coli SSB) to the complex induces stable denaturation of ori inan ill-defined
process and results in bidirectional unwinding of DNA
outwardfromoribecauseof the DNA helicase activity ofT antigen (8, 14, 15, 26).
Chemical probing of the ATP-dependent complex has
*Corresponding author.
t Present address: Department ofBiochemistry, New York Uni-versity Medical Center, New York, NY 10016.
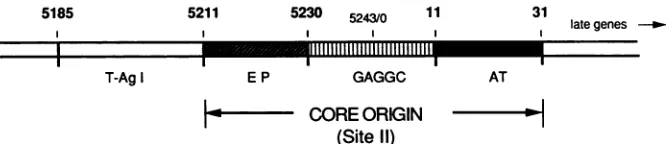
shown that the binding of T antigen induces structural alterations oftwoDNAelements in thecoreori (Fig. 1)(3, 21). In the early-gene side ofori, T antigen melts approxi-mately 8 bp of DNA partially contained withinone armofa
15-bp imperfect inverted repeat (the early-palindrome ele-ment).Thesecondsignificant changeisfoundwithina17-bp
AT tract that becomes structurally distorted, although the DNA remains essentially double stranded in character. These two elements flank a central region containing four conserved GAGGC repeats that are thought to bind and properly position T antigen for induction of the structural changesand for theunwinding ofori (3, 21). Each ofthese three regions is essential for events occurring during the initiation ofreplication; mutation ofanyelement is deleteri-ousboth toSV40DNAreplicationand tounwindingofDNA containing ori (7, 12).
Topologicalassaysmeasuringother aspectsof DNA struc-ture, namely untwisting of the DNA helix mediated by T antigen, have also been developed (9a, 22). These studies show that T antigen significantly untwists the DNA helix (increasesthe numberof basepairsperhelicalturn)
concur-rently with theformation of the ATP-dependent complex. While certain of the structural changes found by chemical probingwould be expectedtountwistthe DNAhelix (e.g., DNA melting), the relationship between DNA structural changes detected chemically and those detected by alter-ations in DNAtopology has notbeen determined.
We haveexamined theinfluence of ATP levels, tempera-ture, and other factors on the structural distortion of ori mediated by Tantigen. We show that the ATP-dependent complex of Tantigenboundto oricandifferentially induce
localized structural distortions withintwocritical regions of ori. We also show thatchanges in DNAtopology generally parallel DNA structural alterations that are detectable by
chemical probing. These results suggest that changes in DNA topology and structural alterations in the early palin-drome and AT-tract regions arerelated events arisingfrom thebinding of Tantigen.
1228
on November 10, 2019 by guest
http://jvi.asm.org/
5185
T-Ag
5211 5230 5243/0 11 31
late genes
-_.-I I I r
E P GAGGC AT
H- CORE
ORIGIN
- H(Site 11)
FIG. 1. Schematic of theSV40 origin of replication. TheSV40coreoriextends fromSV40positions 5211 to 31 (10) of the 5,243-bpSV40 genome. The core ori is considered here to be identical to T-antigen-binding site II. The three regions within the oricritical for DNA replicationare theearly palindrome element (EP), a central palindrome containing four GAGGC repeats (GAGGC), and a 17-bp AT tract (AT).Located outside the coreoriisT-antigen-binding site I.Theapproximate location of each region is indicated by the SV40sequence positionsabove the diagram. The late-gene side oforiis tothe right.
MATERIALS AND METHODS
Preparation ofT antigen. Tantigen was purified from Sf9
insect cells infected with recombinant baculovirus strain vEV55SVT (20). Forty-eight hours postinfection, the Sf9 cellswerecollected andpelleted. The cellswere suspended
in5 ml of lysis buffer(20 mMTris-HCl [pH9.0], 0.3 MNaCl, 1 mM EDTA, 10% glycerol, 0.5% Nonidet P-40, 1 mM dithiothreitol, 0.2pLgof aprotininperml, 0.2 ,ug of leupeptin
perml, 0.1 ,g ofantipain per ml, 50 ,uM EGTA)per mlof pellet volume. After incubation at 0°C for 15 min, the suspension wascentrifuged at40,000 x g for30min (4°C).
Thesupernatant was neutralized with aone-halfvolumeof
neutralization buffer (lysis buffer containing 100 mM Tris-HCl [pH 6.8]), and the lysatewas then stored at -80°C. T antigenwasimmunoaffinity purified from this lysate accord-ing tothe method of Mastrangelo et al. (17).
KMnO4footprinting of DNA. Standard reaction mixtures (30
[lI)
contained 40 mMcreatinephosphate (di-Trissalt,pH 7.8), 7 mM MgCl2, 0.5 mM dithiothreitol, 0.5 ,ug of pSVO1AEP DNA (25), 4 mM ATP, 23.3 ,ug of creatine phosphokinase per ml, and 1.0 ,ug ofT antigen. Reactionswere incubatedat 37°C for 60min priortomodification with KMnO4. KMnO4 was added from afreshly prepared stock
solution (60 or 200 mM) to give a final concentration of 6 mM,and thereactionwasincubatedanadditional4min. The primer extension and gel electrophoresis conditions were essentially as described by Borowiec and Hurwitz (3). For
kineticexaminationof changes in DNAstructureinduced by Tantigen, the concentration of KMnO4 used to modify the DNA was increased to 20 mM (final), and the incubation time withthis chemical probewas shortenedto 30s.
Thelevelof KMnO4 oxidation in each regionwas
quanti-tated by densitometric scanning of autoradiographs with a
Helena Quick Scan densitometer. Autoradiographs with various exposure times were used to ensure that the band intensitywaswithinthe linearrangeoffilm sensitivity. The extent of KMnO4 oxidation of the early palindrome was
determinedbyquantitatingtheintensityoftheregion corre-sponding to top-strand thymine residues 5210, 5214, 5217, and5218. For the AT tract, bandscorrespondingtothymine residues16, 20, 29, and 30 of the top strandwereused.The overall intensity ofeach region was separately normalized by comparison with the intensity oftwo control thymine residues(SV40 positions5159 and5163)located outside both the SV40coreori and T-antigen-binding site I (adjacentto the core ori). The maximum normalized value fortheearly palindromeand the ATtractfor each titrationwassetat 1.0. Thefraction of maximum oxidationwas then calculated for eachpoint.When akineticanalysiswasperformed, regions
of the AT tract proximaland distal to the central GAGGC elementwereexaminedseparately.Theproximal regionwas
considered tominimallycontain the region encompassed by thymine residues located at SV40 positions 16 and 20; the distal region contained thymines at positions 29 and 30.
Topoisomer shift assay (DNA untwisting). Standard
reac-tion mixtures (30
RI)
containing40 mM creatine phosphate(di-Tris salt, pH 7.7), 7 mM MgCl2, 1 mM dithiothreitol, 4 mM ATP, 10 U of HeLatopoisomerase I, 0.9
jig
of bovine serumalbumin, 0.3jig
ofrelaxedpSV01AEP DNA, and 0.75 ,ugofTantigenwereincubated for 45 min at 37°C.Reactionswereterminated by the addition of a solution (5 ,ul) contain-ing 2% sodium dodecyl sulfate (SDS), 0.1 mM EDTA, 1 ,ug
of glycogen, and 5
pLg
of proteinase K followed by anadditional incubation for 30 min at37°C. Following ethanol
precipitation,theproducts were subjected toelectrophoresis
through agarose gels (1.8%) containing chloroquine phos-phate (2 to4,ug/ml).The ethidium bromide-stainedgelswere
photographed, and thephotographic negativeswerescanned
with an LKB 2202 Ultrascan laser densitometer. Therelative
amount of DNA contained in each topoisomer band was
determined, and the (relative) average linking numberwas calculated. The change in the average linking number was thencalculated andnormalizedwith respecttothemaximum
linkingnumberdifference determined foreachexperiment.
RESULTS
Weexaminedtheeffectsof various reactionconditionson the ability of T antigen to distort the structure of DNA.
Conformational changes in oriwereexamined bytwo
com-plementary methods. Local structural transitions were
probedbychemical modification withpotassium permanga-nate (KMnO4), whiletheamount ofDNAuntwisting gener-atedbyT antigenwas examinedby quantitating changes in thedistribution ofplasmid topoisomers.
DNA structuralchangesas afunctionof ATPconcentration. The effect of ATP concentration on
T-antigen-mediated
DNA structural changes was examined (Fig. 2). Localized
structural distortions in the early-palindrome element and the ATtract can be detected by
potassium
permanganate(KMnO4) oxidation ofT-antigen-ori complexes (3). While
KMnO4 reacts poorly with DNAin a
typical
B-DNAcon-formation,
significant changes
in the structure ofthe DNAduplex (e.g., DNA
melting
orpronounced
DNAbending)
leadtohypersensitivitytoKMnO4 oxidation,
predominantly
at thymineresidues (4).T-antigen-cri
complexes wereformed in the presence ofincreasing amounts ofATP, and the complexes were then modified with
KMnO4.
The reactive sites within ori weredetermined by the extension of a 32P-end-labeled DNA
primerthroughthe ori
region
with the Klenowfragment
ofE. coli DNApolymerase I, which stalls atoxidizedthymines.
The sites of
modification,
corresponding
toregions
ofDNAon November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.137.470.72.144.2]4- + + ± 4- 4- + + + __
C C)D CD) C C)
O)C)-= cC) C) C)C) C)- Cco .-CD C)C) C)Cm
B
T A n ,C + + 4- --r ... ±+
A1TP c.,2fl(D
-
5217--
5225-*...:a
4-16 _..
209.-29._ 30"
so MO
EP
GAGGC
-1r
e- 3 4 _ 9_0
AT
1 2 3 4 5 6 7 8 9 10
C
~0
om
Xz
x na
o s
NtT _
O--o (D
1l 1.el00 0 1000
:001-[ATP], ,M
with significant distortion, were visualized by autoradiogra-phy ofthe primer extension products separated on aDNA sequencing gel. While modification of either strand can be
examined by using an appropriate oligonucleotide primer, we have found that a primer complementary to the top
strand (the adenine-rich strand in the AT tract) is most
informative for examination oforiDNA conformation. ori-containing DNA incubated with T antigen in the ab-sence ofATP was poorly oxidized by KMnO4, consistent
withpastprobing studies(Fig. 2A, lane 1) (3). As the level of ATP in the reaction was increased from 0 to 3 mM, the
early-palindrome and AT-tract elements were found to
re-spond at different nucleotide concentrations. The early-palindrome element was reactive to the chemical probe at lower concentrations of ATP, with significant reactivity observedat 10 ,uM ATP. Significant modificationof the AT tractwas notobservedatconcentrations of ATP below 100 ,uM. Theearly-palindrome elementthusrequireslower ATP levels than the AT tract to undergo structural transitions detectable by this method. In addition, while the overall sensitivity of each ori regiontochemical attackwasstrongly
[image:3.612.68.300.80.480.2]influenced by the ATP concentration, the relative intensity
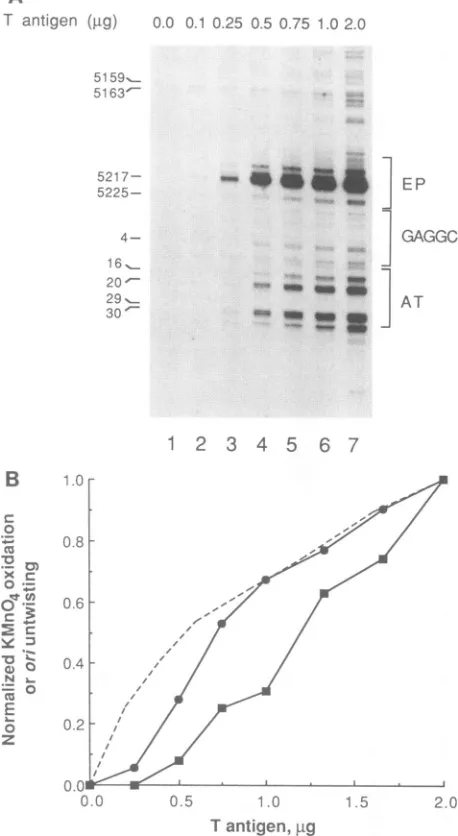
FIG. 2. Response ofT-antigen-dependent structural changes in oritoATP concentration.(A) Representativeautoradiograph show-ingtheKMnO4oxidation of orias afunction of ATP concentration. FormIpSVO1AEP (containingtheSV40ori) (25)wasincubated with
1.0,ugof Tantigenandvarious concentrations of ATP(asindicated)
at37°Cfor 1 h. The DNAwasthen modified withKMnO4,and the sitesof modificationwereprobedby primer extensionasdescribed
in Materials and Methods. The locations of the three critical ori regionsare shownon the right side of theautoradiograph andare
designated as follows: EP, early palindrome; GAGGC, central element containing four GAGGC repeats; AT, 17-bp AT tract.
Various SV40sequence positions are shownon the left. (B) DNA untwistingmediated byTantigen. Form IpSV01lEP(0.3 ,ug)was
incubated with 0.75 jigofT antigen and various concentrations of
ATP (as indicated). Topoisomerase I was also present in each reactiontorelieveany DNAsupercoils generated by thebinding of
T antigen. After incubation, the product DNA was subjected to agarose gel electrophoresis in the presenceof2 ,ug ofchloroquine phosphateperml. ThepositionofformII(nicked) DNA is indicated
ontheleft. (C) Quantitation of ori structural changes. The levels of DNAuntwisting and KMnO4 oxidation in the early palindrome and
ATtractsweredetermined and normalizedasdescribed inMaterials
andMethods. Symbols:0,early palindromeoxidation; *, AT-tract
oxidation; A, DNA untwisting. Maximum DNA untwisting was
foundat 3 mMATP,atwhichpoint 1.3 turnswereremoved.
of each thyminewithinasingleelementwas notaffected by nucleotide concentration. This suggeststhat thechanges in ATP level do notcause distinct structural transitions, but rather affecteither the fraction of ori molecules undergoinga structuraltransition orthe degree ofdistortion within each orimolecule.
The extent of KMnO4 modification in the early-palin-drome and AT-tract regionswasquantitated by densitomet-ric scanningof the autoradiographs. The levels of modifica-tionwereseparately normalizedby comparisonwithcontrol
bands located outside the ori region. Thefraction of maxi-mum KMnO4 oxidation in each region was plotted as a function of ATP concentration (Fig 2C). The structural distortions within each elementhadasigmoidal dependence onATP levels andwerefoundtoplateauat high concentra-tions ofnucleotide.Comparison ofthetwocurvesconfirmed
thedifferentialresponseofeach regiontoATP. The
concen-tration of ATP resulting in 50% maximal reactivity was
found tobe 35 ,uMfor theearlypalindrome, while fivefold-higherlevelsof ATP(175 ,iM)wererequiredtoachieve 50% maximum reactivity in the ATtract.
Theeffect of ATPconcentrationontopological changesin
DNA structure induced by T antigen was examined. ori-containing plasmid DNA wasincubated with Tantigen and variousconcentrations ofATP. HeLa topoisomerase I was
A
T Antigerr
[ATP], piM
51 59..
5163-C."' C',
...+
on November 10, 2019 by guest
http://jvi.asm.org/
A
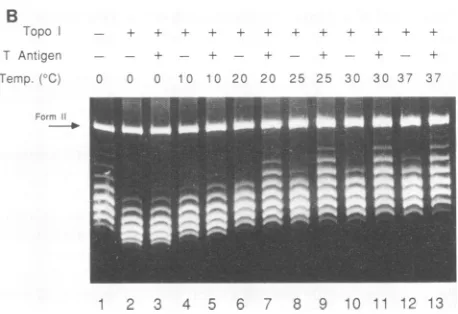
T Antigen
Temp. (0C)
+ + + + +
+-0 10 20 25 30 37 37
5159...
51
63-5217- tj o_ *
-5225- _ _ Om
4-16 ,
20-29._.. 30
.-_ bK _
ar-B
ToC.! -e - -+
TAr iger - -- -r - -. -- +
Tero. C u 0 IC '"d 2C 20 25 25 30 30 37 37
EP
1 2 3 4 5 6 71 8 9 10 11 112 13 GAGGC
AT
1
2
3
4
5
6
7
present in each react generated by T-antige catalyzegross DNA cules in thesereactioi incubation, the react subjected to agarose
chloroquine to allow topoisomer distributic Increases in the cc
caused the distributi broaden and shiftupA lane 9 and 10).Since Tantigeninanucleor
shift in topoisomer d antigen untwisting th topological changes q
crease in the numbe
FIG. 3. Influence oftemperature on T-antigen-dependent struc-tural changes in ori. (A) Representative autoradiograph of the KMnO4 oxidationpattern inori as a function of incubation temper-ature. Form I pSV01AEP(0.5 ,ug) wasincubated with 1.0 ,ug of T antigen and4mMATPfor1 h atvarious temperatures (as indicat-ed). The DNA was then modified with KMnO4, and the sites of modification were probed by primer extension as described in MaterialsandMethods. SV40 sequence positions are indicated as described inthelegend to Fig. 2. (B) DNA untwistingmediated by Tantigen.Form IpSV01AEP(0.3 ,ug) wasincubatedwith 0.75 ,ug of T antigen and 4 mM ATPfor 60 min atvarious temperatures (as indicated). TopoisomeraseIwas presentin all reactions except that of lane 1. After incubation, the product DNA was subjected to agarosegelelectrophoresis in the presenceof2
jig
ofchloroquine phosphateperml. Theposition of form II (nicked)DNAis indicated ontheleft. (C) Quantitation oforistructural changes. The levelsof DNAuntwistingand KMnO4oxidationin theearly palindrome and AT tracts weredetermined andnormalizedasdescribedinMaterials andMethods.Symbols:0,early palindrome oxidation;*, AT-tract oxidation; A, DNA untwisting. Maximum DNA untwisting was foundat30°C,atwhichpoint0.9turnwasremoved.opposedtochanges inthedirectionof thehelicalaxis, such
asthebendingofDNA).
Theamount of DNAuntwisting wascalculatedby
densi--A tometric scanning of the gel and quantitation of the
differ-ence between the center of each topological distribution. The fraction ofmaximal DNA untwisting was plotted as a
10 20 3 function of ATP concentration (Fig. 2C). The untwistingof
oribyTantigenwasfoundto haveasigmoidal dependence
Temperature( C) on ATP concentration. DNA untwisting approached a
pla-teau at thehighest levelsofATP, at whichpoint 1.3 helical turns were removed. The level of ATP giving rise to 50% maximaluntwistingwascalculatedtobe140R,M. The DNA tion to relieve any DNA supercoiling untwistingcurve appearedtoparalleltheKMnO4 reactivity nbindingtotheori. Tantigendoesnot ofthe two elements, with a particularly close overlap be-unwinding of the plasmidDNA mole- tween DNAuntwisting and modification of the ATtract. ns,sinceanSSB wasnotadded. After Sensitivityof theearly palindromeand ATtract to
temper-ion mixtures were deproteinized and ature. The response of DNA structural changes to incuba-gel electrophoresis in the presence of tion temperature was examined. T-antigen-ori complexes i visualization of the entire plasmid were formed at various temperatures in the presence of 4
rn(Fig. 2B). mM ATP and then chemically probed with KMnO4. A
ncentration of ATP from 0 to 3 mM representativeautoradiographoftheseparatedprimer
exten-Lon of plasmid DNA topoisomers to sion products is shown in Fig. 3A. The autoradiograph vardsbyoneto twobands (Fig. 2B,cf. indicated that the early palindrome and AT-tract regions theorigin DNAdoesnotwraparound were differentially sensitive to temperature. Surprisingly, somelikeconfiguration (9), theupward the AT tract was modified at lowertemperatures than the
listribution most likely results from T early palindrome element. This response was opposite to .e DNA helix. We will refer to such thatobservedwithATP,when theearly palindromewasthe is DNA untwisting, defined as an in- moresensitiveof the two elements at low concentrations of -r of base pairs per helical turn (as nucleotide.
C
c
0
o
o.
OE_:
--0
N
-*- o
E
z
=cr~
-- -- ---- r,---- r,--
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.320.549.74.231.2]Thelevel of KMnO4 attack in each of the two regions was
quantitated, and the normalized oxidation of each region was
determinedas afunction of temperature (Fig. 3C). The level of KMnO4oxidation in the AT tract increased sharply from 0 to 10°C and then decreased gradually from these peak
levels. The behavior of the early palindrome element with respect to temperature was quite different, verifying the
differential sensitivity of the two ori elements to tempera-ture. The early palindrome was poorly reactive to KMnO4
below20°C but became increasingly reactive to the chemical
probe up to the highest temperatures tested. The tempera-turesthat resulted in 50% maximumKMnO4oxidationwere determined for each of the two regions. The thermal
mid-points
werefound todifferbyover25°C, with the midpoints calculated to be 2 and 28°C for the AT-tract and earlypalindrome elements, respectively.
Parallel experiments were performed to determine the response of DNA untwisting to temperature. T antigen was
incubatedatvarious temperatures withori-containing DNA, 4 mM ATP, and topoisomerase I. The DNAs were then
subjected to agarose gel electrophoresis to visualize their
topoisomer distribution (Fig. 3B). Since the helical twist of DNAis temperature dependent (13), control reactions
lack-ingTantigen were performed at each temperature. Note that the HeLatopoisomerase I was active at the lowest temper-atures tested (i.e., 0°C; Fig. 3B, cf. lanes 1 and 2). The amount of T-antigen-dependent DNA untwisting at each temperature wascalculated and plotted (Fig. 3C). The
T-an-tigen-mediated DNA untwisting was strongly dependent on temperature,risingsharply from the minimal levels observed in the low-temperature range (s100C). DNA untwisting
plateaued at temperatures of 25 to 30°C and then declined
slightlyat37°C.The thermalmidpoint for DNA untwisting in theincreasing portion of the curve was calculated to be 17°C.
Comparison of DNA untwisting with KMnO4 oxidation
indicated thatuntwisting, in general, paralleled the response of the early palindrome elementbut had athermal midpoint thatwasintermediate between those calculated for the early
palindromeand AT-tract elements.
Titration of T antigen. The response of DNA structural
changes to the amount of added T antigen was examined.
Since ori-dependent DNAuntwistinghas been examined in
parallel
studies(9a), weprobed structural changes in ori byKMnO4 modification. ori DNA was incubated with 4 mM ATPandvariousamountsofTantigen at 37°C, and the DNA was then modified with KMnO4. Autoradiography showed that structuralchanges induced in ori were highly dependent on theamount ofTantigen added, a result consistent with those of preliminary studies (Fig. 4A) (3). The level of oxidation in each region was quantified and separately
normalized,and these levels were plotted as a function of the amountofT antigen added (Fig. 4B). DNA untwisting as a
functionofTantigen is also presented (9a).
Structural changes induced in both the early palindrome andAT tractwerefound to have a roughly linear response to addedTantigen. Noobvious plateau was observed in either curve up tothe highest levels ofT antigen tested (2.0 F.g).
DNA untwisting showed a similar profile, though at the lowestT-antigen levels, DNA untwisting was detected with-outsignificantKMnO4modification. The levels of T antigen
causing
50% maximal oxidation or untwisting were deter-mined by using the data obtained with 2.0 ,ug as the maxi-mum.Themidpointswerefound to be at 0.7 and 1.2[Lg
of Tantigen for structural changes in the early palindrome and AT tracts, respectively, and at 0.6 ,ug of T antigen for DNA
untwisting.
A
T antigen (ug) 0.0 0.1 0.25 0.5 0.75 1.0 2.0
5 1 59...
51
63- 5217-
5225-
4-16 ,
29-
30-"a aiN
nwe
_- _NWa
EP
GAGGC
AT
1 2 3 4 5 6 7
B
0
N.
0
X
nr
o
c
Ya
.N
O
m
z
0.0 0.5 1.0 1.5 2.0
Tantigen, uag
FIG. 4. Influence of T-antigen concentration on structural changes inori. (A) Representative autoradiographyoftheKMnO4 oxidation pattern in orias a function of T-antigen levels. Form I pSVO1AEP(0.5p.g)wasincubatedwithTantigen(asindicated) and 4 mM ATP for 1 h at 37°C. The DNA was then modified with KMnO4, and the sites of modification were probed by primer extension as described in MaterialsandMethods. SV40 sequence positions are indicated as described in the legend to Fig. 2. (B) Quantitation oforistructuralchanges.KMnO4oxidationin the early palindrome and AT tracts were determined and normalized as described in MaterialsandMethods.Symbols: *, early palindrome oxidation;*,AT-tractoxidation;- - --, DNA untwisting. The DNA untwistingcurve isfrom Deanand Hurwitz (9a). Maximum DNA untwisting was found at 2.0 ,ug of T antigen, at which point 1.9 helical turns wereremoved.
These responses were somewhat unanticipated because at the highest levels of protein (i.e., 2.0 ,ugof T antigen), the molar ratio of T antigen to DNA was greater than 80 to 1. Assuming that 3 molecules of T antigen can bind to site I (located outside the core ori) and up to 12 monomers can bind coreori-containing site 11 (17, 18), this level of T antigen nominally represents a fivefold excess over that needed to
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.328.557.85.503.2]completely occupy both sites. The linear response to the high T-antigen levels may reflect a low percentage of active molecules in the population. The DNA replication activity of low concentrations of similar T-antigen preparations showed a 2- to 4-fold stimulation when T antigen was treated with cellular protein phosphatase 2A and a 1.5-fold stimulation when T antigen was treated with human cdc2 kinase (cdc2 kinase generously provided by D. Beach) (9b, 19, 24). We note, though, that formation of ATP-dependent hexamers by T antigen in the absence of DNA occurs with more than 95% of the available T antigen in these and similar preparations of T antigen (9a, 17). Thus, a more likely hypothesis is that the core
ori
molecules are not saturated even at high levels of T antigen.Kinetics of formation of structural changes within
on.
The kinetics of induction of the ori structural changes was examined. To detect events occurring within the first few minutes of T-antigen binding toori, we used higher concen-trations ofKMnO4
and decreased the modification time relative to the conditions used above (see Materials and Methods). Individual reaction mixtures were also pre-warmed to37°C
in the absence of T antigen to reduce any effects of thermal equilibration during the initial period of T-antigen binding.T antigen was added to each reaction, and after various periods of incubation, the DNA was probedwith
KMnO4.
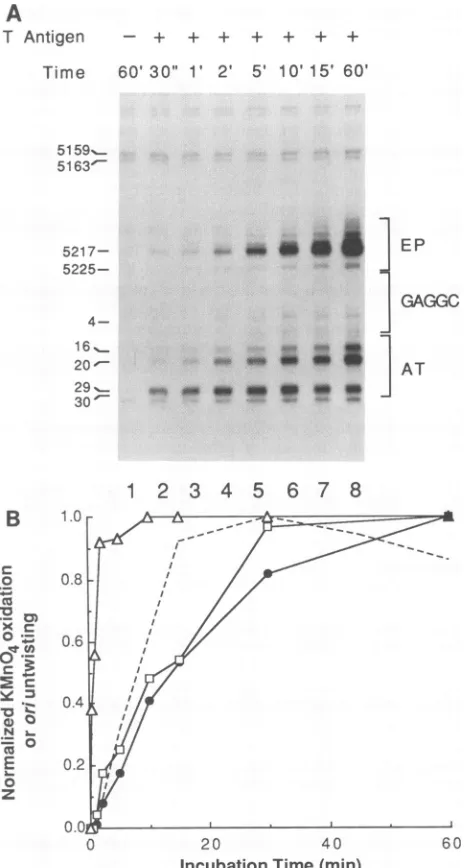
Autoradiography of the primer extension products re-vealed two distinct structural events within the AT tract distinguishable by their dissimilar kinetics of formation (Fig. 5A). The earliest structural transition occurred within the portion of the AT tract distal from the central GAGGC element minimally containing nucleotides located at SV40
positions 29 and 30. Strong KMnO4 modification of this region was found in the first minute of binding, when only minor reactivity of the portion of the AT tract proximal to the GAGGC element (containing nucleotides 16 and 20) was detected. Structural changes in the proximal region of the AT tract as well as in the early palindrome element occurred at a slower rate, increasing gradually over the first 30min of binding. Thus, it appears that separate conformational changes can occur in discrete regions of the AT tract during the first few minutes of T-antigen binding.
Levels of modification of the proximal and distal regions of the AT tract and within the early palindrome element were separately quantified and plotted as a function of incubation time (Fig.
5B).
The time course of DNA untwisting is presented for comparative purposes (9a). Structural changes in the two AT-tract regions clearly differed, with 50% maximal oxidation occurring at 50 s and 11.5 min for the distal and proximal regions, respectively. The modification of the early palindrome element and the proximal AT tract occurred at a similar rate, with50%maximal oxidation of the early palindrome calculated to occur at 13.5min. The kinetic midpoint for DNA untwisting was calculated to be 8 min.DISCUSSION
Conformational changes induced by T antigen in two critical regions ofori DNA, the early palindrome and AT-tract elements, have been demonstrated to be highly and differentially sensitive to reaction conditions. The induction of these structural changes was found to be surprisingly complex. The early palindrome can undergo significant structural transitions at relatively low concentrations of ATP, but these require incubation temperatures above25°C.
The AT tract, on the other hand, can be deformed by T
A
T Antigen - + + + + + +
Time 60'30"
1V
2' 5' 10'15' 60'5159_...
5163'-
5217-
5225- 4-16_
20
-29_...
30
-7
m* a
a *w
ASp, a 0
.0 4. Om o
EP
GAGGC
AT
1
2 3
4
5
6
7 8
B
1.0c 0
:
ns
Xc
0l;,
o ..D
CU)
-0
z
60
20 40
[image:6.612.317.549.75.509.2]Incubation Time
(min)
FIG. 5. Time course of induction of structural changes within ori. (A) Representative autoradiograph of the KMnO4 oxidation pattern in ori as a function ofthelength of incubation ofTantigen with ori.Form IpSVO1AEP(0.5,ug)wasincubatedwith 1.0,ugofT antigen and 4 mMATP for various times (as indicated)at37°C. The DNA was then modified with KMnO4, and the modification sites were probed by primer extension as described in Materials and Methods.SV40 sequence positionsareindicatedasdescribed in the legend to Fig. 2. (B)Quantitationofori structuralchanges. KMnO4 oxidation in the early palindromeand AT-tract regionsproximaland distal to the central GAGGC elementweredetermined and normal-ized as described in Materials and Methods. Symbols: 0, early
palindrome oxidation;O, proximal AT-tract oxidation; A, distal AT-tract oxidation; ----, DNA untwisting. The DNA untwisting curve is from Dean and Hurwitz (9a). Maximum DNA untwisting was found at 30min, at which point 1.0 turn wasremoved.
antigen at low temperatures (5°C), but these conformational changesrequired levels of ATProughly fivefoldhigher than that needed to induce significant changes in the
early-palindrome element. Thus, structural transitions can occur
on November 10, 2019 by guest
http://jvi.asm.org/
in one region even in the absence ofcorresponding changes in the opposite flanking region.
The differential induction ofthe structural transitions in the two flanking ori elements demonstrates that each ele-menthas separateenvironmentalrequirementsforbecoming
distorted.This likely reflectsthat eachelementiscomposed
of different sequence tractsandthat each elementundergoes
distinct conformational changes. The early palindrome ele-ment, containing mainly cytosines and thymines on one of the strands, becomessignificantlymeltedbythe
ATP-depen-dent complex. Structural changes in the AT tract, on the other hand, do not appear to resultfrom DNA melting but rather represent adistortion of double-stranded DNA (3, 21). The sharp temperature dependence ofKMnO4 hypersen-sitivity within the early palindrome element is most likely
due to the need for sufficient thermal input for T antigen to melt this DNA. A similar temperature dependence has been observedformeltingof the E. coliorigin of replication(oriC)
bythe DnaAprotein (5). We note that the thermalprofiles of
oriC melting and early palindrome modification are nearly identical both in shape and temperature range. This pro-nounced thermal dependence is also reminiscent of the
profiles seen for melting ofpromoter elements by E. coli
RNApolymerase (16). The similarities in response to tem-perature in these three diverse systems suggest that such sharp thermal responses may be typical of many protein-mediated DNA melting events such as at other origins of replication or, possibly, within eukaryotic transcription
complexes.
Formation of the ATP-dependent complex rendered the AT tract susceptible to KMnO4 oxidation over a broad temperaturerange,with a thermal profile quite distinct from that of the early palindrome (Fig. 3C). The structural
tran-sitions occurring in the AT tract thus do not appear to require a high thermal input. The actual conformational
change taking placewithin the ATtractisatpresentpoorly understood, though the lack of pronounced thermal depen-dence again indicates that the primary conformational
changeis not DNA melting. We have previously suggested that the DNA becomes untwisted by T antigen (3), and this proposal would be consistent with the observation that T
antigen canuntwist theSV40 ori.
TheT-antigen-mediatedinduction of structural changes in the AT tract was found to require fivefold-higher levels of ATP thanchangesinduced in the early palindrome. Because theTantigen bound to each flanking element contacts DNA of dissimilar sequence, these interactions may induce T
antigen to have different affinities for ATP. An alternative explanation is that distortion of the AT tract requires a
higherenergetic input by T antigen than melting of the early
palindromeelement and that this extra "work" is driven by the presenceof higherconcentrations of ATP. In support of this model, melting of the early palindrome element by T antigen has been shown to take place efficiently in the presence of nonhydrolyzable analogs of ATP, while
com-plete distortionof the AT tract requires ATP hydrolysis (3).
The levels of ori DNA untwisting andKMnO4 oxidation of the early palindrome and AT-tract elements were each linearly dependent on the amount of added T antigen. No plateau was found for any of these three quantities uptothe highest levels of T antigen tested. The higher levels of T antigen (i.e., 2.0 ,ug) nominally represent a fivefold molar excess overthatrequiredtocompletely fill T-antigen sites I and II (littlebinding to site III occurs under these conditions) (2). The linear dependence of ori structural changes on T antigen indicates that the core ori is not saturated with T antigen,evenin the upper rangeof theprotein titration. This conclusion is reinforcedby the observations of Mastrangelo et al. (17), who showed that while a double hexamer of T antigen is the maximal ATP-dependent complex bound to the core ori, themajority of T-antigen-ori complexes found atmoderate levels of Tantigen (i.e., 0.4 ,ug)aresmaller. An increase in the amount ofT antigen might therefore shift moreof the protein into the double-hexamer form, leadingto greater distortion of ori structure. Whether T antigen
con-tained in structures smaller than a double hexamer can
distortthe oristructureis as yetunclear,butquite possible.
Changes in reaction conditions had similar effects on T-antigen-mediated DNA untwisting andonthe induction of structural transitions in the early palindrome and AT-tract
regions. In each ofthe four reaction parameters studied,
DNA untwisting was found generally to parallel or fall intermediate between KMnO4 oxidation of thetwoflanking
ori regions (three regions in the time course ofT-antigen
binding).These results suggest that DNAuntwistingand the localized structural distortions are closely related manifes-tationsof thechangesthatoccurwhenTantigenbindstoori. Themeltingof 8bpin theearly palindrome element and the distortion of the ATtractover aregion of16bpappearstobe
consistent withthe untwisting ofone to two helicalturns. The formation of the T-antigen ATP-dependent complex
with the SV40 ori occurred in at least two discrete steps. In thefirst step, Tantigenboundtooriinducedconformational
changesin the distalportionof the ATtract.Tantigenthen
structurally deformedtheremaining portion of the ATtract and the early palindrome element. The first step likely requiresthepreviousorsimultaneous binding of Tantigento the centralGAGGC repeats.Othershave shown that
distor-tion of the AT tract is not,observed in the absence of the centralGAGGCelement(21). The T-antigen complex there-foreminimally spans the entire AT tract in this initial step. As this step precedes complete distortion of the complete AT tract,distortion ofthedistalportion ofthe AT tract may be a required substep serving to initiate changes in the remainderofthe tract.
Theearlypalindromeelement did not show any evidence
ofmultiple structural transitions duringthe initial few min-utesofbinding.Thekineticprofile ofKMnO4 modification in theearly palindrome instead resembled the curve obtained for theproximalATtract(Fig.5). Structuraldistortion of the distal AT tract does not appear to be a prerequisite for
distortionof theearly palindrome element, since Parsons et
ATP Tantigen V
SV40ori
Disortion of
distalAT-tract Limited DNAuntwisting
Distortion of
proximal AT-tract SSB
o
Melting
of 2. P- DNA unwinding - 0- DNAsynthesisearly palindrome
Greater DNAuntwisting
FIG. 6. Model of events occurring during the initial steps of SV40 DNA replication.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.94.537.658.713.2]al. (21) have shown that T antigen can melt the early palindrome in the absence of the AT tract and, at high concentrationsof T antigen, in DNA constructs lacking both the AT tract and central GAGGC elements. These investi-gators also postulated that early palindrome melting pre-cedes distortion of the AT tract. However, in light of the kinetic experiments presented above, this suggestion may require revision.
These data combined with work by a number of investi-gators (see references 1, 6, and 23 and references therein) lead to an updated model of the events occurring during the initial steps of SV40 DNA replication (Fig. 6). T antigen in the presence of ATP binds to the central GAGGC element of the
SV40
ori. Simultaneously with or soon after this initial binding, T antigen contacts and distorts the distal portion of the AT tract and causes limited untwisting of the helical structure ofori. Conformational changes then occur in the T-antigen-oricomplex that result in distortion of the remain-der of the double-stranded AT tract, melting of the early palindrome, and more-significant untwisting of the DNA. This complex contains two structural lobes which, in the presence of human SSB, separate andbidirectionally unwind the DNA outward from ori. The SSB constrains the single strands, allowing the DNA polymerase machinery entry for catalysis of nascent-strand synthesis.Conversion of DNA into a template suitable for DNA synthesis is seen to be an intricate process. Large-scale denaturation of the DNA helix entails an elaborateseries of protein-protein and protein-DNA interactions. The SV40 model system is remarkable in that these complex reactions are catalyzed predominantly by a single protein, T antigen.
ACKNOWLEDGMENTS
We thank Peter Bullock, Monika Lusky, Warren Jelinek, Daniel Atkinson, and Dhruba Sen Gupta for helpful discussions. We appreciate the technical assistance provided by Nilda Belgado and Barbara Phillips and the preparation of artwork by David Valentin. This work was supported by Public Health Service grantSRO1 GM34559 from the National Institutes of Health. J.B. was supported by a postdoctoral fellowship from the National Institutes of Health.
REFERENCES
1. Borowiec, J. A., F. B. Dean, P. A. Bullock, and J. Hurwitz. 1990. Binding and unwinding-how T antigen engages theSV40 origin of DNA replication. Cell60:181-184.
2. Borowiec, J. A., and J. Hurwitz. 1988. ATP stimulates the binding of simian virus 40 (SV40) large tumor antigen to the SV40 origin of replication. Proc. Natl. Acad. Sci. USA 85:64-68.
3. Borowiec, J. A., and J. Hurwitz. 1988. Localized melting and structural changes in the SV40 origin of DNA replication induced by T antigen. EMBO J. 7:3149-3158.
4. Borowiec, J. A., L.Zhang,S. Sasse-Dwight, and J.Gralla.1987. DNA supercoiling promotes formation of a bent repression loop in lac DNA. J. Mol. Biol. 196:101-111.
5. Bramhill, D., and A. Kornberg. 1988. Duplex opening bydnaA protein at novel sequences in initiation of replication at the origin of the E. coli chromosome. Cell 52:743-755.
6. Challberg, M., and T. Kelly. 1989. Animal virus DNA replica-tion. Annu. Rev. Biochem. 58:671-717.
7. Dean, F., J.
Borowiec,
Y. Ishimi, S. Deb, P. Tegtmeyer, andJ. Hurwitz. 1987. Simian virus 40 large tumor antigen requires three core replication origin domains for DNA unwinding and replication in vitro. Proc. Natl. Acad. Sci. USA84:8267-8271. 8. Dean, F., P. Bullock, Y. Murakami, C. R. Wobbe, L. Weissbach, and J. Hurwitz. 1987. Simian virus 40(SV40) DNA replication:SV40large Tantigen unwindsDNAcontainingthe SV40origin ofreplication. Proc.Natl. Acad. Sci. USA 84:16-20.
9. Dean, F. B., M. Dodson, H. Echols, and J. Hurwitz. 1987. ATP-dependent formation ofaspecializednucleoprotein
struc-ture bysimianvirus 40(SV40)largetumorantigenatthe SV40 replicationorigin. Proc.Natl. Acad. Sci. USA 84:8981-8985. 9a.Dean, F. B., and J. Hurwitz. J. Biol. Chem., inpress. 9b.Dean, F. B., and J. Hurwitz.Unpublished data.
10. Deb, S., A. L. DeLucia, C. Baur, A. Koff, and P. Tegtmeyer. 1986. Domain structure of the simian virus 40core origin of replication. Mol. Cell. Biol. 6:1663-1670.
11. Deb, S., and P. Tegtmeyer. 1987. ATPenhancesthebindingof simian virus 40 large Tantigen tothe origin ofreplication. J. Virol.61:3649-3654.
12. Deb, S., S. Tsui, A. Koff, A. DeLucia, R. Parsons, and P.
Tegtmeyer. 1987. The T-antigen-bindingdomain of the simian virus 40core origin ofreplication.J. Virol. 61:2143-2149. 13. Depew, R. E., and J. C. Wang. 1975. Conformational
fluctua-tions ofDNAhelix. Proc.Natl. Acad.Sci. USA 72:4275-4279. 14. Dodson, M., F. Dean, P. Bullock, H. Echols, andJ. Hurwitz. 1987. Unwinding of duplex DNA from the SV40
origin
of replication by Tantigen. Science238:964-967.15. Kenny, M., S.-H. Lee, andJ.Hurwitz. 1989.Multiplefunctions ofhumansingle-stranded-DNAbindingprotein insimian virus 40 DNAreplication: single-strand stabilization and stimulation ofDNA polymerases a and B. Proc. Natl. Acad. Sci. USA 86:9757-9761.
16. Kirkegaard, K., H. Buc, A. Spassky, and J. C. Wang. 1983. Mapping of single-stranded regions in duplex DNA at the sequence level: single-strand-specific cytosine methylation in RNApolymerase-promoter complexes. Proc. Natl. Acad. Sci. USA80:2544-2548.
17. Mastrangelo, I.A., P. V. C.Hough,J.S.Wall,M.Dodson,F.B.
Dean,andJ. Hurwitz. 1989. ATP-dependentassemblyof double hexamersofSV40 Tantigen atthe viraloriginofDNA
replica-tion. Nature(London)338:658-662.18. Mastrangelo, I. A., P. V. C. Hough, V. Wilson, J. Wall, J. Hainfeld, and P.Tegtmeyer. 1985. Monomersthroughtrimersof large tumor antigen bind in region I and monomers
through
tetramersbindinregionIIof simian virus40originof
replication
DNA as stable structures in solution. Proc. Natl. Acad. Sci. USA 82:3626-3630.
19. McVey, D., L.Brizuela, I. Mohr,D. R. Marshak,Y.Gluzman,
and D. Beach. 1989.Phosphorylationoflargetumour
antigen by
cdc2stimulates SV40DNAreplication. Nature (London) 341: 503-507.
20. O'Reilly, D. R., andL.K.Miller. 1988.Expressionand
complex
formation of simianvirus 40large Tantigenand mouse
p53
in insectcells. J.Virol. 62:3109-3119.21. Parsons, R., M. E. Anderson, and P. Tegtmeyer. 1990. Three domains in the simian virus 40 core origin orchestrate the binding, melting, and DNA helicase activities ofT
antigen.
J. Virol. 64:509-518.22. Roberts, J. 1989. Simian virus 40 (SV40) large tumor
antigen
causes stepwise changes in SV40origin structure
during
initia-tion of DNAreplication. Proc.Natl. Acad. Sci. USA 86:3939-3943.23. Stillmann, B.1989. Initiation ofeukaryoticDNA
replication
in vitro. Annu. Rev.Cell Biol. 5:197-245.24. Virshup, D.M.,M.G.Kauffman,andT.Kelly.1989. Activation ofSV40 DNA replication in vitro
by
cellularprotein
phos-phatase 2A. EMBOJ. 8:3891-3898.25. Wobbe, C. R.,F.Dean,L.Weissbach, andJ.Hurwitz. 1985.In
vitroreplication ofduplexcircular DNA
containing
thesimian virus 40DNAorigin site. Proc. Natl. Acad. Sci. USA 82:5710-5714.26. Wold, M.,J.Li,andT.Kelly.1987.Initiationofsimianvirus 40 DNA replication in vitro:
large-tumor-antigen-
andorigin-de-pendentunwindingof thetemplate. Proc.Natl.Acad. Sci. USA 84:3643-3647.