JOURNAL OFVIROLOGY, Apr.1975, p. 1042-1046 Copyright i 1975 AmericanSocietyforMicrobiology
Vol.15,No.4 Printed in U.S.A.
Purification and Properties
of
a
Bacteriophage
T5-Modified
Form
of
Escherichia
coli RNA
Polymerase
C. SZABO1 AND R. W. MOYER*
DepartmentofBiochemistry, College of PhysiciansandSurgeons,Columbia University, NewYork, New York 10032
Received forpublication 25 November 1974
Amodified form of Escherichia coli RNApolymerase that containsoneofthe
three T5-specific polypeptides known to interact with the host enzyme was
purified from bacteriophage T5-infected cells. The properties of this
T5-modi-fied enzyme appeared identical to those of the RNA polymerase derived from
uninfected non-colicinogenic cells and toafullyactiveenzymeisolated from
T5-infected ColIb+ cells after the limited in vivotranscription of T5 genesallowed
by the plasmid had ceased.
The RNA polymerase ofthehost cell Esche-richia coli isresponsibleforthesynthesisof the
three distinct classes of bacteriophage
T5-specific RNA formed during T5 infection. Byimmunoprecipitation studieswehaveshown
thataclassI(pre-early)T5-specific polypeptide
of 11,000 daltons and two classII (early)
poly-peptides of 90,000 and 15,000
daltons,
respec-tively, become associated with theRNA
polym-erase during the infectious cycle (C.
Szabo,
B.Dharmgrongartama, and R. W.
Moyer,
Bio-chemistry, inpress). Wehavepresented datato
suggest that at least one of the three
polypep-tides, the 90,000-dalton species, tentatively
re-ferred to as aT5-specific "a'factor, is involved
in the in vivo regulation of class III (late)
transcription and is likely the product of gene
C2.Ifthehost E. colicell contains the
colicino-genic factor ColIb (ColIb+ cells), T5-specific
classIRNAis formednormally.
However,
afterthe initiation of class IIphage RNA synthesis,
transcription is arrested and the infection aborts. As expected, infectedcolicinogenic cells show
only
the pre-early T5peptide
of 11,000 daltons associated with the polymerase (C. Szaboetal.,inpress).We have purified RNA polymerase from
T6-infected non-colicinogenic cells 20 min after
infection, using the standard procedures as
described by Burgess and Travers (2). Before
purification, the infected cultures had been
continuously labeled with [3H
Ileucine
underconditions that label all three classes of T5
proteins, including those we find bound to the
polymerase. After each step ofpurification, a
portion of the partially purified enzyme was
'Present address: Lawrence Berkeley Laboratory, Uni-versityofCalifornia,Berkeley, Calif. 94720.
immunoprecipitated with antisera prepared
against theuninfected E. coliRNApolymerase holoenzyme, and the presence of the three
phage-specific proteins in the
immunoprecipi-tate was quantitated by analysis on sodium
dodecylsufate (SDS)-polyacrylamide gels. The
first step of the polymerase purification is the
precipitation of the enzyme by ammonium
sulfate such as we routinely used in our earlier
immunoprecipitation experiments (Szabo et
al., in press) and which, we have observed,
has no effect on the number or amount of the
phage polypeptides bound to the polymerase
(datanotshown). The RNApolymerase precipi-tated in the ammonium sulfate step is then
purified by DEAE-cellulose chromatography,
which removes all but the 15,000-daltonclass II
polypeptide from the enzyme (Fig. 1A). This remaining T5 polypeptide was rather strongly associated with the polymerase and remained
bound through a subsequent purification step
utilizing glycerol gradient centrifugation (Fig.
iB). After theglycerol gradient step of
purifica-tion, ourpreparationsofRNApolymerasewere
greater than 95'% homogeneous and contained
anequivalent amount ofthe 15,000-dalton
sub-unit per mol of enzyme as was originally
present in the immunoprecipitates prepared
from crudeextracts. We will refer to this
puri-fied form of RNA polymerase that contains a
single T5 polypeptide of 15,000daltonsisolated from infected non-colicinogenic cells as
RNAP(T5)(15), in agreement with our
sug-gestednomenclature (Szaboetal., in press).
Asample of theRNAP(T5)(15)from cells was
then analyzed on an SDS-polyacrylamide gel
and the total subunit pattern was compared
with that of a preparation ofRNA polymerase
purified from uninfectedcells. Coomassie
blue-1042
on November 10, 2019 by guest
http://jvi.asm.org/
VOL.15, 1975N
10 20 30 40 50 60 70
Slice number
FIG. 1. SDS-polyacrylamide gel electrophoresisof
'H-labeled T5-specific proteins bound to purified
RNA polymerase. A 100-mlsample ofunstarved E.
coliK-12W3110thy cells in T-MGM medium(6)was
infectedwith T5st(0), aheat-stabledeletionmutant
of T5+, at a multiplicity ofinfection of10.Leucine
wasthenaddedto afinal concentrationof0.1
,g/ml,
and a total of500
MlCi
of [4,5-3HJleucine (50 mCi!Mumol) (New England Nuclear Corp.) was added in
aliquots to the infected culture at 1-min intervals
from 5 to 14minafterinfection.At15min, the [3H]-leucinewas chased with 0.2 mgofunlabeledleucine
perml,andafter20minofinfectionchloramphenicol
(0.1 mg/ml final) was added and the culture was
harvested by centrifugation. The 3H-labeled cells
were added to8gof identicallygrownand infected
unlabeled carriercells,and theRNApolymerasewas
purified bytheprocedure of Burgessand Travers(2)
and assayed aspreviouslydescribed (1). Aftereach
stepofpurification, aportion ofthe RNApolymerase
was immunoprecipitatedas described (Szabo etal.,
in press). The 'H-labeled, precipitated samples
wereanalyzedbySDS-polyacrylamide gel
electropho-stained polypeptide bands with mobilities
cor-respondingtothe ,B, ,B', a, and a subunits of the
hostenzyme were detected in bothpreparations
(Fig. 2). Since we have shown that all of the
radioactivity associated with the phage
poly-peptide of90,000 daltons was lost during this
method of purification (Fig. 1), the stained
polypeptide of 90,000 to95,000 daltons present in the RNA polymerase purified from infected cellswasprobably thehost a factor,whichhas a molecular weight of 95,000. From these results it
appearsthat
T5
infection does notpermanentlydestroy the host a protein, and furthermore neither the association of the 15,000-dalton class II phage polypeptide nor any other
T5
modification permanently prevents the host afactor from associating rather stronglywith the
enzyme.It is notknownatthistime whether the
binding of the
T5-specific
"a" factor and the host a factor are mutually exclusive.We compared the in vitro transcriptional propertiesof
RNAP(T5)(15)
with those of threeother purified enzyme preparations: (i) RNA
polymerase from uninfected non-colicinogenic cells; (ii) RNA polymerase from uninfected
ColIb+
cells; and (iii)RNApolymerase obtainedfrom T6-infected ColIb+ cells that had ceased
all invivo transcription. The properties ofthe RNA polymerase purified from T5-infected ColIb+ cells seemed particularly pertinent for
this study. Althoughno
ColIb
factorprotein has been found associated with RNA polymerase(Szabo
et al., in press), it is still quitecon-ceivable that the plasmid might antogonize
T5
transcription as aresultof acatalytic
modifica-tionofthepolymerase, which might be detected by an alteration inthe in vitro transcriptional
properties of the enzyme.The RNApolymerase
derived from each of the four preparations chromatographedonDEAE-celluloseas asingle peak that eluted at 0.19 M KCI and after low-salt glycerol gradient centrifugationwas at
least
95%
homogeneous. Fully active enzymes were recovered from all preparations including the one from infectedColIb+
cells. All the purified preparations, when analyzed onSDS-polyacrylamide
gels, appearedtohavethe host ar subunit (Fig. 2;unpublished data).
Asum-mary of the enzymatic activities of the four
resis as described by Weber and Osborn (10). The dataarepresented as 'H countsper minute pergel slice where theAbh and Ab, symbols indicate the positions in the gel of the stained immunoglobulin heavy (55,000 daltons) and light chains (25,000 dal-tons),respectively.(A)DEAE-cellulose-purifiedRNA polymerase; (B) low-salt glycerol gradient-purified RNApolymerase.
1043
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.507.49.239.53.446.2]NOTES
SDS-polyacrylamide gel analysis ofpurified
RNA polymerase from late T5-infected
non-colicinogenic cells suggested that the enzyme
still contained the host a factor (Fig. 2A).
Although the a subunit of RNA polymerase is
required for the efficient transcription of T4
DNA, transcription
proceeded quite
effectively
with calf thymusDNA in thecompleteabsence
of the a subunit. Thus, the ratio of template
activities with T4 and calf thymus DNA has
been defined as a measure of functional a
subunit activity (8). This ratio with enzyme
containing the a subunit derived from
unin-fected cellswas approximately 2, and theratio
did not appear to change either as a result of
infection with T5orfromanyeffect of the ColIb
plasmid (Table I).These datamaybecompared
with those obtained for the RNA polymerase
lacking theasubunitthatwasisolated after T4
infection and had an activityratio with T4 and
calf thymus DNA of 0.2. Our results suggest
[image:3.507.70.250.78.473.2]that each of these preparations contains a
FIG. 2. SDS-polyacrylamide gel electrophoresis of
RNApolymerase purified throughalow-saltglycerol
gradient. The preparations were electrophoresed on
7.5%SDS-polyacrylamide gels for8 hat8mApergel.
After electrophoresis the protein bands were
visual-ized with Coomassie blue. (A) RNA polymerase
purified from uninfected non-colicinogenic E. coli
K-12 W31IOthy-; (B) RNA polymerase purified from
T5-infectednon-colicinogenic E. coli K-12 W31IOthy
cells after20min ofinfection.
purified enzyme preparations, utilizing various
templates, is presented in Table 1. All four
preparations yielded approximately the same
total numberofenzymeunits (data notshown) and had similar specificactivities with agiven
DNAtemplate. T5 DNA was foundtogive the
maximum in vitroactivity.
TABLE 1. Template activitiesof RNApolymerase purified from uninfected and T5-infected cellsa
Spact(U/mgwith
variousDNA Ratio templates
Enzymesource T4/
T5b T4 Calf T5/ calf
T5b T4c thy-dT4 ty
musd ty
mus
Uninfected E. coliK-12
W3110thy. ... 1,215 987 475 1.2 2.1 T5-infected E. coliK-12
W31l0thy. ... 965 749 361 1.3 2.1 Uninfected E. coliK-12
W3110thy- (ColIb) 995 723 351 1.3 2.1 T5-infectedE.coli K-12
W3110thy- (ColIb) 1,184 1,045 514 1.1 2.0
aRNApolymerasefromeachsource waspurified through thelow-salt glycerol gradient step as describedby Burgess and Travers(2)togreaterthan95%homogeneity.Theassay mix contained (in 0.25 ml) Tris-hydrochloride (0.04 M), MgCl2(0.01 M), EDTA(0.1 mM), dithiothreitol (0.1 mM), KCl(0.15M), potassiumphosphate(0.4mM), bovineserum albumin (0.5 mg/ml), undegraded DNA, UTP, GTP, CTP, and ATP (0.1 mM each) (P-L Biochemicals), and RNA polymerase(2 to 54gper assay) at a final pHof7.9.Each assay also contained 2 MCi of [8-1C 1ATP (New England Nuclear Corp.). After incubation for 10 min at 37 C, the reaction was terminated bythe addition of 2.5 ml of cold trichloroaceticacid(5%, wt/vol)containingsodium pyrophos-phate(0.1mM).The samples were allowedtostand on ice for 15minand the precipitate was collected onglassfiber filters (Schleicherand Schuellno. 29), washed with a solution of trichloroacetic acid (2%, wt/vol) that contained sodium
pyrophosphate (1.01M), dried, and counted. h 10MgofundegradedT5 DNAwasusedperassay.
10MgofundegradedT4 DNA was usedperassay. d 25 ug ofcalfthymusDNAwas used per assay.
1044 J. VIROL.
.A..
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.507.271.463.330.477.2]VOL. 15, 1975
functionalhostfactor.Additionalevidence
sup-porting this hypothesis is the effect ofKCI on
the in vitro enzyme activity. Host core
polym-erase activity (lacking the a subunit) is only
negligibly affected by KCI, whereas the
holo-enzyme activity is stimulated by KCl up to a
concentration of 0.2 M (9). All of our
prepara-tions of RNA polymerase required 0.2 M KCI
formaximum activity.
Pispa and Buchanan (7) have studied the
specificity of the in vitro transcription of T5
DNA by host RNA polymerase purified from
uninfected cells. They found that the host a
enzyme transcribed primarily class I and class
IIsequencesand onlyasmall amount of classIII
sequences. We have shown that the net
en-zymatic activity of purified RNA polymerase
doesnotappear tobe altered either by the ColIb
plasmid or by T5 infection, However, RNA polymerase from late T5-infected cells, under
theproperconditions, mayrecognizeand
tran-scribe predominantly
late T5 gene sequences,which are not efficiently read by the enzyme fromuninfectedcells. Likewise, RNA
polymer-ase purified from T5-infected ColIb+ cells
pos-sessesnormal net activity, althoughthe
possi-bilityexiststhatthis enzyme may have lost the
abilitytotranscribeclassII T5 genesequences.
To test thesepossibilities, thespecificityofthe
in vitro transcription of native T5 DNA by all
four of the RNA polymerase preparations we
havedescribedwasexamined by hybridization-competition analysis.
3H-labeled T5 RNAthat was synthesized in
vitro washybridizedto
T5
DNA in thepresenceofunlabeled T5 RNA
synthesized
in vivo. The competitionofthelabeledin vitroRNA with anexcess of the in vivo RNA isolated at various
timesafterinfectionmeasuresthe proportionof
each class of RNA synthesized in vitro. The
specificity of the in vitro transcription is thus determined,sinceeach temporalclass of in vivo
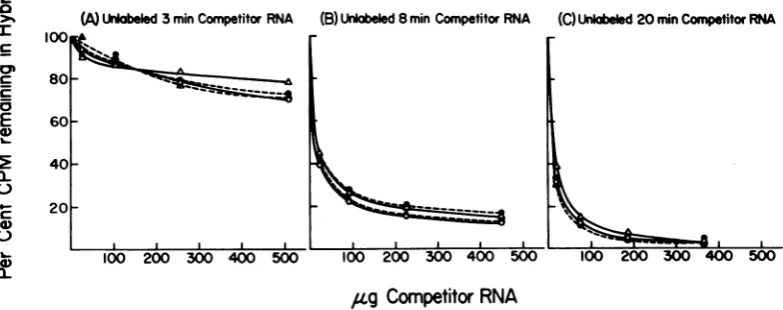
T5 RNA istranscribedfrom aunique set of T5 gene sequences. Figure 3 illustrates the results ofthehybridization-competition analyses, and the data show that all four of the enzyme
preparations described above
transcribed
T5 DNA with the same specificity. Unlabeled(A)Urkabeled3minCompetitorRNA (B) Unlabeled 8minCompetitor RNA (C)Unlabeled20min CompetitorRNA
100 200 300 400 500 100 200
300
400 500 100 200 300 400 500 [image:4.507.54.446.340.495.2]/.g
Competitor RNA
FIG. 3. Hybridization-competition analysis ofT5 RNA synthesized in vitro by differentpreparations of purifiedRNApolymerase. Labeled in vitro RNA and unlabeledinvivoRNA werepurifiedby the hotphenol
method described by Lembach and Buchanan (4). Unlabeled competitor preparations were isolated from
infectedcellsat3, 8,and 20min afterinfection. [3HJRNA wassynthesizedinvitro byaslight modification of
the standard RNApolymeraseassaysystem of Burgess (1). Each mixture contained Tris-hydrochloride (0.04
M),MgCl2 (0.01 M),EDTA(0.1mM), dithiothreitol(0.1mMCalbiochem),KCI(0.2 M),potassium phosphate (0.4 mM),bovineserumalbumin(0.5 Mg/ml),nativeundegradedT5 DNA (20 ug), UTP, GTP, CTP,and ATP (0.1 mMeach) (P-L Biochemicals), and RNA polymerase(10 Mg) inafinalvolume of0.5ml. TheRNA was
labeled by the addition of [5,6-3H]UTP (New England Nuclear Corp.) at a final specific activity of 125 mCi/mmol. The reaction was carried out at 37 C for 15 min before isolation of the RNA. [3H]RNA was synthesized in vitro by RNA polymerase isolated from either uninfected non-colicinogenic E. coli K-12 W3110thy- (0), or colicinogenic E. coli K-12 W3110thy- (ColIb+) (A) and T5-infected non-colicinogenic E. coli K-12 W3110thy- (0) orcolicinogenic E. coli K-12 W3110thy- (ColIb+) cells (A). The labeled RNA
washybridizedtoT5DNA that had been adsorbedto membranefilters(MilliporeCorp.) (0.3 MgofDNAper filter) asdescribedbyCooper(3).Eachhybridizationmixture contained['HJRNA (10,000counts/min),0.3 Mg ofT5DNA,andthe indicatedamountofunlabeled T5 in vivo competitorRNA isolated at: (A) 3min; (B) 8
min;and(C) 20minafterinfection. 100
80 .0 I
c
C. c
.a
0.
0
C
L-c
0-0) 0~
60F
40F
20
I - . .
.--I --- -- -I ---I - - -I .-- -- -2 lr%lf%IIfA^^ Am^^I I aON^ 129%^ Ar%^ wr%^Il
1045 NOTES
I
I
on November 10, 2019 by guest
http://jvi.asm.org/
NOTES
phage RNA isolated 3 min after infection
con-sisted ofonly class I species andwas partially
competitive with the [3H]RNA synthesized in
vitro, indicatingthat at least 25% of the in vitro
RNA in each case wascomposedofclass I gene
sequences (Fig. 3A). Phage RNA isolated 8 min
after infection consists ofboth class I and class
II genesequences (5). Whenthis RNAwasused
as the competitor, only 20% of the in vitro
[3H]RNA remained in
hybrid
form(Fig.
3). Therefore, the greatest proportion of the RNAsynthesized in vitrowith all four enzyme
prepa-rations consisted of early sequences. All three
classesofphage RNA werepresent inthe RNA
isolated 20 min after infection.Although classI
RNA wasnotsynthesizedfrom8 to20 minafter
infection, hybridizablesequences ofclass I RNA
were present inthe cells, eventhoughthis class
ofRNA was no longer metabolically active. All
the [3H ]RNA was displacedfromthe hybrid by
20-min competitor RNA, indicating that all
RNA species synthesized in vitroare also
pres-ent in infected cells. All four enzyme
prepara-tions transcribed class I and class II sequences
more efficiently than class III sequences, since
onlyabout 105% of the in vitro RNA consisted of class III sequences.
The similar catalytic properties of these four
preparations fail to suggest any independent
role for the T5 polypeptide of 15,000 daltons
that is found associated only with the RNA
polymerase purified from infected
non-colicino-genic cells.Furthermore, the isolation of a fully
active preparation from infected ColIb+ cells
with properties similar to those for control preparations suggests that either any
plasmid-directed inactivation of RNA polymerase
activ-ity toward T5 DNA is readily reversible or that
the inactivation of transcription may be the
indirect consequence of some other
plasmid-directed modification ofcellularproperties.
(This work is taken in part from a thesis
submitted
by C.S. to the Faculty of PureScience;
ColumbiaUniverstiy, inpartialfulfill-mentofthe
requirements
for thePh.D.degree.)
ThisinvestigationwassupportedbyPublicHealth Service grant AI-10302from the National Institute ofAllergy and Infections Diseases and grant VC- 142 from the American CancerSociety.C.S.wassupported byPublicHealth Service training grant GM00255 from the National Institute of General Medical SciencestotheDepartmentof Biochemis-try. R.W.M. is supported by Public Health Servicecareer
developmentawardGM34354 from the National Institute of General Medical Sciences.
We acknowledge, with thanks, the gift of antisera
pre-pared against RNA polymerase noloenzyme from E. coli BprovidedbyP. R.Srinivasan and B.Dharmgrongartama.
LITERATURE CITED
1. Burgess, R. R. 1969. Anewmethod for the largescale
purificationofEscherichia colideoxyribonucleic acid-dependentribonucleic acidpolymerase.J. Biol. Chem. 244:6160-6167.
2. Burgess,R.R.,andA. A.Travers.1971.DNAdependent RNA polymerases (EC2.7.7.6), p. 851-853. In G. L. Cantoni and D. R. Davies(ed.), proceduresin nucleic acid research, vol. 2. Harper and Row Publishers,
NewYork.
3. Cooper, T. G. 1972. Procedures for uniform binding of DNA to nitrocellulose filters. Anal. Biochem. 47:481-486.
4. Lembach, K.J., andJ.M.Buchanan. 1969. The relation-shipofproteinsynthesistoearly transcriptioneventsin
bacteriophageT4-infected Escherichia coli B. J. Biol. Chem.245:1575-1587.
5. Moyer, R. W., and J. M. Buchanan. 1969.Patternsof RNAsynthesisinT5-infected cells. Proc. Natl. Acad. Sci.U.S.A.64:1249-1256.
6. Moyer, R. W., and J. M. Buchanan. 1970. Effect of calcium ions on synthesis of T5-specific proteins. J. Biol.Chem.245:5897-5903.
7. Pispa, J. P., and J. Buchanan.1971.Synthesisof
bacte-riophage T5-specific RNA in vitro.Biochim. Biophys.
Acta247:181-184.
8. Schachner, M., W. Seifert, and W. Zillig. 1971. A correlation of changes in host and T4 bacteriophage
specificRNAsynthesiswithchangesofDNA depend-entRNApolymerasesinEscherichia coliinfectedwith bacteriophage T4. Eur.J. Biochem. 22:520-528. 9. Stevens, A. 1974. Deoxynucleic acid dependent
ribonu-cleic acid polymerase from two T4 phage-infected
systems.Biochemistry 13:493-502.
10. Weber, K., and M. Osborn. 1969. The reliability of molecular weight determinations by dodecyl sulfate-polyacrylamide gel electrophoresis. J. Biol. Chem. 244:4406-4412.
1046 J.VIROL.