0022-538X/78/0027-0738$02.00/0
Copyright©1978 AmericanSociety for Microbiology Printed in U.S.A.
Evolution of
Bacteriophage
45X174
IV. Restriction Enzyme Cleavage
Map
of
St-1
J. N. GRINDLEYt* ANDG. N. GODSON
Radiobiology Laboratories, Yale MedicalSchool,NewHaven,Connecticut06510
Received for publication7May1977
The St-1 genome is about 6,050 base pairs in size, approximately 10% larger than 4X174 (5,375 base pairs). The DNA fragmentsobtained by HincII, HaeIII, and EcoRIdigestionwereordered andaligned intoa
colinear
map,and thesingle BglI cleavage sitewaslocated.St-1 is a
small,
icosahedral, single-stranded
DNA-containingbacteriophage whichwasfirst isolated by Bradley (4) and was further charac-terized by Bowes and Dowell (3). It infects K-12
strains of
Escherichia coli and is serologically
unrelated to 4X174
(4,
22).
Preliminary experiments
inthis
laboratory
have indicated that St-1 codes for proteins sim-ilar in number and size to those directed by
4X174, although
theSt-1products presumed
tobe
equivalent
totheOX
geneA, F,
andGprod-ucts are
larger.
However, St-1 has much simpler
requirements for host proteins
involved in DNA
replication than 4X174. The in vitro conversion
ofSt-1 viral
single-stranded
DNA tothe RFII
(open circular,
double-strandedform)
requiresonly the dnaG and dnaE
geneproducts
plus
DNA
unwinding protein and
elongation factors
I
and
11(25). Inthis,
St-1
resembles G4
(18, 25).In
vivo, however, both
St-1 Rf replication and
viral strand synthesis
can takeplace
at thenon-permissive
temperature inhosts carrying
tem-perature-sensitive lesions
inthe
dnaB ordnaC/D
genes(2;
G. N. Godson and J. N.
Grindley, unpublished
data).
Both
G4 and
OX174
areunable
toreplicate under such
con-ditions (7, 12,
14,23,
26).
As a
preliminary
toinvestigating further the
differences
amongbacteriophages
4X174,
G4,
and
St-1,
wehave constructed
acleavage
map ofSt-1 DNA,using
the restrictionendonucleases
from
Haemophilus influenzae
(HincII),
whichis an
isoschizomer
ofHindII,
fromHaemophilus
aegyptius
(HaeIII),and that
specified
by the Rfactor RI
(EcoRI).
We have alsolocated
thesingle
cutproduced by
theenzyme from Bacillusglobiggi (BglI).
MATERIALS AND METHODS
Phageandbacterial stocks.Bacteriophage St-1
wasobtained from C. E.
Dowell.
The Escherichia colitPresent address:DepartmentofBiochemistry,School of Medicine, UniversityofPittsburgh,Pittsburgh,PA 15261.
K-12 host strainW3110wasobtained from K. Brooks Low.
Preparation of32P-labeled RFI DNA. W3110 was grownat40°C in TPG aminoacid, low phosphate-containing medium (19)toabout4 x
10'
cells per ml andinfected withSt-1at amultiplicity of infection of3. After2to 3min, 30,ugofchloramphenicol per ml was added toinhibitsingle-strand DNA synthesis. The cells were labeled with10
,uCi
of[32P]phosphate (New EnglandNuclear)perml10minafterphageinfection andharvested65minlater.Thecellsweresuspended in 10% sucrose (wt/vol) with 50 mMTris-hydrochloride (pH 8.0), treated with lysozyme in the presence of EDTA, and lysed with Sarkosyl in0.2MNaClasdescribed by Godson (10). Theclear viscouscell lysatewascentrifuged at 80,000
xgfor 45 min topelletmost of the host chromosomal DNA. The supernatant was treated with 25 yg of RNaseperml andphenol extracted and the DNAwas
precipitated with ethanol.St-lRFwasfurtherpurified on 5 to20% neutralsucrosegradients.
Preparation of restriction endonucleases.
HincllwasisolatedasdescribedbySmith and Wilcox (20) fromcells obtained from New England Biolabs. HaeIII and Bglwereprepared in thislaboratory by the methods ofSmith and Wilcox (20).EcoRIwas a
gift of W. Summers.
Enzymedigestion. Preparationofpartial and
ter-minal digestion products wasessentiallyasdescribed by Godson (9).Enzyme digests were analyzed on 3 to 5%acrylamide gelsby the buffer system of Maniatis (15) or on 0.7 to 2% agarose gels as described by Sugdenetal.(21).
DNA fragments were extracted from the gel by macerating the gel slice and soaking it in 0.2 M
NaCl-10mM Tris (pH 7.4). The acrylamidewas
re-movedby passage of the solutionthrough glass wool. SolutionscontainingDNA wereextracted withphenol, and the DNAwas precipitated with ethanol in the presence of tRNA as carrier where necessary. The DNAwasresuspended in enzyme digestion buffer for subsequent digestions.
Nomenclature. For
4X174,
the nomenclature ofEdgelletal. (8) for HincIIproducts and Middletonet
al.(16) for HaeIIIproductswasused. The limitdigest fragments of
St-i
were designated Hinclll to -9, HaeIII1to-12,andasindicatedintheResults section for theproducts ofotherenzymes.738
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 27, 1978
RESULTS
Size
and order of
St-1
fragments
pro-duced by the H.
influenzae (HincH)
restric-tion
enzyme.The
HinclI
enzyme cutSt-i
DNAinto nine
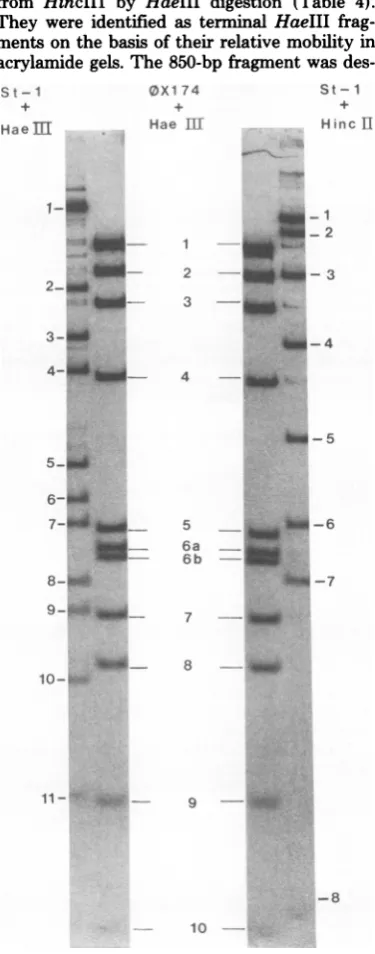
fragments, although only eight bands
were visible in the autoradiograph (Fig. 1) be-cause
the
smallest fragment, HincII9,
was notretained
onthe
gel. Most of the
fragment sizes
shown in
Table
1 wereobtained
by comparing
their
mobility
with that of the DNA
fragments
generated by
HaeIII and HincII from
4X174,
whose sizes
arewell documented
(13, 16, 17).However,
because the
largest
OX174
fragment
isonly 1,200 base pairs
(bp)
and because the
rela-tionship between
log molecular weight and
mo-bility
on4%
polyacrylamide
gels
ceases tobe
astrictly linear function for fragments
greaterthan
1,000bp
(9),the
molecular
weights of suchfragments
arebased
where possible
onthe sumof
the sizes of
fragments produced from the large
fragment by another
enzyme. We wereunable
to
determine the
exactmolecular
weight of
HincII3 because it
was notsubcut
by
the other
enzymes
used. With this
reservation, the total
nucleotide length of St-1 RF from addition of
the
HincII fragment sizes
wasabout
6,120nu-cleotides.
Incomplete digestion of
St-1 with HincII
gave10
partial fragments whose terminal digestion
products
arelisted in Table
2.From
these data
the order
1 2 -9-5 6 4 7 8 3for
theHincIl
ter-minal
fragments
wasestablished.
Size and order of
St-1
fragments
pro-duced
by the H.
aegyptius
(Haell
restric-tion
enzyme.St-1 RF
wascleaved
by
HaeIII
into
12fragments whose nucleotide
lengths,
in-cluding that of
HaeIII12,
which is
notvisible in
Fig.
1, gave atotal of
about 6,010nucleotides
(Table 1). The terminal digestion products
(Ta-ble 3) obtained from overlapping partial
frag-ments
indicated that the
order of the HaeIII
fragments
was 1 3 4 8 10, 5 9 2 7 6 11.No
partial
which
overlapped
both
10and
5 wasisolated, but these
werededuced
tobe
contig-uous
from data described
below. The position of
HaeIII12
could
notbe
deduced from
partial
mapping,
norcould
1 and 11 be shown to beadjacent.
Correlation
of
the
HincH
and HaeM
re-striction
maps.(i)
Location ofHaeT
sub-cuts
within
Hincd
fragments. Table
4showsthe
fragments
obtained when HincII terminalproducts
are digested withHaeIII.
Fractionationof
St-1
DNAdigested
with bothHinclI
andHaeIII
endonucleases
(data
notshown)
con-firmed
that certainof
theHincII
or HaeIIIfragments
were cutby
HaeIII
orHincII,
respec-tively,
and were thereforemissing,
and that newbands, those designated
asdeletions,
weregen-erated.
Three intact HaeIII
fragments
wereproduced
from
HincId1
by HaeIII digestion(Table
4).They were
identified
asterminal
HaeIIIfrag-ments on
the
basis of their relative
mobility
in
acrylamide
gels. The 850-bp fragment
wasdes-St-1
Hae
mU
0X1 74
+
Hae
ruI
2-O
I-m4
-IS
6-*7
8-me
92__0tw
10- .-;
St-i
Hiiincc
_-1
2
*i*
v-V2 3
4
-em-_-5
5 _ _* -6
6a _
6b
--7
7 ----
4
4bio_ 8 --- 1
11- '' ,. - 9 -
A
-8
10--FIG. 1.Analysisofthe HincII and HaeIII cleav-ageproducts of
St-i
DNA on a4%polyacrylamide
gel. ,X174DNAdigestedwith thesameendonucleasewas fractionated in parallel, although
only
the HaeIIIdigestis shown. The autoradiographof
the driedgelwasscanned withaJoyceLoebl microden-sitometer,and the molarratiosofthefragments from eachdigestwereestimatedfromtheprofiles.on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.501.261.449.114.592.2]TABLE 1. Sizeof HincII, HaeIII restriction enzyme fragments of St-la
HinclI HaeIII
Product Size Product Size
1 1,710 1 1,840
2 1,460 2 920
3 1,050 3 715
4 720 4 600
5 460 5 410
6 328 6 355
7 263 7 323
8 78 8 258
9 50 9 230
10 175
11 115
12 70
aThe sizes in basepairsofmostof theseterminal
digestionproductswerededuced frommeasurements of theirmigration ina4%polyacrylamidegel,relative tothoseof4X174 digested bythesameenzyme (Fig.
1).The size ofHincdII and-2andHaeIII1,-2, and-3
wereobtained by addition of the sizes ofsubfragments
generated from thembyredigestionwith another re-striction enzyme(Tables4and5).
ignated as a deletion of HaeIII2 because the latter fragment mapped next to
HaeIII7
and becauseHaeIII2 has been shownto generatea fragment of this size when digested with HinclI (Table 5). The fourth fragment, 70 bp long, we designated HaeIII12. The map location ofHaeIIL12 wasnotresolvedbypartial mapping, but the sole position remaining was between HaeIII1 and -11. HaeIII12 probably does not
containanHinclI cleavage site becausea
frag-ment with the same mobility asHaeIII12 was
presentinthe
HincII
plus HaeIII digest of St-1. Under the conditions offractionation used (7% polyacrylamide gels),twofragments which differ by as little as 5 bp, which is the minimum possible distance between the cleavage sites of thetwoendonucleases, should have distinguish-able mobilities. TheHaeIII cleavage site demar-cating HaeIIIl from -12 probably is located within HincII3 andnot-1,asclosetothe bound-aryof thesetwoHincll
fragmentsaspossible. If this location is correct,afurthertinyfragment, possibly only 5 bp, which isadeletionofHincII1,
[image:3.501.57.455.341.457.2]mustbeproduced. Sucha fragment wouldnot
TABLE 2. Redigestion ofHincII partial digestion products withHincII
Partial size (bp) Observed redigestion fragments' Sum of fragment sizes (bp)
P1 1 2 1,710 + 1,460=3,170
P2 1 3 1,710 + 1,050=2,760
P3 2 9 5 1,460+50+460= 1,970
P4 6 4 7 328 + 720 + 263=1,311
P5 8 3 78+ 1,050=1,125
P6 970 4 7 720+263 =963
P7 850 9 5 6 50+460 + 328=838
P8 800 5 6 460+328=788
P9 535 9 5 50 + 460=510
Plo 345 7 8 263 + 78=341
1 2 9 5 6 4 7 8 3
a The composition of the HincII partial fragments was deduced from the molar ratios of the terminal fragments generated upon redigestion with HincII. The sizes, given in base pairs, of the partial fragments were measureddirectly from the gel.
bFragment deduced from a partial fragment present in the redigestion product.
TABLE 3. Redigestion of HaeIII partial digestion products with HaeIII
Partialsize (bp) Observed redigestionfragmentsa Sum of fragment sizes (bp)
P1 1 3 1,840+715= 2,555
P2 3 4 715+600= 1,315
P3 2 7 920+323= 1,243
P4 1,130 9 2 230+920= 1,150
P5 1,020 4 8 10 600 +258+175= 1,033
P6 860 4 8 600+ 258=858
P7 795 7 6 11 355+323+115=793
P8 670 7 6 355+323=678
P9 630 5 9 410+230=640
P10 480 6 11 355+115=470
Pll 440 8 10 258+ 175=433
1 3 4 8 10 5 9 2 7 6 11
aTheidentity of the terminal digestion products generated from eachpartial fragmentwasdeduced from their relative mobilitieson 4%polyacrylamide gels.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.56.456.515.651.2]TABLE 4. Digestion ofHincII fragments with HaeIII
HincII fragment (base Fragments obtained bydigestion
plair)a with HaeHI
HincIld
HincII2
HincII3(1,053) HincII4 (720)
HincII5(460)
HincII6 (328)
HincII7 (263)
850AHaeIII2
355 (HaeIII6)
323 (HaeIII7)
115 (HaeIII11)
70 (HaeIII12)b Total1,713
310AHaeIII4 410 (HaeIII5)
258 (HaeIII8) 230 (HaeIII9)
175 (HaeIII10) 75AHaeIII2 Total 1,458
Uncutb
573AHaeIII1
153AHaeIIf3
Total 726
228AHaeIII4
228AHaeIII3 Total 456
Uncut
Uncut
aThe size ofeachfragment produced bydigestion
bythe secondenzyme(HaeIII)wasobtained by
meas-uringitsmobilityonthegel,relativetothoseof St-I
HinclI orHaeIIIfragments. Some second digestion
products were presumed to be certain HaeIII frag-ments,asindicated in parentheses, ifthey coelectro-phoresed with that fragment. Other products were
inferred (see text) tobe deletions, indicated by A, of St-i HaeIIIfragments.
bSeetextfordetails.
be detected inourexperiments. However,a
pos-sibility remains that the 70-bp fragment is a deletion ofHaeIII12 thatretains themobilityof
the intact fragment. In this case the
HincII
cleavage site demarcating 1 from 3 would be within HaeIII12 very close to the HaeIII12/1 border. We could discover no further data to resolve thisquestion.Table 4 showsthat four whole HaeIll
frag-mentswerecoveredby HincII2.The other two productswereprobably deletions oftheHaeIII fragments adjacentto8 and9, that is4and 2, respectively. Moreover, since 8, 10, 5, and9are the only complete HaeIII fragments produced by cleavage of HincII2, HaeIII5and -10 mustbe contiguous.
HincII6 and -7 were uncut by HaeIII and
must be from withinHaeIII fragments greater
than 328 and 263 nucleotides, respectively.
HincII8
and
-9 were notsubjected
toHaeIII
digestion because of their small size, but
ap-peared
uncutin
the
HincII plus
HaeIII
digest of
St-1.
(ii)
Location
of
Hincd
cleavages
within
HaeM fragments. Table
5shows that
7of the
12
HaeIII
fragments did
notpossess anHinclI
cleavage
site.
Since the
uncutfragments
HaeIII7,
-6,and
-11wereshown
tobe
contiguous
(Table
3),
they
mustlie within
anHinclI
frag-mentgreater
than
their
total
length (793
nucleo-tides),
i.e.,
HincIIl,
-2,
or-3.Table
4shows that
these
lie within
Hincdll.
Similarly HaeIII10,
-8,
-5,
and
-9,which
aretogether
onthe
map, mustbe
generated by
HaeIll
cleavages within
afrag-mentgreater
than
1,073nucleotides which
mustbe
HincII2.
HaeIII1
overlaps
HincII7 and
-8.Because the
length of HincII3 could
notbe
accurately
as-sessed, for the
reasonsstated above,
we were notTABLE 5. DigestionofHaeIIIfragmentswith HincII
Fragmentsobtainedby
diges-HaeIII
fragments tion withHincIIa
HaeIIIl 930AHincII3b
573AHincII4 263 (HincII7)
78 (HincII8) Total 1,844
HaeIII2 850AHincII1
75 AHincII2 Total 925
HaeIII3(715) 330 (HincII6) 230AHincII5
153AHincII4 Total 713
HaeIII4 (620) 310AHincII2
228AHincII5
50 (HincII9) Total 588
HaeIII5 (410) Uncut HaeIII6 (355) Uncut HaeIII7 (323) Uncut HaeIII8(258) Uncut
HaeIII9(230) Uncut
HaeIII10 (175) Uncut HaeIII11 (115) Uncut
aThesizes, giveninnucleotides, of the products of
HincIIdigestion of HaeIIIfragmentswerecalculated from theirmobility relativetoSt-1 HincIIorHaeIII terminal digestion fragments. If they coelectropho-resed with knownHincIIfragments, theywere desig-natedassuch(indicatedinparentheses); productsthat
didnotwerethoughttobe deletions ofHincII frag-ments(A).
bSeetextfordetails.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.501.48.241.70.382.2] [image:4.501.256.447.294.589.2]able
todetermine whether
theHinclI site
divid-ing
HincIIl and
-3 waslocated within
HaeIII1
or -12. If
it lies
within HaeIII1, the
fragment,
about
930bp in size, will be intact HincII3,
andan
undetected tiny
fragment which is
adeletion
of the
adjacent
HincId
will also be
produced by
HincII digestion
of HaeIIIl. The second
deletion
fragment
produced from the other end of
HaeIIIl is
adeletion of HincII4. HaeIII12
wasnot
examined
onits
ownfor
anHinclI
cleavage
site but,
asdiscussed
above, appeared
intactwhen St-1 RF
wasdigested
with
both
enzymes.On the basis of these
results,
we wereable
toalign the
tworestriction
maps asshown in
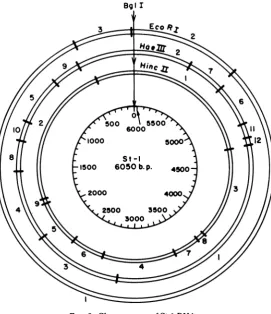
Fig.
2.
EcoRI
and
Bgll
cleavage of
St-i.
The
po-sitions
onthe
mapof the three
EcoRI and
single
Bgl
cuts weredetermined
by
digesting
St-1 RFI
with HincII
orHaeIII
andthen with either
EcoRI
orBglI.
Comparison of the banding
pat-tern obtained
by fractionation of these "double
digests"
onacrylamide
gels with that of the
comparable single
enzymedigest permitted
identification of the
particular fragment
cutby
the
second enzyme. Thesedata
areincluded
onthe
map(Fig.
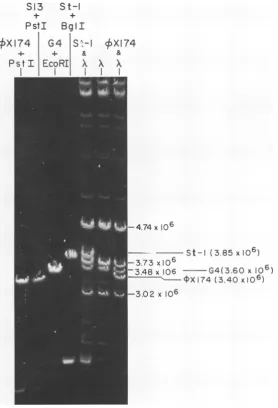
2).Size of
St-1 RF. The
size ofSt-1
RF DNAwas deduced to be about 6,050 bp in two ways:
first,
from the sum of the sizes of fragmentsresulting from digestion of
St-i
RF by theHinclI
or
HaeIII
enzyme; second, relative to that oflinear
4X174, G4,
orS13
(Fig. 3). These werealso compared with
an EcoRI digest ofbacterio-phage
A DNAwhose fragment
sizes are welldocumented
(24).The
fast-moving
bandspres-ent
in the
St-1
+Bgl
andOX174
+ Pst digestswere undigested
St-i
and4X174
RFI,respec-tively.
DISCUSSION
The
analysis of St-1
RF DNAby restriction
endonucleases described
inthis
paperhas
con-firmed
theobservation, based
onsedimentation
values, of Bowes and
Dowell
(3) that the St-1 genomeis
larger than that of
4X174.
Our
obser-vations
suggestthat,
atabout
6,050nucleotide
pairs, the St-1
genomeis 10%
larger
than
OX174.
St-1
DNA isalso
larger than the DNA of the
related isometric
phages G4
andS13 (Fig.
3) [image:5.501.128.400.349.664.2]and,
therefore,
possessesthe
largest
genome ofFIG. 2. Cleavage map of
St-I
DNA.on November 10, 2019 by guest
http://jvi.asm.org/
VOL.27,1978
~~~~~~RESTRICTION
"F 'P
FIG. 3.
Comparison of
themobility
oflinearSt-i DNA(produced
byBglI cleavage)
with thatof
similarly
generated
linearG4,4~X,
and S13 DNAs. An EcoRIdigest of
bacteriophage
A DNAonitsown,ormixed withlinearSt-i1or
4X174
DNAbefore
loading
thegel,
wasfractionated
inparallel (three
slotstoright of figure)
toprovide
the molecularweight
markers noted in thefigure.
The0.7%,o
agarosegel
wasstained with ethidiumbromide
after electrophoresis
andphotographed
with UVillumination.all the isometricphages reportedsofar. The sizes of thefragments producedfrom
St-1 DNA by cleavage with endonuclease HaeIII and Hincll (Fig. 1 and Table 1) arecompletely
different from thoseproducedfrom 4X174DNA. St-i DNA isalso cleavedbyendonucleasesBgl
(once) and EcoRI (three tizmes), which do not
cleave 4X174 DNA.Thus, the St-i genome not
onlyislargerthan that ofOX174,its DNAbase
sequence appears tobe different. However,the viral proteinscoded by St-i appear onsodium
dodecyl sulfate-acrylamidegelstobesimilar to
those codedfor
by 4X174, S13,
andG4(Grindley
and
Godson,
unpublished
data).
Thisimnplies
thatSt-1 has
kept
thesamebasicgenomestrucktureasthe otherisometric
phages
sofarexam-'
mned
(4X174, S13,
andG4).
The extra DNA ofSt-i1
may be due to an increase in size of itsintercistronic spacesor
p'erhaps
tothenonover-lap
inSt-i1
ofthose geneswhich have been shownto
"overlap"
in4X174, viz., D/E
andA/B.
LikeG4
(18),
St-irequires only dnaG, dnaE,
and DNA
binding protein
toinitiate andsynthe-size its
complementary
DNA strand in vitro(14),
VOL. 27,1978
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.109.388.68.477.2]744
but
unlike G4, it
does not require dnaB anddnaC/D
proteins to replicate itsdouble-stranded DNA in vivo (6, 22, 23). At 42°C in
dnaB
anddnaC/D
E. colicells,
St-1 synthesizes
normal amounts
of RF andsingle-stranded
prog-eny DNA (Grindley and Godson,
unpublished
data) and
canform
plaques normally
(2). Thesedifferences
mustultimately
reside indifferences
in the DNA
base sequence and structures ofprotein
recognition
sites. Thegeneration
of theSt-1 restriction endonuclease
cleavage mapde-scribed in
this paper is a prelude to such asequencing study.
ACKNOWLEDGMENTS
This work wassupported by Public Health Service grants CA-06519 from the NationalCancer Institute and5RO1
AI-11633from theNational Institute of Allergy and Infectious Diseases.
LITERATURE CITED
1. Barrell, B. G., G. H. Air, and C. A. Hutchison III. 1976.Overlapping genes in bacteriophage4X174. Na-ture(London) 264:34-41.
2.Bowes, J. M. 1974. Replication ofbacteriophageSt-iin Escherichiacoli strains temperature sensitive in DNA synthesis. J. Virol.13:1400-1403.
3. Bowes, J. M.,andC. E. Dowell. 1974. Purification and some properties of bacteriophage St-1. J. Virol. 13:53-61.
4. Bradley, D. E. 1970. A comparative study of some prop-erties of theOX174type bacteriophages. Can. J. Micro-biol.16:965-971.
5.Chen,C-Y., C. A. Hutchison m, and M. H. Edgell. 1973.Isolation and geneticlocalization of three4X174
promoter regions. Nature (London) New Biol. 243:233. 6. Derstine, P. L., L. B.Dumas, and C. A. Miller. 1976. Bacteriophage G4DNA synthesis in temperature-sen-sitive dna mutants of Escherichia coli. J. Virol. 19:915-924.
7. Dumas, L.B., and C. A. Miller. 1974. Inhibition of bacteriophagefX174DNAreplicationin dnaB mutants ofEscherichia coli C. J. Virol.14:1369-1379. 8. Edgell,M.H., C. A. Hutchisonm,and M.Sclair. 1972.
Specific endonuclease R fragments ofbacteriophage
4OX174
deoxyribonucleicacid. J. Virol.9:574-582.9. Godson,G. N.1975.Evolution ofOX174.II. Acleavage
map of theG4phage genome andcomparisonwith the
cleavagemap of
4OX174.
Virology63:320-335. 10.Godson,G. N. 1977.G4 DNAreplication.III.Synthesisofreplicativeform. J. Mol. Biol. 117:353-367.
11. Godson, G. N., and H. Boyer.1974.Susceptibilityof the
OX-like
phages G4 and G14 toEcoRI endonuclease.Virology62:270-275.
12. Kranias, E.G., and L. B. Dumas. 1975.Replicationof bacteriophage4X174DNA inatemperature-sensitive
dnaC mutant of Escherichia coli C. J. Virol. 13:146-154.
13. Lee,A.S.,and R. L. Sinsheimer.1974. Acleavagemap ofbacteriophage
OX174
genome. Proc.Natl. Acad. Sci. U.S.A. 71:2882-2886.14. McFadden,G., andD. T.Denhardt. 1974.Mechanism ofreplication of4X174single-strandedDNA. IX. Re-quirement for the Escherichia coli dnaGprotein.J. Virol. 14:1070-1075.
15. Maniatis,T.,A.Jeffrey,and H. VandeSande. 1975. Chainlength determinationsof small double- and
sin-gle-stranded DNA molecules by polyacrylamide gel
electrophoresis.Biochemistry14:3787-3794.
16. Middleton,J.H.,M. H.Edgell,andC. A. Hutchison HI.1972.Specific fragmentsof4X174 deoxyribonucleic acidproducedbyarestriction enzyme from
Haemophi-lusaegyptius, endonuclease Zl. J. Virol. 10:42-52. 17.Sanger,G., G.M.Air,B.G.Barrell,N.L.Brown,A.
R.Coulson, J. C.Fiddes,C. A. HutchisonHI,P. M. Slocombe, and M. R. Smith. 1977. Nucleotide se-quence of bacteriophage 4SX174. Nature (London) 265:687-695.
18. Schekman, R., A. Weiner, and A. Kornberg. 1974. Multienzyme system of DNA replication. Science 186:987-993.
19. Sinsheimer, R. L., B. Starman, C. Nagler, and S. Guthrie. 1962. The process of infection with
bacterio-phage
OX174.
I. Evidence fora "replicativeform." J. Mol. Biol. 4:142-160.20. Smith,H.W., andK. W.Wilcox. 1970.Arestriction enzyme fromHaemophilus influenzae.I.Purification andgeneralproperties.J.Mol. Biol. 51:379-391. 21. Sugden, B.,B.DeTroy,R. J. Roberts,and J.
Sam-brook. 1975. Agarose slab-gel electrophoresis
equip-ment. Anal.Biochem. 68:36-46.
22. Taketo, A. 1976. Host factor requirements and some properties of
OXtB.
Mol.Gen. Genet. 148:139-142. 23. Taketo, A.1976.Hostgenes involved in thereplicationofsingle-stranded DNA phage
OK.
Mol. Gen. Genet. 148:848-855.24. Thomas, M., and R. W. Davis. 1975. Studieson the cleavage ofbacteriophage lambda DNA with EcoRI restrictionendonuclease. J. Mol. Biol. 91:315-328. 25. Wickner, S., and J. Hurwitz. 1976. Involvement of
Escherichia coli dnaZ geneproductin DNAelongation
invitro. Proc. Natl. Acad. Sci. U.S.A. 73:1053-1057. 26. Zechel, K.,J-P.Bouche,andA.Kornberg.1975.
Rep-lication ofphageG4:anovel andsimplesystem for the initiation of DNA synthesis. J. Biol. Chem. 250:4684-4689.