Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Will Multiple Coreceptors Need To Be Targeted by Inhibitors
of Human Immunodeficiency Virus Type 1 Entry?
YI-JUN ZHANGANDJOHN P. MOORE*
Aaron Diamond AIDS Research Center, The Rockefeller University, New York, New York Received 15 October 1998/Accepted 14 December 1998
Despite being able to use the Bonzo coreceptor as efficiently as CCR5 in transfected cells, pediatric human immunodeficiency virus type 1 isolate P6 was unable to replicate in peripheral blood mononuclear cells (PBMC) lacking the CCR5 receptor. Furthermore, its replication in wild-type PBMC was completely inhibited by inhibitors of CCR5-mediated entry. Similarly, maternal isolate M6 could use CCR5, CXCR4, Bonzo, and other coreceptors in transfected cells but was completely sensitive to inhibitors of CCR5- and CXCR4-mediated entry when grown in PBMC. The ability of these viruses to use coreceptors in addition to CCR5 and CXCR4 in vitro was, therefore, irrelevant to their drug sensitivity in primary cells. We argue that CCR5 and CXCR4 should remain the primary targets for antiviral drug development, pending strong evidence to the contrary.
The entry of human immunodeficiency virus type 1 (HIV-1) into target cells is now known to involve sequential interactions of the viral envelope glycoproteins with CD4 and a coreceptor (5, 7, 16, 35). The coreceptors are members of the seven-transmembrane-spanning, G-protein-coupled receptor super-family. The first of these proteins identified as being an HIV-1 coreceptor was the CXC chemokine receptor CXCR4, which mediates entry of syncytium-inducing (SI) or T-cell-line-tropic HIV-1 isolates (23). Subsequently, the CC chemokine receptor CCR5 was shown to be the major coreceptor for non-syncyti-um-inducing (NSI) or macrophage-tropic viruses (2, 11, 14, 18, 19). A nomenclature for HIV-1 phenotype based on corecep-tor usage has been proposed, in which viruses able to use CXCR4 are designated X4, those able to use CCR5 are des-ignated R5, and dual-tropic viruses that can use both receptors are called R5X4 (6).
There is strong genetic evidence that CCR5 is the most important coreceptor for the macrophage-tropic viruses that are commonly transmitted between individuals (13, 27, 31, 44). There is also good circumstantial evidence that CXCR4 is the most relevant coreceptor for the T-cell-line-tropic isolates that emerge in a substantial fraction of individuals after several years of HIV-1 infection (8, 12, 28, 49). These coreceptors are, therefore, of clear and obvious interest as targets for antiviral drug development. However, at least 10 other members of the G-protein-coupled receptor superfamily have been shown to have HIV-1 coreceptor activity to greater or lesser extents, when transfected into barren target cells and tested in viral entry and/or fusion assays in vitro. These include CCR2b (18), CCR3 (1, 4, 11, 25, 39), BOB/GPR15 (15, 21, 22), Bonzo/ STRL33/TYMSTR (3, 15, 21, 30, 32), GPR1 (21, 22), CCR8 (26, 41), US28 (38), V28/CX3CR1 (41), APJ (10, 20), and ChemR23 (43). Of these, CCR3 functions most efficiently, with the broadest range of isolates. The question then arises as to whether any among this eclectic gallimaufry of coreceptors is of importance when considering drug development strategies. Will HIV-1 when faced with, e.g., a CCR5-specific inhibitor simply evade the drug by using a different coreceptor in vivo? To a substantial extent, this question can be answered only by
clinical trials of coreceptor-targeted inhibitors in humans or animals. However, to gain some insights into the nature of the problem, we have studied an unusual series of pediatric HIV-1 isolates that are able to use the coreceptors CCR5, Bonzo, and in the case of SI variants, CXCR4, CCR8, V28/CX3CR1, and APJ with approximately equivalent efficiencies in vitro (53). Specifically, we have addressed the issue of whether the ability of these viruses to use Bonzo and other coreceptors affects their sensitivity to inhibitors directed against CCR5 and CXCR4 in primary, CD41T cells.
Growth of Bonzo coreceptor-using HIV-1 in wild-type and
D32-CCR5 peripheral blood mononuclear cells (PBMC).
Vi-ruses designated M6 were isolated from an HIV-1-infected mother who has since died of AIDS (9, 53). They are of the SI phenotype and can use CCR5, CXCR4, Bonzo, CCR8, V28/ CX3CR1, and APJ when these coreceptors are expressed in transfected GHOST or U87-CD4 cells in vitro (53). The P6 isolates are from the mother’s younger, vertically infected child, are of the NSI phenotype, and use both CCR5 and Bonzo in vitro (53). We have previously shown that Bonzo usage by the M6 and P6 isolates is efficient, to an extent comparable with CCR5 use, which is unusual (21, 53).
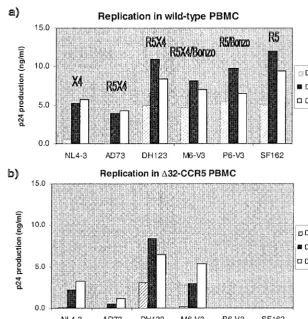
We first addressed whether these isolates were able to rep-licate in PBMC from a human homozygous for defective CCR5 alleles (D32-CCR5), using procedures described previously (51, 53). The maternal (M6) isolate replicated both in the D32-CCR5 cells and in PBMC from a wild-type donor (Fig. 1). The same was also true of other X4 and R5X4 viruses, NL4-3, AD73, and DH123, which is consistent with the ability of all these viruses to use CXCR4, a protein expressed normally on D32-CCR5 cells (31, 51). In contrast, the infant (P6) isolate, like the control SF162 R5 isolate, was completely unable to replicate in theD32-CCR5 cells, although these viruses grew efficiently in wild-type cells (Fig. 1). Similar results were ob-tained in more-stringent cocultivation assays, in that no repli-cation of P6 and SF162 could be detected inD32-CCR5 cells even when they were subsequently cocultivated for 7 days with phytohemagglutinin-activated PBMC from a donor wild type for CCR5 (data not shown). Thus, the ability of the P6 isolate to use Bonzo in transfected cells is irrelevant to its replication in primary PBMC; the absence of CCR5 from theD32-CCR5 cells is clearly not overcome by Bonzo usage.
Sensitivity of Bonzo coreceptor-using HIV-1 to CCR5- and
CXCR4-directed inhibitors.We next assessed the sensitivity of
* Corresponding author. Mailing address: Aaron Diamond AIDS Research Center, 455 1st Ave., 7th Floor, New York, NY 10016. Phone: (212) 725-0018. Fax: (212) 725-1126. E-mail: jmoore@adarc .org.
3443
on November 9, 2019 by guest
http://jvi.asm.org/
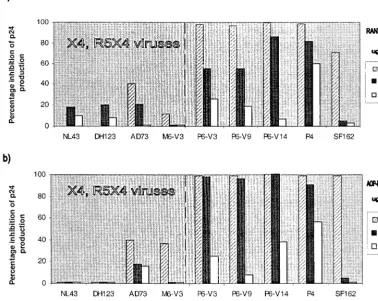
the Bonzo-using P6 and M6 isolates to RANTES and amin-oxypentane-RANTES (AOP-RANTES), two inhibitors of HIV-1 entry via CCR5 (14, 19, 33, 48, 51) (Fig. 2). Like other isolates able to use CXCR4, replication of the M6 isolate in CCR5 wild-type donor PBMC was only weakly inhibited by RANTES and AOP-RANTES, since entry via CXCR4 is un-hindered (Fig. 2). In contrast, the replication of P6 isolates (R5/Bonzo) was as sensitive as the replication of the P4 and SF162 isolates (R5) to RANTES and AOP-RANTES (Fig. 2). The inhibition was dose dependent and eventually complete for all these isolates, showing that Bonzo usage did not permit HIV-1 to evade CCR5-directed inhibitors in PBMC. Of note is the fact that, although the natural ligands for Bonzo are un-known, there is no evidence that Bonzo is a RANTES recep-tor; RANTES does not block P6 replication in GHOST-Bonzo cells (data not shown).
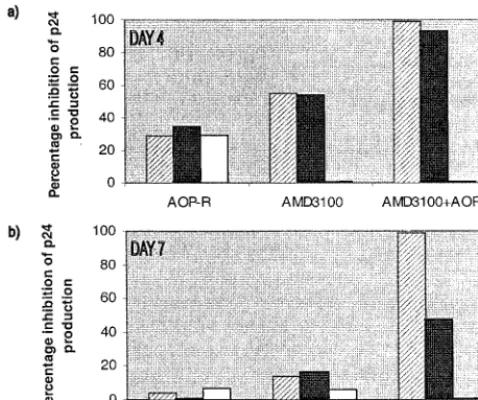
To test the sensitivity of HIV-1 replication to a CXCR4-specific inhibitor, we used the bicyclam AMD3100 (17, 45, 46) (Fig. 3). In PBMC from CCR5 wild-type donors, AMD3100 significantly but only partially inhibited the replication of the dual-tropic M6 isolate, which can still enter the cells via CCR5. The X4 virus NL4-3 was completely inhibited by AMD3100, but the R5/Bonzo virus P6 was insensitive, as expected (Fig. 3a). When PBMC from aD32-CCR5 donor were used, M6 and the control DH123 and NL4-3 isolates were all completely (or almost completely) inhibited by AMD3100 (Fig. 3b). In other experiments, the M6 isolate, which can use Bonzo as well as CXCR4, was indistinguishable from DH123, which cannot use
Bonzo, in its sensitivity to AMD3100 (data not shown). Thus, the ability to use Bonzo does not allow HIV-1 to evade a CXCR4-specific inhibitor. Furthermore, although both DH123 and M6 can also use several other coreceptors in transfected cells, including CCR8, V28/CX3CR1, and APJ (53), they are still sensitive to a CXCR4-specific inhibitor in D32-CCR5 PBMC (Fig. 3b).
The combination of AOP-RANTES with AMD3100 was used to assess the effects of inhibitors directed at both CCR5 and CXCR4 simultaneously on the infection of healthy donor PBMC by the maternal isolate M6 (Fig. 4). Alone, AOP-RANTES and AMD3100 could only partially inhibit M6 infec-tivity, but when they were used together, inhibition was com-plete (Fig. 4). Similar results were obtained with the DH123 isolate (data not shown). Thus, despite the ability of M6 to enter transfected cells via other coreceptors, such as Bonzo, CCR8, V28/CX3CR1, and APJ (53), a combination of a CCR5 inhibitor and a CXCR4 inhibitor was sufficient to block its replication in PBMC from a wild-type CCR5 donor.
Discussion.Although SI, T-cell-line-tropic viruses are often
[image:2.612.149.457.71.390.2]able to use multiple coreceptors for entry into transfected cell lines, the efficient use of coreceptors other than CCR5 by NSI, macrophage-tropic HIV-1 strains is rare (21, 53). CCR3 usage by NSI viruses can be demonstrated in transfected cells (1, 4, 11, 25, 39). However, among the many viruses we have tested, only a single set of NSI isolates (P6 series) from an HIV-1-infected child was able to use a receptor other than CCR5 as efficiently as CCR5 itself could be used; these isolates entered FIG. 1. Replication of HIV-1 isolates in PBMC from wild-type andD32-CCR5 donors. The HIV-1 isolates indicated were tested for their ability to replicate in mitogen-stimulated PBMC from wild-type (a) andD32-CCR5 (b) donors. Virus replication was assessed by p24 antigen production on days 4, 7, and 11, as described previously (51, 53). The coreceptors that can be used by each isolate is given in panel a. For M6-V3 and DH123, this is incomplete, and a more-detailed description is provided elsewhere (53). Similar results were obtained with cells from two donors and when other isolates of the P6 and M6 series were tested.
on November 9, 2019 by guest
http://jvi.asm.org/
Bonzo- or CCR5-expressing cells with approximately equiva-lent efficiency (53). SI isolates (M6 series) from the mother of this child used CCR5, CXCR4, Bonzo, and other coreceptors (CCR8, V28/CX3CR1, and APJ). The availability of these Bonzo-using isolates allowed us to address whether corecep-tors other than CCR5 and CXCR4 are likely to be of signifi-cant importance for antiviral drug development strategies
[image:3.612.116.495.87.388.2]aimed at the HIV-1 coreceptors. For example, had we found that an otherwise effective, CCR5-specific inhibitor such as AOP-RANTES was unable to completely inhibit the replica-tion of the P6 isolates in vitro, it would have implied that Bonzo provided an alternative, unhindered route for HIV-1 entry into PBMC. An inference would then be that the acqui-sition of Bonzo use might be an evolutionary pathway for viral FIG. 2. Sensitivity of HIV-1 isolates to RANTES and AOP-RANTES in wild-type PBMC. The HIV-1 isolates indicated were tested for their ability to replicate in mitogen-stimulated PBMC from a wild-type donor in the presence of the indicated concentrations of RANTES (a) or AOP-RANTES (b). Virus replication was assessed by p24 antigen production on day 7 and related to the amount produced in the absence of inhibitor (defined as 100%). The isolates to the left of the broken line in each panel can use CXCR4; those to the right cannot. Similar results were obtained with cells from two donors and with other isolates of the P6 and M6 series.
FIG. 3. Sensitivity of HIV-1 isolates to AMD3100 in PBMC from wild-type andD32-CCR5 donors. The HIV-1 isolates indicated were tested for their ability to replicate in mitogen-stimulated PBMC from wild-type (a) andD32-CCR5 (b) donors in the presence of the indicated concentrations of AMD3100. Virus replication was assessed by p24 antigen production on day 4 and related to the amount produced in the absence of inhibitor (defined as 100%). Inhibition of M6 replication by AMD3100 exceeded 99% when the cultures were retested on day 7.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.63.540.556.691.2]escape from a CCR5-specific inhibitor. Similar arguments could be made for an SI virus such as M6, under the selection pressure of a combination of CCR5- and CXCR4-specific in-hibitors.
We observed, however, that the ability of the P6 and M6 viruses to use Bonzo did not overtly affect their sensitivity to AOP-RANTES and AMD3100, alone or in combination, when these were used as prototypic inhibitors of entry via CCR5 and CXCR4, respectively. We cannot exclude the possibility of subtle influences of the usage of Bonzo (or other coreceptors) on drug sensitivity, but the P6 and M6 viruses were neither unusually sensitive or insensitive to AOP-RANTES or AMD3100 in our experience. These experiments therefore suggest that the ability of the P6 and M6 isolates to use Bonzo in trans-fected cells is irrelevant to their replication in PBMC. This conclusion is further supported by the observation that P6 viruses were unable to replicate in PBMC from aD32-CCR5 homozygous individual. Bonzo (STRL33) is expressed at the mRNA level in PBMC (15, 21, 30) but is not used for entry of the P6 isolate. Furthermore, the replication of the M6 isolate in the D32-CCR5 PBMC was completely blocked by the CXCR4-specific inhibitor AMD3100, despite M6 also being able to use Bonzo, CCR8, V28/CX3CR1, and APJ in trans-fected cells (53). The same reservations about the relevance of Bonzo usage for HIV-1 replication in PBMC may therefore apply also to CCR8, V28/CX3CR1, and APJ.
Viruses isolated from an HIV-1-infected individual who was homozygous for D32-CCR5 alleles were found to use only CXCR4 when tested against a variety of coreceptor-expressing cell lines in vitro (34). Of note is that the relative levels of mRNAs for several coreceptors in PBMC have been reported to be as follows: CXCR4, 150; CCR5, 100; CCR2b, 15; and CCR3, 10 (36). In addition, fluorescence-activated cell sorting analysis shows that CCR3 is expressed in PBMC at only about 1% of the level of CCR5 expression (42). All of these
obser-vations are consistent with a paramount role of CCR5 and CXCR4 for HIV-1 replication in peripheral blood cells. In principle, any of the coreceptors that have been described as functional for HIV-1 entry in transfected cells could provide an escape route for HIV-1 under drug selection pressure in vivo. However, we could find no reason to believe this is likely to occur. We note that an SDF-1 escape mutant of NL4/3, derived in vitro, still retained CXCR4 usage, in a manner that was less sensitive to the inhibitor (29, 45). Coreceptors other than CCR5 and CXCR4 might be important for HIV-1 replication in nonlymphoid cells, including those found at mucosal sur-faces (20, 24, 25, 36, 40, 47, 50), in minor lymphocyte subsets (42, 54), or in specialist lymphoid tissues, such as the thymus (especially in infants) (52). However, it has been shown that 99% of virus production in HIV-1-infected people is produced by CD41T lymphocytes (37). Inhibition of HIV-1 replication
in these cells is of major clinical benefit, so usage of corecep-tors other than CCR5 and CXCR4 should not be the primary concern in the development of antiviral compounds directed at blocking HIV-1 entry into CD41T cells.
The priority in the development of coreceptor-targeted an-tiviral compounds should, we believe, remain squarely on CCR5 and CXCR4 pending strong evidence to the contrary. A major issue will be whether the use of a CCR5-specific inhib-itor will drive the evolution of an NSI virus towards CXCR4 usage and the consequent acquisition of the SI phenotype. This will require careful evaluation in vitro and in vivo should suit-able drugs be developed. However, it is likely that any core-ceptor-targeted inhibitors will be used in combination with existing antiviral drugs such as protease and reverse transcrip-tase inhibitors. Their suppression of HIV-1 replication will reduce the probability of phenotypic evolution. Nonetheless, the powerful potential of HIV-1 to evade any inhibitors of its replication must always be recognized, so the routes it might take in such evasion need to be carefully defined.
We appreciate the contributions of the donors of HIV-1 isolates and clones used in this study, in particular the participants in the Pediatric AIDS Foundation’s ARIEL Project for isolates M6 and P6. We par-ticularly thank Amanda Proudfoot (Serono Research Institute, Ge-neva, Switzerland) for the gifts of RANTES and AOP-RANTES, Bahige Baroudy (Schering Plough Research Institute, Nutley, N.J.) for AMD3100, and Dan Littman and Vineet KewalRamani for cell lines. We are grateful to Cecilia Cheng-Mayer and Alexandra Trkola for interesting discussions and to Tom Ketas and Ivor Biggun for PBMC preparations.
This study was supported in part by NIH grant AI41420 and by the Pediatric AIDS Foundation, for which J.P.M. is an Elizabeth Glaser Scientist.
REFERENCES
1.Alkhatib, G., E. A. Berger, P. M. Murphy, and J. E. Pease.1997. Determi-nants of HIV-1 co-receptor function on CC chemokine receptor 3. Impor-tance of both extracellular and transmembrane/cytoplasmic regions. J. Biol. Chem.272:20420–20426.
2.Alkhatib, G., C. Combadiere, C. C. Broder, Y. Feng, P. E. Kennedy, P. M. Murphy, and E. A. Berger.1996. CC CKR5: A RANTES, MIP-1a, MIP-1b receptor as a fusion cofactor for macrophage-tropic HIV-1. Science272:
1955–1958.
3.Alkhatib, G., F. Liao, E. A. Berger, J. M. Farber, and K. C. W. Peden.1997. A new SIV coreceptor, STRL33. Nature388:238.
4.Bazan, H. A., G. Alkhatib, C. C. Broder, and E. A. Berger.1998. Patterns of CCR5, CXCR4, and CCR3 usage by envelope glycoproteins from human immunodeficiency virus type 1 primary isolates. J. Virol.72:4485–4491. 5.Berger, E. A.1997. HIV entry and tropism: the chemokine receptor
connec-tion. AIDS11(Suppl. A):S3–S16.
6.Berger, E. A., R. W. Doms, E.-M. Fenyo¨, B. T. M. Korber, D. R. Littman, J. P. Moore, Q. J. Sattentau, H. Schuitemaker, J. Sodroski, and R. A. Weiss.1998. A new classification for HIV-1. Nature391:240.
7.Bieniasz, P. D., and B. R. Cullen.1998. Chemokine receptors and human immunodeficiency virus infection. Front. Biosci.3:44–58.
[image:4.612.53.292.75.275.2]8.Bjo¨rndal, Å., H. Deng, M. Jansson, J. R. Fiore, C. Colognesi, A. Karlsson, J.
FIG. 4. Complete inhibition of HIV-1 by a combination of AOP-RANTES and AMD3100. Replication of the HIV-1 isolate M6-V3 in PBMC from a wild-type donor was tested in the presence of AOP-RANTES (AOP-R) and AMD3100. These compounds were each tested at 1, 0.1, and 0.01 mg/ml, as indicated by the hatched solid, and white bars, respectively. The same final concentrations of each compound were present in the AOP-RANTES plus AMD3100 combination. Virus replication was assessed by p24 antigen produc-tion on days 4 and 7 and related to the amount produced in the absence of inhibitor (defined as 100%).
on November 9, 2019 by guest
http://jvi.asm.org/
Albert, G. Scarlatti, D. R. Littman, and E. M. Fenyo¨.1997. Coreceptor usage of primary human immunodeficiency virus type 1 isolates varies according to biological phenotype. J. Virol.71:7478–7487.
9.Cao, Y., P. Krogstad, B. T. Korber, R. A. Koup, M. Muldoon, C. Macken, J. L. Song, Z. Jin, J.-Q. Zhao, S. Clapp, I. S. Y. Chen, D. D. Ho, A. J. Ammann, and the Ariel Project Investigators.1997. Maternal HIV-1 viral load and vertical transmission of infection: the Ariel Project for the preven-tion of HIV transmission from mother to infant. Nat. Med.3:549–552. 10. Choe, H., M. Farzan, M. Konkel, K. Martin, Y. Sun, L. Marcon, M.
Cay-abyab, M. Berman, M. E. Dorf, N. Gerard, G. Gerard, and J. Sodroski.1998. The orphan seven-transmembrane receptor Apj supports the entry of pri-mary T-cell-line-tropic and dualtropic human immunodeficiency virus type 1. J. Virol.72:6113–6118.
11. Choe, H., M. Farzan, Y. Sun, N. Sullivan, B. Rollins, P. D. Ponath, L. Wu, C. R. Mackay, G. LaRosa, W. Newman, N. Gerard, G. Gerard, and J. Sodroski.1996. The b-chemokine receptors CCR3 and CCR5 facilitate infection by primary HIV-1 isolates. Cell85:1135–1148.
12. Connor, R. I., K. E. Sheridan, D. Ceradini, S. Choe, and N. R. Landau.1997. Change in coreceptor use correlates with disease progression in HIV-1 infected individuals. J. Exp. Med.185:621–628.
13. Dean, M., M. Carrington, C. Winkler, G. A. Huttley, M. W. Smith, R. Allikmets, J. J. Goedert, S. P. Buchbinder, E. Vittinghoff, E. Gomperts, S. Donfield, D. Vlahov, R. Kaslow, A. Saah, C. Rinaldo, R. Detels, Hemophilia Growth and Development Study, Multicenter AIDS Cohort Study, Multi-center Hemophilia Cohort Study, San Francisco City Cohort, ALIVE Study, and S. J. O’Brien.1996. Genetic restriction of HIV-1 infection and progres-sion to AIDS by a deletion of the CKR5 structural allele. Science273:1856– 1862.
14. Deng, H. K., R. Liu, W. Ellmeier, S. Choe, D. Unutmaz, M. Burkhart, P. Di Marzio, S. Marmon, R. E. Sutton, C. M. Hill, S. C. Peiper, T. J. Schall, D. R. Littman, and N. R. Landau.1996. Identification of a major coreceptor 5 for primary isolates of HIV-1. Nature381:661–666.
15. Deng, H. K., D. Unutmaz, V. N. Kewalramani, and D. R. Littman.1997. Expression cloning of new receptors used by simian and human immunode-ficiency viruses. Nature388:296–300.
16. Doms, R. W., and S. C. Peiper.1997. Unwelcome guests with master keys: how HIV uses chemokine receptors for cellular entry. Virology235:179–190. 17. Donzella, G. A., D. Schols, S. W. Lin, J. A. Este´, K. A. Nagashima, P. J. Maddon, G. P. Allaway, T. P. Sakmar, G. Henson, E. De Clercq, and J. P. Moore.1998. AMD3100, a small molecule inhibitor of HIV-1 entry via the CXCR4 co-receptor. Nat. Med.4:72–77.
18. Doranz, B. J., J. Rucker, Y. Yi, R. J. Smyth, M. Samson, S. Peiper, M. Parmentier, R. G. Collman, and R. W. Doms.1996. A dual-tropic, primary HIV-1 isolate that uses fusion and theb-chemokine receptors CKR-5, CKR-2b as fusion cofactors. Cell85:1149–1159.
19. Dragic, T., V. Litwin, G. P. Allaway, S. R. Martin, Y. Huang, K. A. Na-gashima, C. Cayanan, P. J. Maddon, R. A. Koup, J. P. Moore, and W. A. Paxton.1996. HIV-1 entry into CD41cells is mediated by the chemokine
receptor CC-CKR-5. Nature381:667–673.
20.Edinger, A. L., T. L. Hoffman, M. Sharron, B. Lee, Y. Yi, W. Choe, D. C. Kolson, B. Mitrovic, Y. Zhou, D. Faulds, R. G. Collman, J. Hesselgesser, R. Horuk, and R. W. Doms.1998. An orphan seven-transmembrane domain receptor expressed widely in the brain functions as a coreceptor for human immunodeficiency virus type 1 and simian immunodeficiency virus. J. Virol.
72:7934–7940.
21.Edinger, A. L., T. L. Hoffman, M. Sharron, B. Lee, B. O’Dowd, and R. W. Doms.1998. Use of GPR1, GPR15, and STRL33 as co-receptors by diverse human immunodeficiency virus type 1 and simian immunodeficiency virus envelope proteins. Virology249:367–378.
22.Farzan, M., H. Choe, K. Martin, L. Marcon, W. Hofmann, G. Karlsson, Y. Sun, P. Barrett, N. Marchand, N. Sullivan, N. Gerard, C. Gerard, and J. Sodroski.1997. Two orphan seven-transmembrane segment receptors which are expressed in CD4-positive cells support simian immunodeficiency virus infection. J. Exp. Med.186:405–411.
23. Feng, Y., C. C. Broder, P. E. Kennedy, and E. A. Berger.1996. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane G protein coupled receptor. Science272:872–877.
24. Ghorpade, A., M. Q. Xia, B. T. Hyman, Y. Persidsky, A. Nukuna, P. Bock, M. Che, J. Limoges, H. E. Gendelman, and C. R. Mackay.1998. Role of the b-chemokine receptors CCR3 and CCR5 in human immunodeficiency virus type 1 infection of monocytes and microglia. J. Virol.72:3351–3361. 25. He, J., Y. Chen, M. Farzan, H. Choe, A. Ohagen, S. Gartner, J. Busciglio, X.
Yang, W. Hofmann, W. Newman, C. R. Mackay, J. Sodroski, and D. Gabuzda.1997. CCR3 and CCR5 are co-receptors for HIV-1 infection of microglia. Nature385:645–649.
26. Horuk, R., J. Hesselgesser, Y. Zhou, D. Faulds, M. Halks-Miller, S. Harvey, D. Taub, M. Samson, M. Parmentier, J. Rucker, B. J. Doranz, and R. W. Doms.1998. The CC chemokine I-309 inhibits CCR8 dependent infection by diverse HIV-1 strains. J. Biol. Chem.273:386–391.
27. Huang, Y., W. A. Paxton, S. M. Wolinsky, A. U. Neumann, L. Zhang, T. He, S. Kang, D. Ceradini, Z. Jin, K. Yazdanbaksh, K. Kuntsman, D. Erickson, N. R. Landau, J. Phair, D. D. Ho, and R. A. Koup.1996. The role of a mutant
CCR5 allele in HIV-1 transmission and disease progression. Nat. Med.
2:1240–1243.
28. Jansson, M., M. Popovic, A. Karlsson, F. Cocchi, P. Rossi, J. Albert, and H. Wigzell.1996. Sensitivity to inhibition byb-chemokines correlates with bio-logical phenotypes of primary HIV-1 isolates. Proc. Natl. Acad. Sci. USA
93:15382–15387.
29. Labrosse, B., A. Brelot, N. Heveker, N. Sol, D. Schols, E. DeClerq, and M. Alizon.1998. Determinants for sensitivity of human immunodeficiency virus coreceptor CXCR4 to the bicyclam AMD3100. J. Virol.72:6381–6388. 30. Liao, F., G. Alkhatib, K. W. C. Peden, G. Sharma, E. A. Berger, and J. M.
Farber.1997. STRL33, a novel chemokine receptor-like protein, functions as a fusion cofactor for both macrophage-tropic and T cell line-tropic HIV-1. J. Exp. Med.185:2015–2023.
31. Liu, R., W. A. Paxton, S. Choe, D. Ceradini, S. R. Martin, R. Horuk, M. E. MacDonald, H. Stuhlmann, R. A. Koup, and N. R. Landau.1996. Homozy-gous defect in HIV-1 coreceptor accounts for resistance of some multiply-exposed individuals to HIV-1 infection. Cell86:367–378.
32. Loetscher, M., A. Amara, E. Oberlin, N. Brass, D. F. Legler, P. Loetscher, M. D’Apuzzo, E. Meese, D. Rousset, J.-L. Virelizier, M. Baggiolini, F. Arenzana-Seisdedos, and B. Moser.1997. TYMSTR, a putative chemokine receptor selectively expressed in activated T cells, exhibits HIV-1 coreceptor function. Curr. Biol.7:652–660.
33. Mack, M., B. Luckow, P. J. Nelson, J. Lihak, G. Simmons, P. R. Clapham, N. Signoret, M. Marsh, M. Stangassinger, F. Borlat, T. N. C. Wells, D. Schlo¨ndorff, and A. E. I. Proudfoot.1998. Aminooxypentane-RANTES in-duces CCR5 internalization but inhibits recycling: a novel inhibitory mech-anism of HIV infectivity. J. Exp. Med.187:1215–1224.
34. Michael, N. L., J. A. E. Nelson, V. N. KewalRamani, G. Chang, S. J. O’Brien, J. R. Mascola, B. Volsky, M. Louder, G. C. White II, D. R. Littman, R. Swanstrom, and T. R. O’Brien.1998. Exclusive and persistent use of the entry coreceptor CXCR4 by human immunodeficiency virus type 1 from a subject homozygous for CCR5D32. J. Virol.72:6040–6047.
35. Moore, J. P., A. Trkola, and T. Dragic.1997. Co-receptors for HIV-1 entry. Curr. Opin. Immunol.9:551–562.
36. Patterson, B. K., A. Landay, J. Andersson, C. Brown, H. Behbahani, D. Jiyamapa, Z. Burki, D. Stanislawski, M. A. Czerniewski, and P. Barcia.1998. Repertoire of chemokine receptor expression in the female genital tract. Am. J. Pathol.153:481–490.
37. Perelson, A., A. U. Neumann, M. Markowitz, J. Leonard, and D. D. Ho.1996. HIV-1 dynamics in vivo: virion clearance rate, infected cell life-span, and viral generation time. Science271:1582–1586.
38. Pleskoff, O., C. Treboute, A. Brelot, N. Heveker, M. Seman, and M. Alizon.
1997. Identification of a chemokine receptor encoded by human cytomega-lovirus as a cofactor for HIV-1 entry. Science276:1874–1878.
39. Ross, T. M., and B. R. Cullen. 1998. The ability of HIV type 1 to use CCR-3 as a coreceptor is controlled by envelope V1/V2 sequences acting in conjunction with a CCR-5 tropic V3 loop. Proc. Natl. Acad. Sci. USA95:7682–7686. 40. Rubbert, A., C. Combadiere, M. Ostrowski, J. Arthos, M. Dybul, E.
Machado, M. A. Cohn, J. A. Hoxie, P. M. Murphy, A. S. Fauci, and D. Weissman.1998. Dendritic cells express multiple chemokine receptors used as coreceptors for HIV entry. J. Immunol.160:3933–3941.
41. Rucker, J., A. L. Edinger, M. Sharron, M. Samson, B. Lee, J. F. Berson, Y. Yi, B. Margulies, R. G. Collman, B. J. Doranz, M. Parmentier, and R. W. Doms.1997. Utilization of chemokine receptors, orphan receptors, and her-pesvirus-encoded receptors by diverse human and simian immunodeficiency viruses. J. Virol.71:8999–9007.
42. Sallusto, F., C. R. Mackay, and A. Lanzavecchia.1997. Selective expression of the eotaxin receptor CCR3 by human T helper 2 cells. Science277:2005– 2007.
43. Samson, M., A. L. Edinger, P. Stordeur, J. Rucker, V. Verhasselt, M. Shar-ron, C. Govaerts, C. Mollereau, G. Vassart, R. W. Doms, and M. Parmentier.
1998. ChemR23, a putative chemoattractant receptor, is expressed in mono-cyte-derived dendritic cells and macrophages and is a coreceptor for SIV and some primary HIV-1 strains. Eur. J. Immunol.28:1689–1700.
44. Samson, M., F. Libert, B. J. Doranz, J. Rucker, C. Liesnard, C.-M. Farber, S. Saragosti, C. Lapoume`roulie, J. Cogniaux, C. Forceille, G. Muyldermans, C. Verhofstede, G. Burtonboy, M. Georges, T. Imai, S. Rana, Y. Yi, R. J. Smyth, R. G. Collman, R. W. Doms, G. Vassart, and M. Parmentier.1996. Resistance to HIV-1 infection of Caucasian individuals bearing mutant al-leles of the CCR5 chemokine receptor gene. Nature382:722–725. 45. Schols, D., J. A. Este, C. Cabrera, and E. DeClerq.1998. T-cell-line-tropic
human immunodeficiency virus type 1 that is made resistant to stromal cell-derived factor 1acontains mutations in the envelope gp120 but does not show a switch in coreceptor use. J. Virol.72:4032–4037.
46. Schols, D., S. Struyf, J. Van Damme, J. A. Este´, G. Henson, and E. De Clerq.
1997. Inhibition of T-tropic HIV strains by selective antagonization of the chemokine receptor CXCR4. J. Exp. Med.186:1383–1388.
47. Shieh, J. T. C., A. V. Albright, M. Sharron, S. Gartner, J. Strizki, R. W. Doms, and F. Gonzalez-Scarano.1998. Chemokine receptor utilization by human immunodeficiency virus type 1 isolates that replicate in microglia. J. Virol.72:4243–4249.
48. Simmons, G., P. R. Clapham, C. Picard, R. E. Offord, M. M. Rosenkilde,
on November 9, 2019 by guest
http://jvi.asm.org/
T. W. Schwartz, R. Buser, T. N. C. Wells, and A. E. I. Proudfoot.1997. Potent inhibition of HIV-1 infectivity in macrophages and lymphocytes by a novel CCR5 antagonist. Science276:276–279.
49.Simmons, G., D. Wilkinson, J. D. Reeves, M. T. Dittmar, S. Beddows, J. Weber, G. Carnegie, U. Desselberger, P. W. Gray, R. A. Weiss, and P. R. Clapham.1996. Primary, syncytium-inducing human immunodeficiency virus type 1 isolates are dual-tropic and most can use either Lestr or CCR5 as coreceptors for virus entry. J. Virol.70:8355–8360.
50. Spira, A. I., P. A. Marx, B. K. Patterson, J. Mahone, R. A. Koup, S. M. Wolinsky, and D. D. Ho.1996. Cellular targets of infection and route of viral dissemination following intravaginal inoculation of SIV into rhesus ma-caques. J. Exp. Med.183:215–225.
51. Trkola, A., W. A. Paxton, S. P. Monard, J. A. Hoxie, M. A. Siani, D. A. Thompson, L. Wu, C. R. Mackay, R. Horuk, and J. P. Moore.1998. Genetic
subtype-independent inhibition of human immunodeficiency virus type 1 replication by CC and CXC chemokines. J. Virol.72:396–404.
52.Zaitseva, M. B., S. Lee, R. L. Rabin, H. L. Tiffany, J. M. Farber, K. W. C. Peden, P. M. Murphy, and H. Golding.1998. CXCR4 and CCR5 on human thymocytes: biological function and role in HIV-1 infection. J. Immunol.
161:3103–3113.
53.Zhang, Y.-J., T. Dragic, Y. Cao, L. Kostrikis, D. S. Kwon, D. R. Littman, V. N. KewalRamani, and J. P. Moore.Use of coreceptors other than CCR5 by non-syncytium-inducing adult and pediatric isolates of human immuno-deficiency virus type 1 is rare in vitro. J. Virol.72:9337–9344.
54.Zingoni, A., H. Soto, J. A. Hedrick, F. Stoppacciaro, C. T. Storlazzi, F. Sinigaglia, D. D’Ambrosio, A. O’Garra, D. Robinson, M. Rocchi, A. Santoni, A. Zlotnick, and M. Napolitano.1998. The chemokine receptor CCR8 is preferentially expressed in Th2 but not Th1 cells. J. Immunol.161:547–551.