JOURNALOF VIROLOGY, Jan.1980,p. 18-27
0022-538X/80/01-0018/10$02.00/0 Vol. 33,No. 1
Frog Virus
3
Replication:
Analysis
of
Structural
and
Nonstructural Polypeptides
in Infected
BHK Cells by
Acidic
and Basic Two-Dimensional
Gel Electrophoresis
R. M.ELLIOTT,'t R. BRAVO,2t ANDD. C. KELLY`*
Natural Environment Research Council, Unit of InvertebrateVirology, Oxford,OXI 3UB,' and Department
of
Biochemistry,
University of Oxford,
Oxford, OXI
3QU,2
UnitedKingdom
Analysis of frog virus 3-infected BHKcellsby two-dimensional,acidic and basic
gel electrophoresis showed that at least 90 infected cell-specific polypeptides
could be detected. These polypeptides represent between 70 and 85% of the
coding capacity of the viralgenome.The polypeptidesweresequentiallyinduced
in atleast three phases. The virus gradually suppressed host cell polypeptide
synthesisduring infection, although the synthesisofafew cell polypeptidesmay
be "switched off' earlyin infection.
Frog virus 3 (FV3) is alargeicosahedral
DNA-containing virus which replicates in the cyto-plasm butrequiresa functional cellnucleusfor
replicationto occur(8,9,11).Theviruspossesses a linear double-stranded DNAgenomehavinga molecular weight of about
108
(10) and so hasthe potential tosynthesize about 100
polypep-tides ofaverage size. Recent studies inour
lab-oratory with sensitive high-resolution
one-di-mensional gel
electrophoresis
resolved 29in-fectedcell-specific polypeptides(ICPs),ofwhich
22 comigrated with virus structural
polypep-tides. The ICPs appeared to be coordinately
regulatedin acascade fashion, with three
tem-poral phases (a, /3, and y) being observed (6).
Thesynthesis ofy
polypeptides
requiressynthe-sisof viralprogenyDNAto occur(5, 6).In
baby
hamsterkidney (BHK 21/13) cellstheaphase
polypeptides are detected from 2 hafter
infec-tion, the 18
polypeptides
are detected from4 h,and the-y-phasepolypeptidesaredetected from
6h. Thepolypeptides detectedrepresent 30% of
thecodingcapacity of the viral genome.
Thedevelopmentoftwo-dimensionalgel
elec-trophoretic
techniques
to resolveproteins by
separating proteins
inonedimensionaccording
to charge with isoelectric focusing (IEF) and
then separating them in the second dimension
accordingtosize with conventional
electropho-resis in the presence of sodium
dodecyl
sulfate(SDS)andreducingagents hasproduceda
pow-erful tool to
explore
complex proteins
(13, 14).In this research we used this
methodology
toresolve virus-induced polypeptides in
FV3-in-t PresenFV3-in-t-'address: DeparFV3-in-tmenFV3-in-t of Microbiology, MounFV3-in-t SinaiSchool9fMedicine, CityUniversityof NewYork,New
York,NY100,9,
$ Present address: Kemisk Institut, Landgelansgade 140, DK-8000 ArhusD,Denmark.
fected cells and soexaminethenumber of
poly-peptides presentin infectedcells, to determine the effect of the virus on host cellpolypeptides,
and also to confirm the temporal induction of FV3polypeptides.
MATERIALS AND METHODS
Materials. L-[35S]methionine (840 Ci/mmol) and
125I
(100 mCi/ml) were purchased from the Radi-ochemical Centre, Amersham, England. Ampholineswerepurchased from L.K.B.Ltd.,Croydon, England.
DNase I and RNase Awereobtained from theSigma
Chemical Co.Ltd., London,England.
Cells and virus.Babyhamsterkidney (BHK21/
13)cellsand fathead minnow (FHM)cellsweregrown
and maintainedaspreviously described,and FV3was
grown and titrated in FHM cells as previously
de-scribed(4).
Radiolabeling of cellular polypeptides with
[3"S]methionine.
BHK cells were grown in small plastic petri dishes (30 mm; 3 x 105 cells) at 37°Covernight. Cellswereinfectedat25to30PFU of FV3
percellbyadsorbingat roomtemperaturefor1hand thenwashingwith theGlasgowmodification ofEagle
minimal essential medium. Theinfectionwasallowed
toproceedat28°C.Cellswerepretreatedwith radio-labeling medium (2% dialyzed calf serum in Earle balanced saltsolution)for15minbeforebeing pulsed
with 100,uCiofL-[35S]methionine (in1ml)for1hat
28°C. The isotope was removed, and the cellswere
washed three times in ice-cold phosphate-buffered
saline and thenpelleted.The cellswereresuspended
in 100
p1
of 10 mM Tris-hydrochloride (pH 7.4)-50mMNaCl-5mMMgCl2 containing25,ug ofDNase I
perml and25,ugofRNase A perml. The cellswere
sonicated andthenincubatedonice for 30 min. The cellswereimmediatelyfreeze dried.
Two-dimensionalgel electrophoresis of poly-peptides. The procedures followed wereessentially
thosederivedbyO'Farrelland co-workers(13, 14).
Freeze-driedcellswereresuspendedin 100,lIoflysis
buffer(9.4Murea,2% NonidetP-40,1.6%ampholines
18
on November 10, 2019 by guest
http://jvi.asm.org/
ANALYSIS OF FV3 POLYPEPTIDES 19
[pH 5 to 7], 0.4% ampholines [pH3.5 to 10], 5%
/-mercaptoethanol), and 250,000cpmof material(about
20
Ad)
wasappliedtoeachelectrofocusing gel. The first-dimension separations were performedeitheras(i)acidic 4% polyacrylamide gels (130 by2.5
mm) containing2%ampholines (1.6% pH5to7,0.4% pH3.5to10)or(ii) basicnonequilibrium pH gradient
electrophoresis (NEPHGE) 4% polyacrylamide gels (130 by2.5mm) containing2%ampholines (1% pH7
to9, 1%pH8to9.5) exactlyasdescribedbyO'Farrell
and co-workers(13, 14).Both IEFandNEPHGEgels
werethen immersed in equilibrium buffer(0.06Tris,
pH 6.8,4% SDS, 10%,8-mercaptoethanol,20% glycerol)
and incubatedatroomtemperaturefor30min before
being applied tothesecondSDS-polyacrylamide di-mension. Gelstobe stored frozenwereincubated for
15min only,andthe incubationwascompleted after
defrosting.
The second-dimension separations by SDS-poly-acrylamide gel electrophoresis of the proteins
con-tained in theIEFgelswereaccomplished by layering
theequilibrated gelsontoastacking gel (4.75%
acryl-amide,0.024%bisacrylamide, 0.1% SDS,0.0625M Tris-hydrochloride, pH 6.8) which overlaid a 15%
acryl-amidegel (0.1% SDS,0.375 MTris-hydrochloride, pH
8.8). A 1% agarose gel made up in electrophoresis
buffer andcontaining0.01%bromophenol bluesecured
thegelinplace.Theelectrophoresis buffer contained
0.025 MTris,0.192 Mglycine,and0.1% SDS, pH8.8.
Electrophoresiswasperformedat6to10mA for14to 18h. The slab measured19by16cm.The gelswere
processedforfluorography (2) by being fixed in 45% methanol-7.5%acetic acid-47.5%waterfor1h,
incor-poratingPPO(2,5-diphenyloxazole), drying,and then
being exposedto Fuji X-ray filmasdescribed inan
accompanyingpaper(6).
The isoelectricpoints of individual polypeptidesare
givento thenearest0.01 pH unit for convenience of
identification. The valueswere obtained by
extrapo-lation and do notrepresent measurements made to
thenearest0.01pHunit.
One-dimensional SDS-polyacrylamide gel
electrophoresis.This wasbased onthe second
di-mensiondescribed above and followedprocedures
de-scribed inanaccompanyingpaper(6),exceptthat the
reservoir bufferwastwice concentratedand the
acryl-amide-bisacrylamideratiowasincreasedto200:1from
75:1; 15%acrylamide gelswere run.The samples
con-tained ampholines as described for the
two-dimen-sional work andweremixed withanequal volume of
10%SDS-10% /3-mercaptoethanol-15%glycerol-0.02%
bromophenolbluein 1 MTris-hydrochloride,pH6.8.
lodination of FV3 particles with 125I using
chloramine T. This was performed exactly as
de-scribedbyMooreetal.(12).
RESULTS
Incorporation of [35S]methionine into
tri-chloroacetic acid-insoluble material. It has
been claimed that FV3 is apotent inhibitor of
protein synthesis in cells infected with live or
inactivated virus(3). Wewereunabletoconfirm these observations since the incorporation of [35S]methionine into trichloroacetic
acid-insolu-ble materialwasatanequivalentorhigherlevel
than control cells formost of the infection(Table 1). The data reported indicate the results
ob-tained with materialrunonboth the
one-dimen-sional and the two-dimenone-dimen-sional gels. Similar
re-sults have been obtained in numerous other
experiments.
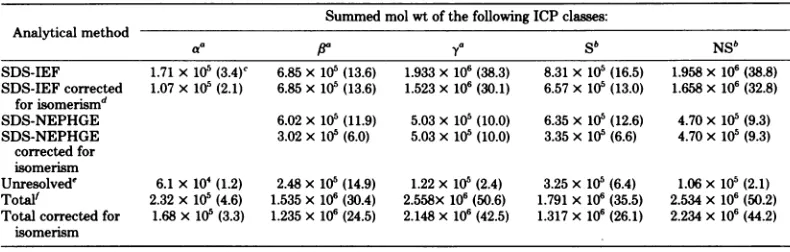
One-dimensional SDS-polyacrylamide gel electrophoresis of polypeptidesin
FV3-infected BHKcells. Thesequential induction
of FV3 ICPs on a gel system identical to the
two-dimensional gel electrophoretic second
di-mension is showninFig. 1;apolypeptideswere
detected from 2 h after infection, polypeptides
were detected from 4 h, and the onset of y
synthesis was at 6 h. In thisparticular
experi-mentthe and y phases were slightly late, so
the
,8
polypeptides predominated at6 h and the-ypolypeptides, whichwerebarelydetected at 6
h, predominated later. Basically the same
pat-ternofpolypeptideswasobservedonthosegels
as on ourstandardSDS-polyacrylamide gel
elec-trophoresis gels (6), although two additional bands are detected in purified virus
(polypep-tides X and88) whicharenotnormallydetected
and X isnotpresentas anICP.There arealso
"anomalies" in therunning of certain polypep-tides inthe molecularweightrangeof3x 104to
6 x 104. In these experiments the acrylamide-bisacrylamide ratiowasdifferent than that used
forourstandardconditions (6), and the samples
contained 4 M urea and 0.5% ampholines, so
some anomalies may have been due to these differences. The ICPsare labeled according to
the nomenclatureused inan accompanying pa-per(6).
Nomenclaturefortwo-dimensionalICPs.
The ICPs detected by IEF and
SDS-TABLE 1. IncorporationofL
-[35S]methionine
into trichloroacetic acid-insoluble material in FV3-infectedBHK cellsatvarioustimesafter infectionTrichloroacetic
acid-msolublecounts'
Control 45,529
FV3 2 55,018
FV3 4 41,647
FV3 6 54,543
FV3 9 44,879
FV3 12 42,748
FV3 24 46,997
aCountsperminuteobtainedafterprecipitationof
material in 6 x 104cells infectedat25 to30PFUof
virusper cell andpulse-radiolabeled with 5MCiof
L-[35S]methioninefor1h.Precipitationwasachievedby addinganequalvolume of ice-cold20%trichloroacetic acidandincubatingonicefor60min.Thecontrolwas
uninfected cells held at the infection temperature
(280C).
VOL. 33, 1980
on November 10, 2019 by guest
http://jvi.asm.org/
20 ELLIOTT, BRAVO, AND KELLY
tCP
U
2
4
6
9
12 24
FV3
VPiNS
_-'' '
''--14 98
88 ---78
r
SESfe
_-
---69 i ---4-4 4
[image:3.504.68.257.69.368.2] [image:3.504.270.462.285.610.2]dlii Sb
~~~~~~~~~~~~~~~~~~~.
-x ....-3--5 4
---34
}; -29
-27 -23
22
18
15 16
2
lb
a-FIG. 1. Sequential induction of ICPs in BHK cells
infected with FV3. Cells were pulse-labeled with
[35S/methioninefor1hatthetimes indicated across the top of thefigure (in hours). Purified FV3 was
co-run to indicate structural polypeptides. ICPs are
identified by estimates of their molecular weights
(xl103) (6)and are assignedasvirus particle (VP) or nonstructural(NS). U, Uninfected.
NEPHGEare shown in
Fig.
2andcataloged
inTable 2. The
polypeptides
are identifiedby
acombination of molecular
weight (to
thenearest1,000) andisoelectric point (to thenearest 0.01
pH unit), so that the
major
structuralpoly-peptide
ofFV3 is58,5.58 and themajor
nonstruc-tural basic
polypeptide
is35,7-10. ThepH
rangeis indicated for basic polypeptides because
nonequilibrium
electrophoresis
wasperformed
(14).
The various polypeptides were designated a,
fi,
and y forcomparison
with one-dimensionalgel polypeptides (6). The
assignment
was notrigorous because the
requisite
"function-block-ing" experiments had not been
performed
yet(6). For the purpose of thispaper, the timing
andapparentamountof
synthesis
wereused forassignment;
apolypeptides
were synthesizedfrom 2 h
(although
apparent isomers ofpoly-peptide 16,5.48weredetectedfrom4
h),
,8
poly-peptides were detected at 4 or 6 h and subse-quently synthesized in similar amounts, and y polypeptides were synthesized at 6 or 9 h and predominated late in infection.
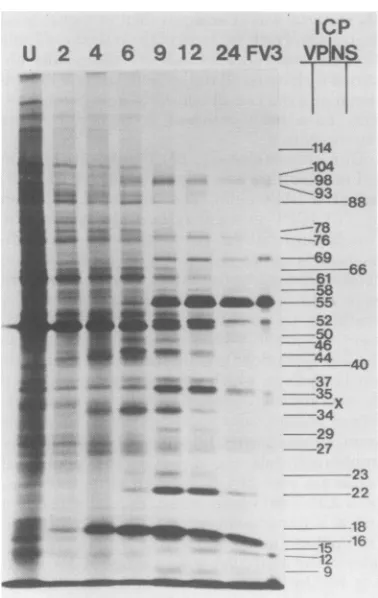
Acid IEF and SDS-acrylamide gel
elec-trophoresis of FV3 ICPs. Uninfected BHK
cells contained well in excess of 200polypeptides
resolvedby SDS-IEF (Fig. 3,gel i). By 2 h after
infection three ICPs were detected, all of which
were nonstructuralpolypeptides (Fig. 3, gel ii).
These three polypeptides (ICP 46,5.55, ICP
45,5.47, ICP 16,5.48) are by definition a
polypep-tides. Most cell polypeptides continued to be
synthesized,
albeit at a reduced rate, and somemajor cell
polypeptides were markedly reduced.At 4 h after infectionthese three a polypeptides
were synthesized in large amounts, and an
ad-ditional fourpresumptive isomers of polypeptide
16,5.48 were detected (Fig. 3, gel iii and Table
2). Seven additional polypeptides,
all
presump-pH
0
I-S 2
C:
4c
-J
Lu
-5
0
I
9
K114
T100 0
_
-76
2
0
.
Lum 4 38 D
35 a
0
2
-18 3
FIG. 2. Diagrammatic representation ofICPs de-tectedon(A) SDS-IEF and(B) SDS-NEPHGE
two-dimensionalgels. ICPs tentativelyassignedas
struc-turalpolypeptides are shown asclosed areas, and nonstructuralpolypeptidesareshownasopenareas
or areindicatedNS.
J. VIROL.
;,.h:-::%:- MOO*
:.4--s'. -Zx
32 .-!:;:,!!:.- .-
'--No
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 33, 1980 ANALYSIS OF FV3 POLYPEPTIDES
21
TABLE 2. Catalog ofICPsdetected inFV3-infectedcellsbytwo-dimensionalgelelectrophoresis
Structural Period Equivalent Structural Period Equivalent
Polypeptide or nonstruc- detected a,13'
one-dimen-
| Polypeptide ornonstruc- detected a,orone-dimen-turala (h) or-y sionalgel tural'~ (h) or-y sionalgel
I
I~__
Polypeptideit
J j polypeptideNS NS S NS S NS S NS NS NS S NS S S S S NS NS NS NS NS NS S NS S S NS NS NS NS NS NS NS S NS NS NS S NS NS NS NS NS NS 9-24 9-24 9-24 9-24 6-24 9-24 4-24 9-24 9-24 9-24 6-24 9-24 6-24 9-24 6-24 9-24 9-24 6-24 4-24 9-24 9-24 9-24 6-24 6-9 12-24 12-24 12-24 4-12 2-24 4-? 2-24 12-24 12-24 9-24 12-24 6-24 12-24 6-24 6-9 12-24 12-24 12-24 9-24 9-24 Y y Y y y y 0 y y y y y y y y y y y /3 y y y .7 y y Y a a y y y y p y y y y y y .7 ICP 76yVP orICP 78yVP ICP69,BVP ICP 55-yVP ICP52NVP ICP46NVP ICP40-yNS ICP 37yVP 29,4.14 29,4.18 29,4.46 27/28,4.50 24,4.76 24,4.88 24,6.37 23,6.46 22,4.76 22,4.88 22,5.23 22,6.53 18,5.91 17,7.00 16,4.931 16,5.22 16,5.30 16,5.48 16,5.70 16,7.00 15,4.86 15,6.52 15,6.80 14,4.14 14,4.57 13,6.25 Basic 115,7-10' 114,7-10 100,7-10 100,7-lObi 100,7-lObii 100,7-10biii 76,7-10 67,7-10 55,7-10 47,7-10 38,7-10 38,7-lOb 35,7-10 24,7-10 24,7-lOb 18,7-10 15,7-10 14,7-10 13,7-10 12,7-10 NS S? S NS NS NS NS NS NS NS NS NS NS NS NS NS NS NS NS S NS NS NS NS S S S NS S S S S NS NS NS NS S NS NS NS S S S NS S S 9-24 9-24 6-24 4-9 6-24 6-24 6-24 6-24 6-24 6-24 6-24 6-24 6-24 6-24 4-24 4-24 4-24 2-24 4-24 6-24 4-9 4-24 6-24 9-24 6-24 12-24 9-24 12-24 6-24 6-24 6-24 6-24 9-24 4-24 9-24 9-24 6-24 4-24 4-24 9-24 6-24 9-24 9-24 12-24 9-24 6-24 Y Y /3 1 /3 / /3 / 13 1 Y / 1 1 a a a a a / 13 / Y Y Y Y Y Y .7 0 0 y /3 y Y /3 p Y Y 0 y y Y y Y I Y3 Y3 Y7 ICP27yVP ICP29,BVP ICP23,8NS ICP23,BNS ICP23,BNS ICP2313NS ICP22yNS ICP 16aNS ICP 12yVP ICP 114yVP ICP98,BVP ICP66,8NS ICP44,/VP? ICP35flNSd ICP12yVP Acidic 86,5.76b 82,5.76 76,6.85 72,5.53 72,5.63 72,5.73 69,6.93 59,5.93j 59,6.11 59,6.22 59,6.35 59,6.5U 58,5.411 58,5.48 58,5.58 58,5.69J 58,6.99 55,5.39 55,6.93 54,5.82 54,5.94 54,6.12 52,5.39 49,4.76 49,5.13 49,5.23 49,4.35 47,5.45 46,5.55 46,5.63 45,5.47 43,5.60 41,5.57 38,5.30 38,5.56 35,4.57 34,5.56 34,6.56 34,6.70 33,5.35 33,5.65 32,5.53 31,4.64 31,5.35
S,Structural; NS,nonstructural.The brackets onthetableindicateapparentcharge isomers.
b For the acidicpolypeptidesthefirst number isthe molecularweight
(x103),
andthe second number is the isoelectric point(pH).
Forthebasicpolypeptidesthe firstnumber is themolecularweight(XI03),and the second numberis thepHrangeresolved.
dSeetext.
tive
f polypeptides,
wereobserved, including
the anumber ofadditional
ypolypeptideswere de-structural polypeptide 69,6.93. Little further tected (Table 2). Minor yICPsweredetected atsuppression of cellular synthesisoccurred.By6 12and 24 h(Fig.3,gels vi and vii). At 24 h after
h ,Bpolypeptide synthesis waswell under way, infection,anumberof /3and y polypeptides had
and an additional 15
,B
ICPsweredetected(Fig.declined
in synthesis, although the main virus3,gelivand Table 2).At 6 height-ypolypeptides structural polypeptides werestillsynthesized in
were detected, includingthe major virusstruc- quantity (Fig. 3, gel vii). About 70 ICPs were
tural
polypeptide
58,5.58.By 9h (Fig. 3, gelv) resolved during infection.these
polypeptides
became the prominent -y The structural polypeptides contained in FV3polypeptides synthesized, althoughatthis time and resolvedby SDS-IEF are shown in Fig. 3,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.56.447.80.513.2]22 ELLIOTT, BRAVO, AND KELLY
4
MW
pH 5
tni7- T'',
76-58- 2
34-
24-
16-
76-4w._
58-
34-"':
24-J. VIROL.
pH
*-_tM #* -16
16-
16-
_
~~~~~~~~~~~~~~~...
'-,,.FIG. 3. Distribution of[35S]methionine-labeled polypeptides in FV3-infected BHK cells analyzed by SDS-IEF.Gel i, Uninfected cells radiolabeled for I hat28°C; gel ii, infected cells radiolabeledat2to3h; gel iii, infected cells radiolabeled at4to 5h; gel iv, infectedcellsradiolabeled at6to 7h; gelv, infected cells
radiolabeledat9to10h; gel vi, infected cells radiolabeledat12to13h; gel vii, infected cells radiolabeledat 24 to25h; gelviii, purified[35S]methionine-labeled FV3 structural polypeptides. The ordinaterepresents
molecularweight (MW)(xl3).
gel viii. Mostpolypeptides detected in virus
par-ticles possessed ICP counterparts, although a
few hadnoobviousICPorcellularpolypeptide
counterpart.Virusparticles presented consistent
profiles, and so this discrepancy may be due
eithertoenrichment ofminorinfectedcell
pro-teins in the virus particles or to processing on
encapsidation and release.
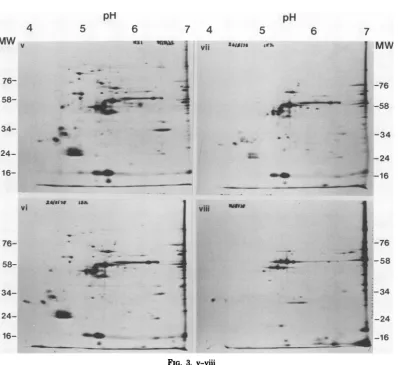
Basic NEPHGE and SDS-acrylamide gel electrophoresis ofFV3 ICPs. In contrast to
the acidicsystem,fewICPsweredetected in the
basicpH7 to pH 10 analysis. The 20 polypep-tides detected are shown diagrammatically in
Fig.2 andsequentially inFig.4. Asummaryof
theirproperties isgiveninTable 2.
NoICPsweredetected2 hafterinfection(Fig.
4, gel ii), and therewasnotamarked alteration
in the synthesis of cell polypeptides, although the synthesis of certain cell polypeptides was
obviously becoming suppressed. At4h the
syn-thesis of three nonstructural ,B polypeptides (ICP 67,7-10; ICP 38,7-lOb; ICP 35,7-10) was
detected,and by 6 hanadditional four structural
B8polypeptidesweredetected (Fig. 4, gels iii and
iv), which increased in relative amountof
syn-thesisat9 h(Fig. 4, gel v). At 6 honestructural
ypolypeptide (ICP 12,7-10) wasdetected.
Sub-sequentlyat12and 24 hlittlealteration in ICP synthesis occurred, although cell polypeptide
synthesiswasfurther suppressed.
Identification of structuraland nonstructural polypeptides by SDS-NEPHGEwas aproblem.
6 7 4
iii
6
14 im
iv IAf j~ 'a'!
VW 4
0-MW
-76.
-58
-34
-24
-16
-76
-58
-34
..-24
I
:
;..4
on November 10, 2019 by guest
http://jvi.asm.org/
ANALYSIS OF FV3 POLYPEPTIDES 23
pH
5
"Isan if,.
6 7
MW
-76
-58
-34
-24
-16
-76
-58
2-34
-24 4
uWO
4
4
[image:6.504.53.451.64.429.2]* -16
...
--[35S]methionine-labeledFV3particlesdisrupted with lysis bufferor lysis buffer plus0.1% SDS
failedtoproduce reproducible patterns.Purified
FV3 subsequently radiolabeled with
"25I
using chloramine T demonstrated that three major polypeptides detected late in infection werebasic structural polypeptides. Considerably more '25I-labeled polypeptides were present in
virus particles than were observed as ICPs in
infected cells late in infection (and in
SDS-NEPHGEresolving purified
[35S]methionine-la-beledvirus), and thisweascribein part to
chlor-amineT-inducedchargealterations in the poly-peptidesand in part todifferences inthe methi-onineandtyrosinecontentsof thepolypeptides. There is also the possibility ofunlabeled
con-taminating host proteins becoming iodinated by chloramineT.
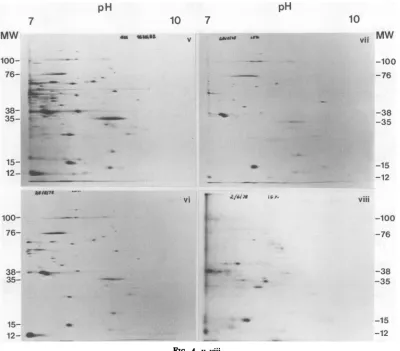
Comparison of SDS-IEF and SDS-NEPHGE. The ICPs detected over the pH range4 to10areshowndiagrammaticallyinFig.
2 and in Fig. 5. A total of 70 acidic and 20basic
polypeptidesweredetected.Thetotalmolecular
weight of these polypeptides was 3.89 x
106,
whichrepresents77% of the coding capacityof theFV3genome (Table 3).
A number of ICPs detected by
one-dimen-sional SDS-PAGE analysis appearednot to be
detected by SDS-IEF and
SDS-NEPHGE.
ThesefrogFV3 ICPsincludedICPs104,93,88, 61, 58, 18, and 9 (whichare all minora, fi, and
ypolypeptides whichco-electrophoresewith
cel-lularpolypeptides) and ICP 34, amajor
struc-tural
ft
polypeptide. With the exceptionofICP34, thesepolypeptides eithermayhavean
isoe-lectric point of pH 7.00 andso arenotdetected
atthe first-dimension gel origin (pH 7.0) above material which hasnotentered the gel ormay
haveisoelectric pointsoutside the pHrange4 to
10. The total molecular weightofthe ICPs
de-tected bytwo-dimensional gelanalysis andthe unresolved polypeptides (with the exception of
pH
5 6 7 4
4
MW
v
76-
58-
34-vii
....
....IL
*0%~ ~ ~ ~ ,
..k
4'
24-
16-vi
76-
58--._ ,@ - .
* II
viii
34-
24-16-
Ab4
.-FIG. 3. v-Vini VOL. 33, 1980
- %4
...
--A-.
on November 10, 2019 by guest
http://jvi.asm.org/
24 ELLIOTT, BRAVO, AND KELLY
pH
10 7
i I4mor"
am 4W
iii MW
--loc' -76
-38
-35
-15 _---12
ii iv
-100
-76
...
W..-.N
l_..4.
_ .,A W
.
....
o 1. - .,.
I' B. M ~
:
. _ -38
-35
-15
-12
FIG. 4. Distributionof[35S]methionine-labeledpolypeptidesinFV3-infected BHK cells analyzed by SDS-NEPHGE. Geli, Uninfectedcellsradiolabeledfor1hat28°C; gel ii,infected cells radiolabeledat2to3h;
gel iii, infectedcells radiolabeledat4to5h;gel iv,infectedcellsradiolabeledat6to 7h; gelv,infected cells
radiolabeledat9to10h; gel vi,infectedcellsradiolabeledat12to13h;gel vii, infected cells radiolabeledat 24to 25h; gel viii,purified FV3 structuralpolypeptides 125I iodinatedusingchloramine T. The ordinate representsmolecularweight (MW) (x103).
ICP 34 [see below]) was 4.325 x 106, which is
equivalentto85.6% of the virusgenome (Table
3).
Effect of FV3oncellularpolypeptide
syn-thesis. Over400polypeptidesweredetected in
uninfected BHK cellsandaretoonumerousto
realistically andprofitably catalog. At 24 h after infection 50 cell-specific polypeptides were
de-tected. Since FV3hasnomarked effectontotal
protein synthesis in BHK cells (Table 1), it is obviousthatthe virussubverts thecapacity of
thecellprotein-synthesizingsystems to
synthe-size itsownvirus-specific proteins. Thisappears
to occur by suppressing cell protein synthesis
ratherthanby actively switching offcell
synthe-sis. Most cell polypeptides appear to become gradually undetectable with respect to time,
which isindicative ofsuccessfulcompetition of virussynthesis with cell synthesis. Certain poly-peptides do, however, declinerapidly in synthe-sis(e.g.,polypeptidesatposition 60,5.01 and four polypeptides withintherange80 to 100,4to5).
DISCUSSION
Ourresults demonstrate thata high
propor-tion of the FV3 genome appears to be
tran-scribed and translated into presumptive virus-specific proteins during a productive infection.
Theresults also showthattwo-dimensionalgel electrophoretic analysis ofFV3 ICPs is vastly superiortoconventional one-dimensional anal-ysis and that it could be the method of choice
for the futureanalysis ofFV3polypeptide
me-tabolism.
7
MWr
100-
76- 38-
35-zF. ...
15- W
12- 0
100-
76- 38-
35-
15-
12--J. VIROL.
W..!:".4..,::!"
jow., :X....b,.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.70.460.72.417.2]ANALYSIS OF FV3 POLYPEPTIDES
7
MW
100-76-;
38-35- t
*
12.
12* ...
100- .
76-.e
38-
-
35- 15-12-
O-ab
10
7
V EaS* ,
pH
10
..
MW
-100 -76
-38 -35
Theresultsobtainedontwo-dimensional gels
confirm that there are atleast three temporal
phases ofpolypeptide synthesis in FV3-infected BHKcells, substantiatingouraccompanying
ob-servations(6).This is atvariance with thereport by Granoff (9) that all FV3ICPsaresynthesized
from the initiation of infection
(although
therearesubsets ofICPsdistinguished by theirrate
ofsynthesis [15]). Autoradiographic
overexpo-sure of the gels shown in this paper failed to
demonstrate any of the /8 or y ICPs at times
earlierthanthosereported (unpublished data),
and so we are confident that there was not a
detectionproblem.
Theseparation of FV3ICPsaccordingtotheir isoelectric pointsintheIEF and NEPHGE di-mensions allowedacatalog of isoelectricpoints
tobecompiled (Table 2), and this is particularly useful in the IEF separation. Our information showsthatthebulk of theFV3ICPshave isoe-lectric points in the pH 4to 7range, including
the major structuralpolypeptide (ICP 58,5.48). Observations on the major structural
poly-peptide showthatthere is apparent isomerism ofthispolypeptide withpolypeptidesofdifferent charges but similar molecularweights appearing in the cell. Thispolypeptide is phosphorylated butisnotdetectably sulfatedorglycosylated (6),
and so some isomerism may be attributed to
phosphate-created charge differences. Not all
phosphorylated ICPsshowisomerism (e.g.,ICP
69,6.93) andso someisomerism maybe caused
byas-yet-unidentified posttranslational
modifi-cation. Anumber of other polypeptidesappear
to show charge isomerism and are shown in
Table 2.
Onone-dimensionalgelsICP34(a majorICP)
appears to be a structural polypeptide,
al-thoughontwo-dimensionalgels nocounterpart
is found. Acomparison of IEF and SDS-NEPHGEindicatesthatamajorftnonstructural
polypeptide (ICP 34,7-10) is found on
SDS-pH
_ Isas
e
.a
Vi .1/6/f ; J.
-15 -12
_K:
E...
Viii
-100
-76
-38 -35
a4
[image:8.504.50.450.76.427.2]i
FIG. 4. v-vuii
-15
-12
25
VOL. 33, 1980
L-
... .won November 10, 2019 by guest
http://jvi.asm.org/
26 ELLIOTT, BRAVO, AND KELLY
4
MW 1
ab .
76-58- - ._
34- r
24-
16-Iv
-100
-76
-38 -35
-15
-12
S.A. . ....-.
FIG. 5. Distribution oftotal[35S]methionine-labeledpolypeptidesinFV3-infectedBHK cells;acomparison
of SDS-IEF and SDS-NEPHGEat4and12hafter infection.Gels i and iii, SDS-IEF and SDS-NEPHGE,
respectively, 4h; gel ii and iv, SDS-IEF and SDS-NEPHGE, respectively, 12 h. The ordinate represents
molecularweight (MW) (x103).
NEPHGEgels from 4to24hand thataminor
y structural polypeptide (ICP 34,6.56) occurs
from 6 to 24 h on SDS-IEF gels; therefore, it
appears that ICP 34 resolved on
one-dimen-sionalgelsrepresents twopolypeptides and that the structuralcomponentof the virus particle is
anacidicypolypeptide.
Early in infectionnobasicapolypeptidesare
detected, which may be considered surprising
since thevirusappearstohaveaneffectonthe
expression ofthe cell genome,and the interac-tion of basicproteins with DNAorRNAorboth
may provide a mechanism of inhibition. The
absence ofproteins with basic isoelectric points doesnotpreclude the possibility thatsomeother
ICPs have basic regions capable of such inter-action.
Ourdata indicate that reasonable quantities of FV3 (about 30 PFU/cell, 20 virus particles
perPFU) failtocausecellpolypeptide synthesis
to be switched offrapidly, although there is a
gradual suppression of cell polypeptide synthesis
asthe infectionprogresses.Thiscontradicts
ear-lierpublishedwork(8),but is inagreementwith
observations ofJ.C. S. Clegg and S. I. T. Ken-nedy (personal communication) and R. W. Schlesinger (personal communication). FV3, like herpesvirus (7), undoubtedly possesses
struc-turalpolypeptides which can inhibit cell RNA
and protein synthesis, as elegantly shown by
pH
7
J. VIROL.
_pu, w0
10
...
MW
III
-100 -76
40-, .:...
t
ii
-38 -35
-ao
*w .
-15 -12
76-
58-34-. w
.m
24--
16-I
wgl.
jr..%-,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.504.73.463.74.432.2]ANALYSIS OF FV3 POLYPEPTIDES 27
TABLE 3. Total molecularweights ofFV3-induced classesofICPs detectedbyvariousgel analytical
methods and the percentagesofthe FV3 genomerequiredfortheirsyntheses
Summedmol wt of thefollowingICPclasses:
Analytical method
,a a ya Sb NSb
SDS-IEF 1.71 x105(3.4)c 6.85x105 (13.6) 1.933x106(38.3) 8.31 x105(16.5) 1.958 x10W (38.8) SDS-IEF corrected 1.07x105(2.1) 6.85x105 (13.6) 1.523x10W (30.1) 6.57x105(13.0) 1.658x10W (32.8)
for isomerismd
SDS-NEPHGE 6.02 x105 (11.9) 5.03 x105(10.0) 6.35 x105(12.6) 4.70x 105(9.3) SDS-NEPHGE 3.02 x105(6.0) 5.03 x105(10.0) 3.35 x105(6.6) 4.70 x105(9.3)
corrected for isomerism
Unresolved' 6.1x104(1.2) 2.48x105(14.9) 1.22 x105(2.4) 3.25x 105(6.4) 1.06 x105(2.1)
Totalf 2.32 xi05(4.6) 1.535 x10"(30.4) 2.558x106(50.6) 1.791 x10W (35.5) 2.534x106(50.2)
Total corrected for 1.68 x105 (3.3) 1.235x106(24.5) 2.148x106 (42.5) 1.317 x106 (26.1) 2.234x106(44.2) isomerism
aa,
fi,
andy refertothetemporalclasses ofICPs resolved (6,7).bSStructuralpolypeptides; NS,nonstructuralpolypeptides.
'Thenumbers in parentheses are the percentages of the FV3 genome(molecular weight, 1x 108[12]) responsiblefor the synthesis ofindividual classes ofICPs, assuming that the whole genome is available for translation and transcription and that no overlapping genes occur.
d Forcorrections forpossible isomerism,see textand Table 2.
'Unresolvedpolypeptides are ICPs which are resolved on one-dimensional gels but havenotwo-dimensional counterparts (see text).
fThe total molecularweight for a plus f plus y polypeptides was 4.325x 106or85.6% of the FV3 genome(3.551x106and 70.3%correctedforisomerism). Forstructuralplusnonstructuralpolypeptidesthemolecularweight totalswerethesame as
those for aplus fiplusypolypeptides.
Aubertinetal. (1), but it is doubtful that
suffi-cientquantitiesof theseproteinsareintroduced
into cells undernormalphysiologicalconditions tosuppresshostpolypeptide synthesis
dramati-cally.
ACKNOWLEDGMENTS
Wethank J. S.Knowland for his interest in thesestudies.
R.M.E. received a Science Research Council postgraduate studentship. R.B. thanks the Comision Nacional de Investi-gacionCientificayTechnologica deChile, the Royal Society
of London for an Exchange Fellowship, and the Medical ResearchCouncil forfinancial assistance.
LITERATURE CITED
1. Aubertin, A. M., C. Hirth, C. Travo, H. Nonemacher, andA.Kirn.1973.Preparation and properties ofan inhibitoryextractfrom frog virus3particles.J.Virol. 11:694-701.
2. Bonner, W.M., and R. A. Laskey.1974. Afilmdetection
method fortritium labeled proteins andnucleic acids in
polyacrylamide gels. Eur. J. Biochem.46:83-88. 3. Drillien, R.,D.Sephner, and A.Ki-n.1977.Cell killing
by frog virus3:evidence forcellkillingby single viral
particlesorsingle viral units. Biochem. Biophys. Res. Commun.79:105-111.
4. Elliott,R.M., M. K. Arnold, and D. C.Kelly. 1979. The
replication of frog virus 3 inan amphibian cell line
(XTC-2) derived fromXenopus laevis. J. Gen. Virol.
44:89-98.
5. Elliott, R. M., A. Bateson, and D. C. Kelly. 1980.
Phosphonoacetic acid inhibition offrogvirus 3 replica-tion. J. Virol. 33:539-542.
6. Elliott, R. M., and D. C. Kelly. 1980. Frog virus 3
replication: induction and intracellular distribution of
polypeptides in infected cells. J. Virol. 33:28-51.
7. Fenwick,M.L.,and M. J. Walker. 1978.Suppression
and synthesis ofcellular macromolecules by herpes simplex virus. J. Gen. Virol. 41:37-51.
8. Goorha, R., D.Willis, and A. Granoff. 1977. Macro-molecularsynthesisin cellsinfected withfrog virus 3. VI. Frog virus3replication is dependent on thecell nucleus. J. Virol.21:802-805.
9. Granoff,A.1969.Viruses ofamphibian.Curr.Top.
Mi-crobiol. Immunol. 50:107-137.
10. Kelly, D.C.,and R. J.Avery.1974.Frogvirus3 deoxy-ribonucleicacid. J. Gen. Virol. 24:339-348.
11.Kelly, D. C.,and J. S. Robertson. 1973. Icosahedral
cytoplasmicdeoxyriboviruses.J. Gen. Virol. 20:17-41.
12.Moore, N.F.,J. M.Kelley,and R. R.Wagner. 1974.
Envelopeproteinsof vesicular stomatitis virions:
acces-sibilitytoiodination.Virology61:292-296.
13.O'Farrell, P. H. 1975.Highresolutiontwo-dimensional
electrophoresis ofproteins.J. Biol. Chem.
250:4007-4021.
14. O'Farrell,P.Z.,H. M.Goodman,and P. H.O'Farrell.
1977.Highresolution two dimensionalelectrophoresis of basicaswellasacidicproteins. Cell 12:1133-1142.
15. Willis,D.B., R.Goorha,M.Miles, and A.Granoff.
1977.Macromolecularsynthesis in cells infected by frog
virus 3. VII. Transcriptional and posttranscriptional
regulation of virus gene expression. J. Virol.
24:326-342.
VOL. 33, 1980

![FIG. 4.gelrepresents24NEPHGE.radiolabeled to Distribution of [35S]methionine-labeled polypeptides in FV3-infected BHK cells analyzed by SDS- Gel i, Uninfected cells radiolabeled for 1 h at 28°C; gel ii, infected cells radiolabeled at 2 to 3 h; iii, infecte](https://thumb-us.123doks.com/thumbv2/123dok_us/1500608.102791/7.504.70.460.72.417/gelrepresents-radiolabeled-distribution-methionine-polypeptides-uninfected-radiolabeled-radiolabeled.webp)
![FIG. 5.molecularofrespectively, SDS-IEF Distribution oftotal [35S]methionine-labeledpolypeptides in FV3-infected BHK cells; a comparison and SDS-NEPHGE at 4 and 12 h after infection](https://thumb-us.123doks.com/thumbv2/123dok_us/1500608.102791/9.504.73.463.74.432/molecularofrespectively-distribution-methionine-labeledpolypeptides-infected-comparison-nephge-infection.webp)