Tethering by EBNA1 to Host Chromosomes
Theresa L. Hodin, Tanbir Najrana, John L. Yates
Roswell Park Cancer Institute, Department of Cancer Genetics, Buffalo, New York, USA
The EBNA1 protein of Epstein-Barr virus enables plasmids carryingoriPboth to duplicate and to segregate efficiently in prolif-erating cells. EBNA1 recruits the origin recognition complex (ORC) to establish a replication origin at one element oforiP, DS (dyadsymmetry); at another element, FR (family ofrepeats), EBNA1 binds to an array of sites from which it tethers plasmids to host chromosomes for mitotic stability. We report experiments leading to the conclusion that tethering by EBNA1 to host chro-mosomes is also needed within interphase nuclei in order for plasmids to be replicated efficiently fromoriP. The DNA-binding domain of EBNA1, which lacks chromosome-binding ability, was found to support weak, DS-specific replication in HEK293 cells after transient transfection, being 17% as active as wild-type EBNA1. The low efficiency of replication was not due to the failure of the DNA-binding domain to retain plasmids within nuclei, because plasmids were recovered in similar amounts and entirely from the nuclear fraction of these transiently transfected cells. A derivative of EBNA1 with its chromosome-tethering domains replaced by a 22-amino-acid nucleosome-binding domain was fully active in supportingoriPfunctions. The implication is that EBNA1’s DNA-binding domain is able to recruit ORC to DS, but either this step or subsequent replication is only efficient if the plasmid is tethered to a host chromosome. Finally, with some cell lines, DS can hardly support even transient plasmid replica-tion without FR. A loss of plasmids lacking FR from nuclei cannot account for this requirement, suggesting that the stronger tethering to chromosomes by FR is needed for plasmid replication within the nuclei of such cells.
T
wo distantly related gammaherpesviruses, Epstein-Barr virus (EBV) and Kaposi’s sarcoma-associated herpesvirus (KSHV), are each associated with diverse human tumors, infect cells la-tently as part of their normal life cycles, and can drive these cells to proliferate (1,2). In latent infections, the circularized viral chro-mosomes are replicated during S phase of the cell cycle and asso-ciate with condensed human chromosomes through mitosis to ensure safe passage of viral chromosomes to daughter nuclei. A single virus-encoded protein, EBNA1 of EBV or LANA of KSHV, forms a tether holding the viral chromosome to human chromo-somes. EBNA1 and LANA also recruit the cellular origin recogni-tion complex, ORC, to specific sites on their respective viral chro-mosome where replication may initiate during S phase (3, 4). EBNA1 and LANA have no sequence homology and at most share a very distant common ancestor for their DNA-binding domains (5). EBNA1 and LANA bind to unrelated sequences, they bind to chromosomes using different mechanisms (6,7), and they may recruit ORC differently as well. This parallel evolution raises an interesting question: why for each virus did a single protein evolve to perform two functions that lack any obvious connection, de-fining a replication origin by recruiting ORC and tethering to human chromosomes for mitotic segregation?It has long been suspected that plasmids derived from EBV and KSHV need to be tethered to human chromosomes in the nucleus during S phase in order to replicate efficiently, because removal of the chromosome-binding domains from EBNA1 or LANA greatly reduces the accumulation of replicated plasmids in transiently transfected cells. It is important to know whether such a func-tional relationship truly exists, however, or is merely apparent. Tethered plasmids are recovered at higher levels from transfected cells in large part because they survive better, an effect that is obvious in several published experiments but seldom acknowl-edged. In addition, untethered plasmids would have less opportu-nity to be replicated than tethered plasmids if they have spent less
time within nuclei—an issue that has not been addressed. The published evidence for EBV is discussed below (for KSHV, see Discussion).
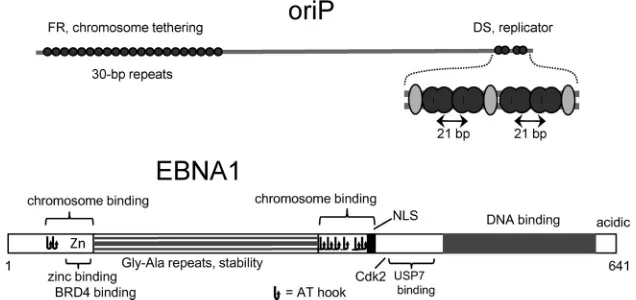
EBNA1 acts at a bipartite locus,oriP, to maintain EBV chro-mosomes in mitotically active, latently infected cells (Fig. 1) (for a review, see reference4). By binding to FR (family ofrepeats) of
oriP, an array of⬃20 high-affinity EBNA1-binding sites, EBNA1 confers mitotic stability to plasmids (8) by tethering them to mi-totic chromosomes (7,9,10). The other component oforiP, called DS (dyadsymmetry), functions as an EBNA1-dependent replica-tor (11–13), which although essential for small plasmids, is redun-dant for the EBV chromosome (14). With four EBNA1-binding sites, DS has only weak tethering ability (13). What defines DS as a replicator is a 21-bp center-to-center spacing within pairs of EBNA1-binding sites (15,16), although binding by TRF2 (t elo-mererepeatfactor2) at three sites surrounding the pairs of EBNA1 sites plays an ancillary role (17,18). While the C-terminal DNA-binding domain binds to sites atoriP, two N-terminal domains called LR1 and LR2 (linkingregions1and2) bind to mitotic chro-mosomes (7,19). Both LR1 and LR2 have AT hook motifs and activity for binding AT-rich DNA (20), although the region of LR1 that binds zinc and the cellular factor bdr4 (21,22) also contrib-utes to chromosome binding (7). The arginine-rich regions with AT hook activity also bind RNA, and it has been proposed that binding to RNA mediates both chromosome tethering and re-cruitment of ORC (23,24).
Received13 June 2013 Accepted19 September 2013
Published ahead of print25 September 2013
Address correspondence to John L. Yates, [email protected].
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.01606-13
on November 7, 2019 by guest
http://jvi.asm.org/
Four published studies have correlated deletions of LR1 and LR2 from EBNA1 with a loss of the capacity to support short-term replication of plasmids carryingoriP(10, 19, 25,26). Three of these show Southern assays, from which we may judge the effect of chromosome tethering on the amount of plasmid recovered, in addition to how well the plasmids replicated. In all three studies, a loss of plasmids in the absence of tethering can account for much of the reduction in replicated plasmid, though not all of it: after 4 days in HeLa cells (see Fig. Fig. 7 in reference10), after 3 days in C33A cells (see Fig. 4 in reference26), and after 4 days in HEK293 cells (see Fig. 5 in reference19). In two of the studies, some plas-mid replication was seen in the absence of any tethering potential (10,26), and in the third, high background hybridization might have obscured a modest amount of replication by the poorly re-tained plasmid (19). The results suggest that replication might not be entirely dependent upon tethering, at least in some cell lines. If the transfected plasmids that survived in the cells up to the time of harvesting spent less time within nuclei, a possibility that was not investigated, then this combined with the reduced survival of un-tethered plasmids might account for the full reduction of mea-sured replication. If so, then chromosome tethering need play no real role in plasmid replicationper se.
A lack of clarity on this issue has confounded studies of another aspect of EBNA1’s support oforiPfunction, namely, how EBNA1 recruits ORC to DS, which is not yet understood. Often it has been assumed that N-terminal domains of EBNA1, specifically LR1 and LR2, recruit ORC to DS (e.g., see reference
24). It seemed to us that this too might be wrong for the reasons mentioned above and for other reasons. This led us to perform cleaner and shorter assays for transient plasmid replication that minimize the contribution of tethering to plasmid survival, to examine the distribution of plasmids between nuclear and cy-toplasmic fractions after transfection and to test whether a sim-ple, well-defined chromosome-binding domain could func-tionally replace EBNA1’s N-terminal domains. The results lead to the intriguing conclusion that plasmids must tether to chro-mosomes within interphase nuclei to be replicated efficiently and that the only function of EBNA1’s N-terminal domains that is critical to replication is tethering to chromosomes.
MATERIALS AND METHODS
Cell lines, plasmids, and EBNA1 mutants.HEK293 and 143B cells are available from the American Type Culture Collection, Atlanta, GA, al-though the cells used here, obtained from Bill Sugden, grow and transfect somewhat differently from the same lines obtained from the repository. EBNA1-expressing 143B cells are 143B/SVoB-H2.9 clone 4 (27). All se-quences of plasmids and the primers used for their construction will be made available on request. pHEBo, carryingoriPand thehph (hygromy-cin B phosphotransferase) gene linked to the herpes simplex virus thymi-dine kinase (HSVtk) promoter and its derivatives lacking DS or FR have been described previously (16,28). p803 is pHEBo carrying the EBNA1 gene expressed from the Rous sarcoma virus LTR; it is identical to p367 (29), except that codons 382 and 383 of the EBNA1 gene were changed to synonymous codons to make a BglII site. Mutations in EBNA1 between codons 16 and 376 were introduced using specific PCR primers that in-cluded the AvrII site at codons 13 to 15 and the BglII site. EBNA1 mutant dn1160 was originally called N⌬450 – 641 (30). A plasmid expressing dn1160 (p1160), a comparable plasmid expressing wild-type EBNA1 (p1553), and plasmids encoding the EBNA1 deletion of residues 64 to 89 (p3015) and EBNA1 with randomized codons 394 to 453 (p2774) (31) were obtained from Bill Sugden. Mutant EBNA1 proteins could be tested easily for expression after transient transfection of 293T cells, because the plasmid vector contains the simian virus 40 (SV40) replication origin. EBNA1 was detected by immunoblotting using rabbit IgG against the EBNA1 DNA-binding domain (Cyto-Barr, Zuidhorn, The Netherlands) and peroxidase-conjugated goat anti-rabbit IgG and chemiluminescent detection (Pierce.).
Plasmid replication assays.Adherent cell lines were grown in Iscove’s modified Dulbecco’s medium (IMDM) with 9% fetal bovine serum and gentamicin, all from Life Technologies (Invitrogen). Cells were trans-fected in 6-cm dishes by the calcium phosphate method as described pre-viously (16) with 2.5g DNA per dish when using a single plasmid or 2g of a plasmid withoriPcomponents plus 0.75g of a plasmid to express EBNA1. The cells were shocked with glycerol at a concentration of 20% (143B) or 10% (HEK293) in complete medium for 2.5 min at room tem-perature after 4.5 to 5.5 h of incubation with the calcium phosphate-DNA coprecipitates. Transient transfections were considered to begin with the glycerol shock, since most of the DNA uptake occurs at this point (32). Cells were removed from plates the next day using trypsin, suspended in 4 ml of phosphate-buffered saline (PBS) containing 1 mM EDTA and 1% serum, pelleted at 200⫻g, and then washed again in PBS-EDTA without serum. Cells from one 6-cm plate were transferred to two 10-cm plates, which were combined later for the recovery of plasmids using the alkaline FIG 1Features oforiPand EBNA1. Above,oriPspans⬃1,800 bp, with two functional components, FR and DS. Sites of EBNA1 binding are shown as filled black circles. DS is shown expanded, with dark double ovals representing dimers of the EBNA1 DNA-binding domain at the four binding sites and lighter ovals representing TRF2 molecules. Below are shown features of the 641-amino-acid EBNA1 protein (see the text). NLS, nuclear localization signal (54). Each nonoverlapping occurrence of RGR is indicated as a potential AT hook.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:2.585.133.451.68.218.2]lysis method, as previously described (16). One-quarter of the recovered plasmids was analyzed for resistance to DpnI as described previously (16). To select cells stably carrying plasmids, one-tenth of the transfected cells were transferred to 6-cm dishes the day after transfection, and the following day, hygromycin B (Sigma-Aldrich) was added to the growth medium at nominal concentrations of 250g/ml initially, raised to 280
g/ml after 3 days, and raised to 350g/ml after 3 more days. (The sup-plier listed the active drug concentration as⫾20%, so the true concentra-tion was not known.) This gradual selecconcentra-tion eliminated any viable cells from untransfected control plates by 7 to 8 days, while allowing the colo-nies that grew more slowly with certain plasmids to expand. The colocolo-nies carrying p803 (functionally wild-type EBNA1) covered the surface of the dishes after 1 week of selection; these were kept in the same dishes with frequent changes of medium for another 6 days while waiting for colonies in the other plates to expand. After 13 days of selection, a portion of cells from each plate was transferred to 10-cm plates, grown without selection until the plates were full, and harvested by alkaline lysis 17 to 20 days after transfection. One-fourth of each sample was run on a 0.7% agarose gel, which was stained with ethidium bromide after electrophoresis.
Recovery of plasmids from cytoplasmic and nuclear fractions. Cy-toplasmic and nuclear fractions were prepared using two methods. For method 1, cells grown in two 10-cm plates were lysed and rinsed off the plates using 2 ml per plate of ice-cold high-sucrose buffer A (320 mM sucrose, 5 mM MgCl2, 10 mM Tris HCl [pH 8.0]) containing 1% (vol/vol)
Triton X-100, vortexed at high speed for 5 min in a 4°C cold room, and centrifuged at 260⫻gfor 5 min at 4°C in a swing-out rotor. The super-natant (cytoplasmic) and pellet (nuclear) fractions were then processed as described below. Method 2 followed the procedure of Belgrader et al. (33). Cells were harvested as described above using iso-high-pH buffer (140 mM NaCl, 1.5 mM MgCl2, 10 mM Tris-HCl [pH 8.4]) containing 0.5%
(vol/vol) Nonidet P-40. The nuclear pellets were then washed twice in the same solution, and the nuclei were suspended in 200l of iso-high-pH buffer containing 0.33% sodium and 0.66% Tween 80 and sedimented through a sucrose cushion as described previously (33). The cytoplasmic fractions were adjusted to 200 mM NaCl–5 mM EDTA, treated with 4 ml of a 1:1 mixture of phenol and chloroform, and then combined with an equal volume of isopropanol to precipitate nucleic acids. The precipitates were dissolved in 0.4 ml water, combined with 16l of 5 M NaCl, treated with 200l of phenol-chloroform, and precipitated with 0.8 ml ethanol. Pellets were rinsed with 70% ethanol, dried, and dissolved in 50l of water. The nuclear pellets were suspended in 3.8 ml of 15 mM EDTA (pH 8.0) and then dissolved by adding 0.24 ml of 10% SDS (to a concentration of 0.6%) while quickly mixing. Most of the chromosomal DNA was then removed using the Hirt procedure, by gently mixing in 1 ml of 5 M NaCl, leaving the mixture on ice for 4 to 16 h, and then centrifuging it at 10,000⫻gfor 1 h at 4°C. DNA was purified from the supernatants as described above for the cytoplasmic fractions.
RESULTS
EBNA1’s tethering domains are needed for efficient replication of DS-containing plasmids that are within nuclei.To explore the role of EBNA1’s N-terminal domains in plasmid replication, we sought conditions for a transient transfection assay that would minimize contributions of chromosome tethering by EBNA1 to plasmid survival. First, we chose a cell line, HEK293, that retains transfected plasmids unusually well. Second, we tested plasmids that carry only DS, eliminating the role of FR in retaining plasmids within nuclei. In HEK293 cells, plasmids carrying only DS repli-cate as well as plasmids carryingoriPfor a few days after transfec-tion, not requiring nuclear retention by FR (26). Third, we har-vested plasmids after brief periods that still allow good replication signals, 48 to 63 h after transfection. Finally, we were able to show that essentially all of the plasmids recovered from cells after trans-fections had been taken up by the cells and in fact had been
recov-ered from nuclei. Apparently, any plasmids lost to the cytoplasm are rapidly degraded.
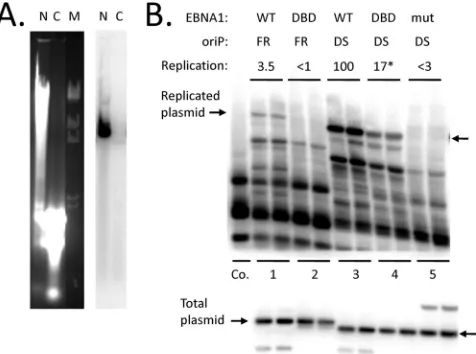
Cells that have been transfected using calcium phosphate co-precipitates can be washed subsequently with PBS-EDTA to dis-solve and remove any external calcium phosphate-DNA particles. When this is done, we find that the plasmids used to transfect cells are recovered almost entirely within the nuclear fraction of cells as early as a few hours after transfection and from then on. Similar results have been obtained using two different, common methods to separate nuclei from the cytosol using nonionic detergents (Materials and Methods.) An example is shown inFig. 2A. More than 99% of a plasmid recovered from HEK293 cells 59 h after transfection appeared in the nuclear fraction, with less than 0.5% appearing in the cytoplasmic fraction. The plasmid used, pHEBo, containsoriP, but EBNA1 was not present to support it. Thus, plasmids that survive within transfected cells have been within the nuclei of cells, effectively for the duration of the experiment, re-gardless of whether they have been tethered or have replicated.
Using this transfection protocol, plasmids carrying only DS or only FR oforiP were tested together with plasmids expressing either functionally wild-type EBNA1 or dn1160, a dominant-neg-ative derivdominant-neg-ative of EBNA1 that is little more than the DNA-bind-ing domain (originally called⌵⌬450 – 641 [30]). Shown inFig. 2B
are Southern analyses of the recovered plasmids cut with HindIII
FIG 2Apart from preventing plasmid loss, N-terminal domains of EBNA1 increase the efficiency oforiP-dependent replication. (A) Recovery of a plas-mid in nuclear (N) and cytoplasmic (C) fractions from HEK293 cells 59 h after transfection, prepared by method 1 (Materials and Methods.) The samples were digested with SpeI to linearize the plasmid (pHEBo) and run on a 0.7% agarose gel, shown stained with ethidium bromide (left). A Southern blot of the gel probed with pHEBo is shown to the right. M, bacteriophage lambda DNA cut with HindIII. (B) Southern blot assay for replicated plasmids recov-ered 63 h after transfection of HEK293 cells. Percentages of replication are shown above. (Top gel) A portion of each sample digested with DpnI plus HindIII; (bottom gel) 40% as much of each digested with HindIII. Arrows indicate the positions of the linear, full-length plasmids derived from pHEBo, carrying only DS or FR oforiP. Co., 2 ng of pHEBo mixed with DNA from mock-transfected cells as a control to check for completeness of DpnI diges-tion. Cotransfected plasmids used to express wild-type (WT) EBNA1 (WT), the EBNA1 DNA-binding domain (DBD) (dn1160), or a mutant EBNA1 un-able to bind DNA (mut) produced the additional bands seen in the lower blot. The plasmid used to express wild-type EBNA1 lacks a HindIII site, so it re-mained primarily in the supercoiled form. *, average of six transfections (SD, 3.5%).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.303.541.67.244.2]to linearize them and with DpnI to reveal replicated, DpnI-resis-tant plasmids (upper image); also shown is a smaller portion of each sample cut with HindIII to reveal the amount of recovered plasmid (lower image). The fraction of plasmids that had repli-cated in each sample was calculated by dividing the signal from the DpnI-resistant form by the signal from the total recovered plas-mid (after correction for loading and, when done on separate blots, in relation to known standards). The plasmid carrying DS of
oriP replicated well in the presence of wild-type EBNA1, with 8.0% of the retained plasmid molecules, on average, being fully resistant to DpnI (lanes 3). dn1160 also supported replication of the plasmid carrying DS, but did so inefficiently; only 1.5% of the retained plasmid DNA had replicated, on average (Fig. 2B, lanes 4). Note that the amounts of plasmids retained by the cells were similar in both cases. From this, it appears that the N-terminal domains of EBNA1 are needed to support efficient replication from DS oforiPon plasmids that are within the nucleus.
It is significant that the DNA-binding domain of EBNA1 alone (mutant dn1160) supported some replication from DS, at 17% of the efficiency seen with functionally wild-type EBNA1 (the aver-age of 6 transfections; standard deviation [SD], 3.5%). This repli-cation was indeed due to dn1160 EBNA1, because a mutant EBNA1 incapable of binding to DNA did not support replication at a detectable level (Fig. 2B, lanes 5). Replication due to dn1160 was specific for DS, since dn1160 did not support detectable rep-lication of a similar plasmid carrying FR of oriP (with its 20 EBNA1-binding sites) but lacking DS (lanes 1). The implication is that the DNA-binding domain of EBNA1 alone has some capacity to recruit ORC to DS.
A third point to note here is that wild-type EBNA1 supported the replication of the plasmid carrying only FR oforiP(without DS) at a very low, but detectable, level (lanes 1), while dn1160 EBNA1, lacking the chromosome-binding domains, did not (lanes 2). Presumably, replication of this plasmid is initiated at nonspecific sites, as was shown for plasmids carrying FR and ran-domly chosen DNA segments from heterologous sources, while depending on FR and EBNA1 for stable plasmid maintenance (34). Yet, under the conditions of this transient replication assay, EBNA1 and FR did not noticeably increase the amount of retained DNA. This suggests that tethering of plasmids to human chromo-somes by means of FR and EBNA1 increases the efficiency of rep-lication initiation at non-DS sites as well.
In some cell lines, DS activity requires FR incis.DS must have some capacity to tether plasmids to host chromosomes in the presence of EBNA1 because the presence of two or more copies of DS allows plasmids lacking FR to be maintained rather stably un-der selection (13). The results presented above and those to follow suggest that this weak tethering by DS is enough to support effi-cient replication in some cell lines in the absence of FR, namely, HEK293 (16) and derivatives of HeLa (12). (In one study, FR was even found to reduce replication efficiency in HEK293 cells [35], but this was probably a context effect of the particular plasmid vector used, since it is not seen with the plasmids used here or in the other studies [12,16].) However, in some cell lines, DS cannot support significant plasmid replication even transiently in the ab-sence of FR oforiP, as was first reported for the EBV-positive Burkitt’s lymphoma cell line, Raji (13). It has not been determined whether this apparent need for FR is a direct one or whether FR is needed simply to maintain the plasmids in nuclei long enough for replicated plasmids to survive and accumulate. The following
ex-periments address this question using the human osteosarcoma-derived cell line 143B.
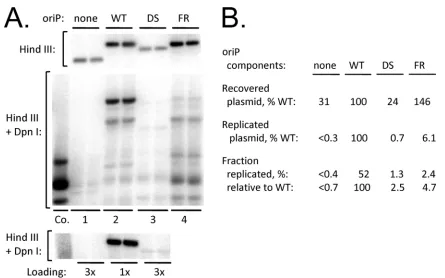
As with HEK293 cells, transfected plasmids are found entirely in the nuclear fraction of 143B cells under the protocol mentioned above (data not shown). The upper image ofFig. 3Ashows a Southern assay for plasmids retained by cells 50 h after transfec-tion of a derivative of 143B that constitutively expresses EBNA1. EBNA1 can increase the apparent transfection efficiency of FR-containing plasmids in some cell lines, particularly if the cells con-tain EBNA1 before the FR-carrying plasmid is introduced. The plasmid pHEBo, which containsoriP, was retained at higher levels than its derivative lacking FR or the vector alone, which were detected at⬃25% and⬃20% of the level of pHEBo, respectively. DpnI resistance assays, seen in the middle and bottom images of the figure, revealed that the plasmid containing DS replicated so poorly that the DpnI-resistant form was difficult to detect, while replication of the vector itself could not be detected. Quantitative analysis of the images revealed that DS supported replication only 2.5% as effectively as didoriP, even after correction for the lower level of plasmid retained by the cells. Clearly FR is required for efficient DS-dependent replication in 143B cells, even for the plas-mids that are retained in nuclei. The results that follow suggest that FR contributes to replication from DS by tethering plasmids to the cell’s chromosomes.
A simple nucleosome-binding domain restores full replica-tion and maintenance activity to the EBNA1 DNA-binding do-main.From the above observations, it seemed possible, and in-deed likely, that the N-terminal domains of EBNA1 contribute to initiation of replication at DS in the same way that they prevent the loss of plasmids from nuclei, by tethering plasmids to cellular chromosomes. To test this idea, we replaced most of the N-termi-nal two-thirds of EBNA1, including both chromosome-binding domains, with a 22-amino-acid segment from the N terminus of the LANA protein of KSHV, which binds to nucleosomes. This small domain of LANA binds stably to chromatin through a well-defined interaction with histones H2A and H2B at an exposed acidic pocket of the nucleosome core (6). It can target green fluo-rescent protein (GFP) to metaphase chromosomes, it enables LANA to tether plasmids containing the KSHV terminal repeats (to which LANA binds at specific sites) to human mitotic chro-mosomes, and it is necessary for the detection of efficient LANA-dependent replication from the KSHV terminal-repeat origin in transient-transfection assays (6, 36, 37). The structures of the LANA-EBNA1 hybrid protein and some derivatives are shown in
Fig. 4A, along with a test for levels of expression inFig. 4B. Dele-tions between amino acids 16 and 376 of EBNA1 in some cases resulted in proteins that were expressed very poorly (e.g., 1020, which lacks both AT hook regions). The substitution of LANA residues 2 to 22 for EBNA1 residues 16 to 376 led to stable expres-sion though.
We first tested the plasmids that express these mutants (and also containoriP) in 143B cells transiently transfected, because of the strict dependency of replication on tethering in these cells. With wild-type EBNA1 and with 1022, in which LANA residues 2 to 22 replace residues 16 to 376 of EBNA1, most of the plasmids recovered from cells 96 h after transfection had replicated (Fig. 4C). In fact, 1022 led to a higher recovery of replicated plasmids than did wild-type EBNA1 in this and similar experiments using transient transfection (data not shown). The human protein IL33 contains near its amino terminus a nucleosome-binding domain
on November 7, 2019 by guest
http://jvi.asm.org/
that is homologous to LANA residues 2 to 22 and binds nucleo-somes similarly (38). Substitution of this homologous region, res-idues 40 to 58 of IL33, for resres-idues 16 to 376 of EBNA did not result in a stable protein. Insertion of a longer region of the IL33 N terminus, residues 1 to 65, gave better expression, although stabil-ity appeared still to be a problem; the protein supported plasmid replication to a significant degree (plasmid 1040) (Fig. 4BandC). A methionine at position 6 and a leucine at position 8 of LANA make hydrophobic contacts in the acidic pocket of the histone H2A-H2B dimer (6), and changing either to an alanine at the homologous positions of IL33 was shown to impair binding to chromosomes (38). Each mutation, when introduced into 1022, nearly eliminated plasmid replication (Fig. 4C, 1029 and 1030.).
Clearly, the nucleosome-binding domain of LANA can func-tionally replace both chromosome-binding domains of EBNA1. Wondering whether chromosome binding is the only essential function that is not provided by the EBNA1 DNA-binding do-main in supporting replication fromoriP, we deleted all but 9 of the 64 amino acids between the nuclear localization sequence and the DNA-binding domain of 1022. The region includes binding sites for USP7 and CK2 and a phosphorylation site for Cdk2 (Fig. 1) (39–41), with evidence that the latter plays a role inoriP func-tion in BJAB cells (40), although deletion of the site of interaction hardly mattered in 143B cells (29). The resulting construct, 1024, containing the LANA nucleosome-binding domain but lacking residues 387 to 450 of EBNA1, was much less active than 1022, although it still showed significant activity. Kennedy et al. ran-domized the amino acids in this region to test whether its apparent
role in transcriptional activation was to provide a flexible joining segment between the DNA-binding domain and N-terminal do-mains that activate transcription (31). This randomized segment of amino acids 393 to 454, when substituted into 1022 resulted in activity similar to that of wild-type EBNA1, although still notice-ably below that of 1022. Neither the deletion nor the randomiza-tion of this region affected the level of protein expression (data not shown), as found previously (31).
To test how well the LANA nucleosome-binding domain would support the EBNA1 DNA-binding domain to maintain the plasmid under genetic selection, 143B cells transfected with p1022 and p803 (wild-type EBNA1) were grown in the presence of hy-gromycin B. The plasmids, derived from p367 (29), carry thehph
(hygromycin phosphotransferase) gene expressed from the HSVtkpromoter, which is very weak in this context but is strongly enhanced by EBNA1 binding to FR oforiP(42). The enhancer activity of FR requires the zinc finger and AT hook regions of regions LR1 and LR2 of EBNA1, which also bind to chromosomes (7,19,20,22,31). With these regions replaced by LANA residues 2 to 22 in p1022, emerging hygromycin B-resistant colonies car-rying this plasmid grew more slowly than colonies carcar-rying p803 expressing wild-type EBNA1. Colonies emerged in very high numbers with both plasmids, though, and at least 20 times more efficiently than occurred with plasmids lacking a functional EBNA1 gene. For comparison, we tested a plasmid for which EBNA1 lacks the zinc-binding region (amino acids 65 to 89), ex-pecting plasmid maintenance and replication at normal levels without any transcriptional activation from EBNA1 (26,31).
Col-FIG 3FR is essential for significantoriP-dependent replication in EBNA1-expressing 143B cells. (A) Duplicate cultures were transfected with plasmids lacking oriP(none), carryingoriP(wild type [WT]), or carrying only DS or FR, as indicated. (Top image) A Southern blot analysis of the plasmids recovered from cells 50 h after transfection, using 1/10 of each sample cut with HindIII. (Middle image) Analysis of 1/4 of each sample digested with HindIII plus DpnI. Co., 2 ng of a plasmid mixed with DNA from mock-transfected cells to check for completeness of DpnI digestion. (Bottom image) A repeat of the DpnI assay using only 1/3 as much of the samples with wild-typeoriP(1/4 or 1/12 of each sample.) (B) Quantitative results from the images.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.72.517.61.339.2]onies carrying this plasmid emerged at a high frequency and grew somewhat more slowly than those obtained with p1022. After 13 days of culture in the presence of hygromycin B, cells were trans-ferred to new plates and grown without selection for 3 to 6 more days before plasmids were harvested.
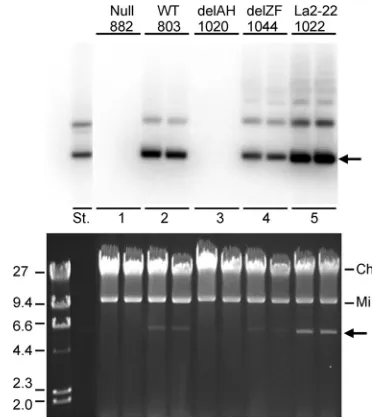
Except for negative controls, the plasmids were maintained at levels high enough to be visible by ethidium bromide staining of an agarose gel following electrophoresis (Fig. 5, lower image). A Southern blot of the gel is shown above. p1022, for which the LANA nucleosome-binding domain replaces the EBNA1 chromo-some-binding domains, was recovered at the highest level, esti-mated to be at least 100 copies per cell by quantification of the radiometric image. Wild-type EBNA1 and EBNA1 lacking the zinc finger region supported plasmid levels of about 40 and 25 copies per cell, respectively.
About 10% of p1022 molecules accumulated as multimers, and the same fraction of multimers accumulated with EBNA1 lacking the zinc finger region. We estimated that at most 2% of p803 molecules, expressing wild-type EBNA1, accumulated as multimers; only a faint band at the position of a plasmid dimer could be resolved, represent-ing much less than this amount. It has not been explored whether the higher proportion of multimeric plasmids arose due to selective pres-sure (for expression of thehphgene or for mitotic stability) or due to a higher rate of recombination between plasmids.
DISCUSSION
Two issues have clouded the long-held suspicion that efficient duplication of EBV-derived plasmids during S phase might re-quire tethering by EBNA1 to host chromosomes. One has been the problem of determining how much of the apparent requirement for EBNA1’s tethering domains might be simply to prevent the
FIG 4The 22-amino-acid nucleosome-binding domain of LANA functionally replaces the N-terminal domains of EBNA1. (A) Diagram of LANA-EBNA1 hybrid proteins. DBD, DNA-binding domain. (For definitions of other abbreviations, see the legend toFig. 1and the text.) Numbers to the left refer to the mutant proteins as well as the plasmids that express them. (B) Expression of the mutant proteins in transiently transfected 293T cells assessed by immunoblotting, using lysates from 50,000, 15,000, and 5,000 cells transfected with p803 (from left to right) and from 50,000 cells for the others. (C) Southern analysis of DpnI-resistant (replicated) plasmids isolated 4 days after transfection of 143B cells, with duplicate tests of each as indicated. Samples were also digested with BamHI to linearize the plasmids. Co., 2 ng of p803 mixed with a sample from mock-transfected cells as a control for DpnI digestion.
FIG 5Efficient replication and maintenance of anoriPplasmid supported by the nucleosome-binding domain of LANA fused to the DNA-binding domain of EBNA1. Images of a Southern analysis (above) and the ethidium-bromide-stained 0.7% agarose gel (below) are shown with an arrow indicating the position of the supercoiledoriP-carrying plasmids. Mi and Ch indicate the 16-kb supercoiled mitochondrial plasmid and residual human chromosomal DNA, respectively. The plasmids expressed EBNA1 with its amino-terminal domains either being wild type (WT) (p803), car-rying a frameshift mutation (null), lacking both AT hook regions (delAH) (p1020), lacking the zinc finger region (delZF) (p1044), or having been replaced by the LANA nucleosome-binding domain (La2-22) (p1022). DNA from alkaline extracts of⬃5 million cells was analyzed from duplicate transfections. To the left were run 0.5g of bacteriophage lambda DNA cut with HindIII (sizes shown in kb) and 1 ng of p803 (lane St.).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.95.485.65.301.2] [image:6.585.325.511.385.594.2]loss of plasmids from nuclei during mitosis. The other has been uncertainty as to whether the same regions of EBNA1 that bind to chromosomes also are needed to recruit ORC. This study resolves both issues. EBV-derived plasmids appear to replicate only very inefficiently unless tethered to chromosomes within the inter-phase nucleus.
While untethered plasmids are mostly lost from cells within a few cell divisions following transfection, we find that the plasmids are detected almost exclusively in the nuclear fraction between a few hours and a few days after transfection. This suggests that any plasmids that are lost to the cytoplasm during mitosis are quickly degraded. We interpret this to mean that untethered plasmids that are recovered from cells a few days after transfection—those that have managed to survive— have spent as much time within nuclei as tethered plasmids. Since plasmids replicate very poorly without tethering, all of this implies that tethering to chromosomes within interphase nuclei plays an important role in plasmid replication. One caveat is that if untethered plasmids were lost to a cytoplas-mic compartment but were for some reason to adhere to nuclei during the separation of cytoplasmic and nuclear fractions, the plasmids would only appear to be within nuclei and the effect of tethering might indeed be entirely that of keeping the plasmids within nuclei. While this seems unlikely, the caveat cannot be ruled out entirely at this time. Consistent with our favored inter-pretation though, Lechardeur et al. found that plasmids directly microinjected into the cytoplasm of HeLa cells were degraded with a half-life of 50 to 90 min (43), and Wang and Sugden found that plasmids were recovered primarily with the nuclear matrix fraction days after transfection, regardless of the presence or ab-sence oforiP(44). In addition, the magnitude of the effect that tethering has on plasmid replication seems to argue against nu-clear retention being the only role of tethering.
The possibility that the chromosome-binding domains of EBNA1 might be required in any direct manner to recruit ORC now seems remote, becauseoriPis fully supported by a derivative of EBNA1 lacking its own chromosome-binding domains but containing the nucleosome-binding domain of LANA of KSHV, which is only 22 amino acids long. Only the DNA-binding domain of EBNA1 is entirely essential to support replication from DS, it can do so inefficiently on its own, and all it appears to need to approach full activity is the addition of any chromosome-binding domain. As discussed previously (15), an exact, tight spacing be-tween adjacent EBNA1-binding sites is what makes DS a replica-tion origin, so a precise structure formed there by adjacent EBNA1 DNA-binding domains would appear to be the key to recruiting ORC.
An insightful study by Hung, Kang, and Kieff in 2001 presaged this conclusion by showing that the DNA-binding domain of EBNA1, if fused to either of two chromosome-binding proteins, histone H1 or HMGA1, would support full replication and main-tenance functions oforiP(9). However, it was subsequently re-ported that HMGA1 can interact with ORC and, when fused to the tet repressor and directed to a cluster of sites on a plasmid, can make a functional replication origin, albeit an inefficient one com-pared to DS (45). Such properties have not been reported for histone H1, though, and in any case are unlikely to be shared by the 22-amino-acid nucleosome-binding domain of LANA.
Moriyama recently reported promisingin vitrostudies of the recruitment of ORC and CDC6 to DS (46). The association with ORC was specific for DS and not FR, and it did not involve an RNA
or DNA intermediate. A functional EBNA1 derivative, having the maltose binding protein (MBP) in place of the Gly-Ala repeats, could recruit ORC to DS, while MBP fused to EBNA1’s DNA-binding domain (amino acids 452 to 641) could not. Without further experiments, it is unclear what regions of EBNA1’s first 451 amino acids are needed for the binding activity in this context or whether MBP might have interfered with ORC recruitment when fused so close the EBNA1’s DNA-binding domain. In any case, when EBNA1’s DNA-binding domain is linked to the 22-amino-acid nucleosome-binding domain of LANA, it requires very little else to be fully functionalin vivo.
With regard to KSHV-derived plasmids supported by LANA, a connection between chromosome tethering and replication has been more apparent than it has for EBV-derived plasmids, be-cause the essential chromosome-binding domain of LANA has been so clearly defined (6). A pair of adjacent LANA-binding sites, present in each⬃800-bp terminal repeat copy on the viral ge-nome, serve both as a tethering site for mitotic stability and, to-gether with some adjacent sequences, as a LANA-dependent rep-lication origin (3). The nucleosome-binding domain within the first 22 amino acids of the amino terminus is essential for plasmid maintenance and targeting of GFP to chromosomes (36). In tests for LANA-dependent transient replication of terminal-repeat-containing plasmids, the nucleosome-binding domain seemed to be essential in BJAB, a B-lymphoma cell line (36), but the DNA-binding domain by itself showed very clear, though weak, activity in HEK293 cells in another study, being 20% as active as the wild type (37). Perhaps the dependence of replication on tethering might be less strict in some cell lines (HEK293) than it is in others, as suggested by our own experiments with EBV-derived plasmids. An intriguing (and all but forgotten) fact is that while DS is fully active without FR in some cell lines (HEK293 and HeLa) (12,
16), in the B-lymphoma cell line Raji, it was virtually inactive without FR (13). Because plasmids lacking FR were retained very poorly by Raji cells, it has not been clear whether FR is needed for replicationper se. Here we found that in an osteosarcoma-derived cell line, 143B, DS supported plasmid replication just 2.5% as well without FR as with FR, even after taking into account the reduced survival of plasmids lacking FR. Presumably, FR is supporting plasmid replication by tethering to chromosomes during inter-phase. FR has no significant activity as a replication origin and does not recruit ORC (11–13, 42, 47), but of course it tethers plasmids to chromosomes during mitosis, which is essential for mitotic stability in all cell lines. DS has weak tethering ability, since multiple copies of it provide stable plasmid maintenance (13). Perhaps the relatively weak tethering by DS is sufficient for repli-cation in HeLa and HEK293 but not in other cell lines, such as Raji and 143B, where a tighter association with chromosomes might be required. This issue has not been explored, but it would not be surprising if the strong dependency of replication on tethering by FR turned out to be representative of more normal cells, given that HEK293 (48) and HeLa (49) cells carry the potent transforming oncogenes of adenovirus 5 and human papillomavirus 18.
A requirement that plasmids be tethered to host chromosomes in order to replicate efficiently suggests that not all spaces within the nucleus are suitable for some aspect of origin function or rep-lication. It suggests that subcompartments exist that are favorable for DNA replication and that untethered plasmids usually fail to reach these sites on their own. In principle this could apply to the recruitment of ORC and origin licensing during G1phase or to
on November 7, 2019 by guest
http://jvi.asm.org/
origin firing and replication during S phase. The former does not seem so likely given the abundance of replication origins on mam-malian chromosomes and the rapid mobility of ORC observed in photobleaching experiments (50). Replication of mammalian chromosomes, though, takes place in numerous concentrated foci, each focus suspected to contain on the order of 6 to 12 con-tiguous replicons firing synchronously on a single chromosome (51–53). As replication follows a temporal pattern, most replicons are inactive most of the time during S phase. Conceivably, an isolated plasmid might miss out if not tethered.
ACKNOWLEDGMENTS
We thank Bill Sugden for providing plasmids with mutant EBNA1 genes and for helpful comments on the manuscript and Liam Coyne for help with the figures and manuscript.
This work was supported by grant CA43122 to J.L.Y. and grant CA16056 to the Biopolymer Facility of the Roswell Park Cancer Institute.
REFERENCES
1.Ganem D.2007. Kaposi’s sarcoma-associated herpesvirus, p 2847–2888. InKnipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE (ed), Fields virology, 5th ed, vol 2. Lippincott Williams & Wilkins, Philadelphia, PA.
2.Kieff ED, Rickinson AB.2007. Epstein-Barr virus and its replication, p 2603–2654.InKnipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE (ed), Fields virology, 5th ed, vol 2. Lippincott Wil-liams & Wilkins, Philadelphia, PA.
3.Ballestas ME, Kaye KM.2011. The latency-associated nuclear antigen, a multifunctional protein central to Kaposi’s sarcoma-associated herpesvi-rus latency. Future Microbiol.6:1399 –1413.
4.Hammerschmidt W, Sugden B.2006. Epstein-Barr virus, p 187–706.In Depamphilis ML (ed), DNA replication and human disease. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
5.Grundhoff A, Ganem D.2003. The latency-associated nuclear antigen of Kaposi’s sarcoma-associated herpesvirus permits replication of terminal repeat-containing plasmids. J. Virol.77:2779 –2783.
6.Barbera AJ, Chodaparambil JV, Kelley-Clarke B, Joukov V, Walter JC, Luger K, Kaye KM.2006. The nucleosomal surface as a docking station for Kaposi’s sarcoma herpesvirus LANA. Science311:856 – 861. 7.Marechal V, Dehee A, Chikhi-Brachet R, Piolot T, Coppey-Moisan M,
Nicolas JC.1999. Mapping EBNA-1 domains involved in binding to metaphase chromosomes. J. Virol.73:4385– 4392.
8.Middleton T, Sugden B.1994. Retention of plasmid DNA in mammalian cells is enhanced by binding of the Epstein-Barr virus replication protein EBNA1. J. Virol.68:4067– 4071.
9.Hung SC, Kang MS, Kieff E.2001. Maintenance of Epstein-Barr virus (EBV) oriP-based episomes requires EBV-encoded nuclear antigen-1 chromosome-binding domains, which can be replaced by high-mobility group-I or histone H1. Proc. Natl. Acad. Sci. U. S. A.98:1865–1870. 10. Kanda T, Otter M, Wahl GM.2001. Coupling of mitotic chromosome
tethering and replication competence in Epstein-Barr virus-based plas-mids. Mol. Cell. Biol.21:3576 –3588.
11. Gahn TA, Schildkraut CL.1989. The Epstein-Barr virus origin of plasmid replication, oriP, contains both the initiation and termination sites of DNA replication. Cell58:527–535.
12. Harrison S, Fisenne K, Hearing J.1994. Sequence requirements of the Epstein-Barr virus latent origin of DNA replication. J. Virol.68:1913–1925. 13. Wysokenski DA, Yates JL.1989. Multiple EBNA1-binding sites are
re-quired to form an EBNA1-dependent enhancer and to activate a minimal replicative origin within oriP of Epstein-Barr virus. J. Virol.63:2657–2666. 14. Norio P, Schildkraut CL, Yates JL.2000. Initiation of DNA replication within oriP is dispensable for stable replication of the latent Epstein-Barr virus chromosome after infection of established cell lines. J. Virol.74:8563– 8574. 15. Bashaw JM, Yates JL.2001. Replication from oriP of Epstein-Barr virus
requires exact spacing of two bound dimers of EBNA1 which bend DNA. J. Virol.75:10603–10611.
16. Yates JL, Camiolo SM, Bashaw JM. 2000. The minimal replicator of Epstein-Barr virus oriP. J. Virol.74:4512– 4522.
17. Deng Z, Atanasiu C, Burg JS, Broccoli D, Lieberman PM.2003.
Telo-mere repeat binding factors TRF1, TRF2, and hRAP1 modulate replica-tion of Epstein-Barr virus OriP. J. Virol.77:11992–12001.
18. Deng Z, Lezina L, Chen CJ, Shtivelband S, So W, Lieberman PM.2002. Telomeric proteins regulate episomal maintenance of Epstein-Barr virus origin of plasmid replication. Mol. Cell9:493–503.
19. Sears J, Kolman J, Wahl GM, Aiyar A.2003. Metaphase chromosome tethering is necessary for the DNA synthesis and maintenance of oriP plasmids but is insufficient for transcription activation by Epstein-Barr nuclear antigen 1. J. Virol.77:11767–11780.
20. Sears J, Ujihara M, Wong S, Ott C, Aiyar A.2004. The amino terminus of Epstein-Barr virus (EBV) nuclear antigen 1 (EBNA1) contains AT hooks that facilitate the replication and partitioning of latent EBV genomes by tethering them to cellular chromosomes. J. Virol.78:11487–11505.
21. Aras S, Singh G, Johnston K, Foster T, Aiyar A.2009. Zinc coordination is required for and regulates transcription activation by Epstein-Barr nuclear antigen 1. PLoS Pathog.5:e1000469. doi:10.1371/journal.ppat.1000469. 22. Lin A, Wang S, Nguyen T, Shire K, Frappier L. 2008. The EBNA1
protein of Epstein-Barr virus functionally interacts with Brd4. J. Virol. 82:12009 –12019.
23. Norseen J, Johnson FB, Lieberman PM.2009. Role for G-quadruplex RNA binding by Epstein-Barr virus nuclear antigen 1 in DNA replication and metaphase chromosome attachment. J. Virol.83:10336 –10346. 24. Norseen J, Thomae A, Sridharan V, Aiyar A, Schepers A, Lieberman
PM.2008. RNA-dependent recruitment of the origin recognition com-plex. EMBO J.27:3024 –3035.
25. Mackey D, Sugden B.1999. The linking regions of EBNA1 are essential for its support of replication and transcription. Mol. Cell. Biol.19:3349 – 3359.
26. Wu H, Kapoor P, Frappier L.2002. Separation of the DNA replication, segregation, and transcriptional activation functions of Epstein-Barr nu-clear antigen 1. J. Virol.76:2480 –2490.
27. Yates JL, Warren N, Sugden B.1985. Stable replication of plasmids derived from Epstein-Barr virus in various mammalian cells. Nature313: 812– 815.
28. Sugden B, Marsh K, Yates J.1985. A vector that replicates as a plasmid and can be efficiently selected in B-lymphoblasts transformed by Epstein-Barr virus. Mol. Cell. Biol.5:410 – 413.
29. Yates JL, Camiolo SM.1988. Dissection of DNA replication and enhancer activation functions of Epstein-Barr virus nuclear antigen 1. Cancer Cells 6:197–205.
30. Kirchmaier AL, Sugden B. 1997. Dominant-negative inhibitors of EBNA-1 of Epstein-Barr virus. J. Virol.71:1766 –1775.
31. Kennedy G, Sugden B. 2003. EBNA-1, a bifunctional transcriptional activator. Mol. Cell. Biol.23:6901– 6908.
32. Frost E, Williams J.1978. Mapping temperature-sensitive and host-range mutations of adenovirus type 5 by marker rescue. Virology91:39 –50. 33. Belgrader P, Cheng J, Zhou X, Stephenson LS, Maquat LE. 1994.
Mammalian nonsense codons can be cis effectors of nuclear mRNA half-life. Mol. Cell. Biol.14:8219 – 8228.
34. Krysan PJ, Haase SB, Calos MP.1989. Isolation of human sequences that replicate autonomously in human cells. Mol. Cell. Biol.9:1026 –1033. 35. Leight ER, Sugden B.2001. The cis-acting family of repeats can inhibit as well
as stimulate establishment of an oriP replicon. J. Virol.75:10709 –10720. 36. Barbera AJ, Ballestas ME, Kaye KM. 2004. The Kaposi’s
sarcoma-associated herpesvirus latency-sarcoma-associated nuclear antigen 1 N terminus is essential for chromosome association, DNA replication, and episome per-sistence. J. Virol.78:294 –301.
37. Hu J, Garber AC, Renne R.2002. The latency-associated nuclear antigen of Kaposi’s sarcoma-associated herpesvirus supports latent DNA replica-tion in dividing cells. J. Virol.76:11677–11687.
38. Roussel L, Erard M, Cayrol C, Girard JP. 2008. Molecular mimicry between IL-33 and KSHV for attachment to chromatin through the H2A-H2B acidic pocket. EMBO Rep.9:1006 –1012.
39. Holowaty MN, Sheng Y, Nguyen T, Arrowsmith C, Frappier L.2003. Protein interaction domains of the ubiquitin-specific protease, USP7/ HAUSP. J. Biol. Chem.278:47753– 47761.
40. Kang MS, Lee EK, Soni V, Lewis TA, Koehler AN, Srinivasan V, Kieff E. 2011. Roscovitine inhibits EBNA1 serine 393 phosphorylation, nuclear local-ization, transcription, and episome maintenance. J. Virol.85:2859 –2868. 41. Sivachandran N, Cao JY, Frappier L.2010. Epstein-Barr virus nuclear
antigen 1 Hijacks the host kinase CK2 to disrupt PML nuclear bodies. J. Virol.84:11113–11123.
42. Reisman D, Yates J, Sugden B.1985. A putative origin of replication of
on November 7, 2019 by guest
http://jvi.asm.org/
plasmids derived from Epstein-Barr virus is composed of two cis-acting components. Mol. Cell. Biol.5:1822–1832.
43. Lechardeur D, Sohn KJ, Haardt M, Joshi PB, Monck M, Graham RW, Beatty B, Squire J, O’Brodovich H, Lukacs GL.1999. Metabolic insta-bility of plasmid DNA in the cytosol: a potential barrier to gene transfer. Gene Ther.6:482– 497.
44. Wang CY, Sugden B.2008. Identifying a property of origins of DNA synthesis required to support plasmids stably in human cells. Proc. Natl. Acad. Sci. U. S. A.105:9639 –9644.
45. Thomae AW, Pich D, Brocher J, Spindler MP, Berens C, Hock R, Hammerschmidt W, Schepers A.2008. Interaction between HMGA1a and the origin recognition complex creates site-specific replication ori-gins. Proc. Natl. Acad. Sci. U. S. A.105:1692–1697.
46. Moriyama K, Yoshizawa-Sugata N, Obuse C, Tsurimoto T, Masai H. 2012. Epstein-Barr nuclear antigen 1 (EBNA1)-dependent recruitment of origin recognition complex (Orc) on oriP of Epstein-Barr virus with pu-rified proteins: stimulation by Cdc6 through its direct interaction with EBNA1. J. Biol. Chem.287:23977–23994.
47. Schepers A, Ritzi M, Bousset K, Kremmer E, Yates JL, Harwood J, Diffley JFX, Hammerschmidt W.2001. Human origin recognition com-plex binds to the region of the latent origin of DNA replication of Epstein-Barr virus. EMBO J.20:4588 – 4602.
48. Graham FL, Smiley J, Russell WC, Nairn R.1977. Characteristics of a human cell line transformed by DNA from human adenovirus type 5. J. Gen. Virol.36:59 –74.
49. Picken RN, Yang HL.1987. The integration of HPV-18 into HeLa cells has involved duplication of part of the viral genome as well as human DNA flank-ing sequences. Nucleic Acids Res.15:10068. doi:10.1093/nar/15.23.10068. 50. McNairn AJ, Okuno Y, Misteli T, Gilbert DM.2005. Chinese hamster
ORC subunits dynamically associate with chromatin throughout the cell-cycle. Exp. Cell Res.308:345–356.
51. Gilbert DM, Stillman B.2006. DNA replication and nuclear architecture, p 179 –196.InDePamphilis ML (ed), DNA replication and human disease. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. 52. Ma H, Samarabandu J, Devdhar RS, Acharya R, Cheng PC, Meng C,
Berezney R.1998. Spatial and temporal dynamics of DNA replication sites in mammalian cells. J. Cell Biol.143:1415–1425.
53. Maya-Mendoza A, Olivares-Chauvet P, Shaw A, Jackson DA.2010. S phase progression in human cells is dictated by the genetic continuity of DNA foci. PLoS Genet.6:e1000900. doi:10.1371/journal.pgen.1000900. 54. Ambinder RF, Mullen MA, Chang YN, Hayward GS, Hayward SD.
1991. Functional domains of Epstein-Barr virus nuclear antigen EBNA-1. J. Virol.65:1466 –1478.