JOURNALOFVIROLOGY, June 1993,p. 3418-3426
0022-538X/93/063418-09$02.00/0
Copyright ©1993, American SocietyforMicrobiology
Identification
of EBNA1 Amino Acid
Sequences Required for
the
Interaction of the Functional
Elements of the
Epstein-Barr
Virus Latent Origin of DNA Replication
KIM GOLDSMITH, LESLIE BENDELL, ANDLORI FRAPPIER* MolecularVirology andImmunology Program, McMaster University,
1200 Main Street West, Hamilton, Ontario, CanadaL8N3Z5 Received 22January 1993/Accepted16 March 1993
Epstein-Barr nuclear antigen 1 (EBNA1) activates DNA replication from the Epstein-Barr virus latent origin, oriP. This activation involvesthedirect interactionof EBNA1 dimers withmultiplesites within thetwo noncontiguous functional elements of the origin, thefamilyof repeats (FR) element and thedyad symmetry (DS) element. The efficient interaction of EBNA1 dimersbound to these two elements in oriP results in the
formation of DNA loops inwhich the FR and DSelements arebound togetherthroughEBNA1.In orderto
elucidate themechanismbywhich EBNA1 induces oriP DNAlooping,wehaveinvestigatedthe DNAsequences and EBNA1 amino acidsrequiredforEBNAl-mediatedDNAlooping. Usingaseries oftruncation mutantsof EBNA1producedinbaculovirusandpurifiedtoapparenthomogeneity,wehave demonstrated that the EBNA1
DNAbinding and dimerizationdomain isnot sufficient to mediate oriP DNAloopingandthatanadditional
region(s) located between amino acids 346 and 450 is required. SingleEBNAl-binding sites, separated by930
bp ofplasmidDNA,werealso shown to support EBNAl-mediatedlooping, indicatingthatthe formation of largeEBNA1complexes, suchasthose observedonoriPFRandDSelements,isnotarequirementforlooping.
EBNA1 is a multifunctional protein essential for the
replication and maintenanceofEpstein-Barr virusgenomes
during latent infection (30). EBNA1 activates the latent
origin ofEpstein-Barr virus DNA replication, oriP, to
ini-tiateasingleround ofreplicationpercellcycle (1,29).This activation involves a direct interaction of EBNA1 with
multiple 18-bp palindromic recognition sites present within the two noncontiguous functional elements of oriP, the
family ofrepeats(FR) and the dyadsymmetry(DS)elements
(19, 22). These elements are separated by approximately 1
kb inoriP, but origin functionisindependentofthisspacing (22). The DS element contains fourEBNA1-binding sites,a
65-bp region of dyad symmetry, and the initiation site for
latent-phase DNAreplication (9, 27). The FR element
con-tains20EBNA1-bindingsites and,when boundbyEBNA1,
activates DNAreplication from the DS element, enhances transcription from avariety ofpromoters, andgoverns the
stable segregation of oriP-containing plasmids during cell
division (12, 21, 25). The mechanism by which EBNA1
functions inanyoftheseprocessesis notyetunderstood.
In order to gain a better understanding of the EBNA1
proteinand themechanism(s) by which it functions,wehave
overproduced EBNA1 by using thebaculovirus expression system(bEBNA1) and purified it to apparent homogeneity (6). PurebEBNA1wasshowntobeadimer in solution and
tobindthemultipleEBNA1-binding sites inoriPas adimer.
bEBNA1binding tothe DS element caused localized struc-tural distortions ofthis element consistent with bEBNA1-induced bending of the DS element (8). Recently, we used electron microscopy to examine the interaction of pure bEBNA1 withorP(7). The resultsindicatedanordertothe
assemblyofbEBNA1 onto orP,with abEBNAl complex first being formed on the FR element of orP. At higher bEBNA1 concentrations, interaction of bEBNA1 with the
* Correspondingauthor.
DS element was observed in the form of looped DNA
molecules, inwhich the FR and DS elements of orPwere
joinedthroughabEBNA1complex, causingtheloopingout
of the 1 kb ofinterveningDNA. EBNA1-mediatedlooping of the FR elementtothe DS elementwasalso reported by Suet
al.(24) andwasfoundtostabilize theinteraction ofEBNA1 with the DS element. These results suggest that
EBNA1-mediated looping oforiP maybe an important step in the
activation of the DS elementbythe FRelement.
DNAloopinghas been described foravarietyof transcrip-tionsystemsas amechanismby whichenhancerorrepressor
elements act at a distance to activate or repress promoter
elements (reviewed inreference 13). DNAloopinghasalso been shownto regulate DNA replication in theEscherichia
coliplasmidsR6K andP1.ForplasmidR6K,thereplication
initiatorproteinmediates DNAlooping betweenthe-yand cX originsto activate replication from the aorigin orbetween
the y and 1Boriginsto activatereplicationfrom the 13origin (15-17). The initiatorprotein ofthe P1 plasmid negatively regulates replicationby mediatingDNAlooping between the
controllocus and thereplication origin(4). Althoughit isnot yetclearwhether similar looping mechanisms playarole in
the initiation of DNA replication ineukaryotes, the multi-component nature of eukaryotic origins suggests that the interaction of origin elements by DNA looping maybe an
important step in the regulation of replication from these origins (reviewed in references 5 and 10). In keeping with
this hypothesis, the UL9 origin-binding protein of herpes simplexvirus type 1 has been shown to mediate looping
betweenbindingsites I and IIwithin
oris
(11).Inorderto further understand EBNA1-mediatedlooping
of oriP and itsfunctionalsignificance,wehave beenmapping
theEBNA1 amino acidsresponsible for theassociationata
distance of DNA-bound EBNA1 dimers. To this end, we
haveoverproducedaseriesoftruncationmutantsofEBNA1
in baculovirus, all of which retain the DNA binding and
dimerization domain (2, 23), and have purified them to 3418
Vol.67,No. 6
on November 9, 2019 by guest
http://jvi.asm.org/
apparent homogeneity. Here we examine their ability to mediate interactions between theoniPFR and DS elements, aswell as between individual EBNA1-binding sites.
MATERIALS ANDMETHODS
DNA. TheoriP-containing DNA used for electron micros-copy wasgenerated by digesting pGEMoriP, which contains the complete oniP sequence in pGEM2 (Promega), with XmnI andHindIl (6). This yields a 3.16-kb DNA fragment containing oriP and a 1.84-kb vector fragment which are easily distinguished by size in the electron microscope. For enhanced ligation efficiency assays, pGEMoriP was linear-ized with MluI, which cuts between the FR and DS elements of oriP. The linearized plasmid was then extracted with phenol-chloroform (1:1) and ethanol precipitated prior to being used in ligation efficiency assays. To construct a DNA fragment containing twoEBNA1-binding sites, pGEMdyad, which contains only the DS element of onP (7), was digested with AvaI to remove a 123-bp fragment containing three EBNA1-binding sites of the DS element. The large (3-kb) AvaI fragment of pGEMdyad was religated to form
pGEMsl, which contains one EBNA1-binding site
corre-sponding to the DS element site 1 (20). pGEMs2 was
generated from pGEMsl by excising the 1.16-kb
PvuII-to-SspIfragment of pGEMsl, containing the EBNA1-binding
site, and ligating it into the unique EcoRI site of pGEMsl (after the recessed 3' ends of pGEMsl had been extended with DNA polymerase I Klenow fragment). The resulting
pGEMs2 plasmid contains two copies of EBNA1-binding
site 1 of the DS element, separated by 930 bp of plasmid DNA. A2.33-kb DNA fragment containing the two EBNA1-bindingsites was excised from pGEMs2 with PvuII and SspI
and purified from low-melting-point agarose prior to being
used in electron microscopy.
Construction ofEBNA1 truncation mutants. EBNA1 DNA fragments coding for bEBNA39,
-346,
and-395
were excised fromplasmid p205 (kindly provided by Bill Sugden [30]) with NcoI, TaqI, and BsaI enzymes, respectively, in addition toMscI.Therestriction fragment used to construct bEBNA330
was generated by partial digestion of p205 with TaqI and
complete digestion withMscI.The ends ofEBNA1
restric-tion fragments were made blunt by extending 3' recessed ends with the DNA polymerase I Klenow fragment (for
bEBNA39,
-330,
and-346) orby removing 5' overhangs withmung bean nuclease (for
bEBNA395).
EBNA1 fragmentswereligated tothebaculovirus transfervectorpVL941-SW
after linearization of thevectorwith Ncol and extension of the 3' recessed ends with the DNA polymerase I Klenow fragment as previously described (6). As a result, the EBNAl-coding sequenceswereplaced next to the ATG start site of the vector, resulting in proteins composed only of
EBNA1 sequences. For
bEBNA451,
the SmaI-to-MscIre-strictionfragmentofp205containing EBNA1 sequences was
ligatedto pVL941-SWafterlinearization of the vector with
XbaI and extension of the 3' recessed ends with the DNA
polymerase I Klenow fragment. The resulting
bEBNA451
proteincontainstwoamino acids(Ala Leu)atitsNterminus
inadditiontoEBNA1 sequences.
bEBNA330619
wasgener-atedbyreplacingtheSacII-to-Spel fragmentofpVL941-SW/
EBNA330, containing the C-terminal 36 amino acids of EBNA1, withaDNA linkercontainingastopcodon. For all
EBNA1 fragments cloned into pVL941-SW, the DNA
se-quences spanning the N-terminal and C-terminal EBNA1 truncationswereverifiedbydideoxysequencingwith
Seque-nase(United StatesBiochemicals).
Generation of recombinant baculoviruses. Recombinant EBNA1 baculoviruses were generated by recombination of pVL941-SW transfer vectors containing EBNA1 sequences with wild-type baculovirus (Autographa californica nuclear polyhedrosis virus) in Spodoptera
ftugiperda
(SF-9) cells as previously described (6). SF-9 cells were propagated as monolayer cultures in Grace's medium (GIBCO Laborato-ries) supplemented with 0.33% yeastolate, 0.33% lactalbu-min hydrolysate, and 10% fetal bovine serum.Purification of bEBNAl mutants from insect cells. SF-9
monolayer cultures were infected with recombinant baculo-virusescontaining EBNA1 fragments and harvested 46 to 48 h postinfection as previously described (6). For
bEBNA395
and -451. infected SF-9 cells were washed in phosphate-buffered saline and then extracted for 30minon ice in 20 mM Tris-HCl (pH 7.5)-135 mM NaCl-1% Nonidet P-40-1 mM
MgCl2-10%glycerol-2 mMphenylmethylsulfonyl fluoride at
107 cells per ml.Extracts were then clarified by centrifuga-tion at12,000 x g for 30minat4°C. bEBNA39,
-330,
-346, and -330-604were
extracted from infected SF-9 nuclei as previ-ouslydescribed forbEBNA1 (6). All bEBNA proteins were purified by chromatography on a heparin-agarose column (Bio-Rad) and then on a DNA affinity column containing EBNA1-binding sites as previously described for bEBNA1 (6). Pure bEBNA1 proteins were then concentrated by loading onto a 1-mlheparin-agarose column in column buffer (20 mM HEPES [N-2-hydroxyethylpiperazine-N'-2-ethane-sulfonic acid] [pH 7.5], 0.5 mM EDTA, 20% glycerol) containing 350 mM NaCl and eluting in column buffer containing 1 M NaCl. The concentrations of pure bEBNA1 proteins were determined by the method of Bradford (3).Electron microscopy. DNA containing oriP (10 fmol as DNA molecules) or two EBNA1-binding sites (20 fmol as DNAmolecules) was incubated with 480 fmol or 1.4 pmol of homogeneousbEBNA1 truncation mutants, respectively, in 20 ,ul of 25 mMTris-HCl (pH 7.5)-300 mM NaCl for 10min at25°C. The higherconcentration ofbEBNA1 proteins used with DNAcontaining two EBNA1-binding sites was neces-sary in order to achieve binding of these proteins to the singleEBNA1 sites, for which the bEBNA1 proteins have an affinity lower than the affinity they have for the FR and DS elements. Reaction mixtures were then diluted sixfold with 20 mM Tris-HCl (pH7.5)-280 mM NaCl-4 mM spermidine and stored on ice (15 to 30 min) prior to being spread on glow-charged carbon-coated copper grids (400 mesh) with-out fixation. Grids were stained with 5% aqueous uranyl
acetate,rotary shadowed with tungsten, and observed with a
JEM-1200EXII electron microscope. Length measurements were performed on projected negatives with a Numonics Graphic Master digitizing tablet.
Enhanced ligation efficiency assays. Homogeneous
bEB-NAl truncation mutants (1 to 4 pmol as dimer) were incu-bated with 45 fmol of a DNAfragment containing the FR and DS elements of oriP in 100 ,ul of 10 mM Tris-HCl (pH 7.5)-300 mMNaCl-20mMMgCl2-2 mMspermidine-lOmM dithiothreitol-2 mM ATP for 10 min at 25°C. Reaction mixtures were diluted twofold prior to the addition of 1 Weiss unit of T4 DNA ligase (GIBCO BRL) and a 5-min incubation at 15°C. Reactions were stopped by the addition ofEDTA to 10 mM and were ethanolprecipitated with 5
jig
of glycogen (Boehringer Mannheim) as a carrier. Samples wereresuspended in 9 ,ul of 1 mMTris-HCl(pH8.0)-0.1 mM EDTA-1% sodium dodecyl sulfate (SDS) and subjected to electrophoresis on a 0.8% agarose gel containing 0.5 ,ug of ethidium bromide per ml.on November 9, 2019 by guest
http://jvi.asm.org/
3420 GOLDSMITH ET AL.
EBNA1
bEBNA1
nuclear DNAbinding/
(GIy-Arg) Gly-Gly-Ala badclocaization dimeazation acidc
5 1 1
90328 458 604 641
66
A
bEBNA
39
bEBNA
330
bEBNA346 bEBNA395
bEBNA
451
bEBNA330-619
330 641
346 641
395 641
451 641
330 619
FIG. 1. bEBNA1 truncation mutants. EBNA1 amino acids presentin eachmutantareshown. Thepositionsof the DNA binding
anddimerization domain, thenuclearlocalizationsignal, and other
salient features ofthe EBNA1 polypeptideare indicated.
RESULTS
Mappingof EBNA1 sequencesessentialfor oriPDNA loop-ing.Inordertomaptheregion(s)of EBNA1which mediates
the association of DNA-bound EBNA1 dimers, we have
constructed a series oftruncations ofEBNA1, all of which retain the DNAbinding anddimerization domain (Fig. 1) (2, 23). Like bEBNA1, all of the truncationmutants lackmost
of the Gly-Gly-Ala repeat region of EBNA1 previously shown to be nonessential for EBNA1 function (28, 30). bEBNA1 mutant proteins were expressed in insect cells, using the baculovirus expression system, and purified to apparent homogeneity. Thepure bEBNA1 mutant proteins
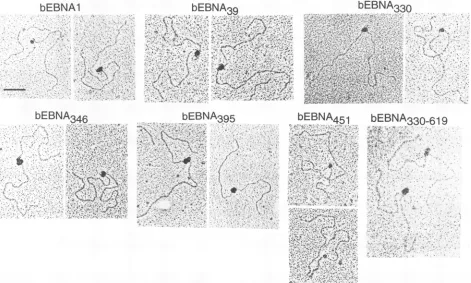
used in subsequent experiments are shown in Fig. 2.
bEBNA395 and bEBNA451 are observedto migrate as dou-bletsinsomeSDS-polyacrylamide gels,indicating
heteroge-neous forms of these proteins. All of the pure bEBNA1
mutantswere confirmed to bind
oniP
FR and DS elements and single EBNA1-binding sites by gel retardation assays(datanot shown).
The interaction of the bEBNA1 mutants with
oniP
wasexamined by electron microscopywith theoriP-containing DNAfragment(10 fmol) shown in Fig. 3 (topdiagram) and equimolar quantities (480 fmol) of the bEBNA1 truncated proteins. This ratio represents a twofold molar excess of
bEBNA1 proteins to EBNA1-binding sites. Like bEBNA1,
bEBNA39
-3309
-346, and -330-619 mediatedtheinteraction of(kda)
z I?
CDCD LO_ o
LLJa) C) sJt a) e
Do C CO CO CO IT CO
97 66
45-31
[image:3.612.69.286.82.219.2]-22
FIG. 2. Purified bEBNA1 mutant proteins. Purified bEBNA1
proteins (1.2
,ug)
weresubjectedtoelectrophoresison an SDS-12%polyacrylamide gel and visualized by Coomassie blue staining.
Molecularsizes (in thousands)are onthe left. Lanes: 39, bEBNA39;
330,bEBNA330; 346, bEBNA346; 395,bEBNA395; 451,bEBNA451;
330-619,bEBNA330
619-FR and DS elements within oniP molecules to
generate
looped structures (Fig. 3), as well as interactionsbetween
onPmolecules resulting in the formation of
multiple
DNAcomplexes (Fig. 4). Although the
frequency
with which looped DNAmolecules were inducedby
bEBNA39,
-330,and-346wasreduced
compared
with thatby
bEBNA1, the totalfraction of DNA molecules bound
by
these mutants wassimilarly reduced (Table 1).When the
frequency
oflooped
molecules was calculated as a percentage of totalprotein-bound DNA molecules, the values obtained for
bEBNA1
and
bEBNA39,
-330,
-346,and-346-619
werevery similar(Table
1, values inparentheses).Therefore,although
amino acids 7 to 346 appear to affect DNAbinding,
they
arenotrequired
for DNA looping, and neither are amino acids 620to 641.bEBNA395
mediated looping ofoniP
molecules at a lower frequency than theother bEBNA1 mutants(Table
1;
Fig.
3)
and did not induce intermolecular interactions
resulting
in the formation of multiple complexes(Table
1).
This result indicates thatbEBNA395
is impairedin itsability
tomediate interactions between FR and DS elements. ForbEBNA451,
looped andmultiple DNA structureswere notobserved ata
frequency above that seen in theabsence of
protein,
and for those that were observed, the DNA interactions did not appear to be mediated by protein (Table1).
These results were not affected by a twofold increase in theconcentrationof bEBNA451 in the binding reaction mixture or
by
adecrease in the NaCl concentration in the reaction mixture to 150 mM (data notshown). We interpret the resultsofthe electron microscopy study to indicate that a
region
of EBNA1 essential for oniPlooping lies between amino acids 346 and 450 and that this region spans amino acid 395.For allbEBNA1 mutants,onPmolecules withabEBNA1 complex on the FR element were the most
commonly
observed protein-DNA complexes(Fig.
5;
Table1).
AsreportedpreviouslyforbEBNA1, all of the bEBNA1 mutant proteins formed a single discretecomplexonthe FR element as opposed to a linear array ofbEBNA1 dimers
spanning
the20 EBNA1-binding sites of the FR element
(7).
Thisobser-vationsuggests that the bEBNAldimersbound to
adjacent
sites within the FR element areassociating
to form ahigher-order bEBNA1 complex. For the bEBNA1 mutants
that were not impaired in the ability to mediate FR-DS
interactions, binding to the DS element within
onP
wasobserved only in looped molecules (Table
1).
LinearoniP
molecules with bEBNA1 bound to both the FR and DS elements were observed with bEBNA1 mutants that aredefective
(bEBNA451)
or impaired(bEBNA395)
for FR-DS interactions.Assessment of
bEBNAl-mediated
interactions oforiPele-ments by enhanced ligation efficiency. A second
approach
taken to assess the ability ofbEBNAl mutants to mediate binding of oriP FR and DS elementswasbased on a
ligation
enhancement assay developed byMukherjee et al.
(16,
17).
For this assay, a DNA fragment in which the FR and DS elements of oniP were located closetoeach endof the DNA molecule was generated by linearizing pGEMoriP between the FR and DS elements of oriP with MluI
(Fig. 6,
top diagram). The interaction of DS and FR elementsthrough
EBNA1 thus brings the cohesive ends of the DNA molecules together, resulting in increased ligation
efficiency.
When these DNA fragments were incubated with the purebEBNAl proteins and incubation was followed by a short
(5-min) ligation reaction, bEBNAl andbEBNA39,
-330,
-346,and
-330-619
were found to induce the formation of DNAconcatemers compared with control ligation reactions
with-out bEBNAl (Fig. 6). In the reactions shown in
Fig. 6,
1J.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.140.222.566.673.2]bEBNA1
bEBNA330
* t;-.'.
X
~ /
.r
-!
bEBNA39
bEBNA346
X...,~~~~~~~~~~~~-6-D
bEBNA395
bEBNA330-619
FIG. 3. LoopedDNA structuresinducedbybEBNAlproteins boundtoboth the FR and DS elements oforiP.bEBNAlproteins were
incubated withaDNAfragment (topdiagram) containingoriP.DNAlengthsareshown in base pairs.Bar= 0.1
itm.
pmol of bEBNA1 represents stoichiometric levels ofbEBNA1 dimerstoEBNA1-bindingsites inoriP. The induc-tionof DNA concatemers inthisassayindicates that multi-ple DNAmolecules arebeingboundtogetherthrough these
bEBNAl mutants. The induction of DNAconcatemers by
bEBNA346
required more protein (4 pmol) than the other bEBNAl mutants. This increasedprotein
requirement islikely due to the unstable DNA binding properties of
bEBNA346, since gel retardation and nitrocellulose filter
binding assays have indicated that the interaction of
bEBNA346 withtheonP elements isconsiderablylessstable
than that ofbEBNAl orthe other bEBNAlmutants
(data
not shown).
bEBNA395
andbEBNA451
did not conferen-hanced ligation efficiency to the DNA molecules at any
concentrationtested. NeitherDNA concatemers,indicative
ofintermolecularinteractions,norcovalently closedcircular
products, indicative of intramolecular interactions, were
induced by these mutants, indicating that bEBNA395 and
bEBNA451
donotmediateFR-FR, FR-DS, orDS-DSinter-actions inthis assay. Identical resultswere obtainedwhen
bEBNA451
was bound to the DNA fragment in 150 asopposed
to300mMNaCl
(data
notshown).
Single
EBNAl-binding
sites support bEBNAl-mediatedDNAlooping. ThebEBNA1-mediated
looping
of DS and FR elementsinvolves bEBNA1complexes
composed
ofmulti-ple bEBNA1 dimers bound to
multiple
EBNA1-binding
sites. In order to determine whether individual EBNA1 dimersbound to
single
EBNA1-binding
sitescould interacttomediate DNA
looping,
we constructed aDNAfragnent
containingtwo
EBNA1-binding
sitesseparated by
930bp
ofplasmid DNA
(Fig.
7, topdiagram).
Whenthis DNAfrag-ment wasincubatedwith
bEBNAl
andobservedby
electronmicroscopy,loopedstructureswereobservedat a
frequency
[image:4.612.165.460.74.356.2]..
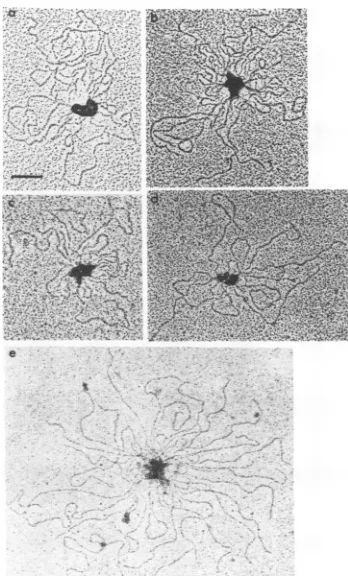
FIG. 4. Multiple
complexes
inducedby bEBNA1proteins
boundtooriP DNA.
bEBNA1 (a),
bEBNA39(b), bEBNA330 (c),bEBNA346
(d), and
bEBNA330--19
(e)wereincubated with the DNAfragmnent
containing
oriP
shown inFig. 3. Bar= 0.1iLm.
FR -I
i
H 70+1101 980 i 600 1300
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.354.528.397.685.2]3422 GOLDSMITH ET AL.
TABLE 1. Complexes formed between bEBNAlmutantproteins and oriP DNAa
bEBNA1 %observedstructure(s) No.of
mutant FR-DS FR-DS Multiple Unbound structures
(looped) (linear) FR DS complex DNA scored
None 0.5 0 0 0 0.5 99.0 200
bEBNA1 14.1 (20.7)b 0 36.5 0 17.4 32.0 241
bEBNA39 7.9(19.9) 0 18.8 0 12.9 60.4 240
bEBNA330 4.6(17.0) 0 13.8 0 8.7 72.9 253
bEBNA346 4.7(16.2) 0 18.2 0 6.1 71.0 214
bEBNA395 1.9(7.6) 1.4 21.0 0 0.5 75.1 209
bEBNA451 0.9(3.0) 3.3 22.3 0.9 0.5 72.1 211
bEBNA330619 10.1(21.3) 0 21.4 0 16.0 52.5 238
abEBNA1proteins(480 fmolasdimer)wereaddedto10 fmolof theoriP-containingDNAfragmentshown inFig.3.
bValues in parentheses indicate the percentage ofprotein-bound DNA that containedDNAloopsbetweentheFRandDSelements.
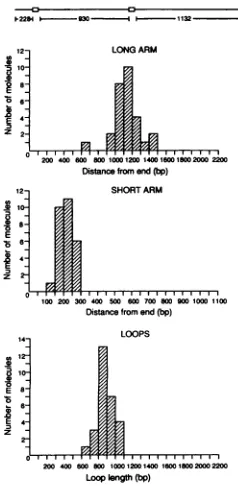
similar to that with oriP DNA (Fig. 8a to c; Table 2). Measurements of the lengths of DNAarms and loops indi-cated thatloopingoccurredthroughthetwoEBNA1-binding sites (Fig. 7). These results indicate thata bEBNA1 dimer bound to a single EBNA1-binding site can stably interact with asecond DNA-bound bEBNA1 dimerata distanceto
formaDNAloop.ThefrequencywithwhichmultipleDNA
molecules were bound togetherthroughbEBNA1,however, was substantially reduced (approximately eightfold)
com-pared with results with oriP DNA (Table 2). In DNA
molecules in which a single EBNA1 site was bound, bEBNA1dimerswereobservedtobindtoeither of thetwo
EBNA1-binding siteswith approximatelyequal frequencies
(Fig. 8d and e; Table2).
When the bEBNA1 mutants were examined for their
abilityto mediate looping of the DNA fragment containing twoEBNA1-binding sites, thefrequency of loops observed for each mutant wasvery similar to the frequency of loops inducedonoriP, with the exception of that forbEBNA395, whichwasnotobserved to induce interactions between the
two single EBNA1 sites(Fig. 8ftom; Table 2). Inkeeping
with the results with oriPDNA,
bEBNA451
was unable to mediate looping on DNA containing two EBNA1-binding sites. Since the small size of thetruncatedbEBNA1 dimerson thesingle binding sites made their detection by electron microscopy difficult or impossible
(bEBNA451
dimers are below the theoretical size limitof detection of50 kDa for this method [26]), we did not quantify linear DNA molecules bound by thebEBNA1 mutantproteins.bEBNA1
.-.... .7 . ..
bEBNA346
bEBNA39
bEBNA395
.'"'',
OrS
'
bEBNA330
..,'.X,...\*.. .
)EBNA451
bEBNA330-619
1" I,r
4, .. ;. . .. .-.
FIG. 5. bEBNA1proteins bound to the FR element ofonPDNA.bEBNA1proteins were incubated with the DNA fragment containing oriPshown inFig.3. Bar = 0.1 j±m.
J.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.74.545.421.704.2]4F
H-710-4 -- 3025 -- F
280-z
bEBNAl 39 330 346 U 395 451 330-619
IrI- A DA
L21
I41
24n i 9 d i 94 1 2 4 1 2 4 L0 2 1 2 4 1 2 4 1 2 4
] *
_ L
_ ccc
FIG. 6. Detection ofmultiple complexes by enhanced ligation efficiency. ADNA fragmentcontaining the FR and DS elements (top diagram)wasincubated with bEBNA1 proteins (1, 2,or4pmolas
dimer)orwithout protein (0). DNA lengthsareshown in base pairs. Afterthe addition of ligase, sampleswere incubated for 5 minat
15°Cpriortoethanolprecipitation andagarosegelelectrophoresis in
0.5,ug of ethidium bromideperml. Thepositions of the linear DNA fragment(L)aswellascovalently closed circular (ccc) and
concate-meric (*)ligation productsareindicated. Lane L contains the linear oniP substrate DNA thatwasused inligation reaction mixtures.
DISCUSSION
Wehave used aseries of truncationmutantsofbEBNA1
and two independent methods (electron microscopy and enhancedligationefficiency)tomap aregion of EBNA1 that
1-2241H 930 --- F1 132
12-5
10-E
° 6
E4
Z
2- 12-.2
10-9
-
-E-'b
6-
i4-I
a-E
3E
LONG ARM
12
12~0140'60ec sLooI100 200 14 It600SW 20002M0
Distance from end(bp)
SHORT ARM
D0 st
fr0o 7e00n
SO 900 0 1100Distance fromend(bp)
200 400
L0o
op1000g12D(14bp
60 20002200Looplength (bp)
FIG. 7. DNAlength measurements oflooped structuresformed between bEBNA1 and DNAcontaining twoEBNA1-binding sites. bEBNA1wasincubated withaDNAfragment (topdiagram) contain-ingtwononcontiguous18-bp EBNA1-bindingsites(open boxes).
*
7-WW
X
A;:'
A: ,': :':
FIG. 8. Complexesformed between bEBNA1 proteins and DNA containingtwoEBNA1-binding sites. The DNA fragment containing thetwoEBNA1-binding sites shown in Fig. 7wasincubated with bEBNA1 (a to e), bEBNA39 (f and g), bEBNA330 (h and i), bEBNA346a and k),orbEBNA330619(1 and m) and then prepared forelectronmicroscopy.Loopedstructures(atocand ftom) and linear structureswith bEBNA1boundtotheend(d)orthemiddle (e) EBNA1-bindingsiteareshown. Bar= 0.1 pm.
is required to mediate the interaction of the DS and FR
elements oforiP. The N-terminal 345 amino acids and the
C-terminal 22 amino acids of EBNA1were notfoundtobe requiredfor intra-orintermolecular interactions between the
FRandDS elementsineitherassay, nor weretheyrequired
to mediate interactions between single EBNA1 sites at a
distance. AbEBNA1mutantthatlackedthefirst450 amino
acids ofEBNA1 butretained the DNAbinding and
dimer-ization domain was found to be defective in mediating
interactions between the FR and DS elements in both
assays, as well as between single EBNA1 sites. These
results indicate that the DNA binding and dimerization domain of EBNA1 is not sufficient to mediate oriP DNA
loopingundertheconditions tested. Anothermutantlacking
thefirst394 amino acids of EBNA1(bEBNA395)wasfound
tohave a limited abilityto mediateDNA looping between FR andDS elements inoniP byelectronmicroscopy,but it did not mediate detectable intramolecular interactions
be-tween the FR and DS elements in the enhanced ligation
efficiency assay. This discrepancy is likely due to the
in-creased separation distance of the FR and DS elements in
the DNAfragmentused in theligationassay.bEBNA395did notmediateintermolecular interactions betweentheFR and
DS elements in eitherassay, and itwasalsonotobservedto
mediate DNA looping between single EBNA1 sites sepa-rated by 930 bp. These results suggest that the affinity of
DNA-bound bEBNA395 dimers for each other is reduced comparedwith thatofbEBNA1. Our resultsareconsistent
with thepossibilitiesthat(i) bEBNA395containsonlypartof
the domain that mediates the interaction of DNA-bound
protein:
(pmol)
L _
ccc _m
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.64.302.75.275.2] [image:6.612.333.550.76.330.2] [image:6.612.128.247.448.691.2]3424 GOLDSMITH ET AL.
TABLE 2. Complexesformed between bEBNA1 mutantproteins and DNA containing two EBNA1-binding sitesa
% observedstructure(s) No. of
bEBNAlbEBNAl
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~structures
mutant Two sites Two sites Middle End Multiple Unbound scored
(looped) (linear) site site complex DNA
None 0.5 0.5 1.9 0.5 0.5 97.1 209
bEBNA1 12.1 3.5 18.2 15.2 2.2 48.8 231
bEBNA39 9.8 224
bEBNA330 3.3 211
bEBNA346 3.0 202
bEBNA395 0 209
bEBNA451 0.5 203
bEBNA330619 8.5 200
a bEBNAl proteins (1.4 pmol asdimer)wereaddedto20 fmol of theDNAfragmentcontainingtwosingleEBNA1sites shown inFig.7.Samplescontaining the bEBNAl truncationmutants wereonly scored for looped DNA molecules(see text).
EBNA1 dimers, (ii) interactions between DNA-bound EBNA1 dimers involvetwo domains, onelocated between
amino acids 346 and 394 and a second located between
amino acids395 and 450, and (iii) theregion thatmediates
interactions betweenDNA-bound EBNA1dimersis located between aminoacids 346and394,but in its absenceasecond
region between amino acids 395 and 450 mediates limited
homologous interactions. Allof the bEBNA1proteinsused
inthisstudyarecomposed solelyof EBNA1sequences(with
theexception ofbEBNA451, which containstwoadditional
amino acidsattheNterminus)andwerepurifiedtoapparent homogeneity to eliminate the possibility that protein
se-quencesfused tobEBNA1 mutants orproteins copurifying
with bEBNA1 proteinsmay beresponsible forthe
interac-tions thatwehave observed.
Milman and Hwang (14a)hadpreviouslyreportedthatan
E. coli-produced fusion protein containing EBNA1 amino acids 450 to 641 and 36 amino acids from the X N protein
caused intermolecular interactions between synthetic EBNA1-binding sites at a low frequency. The equivalent EBNAl fragment (bEBNA451) lacking the X N sequences
and produced in insect cells was not found to mediate
intermolecularinteractions betweenEBNA1-bindingsites in
anyofourassays. Thisdiscrepancycould be duetoanyor
all of thefollowing possibilities: (i)theANproteinsequence
in the EBNA1 fusion protein mediates the interaction of DNAfragments observed by Milman and Hwang, (ii)
differ-encesinposttranslational modificationsof the EBNA1 frag-mentproducedinE. coli and insect cells affect itsabilityto
mediate DNA interactions, and (iii) the protein-mediated
DNAinteractions detectedbytheagarosegel
electrophore-sisassayofMilman andHwangareofanaturedifferentfrom thosedetectedbytheassayspresentedhere. Toaddress the
lastpossibility,wehave testedpartially purified preparations
of theE. coli-produced X N-EBNA1 fusion protein (kindly
provided by David Mackey and Bill Sugden) inourenhanced
ligation efficiencyassayandfound that the fusion protein did notmediate inter-orintramolecular interactions betweenFR
and DS elementsin thisassay(datanotshown). Therefore,
the DNAinteractionsobserved by Milmanand Hwangtobe mediated by the X N-EBNA1 fusion protein are clearly differentfromthebEBNA1-mediatedDNAinteractions pre-sentedhere.
As reported previously for bEBNA1, none of the bEBNA1 mutants that mediated oriP DNA looping were observedby electron microscopytobindtotheDS element within onP inthe absence of DNA looping, althoughallwere abletostablybindto DNAcontaining only theDS element
(7).LinearDNAmolecules with bEBNA1 boundtoboththe
FR and DS elements within oriP were only observed in
mutantsthatwereimpairedfor DNAlooping.These results
are consistent with the hypothesis that the interaction of
bEBNA1 dimersbound to the FRand DS elementswithin
oriP is extremely efficient, occurring concurrent with or
immediately following binding of bEBNAl to the DS
ele-ment. In contrast, the interaction of bEBNA1 dimers in solutionunder identicalconditionswas notdetected by gel filtrationorglycerolgradient analysis (6),suggestingthat the
domain(s) of EBNA1 responsible for the association of
DNA-bound EBNA1 dimers becomes exposed or altered
uponbindingof EBNA1 toDNA.
The enhanced ligation efficiency assay that we used to assessthe ability of bEBNA1 mutants to mediate
interac-tionsbetween FRand DSelementswasbasedontheligation
enhancement assaydeveloped byMukherjee et al.
(16, 17)
that was used to demonstrate the ability of the rr initiator
protein to mediate intramolecular interactionsbetween the
on 1 and on y sequencesof
plasmid
R6K. Theinteractionsthatwedetectedwith bEBNA1 inthis assaywere
intermo-lecular, resultingin the formation of DNAconcatemers.The
reason thatintramolecular associations of FR and DS ele-ments(resultingincovalentlyclosedcircularproducts)were notdetectedin this assayislikely duetothehighefficiency with which DNA molecules bound by these proteins are
sequestered into multiple complexes. Electron microscopy
data of onP DNA boundbythese bEBNA1proteinsindicate
that looped and multiple complexes were observed with
similar frequencies. However, since we have found that
multiple complexes often contain as many as 20 DNA
molecules, they account for many more DNA molecules
than do single looped molecules. Therefore, although bEBNA1 does mediate looping between the FR and DS elements in the DNA moleculeusedforligation assays(8a),
looping within a single molecule resulting in covalently
closed circular products was likely not detected because
mostof theloopedDNA molecules were bound into multiple complexes.
Our electronmicroscopy data indicate thatbEBNA1 can mediate loop formation between individual EBNA1-binding sites separated by 930 bp of DNA (spacing similar to that found between the DS and FR elements in onP). Since
EBNA1binds toitsrecognition site as a dimer (2, 6), these
results indicate that single DNA-bound EBNA1 dimers can interact tomediate DNA looping and that the formation of
largerbEBNA1 complexessuch as those formed on the FR
and DS elements isnot aprerequisiteforbEBNA1-mediated loop formation. Unlike the interaction of
bEBNA1
with onP,in whichoneelement(FR)is boundprior to interaction J. VIROL.on November 9, 2019 by guest
http://jvi.asm.org/
with the second element
(DS),
bEBNA1 was observed tobindtoeither ofthetwo
single binding
siteswithinthe DNAfragment
used for these studies withequal frequency,indi-cating
thattherewas nopreferential
order offillingof thesetwosites. This resultwas
expected,
since thetwoEBNA1-binding
sitesareidentical.Like theinteractionsobservedononiP DNA,
thebinding
ofbEBNA1toboth of thesinglesites withinasingle
molecule in the absence ofloopingwasrare,indicating
the efficient intramolecular interaction of the DNA-bound bEBNA1 dimers. Intermolecular associationsbetween
single
DNA-bound bEBNA1 dimers weresignifi-cantly
reducedcompared
with intermolecular interactions between bEBNA1complexes
formed on DS and FR ele-ments. This result suggests that theaffinity
ofsingle
DNA-boundbEBNA1
dimers for each other is less than that oflarger
bEBNA1complexes
and/or
that each DNA-boundbEBNA1 dimer can be involved in
only
onehomologous
interactionat adistance. Stable intermolecular interactions
between
single
DNA-bound EBNA1 dimers and EBNA1complexes
formedonthe FR element have beenpreviously
reported (14).
Our results suggest that at least part of the
region
ofEBNA1
responsible
for thehomologous
association ofDNA-boundEBNA1 dimersmapsbetween amino acids 346 and 394 and that additional amino acids between
positions
395 and 450 are also involved. This
region
of EBNA1 includes a basicdomain,
a nuclear localizationsignal,
aserine-rich sequence
containing
one or morephosphoryla-tion sites
(18),
andaproline-arginine-rich
sequencespanning
aminoacid395. Weare
currently investigating which,
if any, of these sequences areimportant
for the interaction ofDNA-bound EBNA1 dimers at a distance. At present, we
cannotexclude the
possibility
that theregion
of EBNA1 thatwe have found to mediate DNA
looping
in our bEBNA1 truncationmutantsmaynotberequired
to mediatelooping
in the contextof the
full-length
EBNA1protein.
Thispossi-bility
will be addressed withinternally
deleted andpoint-mutated EBNA1
proteins.
The interaction of distant DNA elements
through
aninitiator
protein by
a DNAlooping
mechanism has beenpreviously
demonstratedtobeessential forreplication
fromal
and03
origins
inplasmid
R6K andcanbe reducedfour-toeightfold by
asingle
amino acidchange (proline
toleucine)
inthe initiator
protein (15).
Similarly,
wepredict
that thedeletion of amino acids essential forEBNA1-mediatedonP
looping
willeliminatetheability
ofEBNA1toactivateDNAreplication
fromonP.
In this respect, it isinteresting
to consider the mutationalanalysis study
ofEBNA1conductedby
Yates and Camiolo(28),
inwhich,
of the amino acids in theregion
ofEBNA1that wehave foundtobe involved in oriPlooping, only
those between amino acids 346 and384,
392 and
396,
and 403 and 432 have not been shown to be nonessentialfor transientreplication
fromonP.Therefore,
ifEBNA1-mediated
looping
ofonPis essential forreplication,
amino acids essential for
looping
would beexpected
tomapwithin these areas.
ACKNOWLEDGMENTS
Weare
grateful
toKlaus Schultes forhelp
withsample
prepara-tion for electronmicroscopy.
We alsothank JimSmiley
and Mike O'Donnell for criticalreading
of themanuscript.
This workwas
supported by
agrant from the National Cancer Institute of Canada andby
aMedical Research CouncilofCanadastudentship (to K.G.).
L.F.isaResearch Scientist of the National Cancer Institute of Canada.REFERENCES
1. Adams,A. 1987. Replication of latent Epstein-Barr virus ge-nomes.J. Virol. 61:1743-1746.
2. Ambinder, R. F., M. Mullen, Y. Chang, G. S. Hayward, and S. D.Hayward. 1991. Functional domains of Epstein-Barrvirus nuclearantigenEBNA-1. J. Virol. 65:1466-1478.
3. Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dyebinding. Anal. Biochem. 72:248-254. 4. Chattoraj,D.K.,R.J. Mason, and S. H. Wickner. 1988. Mini-Pl
plasmid replication: the autoregulation-sequestration paradox. Cell52:551-557.
5. DePamphilis, M. L. 1988. Transcriptional elements as compo-nentsofeukaryotic originsof DNA replication. Cell 52:635-638.
6. Frappier, L.,and M. O'Donnell. 1991. Overproduction, purifi-cation and characterization ofEBNA1, the originbinding pro-tein ofEpstein-Barr virus. J. Biol. Chem. 266:7819-7826.
7. Frappier, L., and M. O'Donnell. 1991. Epstein-Barr nuclear
antigen 1 mediates a DNA loop within the latent replication origin of Epstein-Barr virus. Proc. Natl. Acad. Sci. USA 88:
10875-10879.
8. Frappier, L.,and M.O'Donnell.1992. EBNA1distortsonP, the Epstein-Barrvirus latent replication origin. J. Virol. 66:1786-1790.
8a.Frappier, L., and M. O'Donnell. Unpublishedresults. 9. Gahn, T. A., and C. L. Schildkraut. 1989. The Epstein-Barr
virus origin of plasmid replication, onP, contains both the
initiation and termination sites ofDNAreplication. Cell 58:527-535.
10. Heintz,N.H.,L.Daily,P.Held, and N. Heintz. 1992.Eukaryotic
replicationoriginsaspromotersofbidirectionalDNAsynthesis. TrendsGenet. 8:376-381.
11. Koff, A., J. F. Schwedes, and P. Tegtmeyer. 1991. Herpes simplexvirus origin-bindingprotein (UL9) loops and distorts the viralreplicationorigin. J. Virol. 65:3284-3292.
12. Krysan,P.J.,S. B.Haase, and M. P. Calos. 1989. Isolation of
human sequencesthatreplicate autonomously in human cells. Mol. Cell. Biol. 9:1026-1033.
13. Matthews, K. S. 1992. DNAlooping. Microbiol. Rev. 56:123-136.
14. Middleton, T., and B. Sugden. 1992. EBNA1 can link the
enhancer element to the initiatorelement ofthe Epstein-Barr virusplasmid origin ofDNAreplication. J.Virol. 66:489-495. 14a.Milman,G.,andE. S.Hwang. 1987.Epstein-Barr virusnuclear
antigen forms a complex that binds with high concentration
dependenceto asingleDNA-bindingsite. J.Virol. 61:465-471.
15. Miron, A., S. Mukherjee, and D. Bastia. 1992. Activation of
distant replicationoriginsin vivobyDNAloopingas revealed byanovel form ofaninitiatorprotein defective in cooperativity atadistance. EMBOJ. 11:1205-1216.
16. Mukherjee, S., H. Erickson, and D. Bastia. 1988.
Enhancer-origin interaction inplasmid R6Kinvolves aDNAloop medi-atedbyinitiatorprotein.Cell 52:375-383.
17. Mukherjee, S.,H.Erickson,and D. Bastia. 1988. Detection of
DNAloopingduetosimultaneous interaction ofaDNA-binding
protein with two spatially separated binding sites on DNA.
Proc.Natl.Acad. Sci. USA 85:6287-6291.
18. Polvino-Bodnar, M., J. Kiso, and P. A. Schaffer. 1988.
Muta-tional analysis of Epstein-Barr virus nuclear antigen 1
(EBNA1).Nucleic AcidsRes.16:3415-3435.
19. Polvino-Bodnar, M.,and P.A. Schaffer.1992. DNAbindingis required for EBNA1-dependent transcription activation and DNAreplication. Virology187:591-603.
20. Rawlins,D.R.,G.Milman,S. D.Hayward,andG. S.Hayward.
1985.Sequence-specificDNAbindingof theEpstein-Barrvirus
nuclear antigen (EBNA-1) to clustered sites in the plasmid
maintenanceregion. Cell 42:859-868.
21. Reisman, D., and B. Sugden. 1986. trans Activation of an
Epstein-BarrviraltranscriptionalenhancerbytheEpstein-Barr viral nuclearantigen1. Mol. Cell.Biol. 6:3838-3846.
22. Reisman, D.,J. Yates,and B.Sugden.1985. Aputativeoriginof
on November 9, 2019 by guest
http://jvi.asm.org/
3426 GOLDSMITH ET AL.
replication of plasmids derived from Epstein-Barr virus is composed of two cis-acting components. Mol. Cell. Biol. 5:1822-1832.
23. Shah, W. A., R. F. Ambinder, G. S. Hayward, and S. D. Hayward. 1992.Binding of EBNA-1toDNAcreatesa
protease-resistant domain that encompasses the DNA recognition and dimerization functions. J. Virol.66:3355-3362.
24. Su, W., T. Middleton, B. Sugden, and H. Echols. 1991. DNA looping between the origin of replication of Epstein-Barr virus and its enhancer site: stabilization ofan origin complex with Epstein-Barr nuclear antigen 1. Proc. Natl. Acad. Sci. USA 88:10870-10874.
25. Sugden, B., and N. Warren. 1989. ApromoterofEpstein-Barr virus thatcanfunction during latent infectioncanbe transacti-vated by EBNA1, a viral protein required for viral DNA replication during latentinfection. J. Virol. 63:2644-2649.
26. Thresher, R.,andJ.Griffith. 1992. Electron microscopic visu-alization of DNA and DNAprotein complexesasanadjunctto biochemical studies. MethodsEnzymol.211:481-490.
27. Wysokenski, D.A., and J. L. Yates. 1989. Multiple EBNA1-binding sites are required to form an EBNA1-dependent en-hancer andtoactivateaminimalreplicative origin within oniP of Epstein-Barrvirus.J. Virol. 63:2657-2666.
28. Yates, J. L., and S. M. Camiolo. 1988. Dissection of DNA
replicationandenhancerfunctions ofEpstein-Barrvirus nuclear
antigen1.Cancer Cells 6:197-205.
29. Yates, J. L., and N. Guan. 1991. Epstein-Barr virus-derived plasmids replicate onlyonce percell cycle andarenotamplified
after entryinto cells. J. Virol. 65:483-488.
30. Yates, J. L.,N.Warren,and B.Sugden.1985. Stablereplication
ofplasmidsderived fromEpstein-Barrvirus in various mamma-lian cells.Nature (London) 313:812-815.
J. VIROL.