0022-538X/80/02-0866/11$02.00/0
Characterization
of
an
Extremely
Basic Protein Derived from
Granulosis
Virus
Nucleocapsidst
KATHLEEN A.TWEETEN,' LEE A. BULLA, JR.,2AND RICHARD A. CONSIGLI'*
DivisionofBiology, Sectionof VirologyandOncology,KansasStateUniversity,
Manhattan,
Kansas66506,'andU.S. GrainMarketingResearchLaboratory,Science andEducationAdministration, Manhattan,
Kansas665022
Nucleocapsids
wereisolated frompurified enveloped nucleocapsids
of Plodia interpunctella granulosis virus by treatment with Nonidet P-40. When analyzedon sodium
dodecyl
sulfate-polyacrylamide
gels,
thenucleocapsids
consisted ofeight
polypeptides. One
ofthese,
amajor
componentwithamolecularweight
of12,500 (VP12), was
selectively
extracted from thenucleocapsids
with 0.25 Msulfuric acid. Itselectrophoreticmobilityonacetic acid-ureagelswasintermediate
tothat
of cellular
histones andprotamine.
Amino acidanalysis
showed that 39%of
the amino acid residues ofVP12
werebasic: 27%werearginine
and12% werehistidine. The
remaining residues
consistedprimarily
ofserine,
valine,
andisoleu-cine.
Proteins
ofsimilar
arginine
contentalsowereextracted from thegranulosis
virus
ofPieris
rapaeand from the
nuclear
polyhedrosis
viruses ofSpodoptera
frugiperda
andAutographa californica. The basic polypeptideappeared
tobevirus
specific
because itwasfound innucleocapsids
andvirus-infected cells butnotin
uninfected cells.
VP12was notpresentinpolypeptide profiles
ofgranulosis
viruscapsids,
indicating
that itwas aninternal
or coreprotein
of thenucleocap-sids. Electron
microscopic
observationssuggested
that the basicprotein
wasassociated with the viral DNA in the form ofa
DNA-protein complex.
Granulosis
virus(GV)
and nuclearpolyhedro-sis virus (NPV) are insect viruses
belonging
tothe
family Baculoviridae.
They
arestructurally
complex
virusesconsisting of
enveloped
nucleo-capsids embedded
withinathick
matrix ofpro-tein. The
nucleocapsids
of GV and NPV aremorphologically
similar and consist ofrod-shaped capsids that
containhigh-molecular-weight,
covalently closed supercoiled
DNAmol-ecules
(1,
28,31).
Themechanisms involved in the maturation
and
assembly
ofbaculovirus
nucleocapsids
arenot
well
characterized. This is due, in part, tothe lack
of information
onthepolypeptide
com-position
of thenucleocapsids.
The structuralpolypeptides
ofnucleocapsids
fromonly
a fewNPV (12) and GV (6, 32a) isolates have been
identified.
Inourstudies on the molecular biology of the
GV which infects the Indian meal moth, Plodia
interpunctella,
we have become interested inthe mode ofpackaging of the 80 x
106-dalton
genome ofthe virus (31) into the viral capsid.
Studies with the papovaviruses have
demon-strated that
cellular
histones are tightlyassoci-ated with the viral DNA, which is also circular and
supercoiled,
in the form of DNA-proteint Contribution no.80-97-j, Kansas Agricultural Experiment Station, KansasState University, Manhattan, KS 66506.
complexes. These histones have been found to
haveimportant functions in the viral
replication
process,
including
condensation of the viralDNA (5, 7, 19, 20).
Since
a major structuralpolypeptide
of theGV
nucleocapsids,
VP12, wasobservedtohavea
molecular
weight
similartothat
of
histones,
itwasisolated
and itsbiochem-ical
properties
wereinvestigated
to determinewhether
it washistone-like.
The resultsde-scribed in this report demonstrate
that
VP12 isan
extremely
basic, arginine-rich polypeptide that can beselectively
extracted from GVnu-cleocapsids
with dilute sulfuric acid. Evidencethat this
protein
islocated
inside theviral capsidas a core component also is presented. Similar
basic
polypeptides
wereextracted fromanum-ber of other NPVs and GVs, indicating that
these
proteins
are common tobaculoviruses.MATERIALS AND METHODS
Production andpurification of virus. The GV
of P. interpunctella was produced in a laboratory
colonyof P.interpunctella larvae reared as previously
described(30).Early third-instar larvae were infected
peroswithGV,and the virus was purified by differ-entialcentrifugation, treatment with 1% deoxycholate,
and velocity sedimentation in sucrose gradients (30,
32).
Pieris rapaeGV,whichwas producedinP. rapae larvae, was obtained from R. P. Jacques (Canadian
866
on November 10, 2019 by guest
http://jvi.asm.org/
Department of Agriculture, Harrow, Ontario) in the
form of an insecticide preparation. Sodium dodecyl
sulfate (SDS) was added to a final concentration of
0.5%,and the inert ingredients were allowed to settle
out.The preparation was filtered through Whatman
no. 1 paper, and the virus was then purified by the
method utilized for P.interpunctella GV.
Autographacalifornica NPV was produced in vitro
in aSpodoptera frugiperda cell line (no.
IPLB-SF-21AE)obtained from D.L. Knudson (Yale University
School of Medicine, New Haven, Conn.). The cells
weremaintained in Grace's insect tissue culture
me-dium supplemented with 10% fetal calf serum and 0.2%
tryptose (DifcoLaboratories). The original inoculum
of A. californica NPV consisted of infected tissue
culture supernatant kindly provided by W. F. Hink
(Ohio State University, Columbus). Falcon flasks (75 cm2) were seeded with 107 cells and incubated for 24
h at27°C. The medium was removed, and the cells
wereinfected with NPV inoculum at amultiplicity of
infection of 0.01 PFU/cell. The virus was allowed to
adsorb for 1 hat room temperature with occasional
tilting of the flasks. After adsorption, 12 ml of complete medium was added to each flask. The cultures were
harvested 7 days after infection by scraping the
in-fected flasks. The cells and tissue culture fluid were collected and centrifuged at 10,000 rpm (HB-4 rotor)
for30min.The cell-freesupernatant,which contained
nonoccluded enveloped nucleocapsids, was layered
over 5ml of 30% (vol/vol) glycerol (in 0.01 M Tris-hydrochloride, pH 7.5) and centrifuged at 25,000 rpm
(SW27 rotor) for 1h.Pelleted virus was resuspended
in0.01MTris-hydrochloride, pH 8.5, and utilized for
nucleocapsid isolation. The cell pellet, which
con-tainedpolyhedra,wassuspended indistilledwater and
disrupted with aSorvallOmnimixer. The preparation
waslayered on 40 to 65% (wt/wt) sucrose (indistilled
water)gradients which were then centrifuged at 25,000
rpm(SW27 rotor) for 1 h at 10°C. The band of virus
wasrecovered from the gradients, diluted withdistilled
water, and pelleted
.by
centrifugation at 10,000 rpm(HB-4 rotor) for 30 min. The polyhedra were
sus-pended in water and stored at -20°C.
In vivo-grown S. frugiperda NPV was obtained
from E.Dougherty (U.S. Department of Agriculture,
Science, and Education Administration, Beltsville,
Md.). The occluded virus was washed with 0.5% SDS
for20min, filteredthroughWhatmanno. 1paper,and
pelletedbycentrifugationat10,000 rpm (HB-4 rotor)
for20min. Thepelletwassuspended in1 MNaClin
distilled water, incubated for30 min at room
temper-ature, andpelletedthrougha5-ml 40%(wt/wt)sucrose
(in distilled water) shelf by centrifugation at 25,000
rpm (SW27 rotor) for30min.The pelletwas
resus-pended in distilled water and layered on 40 to 65%
(wt/wt) sucrose (in distilled water) gradients which
werecentrifugedat25,000 rpm(SW27 rotor)for1h at
10°C.Theband of viruswasrecovered fromthe
gra-dient, dilutedwithdistilledwater, andcentrifugedat
10,000rpm (HB-4 rotor) toremovethe sucrose.
Puri-fiedNPV wassuspendedin water andstoredat-20°C.
Preparation of radiolabeled GV andNPV.
Ra-dioactively labeledP.interpunctella GVwasproduced
in vivoby injection of1,ul (0.5
,uCi)
of[3H]thymidine
(Schwarz/Mann) into larvae at 96 and 120 h after
infection. The GVwaspurified from injected larvae 8
days after infection.
Radiolabeled A. californica NPVwaspreparedby
growing infected cells in Grace's insect tissue culture
medium (GIBCO Laboratories) containing 10% fetal
calf serum, 0.2% tryptose, and10,uCiof[3H]thymidine
per ml.Polyhedra and extracellularenveloped
nucleo-capsids were harvested and purified as described
above.
Isolation and purification of nucleocapsids.
Nucleocapsids were isolated by treatment of
enve-loped nucleocapsids with Nonidet P-40 (Shell
Chemi-calCo.). To obtain GV enveloped nucleocapsids,
pu-rified P.interpunctella or P. rapae GV was incubated
in0.05M sodium carbonate-0.05 M NaCl, pH 10.6, for
30 min at room temperature. The dissociated virus
waslayered on 30 to 70%(vol/vol) glycerol (in 0.01 M
Tris-hydrochloride, pH 7.5) gradients whichwere
cen-trifugedat25,000 rpm(SW41rotor) for 1 hat 10°C.
The band ofenvelopednucleocapsidswasrecovered
from the gradients and centrifuged at 25,000 rpm
(SW41 rotor) for1hto removetheglycerol. A similar
procedurewasusedtoobtain NPVenveloped
nucleo-capsids, except the alkalinesolubilization consisted of
incubating the NPVs in 0.1 M sodium carbonate for 2
h at37°C. The dissociated viruswaslayeredon10 to
50% (wt/wt) sucrose in 0.01 MTris-hydrochloride, pH
7.5) gradients whichwere centrifugedat 17,500 rpm
(SW41 rotor) for 30 minat10°C. The bands of
enve-lopednucleocapsids were recovered from the gradients
and pelleted by centrifugation at 25,000 rpm (SW41
rotor) for1h. For the isolation of nucleocapsids, the
NPV andGV envelopednucleocapsids were incubated
in 1%(vol/vol) Nonidet P-40 (in 0.01 M
Tris-hydro-chloride, pH 8.5) for 30 min with stirring at room
temperature.Thenucleocapsids were separated from
thesolubilized envelope proteins by sedimentation on
30to70%(vol/vol)glycerol (in 0.01M
Tris-hydrochlo-ride, pH 8.5) gradients bycentrifugation at 30,000 rpm
(SW41 rotor) for1 h.The band ofnucleocapsids was
recovered, diluted with0.01 MTris-hydrochloride (pH
8.5), andcentrifugedat25,000 rpm(SW41 rotor) for 1
h toremoveglycerol. Freshly preparednucleocapsids
wereutilized for acid extractionorgel electrophoresis.
Capsid isolation.Nucleocapsidswereincubated in
2%Nonidet P-40-0.01MEDTA-1 M NaCl in0.01M
Tris-hydrochloride, pH 8.5, for12hat37°C (29). The
preparation wasthen sedimentedon apreformed
ce-sium chloride gradient made in 0.01 M
Tris-hydro-chloride, pH 8.5, andrangingindensity from 1.20 to
1.50g/cm3. Centrifugationwas at34,000 rpm (SW41
rotor) for2h at10°C. The visible band ofcapsidswas
recovered, diluted with0.01MTris-hydrochloride(pH
7.5), and pelleted by centrifugation at 30,000 rpm
(SW50.1 rotor) for30minat10°C.
Electron microscopy. For examination of intact
nucleocapsids or capsids, samples were placed on
Formvar-coated grids and were negatively stained
with2% uranylacetate. Fordegradation studies,
pu-rified nucleocapsids were air dried onto
Formvar-coatedgrids.Thegridswereincubated for30minin
dropletsof 0.01 M EDTA in 0.01MTris-hydrochloride
(pH 7.0), followedbya30-minincubation indroplets
of5 mM dithiothreitol in 0.01 MTris-hydrochloride
(pH 7.0).Specimenswerethen stained with 2%uranyl
on November 10, 2019 by guest
http://jvi.asm.org/
868
acetate.Gridswereexamined with aPhilips EM 201
electron microscopeat60kV.
Acidextraction. For acidextraction,
nucleocap-sids weresuspended in distilled water, andanequal
volume of cold0.5 Nsulfuric acid wasadded. After
incubationfor16hat4°C, the acid-insoluble protein
was pelleted by centrifugation at 15,000 rpm (SS34
rotor)for1h at4°C. The supernatantwascentrifuged
to insuretheremoval of acid-insolubleproteins. The
supernatantwasrecovered and mixed with4volumes
ofcold 100% ethanol. After incubationovernight at
-20°C, theprecipitatedproteinwascollectedby
cen-trifugationat10,000 rpm(SS34 rotor) for30min. The
resultingpellet, along with the acid-insoluble pellets,
waswashed two times with cold95% ethanol and dried
under a streamofnitrogen.
Acid-soluble proteins were also extracted from
whole cells, and nucleiwereobtained from P.
inter-punctella larvae. Whole cellswere prepared by
ho-mogenizing25fourth-instar larvaein 10mlof cold0.15
MNaCl-0.01 M sodium citrate-0.05 M sodium
bisul-fite, pH 7.8. The homogenate was filtered through
gauzeand centrifugedat5,500 rpm (HB-4 rotor) for
15min. The pellet of cellswassuspended in1 ml of
distilled water, and cold sulfuric acidwasaddedto a
final concentrationof0.4Nfor extraction of the tissue.
The acid-soluble protein wasthen recovered as
de-scribed above. The larvalcellhomogenateswerealso
utilized for nuclei isolation. After centrifugation at
5,500 rpm(HB-4 rotor) for 15min, the cellularpellet
wassuspended in2mlof0.15MNaCl-0.01Msodium
citrate-0.05 M sodium bisulfite-1% TritonX-100,pH
8.0.The mixturewasstirred for10min-at4°C andwas
thencentrifugedat5,500 rpm(HB-4 rotor) for15min.
The nuclearpelletwaswashedtwo timesin 0.15 M
NaCl-0.01 M sodium citrate-0.05 M sodium bisulfite,
pH 7.8, and suspended in1ml ofdistilledwater.With
a 26-gauge needle and syringe, the nuclei were
dis-rupted and were then extracted with 0.4M sulfuric
acidasdescribed above.
SDS-polyacrylamide gel electrophoresis.
Nu-cleocapsids, capsids, acid-soluble proteins, and
acid-insoluble proteins were subjected to electrophoresis
on15%SDS-polyacrylamide slab gels (1.5 by14by18
cm; model SE 500; Hoefer Scientific Instruments),
using the discontinuous buffer system of Laemmli (18).
Sampleswereprepared forelectrophoresis by boiling
for3min in 2%SDS-5% 2-mercaptoethanol-0.0625 M
Tris-hydrochloride (pH 6.8)-10% glycerol.
Electropho-resis was carriedout at 20mA/slab. Gels were stained
overnightat 0.1%Coomassiebrilliantblue R (Sigma
Chemical Co.)-50% methanol-7.5% acetic acid.
De-staining wasin50% methanol-7.5% acetic acid for 1 h
and25%methanol-1.5% acetic acid for 48 h. Molecular
weights were determined by the method of Weber and
Osborne (33), using cytochrome c (molecular weight,
11,700), chymotrypsinogen (molecular weight, 27,500),
ovalbumin(molecular weight, 43,000), and bovine
se-rum albumin (molecular weight, 68,500) (Schwarz/
Mann) asstandards. Gels were dried with an SE-540
HoeferScientificInstrumentsslabgel dryer.
Acetic acid-urea gels. Viral proteins were also
electrophoresedon 15%polyacrylamideslabgels
con-taining 6.25 M urea. Gels were prepared and
pre-electrophoresed bythe method ofPanyimand
Chalk-ley (23) to give a final pH of 3.2. Samples were pre-pared for electrophoresis by dissolving them in 0.9 N
acetic acid-10 M urea-2% 2-mercaptoethanol-10%
glycerol. The buffer was 0.9 N acetic acid, and electro-phoresis was carried out at 20 mA/slab until the methyl green tracking dye eluted. Gels were stained, destained, and dried as described above.
Amino acid analysis.Sampleswerehydrolyzed in
1.0 ml of 6 N hydrochloric acid in evacuated, sealed
tubes at 110°C for24h. After removal of the
hydro-chloric acid byanitrogen stream, amino acid analysis
wasperformed on a Beckman 120C analyzer. Cystine
and cysteine were determined as cysteic acid and methionine was determined as the sulfone after
per-formicacid oxidation (13).
Isoelectricfocusing of VP12. VP12 was acid
ex-tracted from nucleocapsids and precipitated as
de-scribedabove. The isolated protein was suspended in
1% carrier ampholytes (pH 9 to 11; Brinkmann
Instru-mentsInc.)-12.5% sucroseindeionized water.
Isoelec-tric focusing was conducted in 7.5% polyacrylamide
slab gels containing 2% carrier ampholytes (pH 9 to
11),5% urea, 0.1%lysine, and 0.1% arginine. Gels were
prefocused for 60 min at 10 mA. After sample
appli-cation, thegels were electrophoresed at 15 mA until
thevoltage reached500V andthe pH gradient formed.
The voltage was then set at 100 V and increased by 100 V every 15 min until 1,000 V was attained.
Elec-trophoresis wascontinued until the current dropped
to 5 mA.Thegelwasfixed in 20% (vol/vol)
trichloro-acetic acid for90min and washedin25%
methanol-10% acetic acid for 10 min. Staining wasovernight in
0.2%Coomassiebrilliant blue R in 45% methanol-10%
acetic acid. The pH gradient was determined on a
section ofgel removed before fixing and staining with
aDesaga/Brinkmann flat membrane glass electrode.
RESULTS
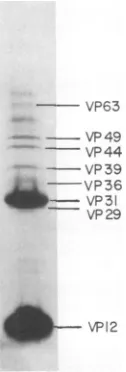
Polypeptide composition
ofnucleocap-sids. When
nucleocapsids
isolated fromthe GVof P.
interpunctella
wereelectrophoresed
onSDS-polyacrylamide gels, eight structural
poly-peptides
wereobserved(Fig.
1).The
nucleocap-sids
werecomposed primarily
of twoof
theseproteins, having molecular
weights of 12,500(VP12) and
31,000(VP31). The
remaining
pro-teinswerepresentin minoramountsand
ranged
inmolecularweight from 30,100 to 64,200. The
nucleocapsids
werefoundtobe free ofenvelope protein contamination by SDS discontinuous andgradient gel electrophoresis
andby
surfaceradioiodination studies (32a). Thus, these eight
structuralpolypeptides areunique constituents
of GV
nucleocapsids.
Acid extraction of
nucleocapsids.
Tode-termine whether any of the structural
polypep-tides were basic in nature, we treated the GV
nucleocapsids
with cold 0.25 M sulfuric acid.Thepreparation was thenseparated into
acid-soluble and acid-inacid-soluble fractions
by
centrifu-gation.
The acid-solublesupernatant
wason November 10, 2019 by guest
http://jvi.asm.org/
BASIC PROTEIN DERIVED FROM GV
869
A
B
H'
-H3- _
H25- m
H2A-H4- -VP63
---VP49 - VP44 - VP39 - ~r VP36 _ VP31
VP29
VP12
FIG. 1. SDS-polyacrylamide gels electrophoresis
ofP.interpunctella GVnucleocapsids. Nucleocapsids
wereisolatedasdescribed in thetextandprepared
forelectrophoresis by boilingin 2%SDS-5%
2-mer-captoethanol-10% glycerol. Numerical designations
refertothe molecular weight (x10-3) of each
poly-peptidedeterminedbyacomparison withmolecular
weightstandards.
covered for characterization by precipitating it
with
ethanol.
When the acid-soluble extractfrom thenucleocapsidswas electrophoresedon
polyacrylamide
gels containing 6.25 M ureaatpH 3,asinglepolypeptide specieswasobserved
(Fig. 2, lane B). To identify which nucleocapsid
polypeptide this protein corresponded to, we
analyzed it bySDS-polyacrylamide gel
electro-phoresis. A polypeptide withamolecular weight
of12,500waspresentintheSDS-polyacrylamide
gels, indicating that the acid-soluble
nucleocap-sidproteinwasVP12 (data described belw).
Theextremebasicity of thisnucleocapsid
pro-teinwasrevealed with the acetic acid-urea gel
system. In these gels, calf thymus histones,
whichrangeinmolecularweight from 11,000to
21,000, had relatively similarelectrophoretic
mo-bilities (Fig. 2, lane A). The much greater
ca-thodic mobility of the 12,500-dalton VP12 in
comparison with the histonessuggested that it
had a higher arginine content than did these
proteins. The electrophoretic mobility ofVP12
wasnot,however,asextremeasthatof thevery
arginine-rich polypeptide protamine sulfate,
which eluted from thegel with the tracking dye
(Fig. 2, arrow).
Amino acid analysis and isoelectric
fo-cusingof VP12. Forafurther evaluationof the
basicity and arginine content of VP12, it was
isolatedfrom GVnucleocapsids by acid
[image:4.514.122.185.78.264.2]extrac-tion,hydrolyzed with6 Nhydrochloric acid,and
FIG. 2. Electrophoresis of the acid extract of
nu-cleocapsids from P. interpunctella GV on an acetic
acid-ureagel.Nucleocapsids were incubated for 16 h
at4°C in 0.25Msulfuric acid. Acid-insoluble proteins
wereremoved by centrifugation at
15,(KX
rpm (SS34rotor), and theacid-soluble proteins in the
superna-tantwereprecipitated with ethanol. The pellet was
dissolved in 0.9 N acetic acid-10 M urea-2%
2-mer-captoethanol-10% glycerol and electrophoresed on
15% polyacrylamide slab gels containing 6.25 M urea
atpH 3.2. (A) Calf thymus histones; (B) acid extract
ofGV nucleocapsids. Arrow indicates the migration
of the dye marker.
TABLE 1. Amino acidanalysis of VP12
Aminoacid residue
Lysine
...Histidine...
Arginine .. ... ... .... ..
Aspartic acid ... ....
Threonine ...
Serine ...
Glutamic acid .... ... ...
Prohne ...
Halfcystine ... ... ..
Glycine
...Alanine ...
Valine .. ... ...
Methionine .. ...
Isoleucine ...
Leucine ...
Tyrosine ... ... ...
Phenylalanine ... ...
mol/100mol
1.31 12.45 26.61 0.97 0.00 16.31 0.39 4.14 0.28 1.65 1.32 16.89 0.00
13.28
0.10 5.11 0.41
analyzed for
itsamino acid
composition.
The
analysis (Table 1)
showed thatmorethanone-third,
approximately 39%,
of
the amino acidresidues
of
VP12 werebasic: 27%werearginine
and
12% werehistidine.Only
trace amountsoflysine
werepresent in thepolypeptide.
Theover-all
amino acidcomposition
of theprotein
wasrelatively
simple,
with the seven amino acidshistidine,
arginine, serine,
valine, isoleucine,
ty-rosine,
andproline
contributing
over90% of theVOL.
33,
1980r
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.514.326.386.78.235.2] [image:4.514.262.453.374.549.2]total amino acid residues. Other than the basic
amino acids, the most
prominent
residues wereserine (16.3%),
valine(16.9%),
and isoleucine(13.3%).
Only
minor amounts ofaspartic
acid,glutamic acid,
glycine,
and alanineweredetected in VP12.The isoelectricpoint of VP12 was obtained by
a direct measurement of the gel pH after
isoe-lectric
focusing.
Analkaline
pH
rangegel
wasneeded to
resolve
VP12which
wasobserved
tohave
anisoelectric
point of
approximately
9.8 to 10.0.Acid extraction
of uninfected and
GV-in-fected
whole cells and nuclei.
Acid extractswere
also
preparedfrom
uninfected andGV-infected P. interpunctella larvae to determine
whether the
arginine-rich
VP12 wasspecific
tovirus
infection.
Toanalyze
for the presence ofVP12, we
subjected
the acid extracts toelectro-phoresis in
acetic acid-urea
gels
(Fig.
3). LanesC
through
Fcontained
increasing
amounts ofthe acid-soluble
proteins
derivedfrom
unin-fected larval cell nuclei.
In no case was aprotein
asbasic as VP12
(Fig.
3,lane
B) observed.
Sim-ilar
results
wereobtained when whole
cells
fromuninfected larvae
wereacid
extracted, indicating
A
B
C
D
E F
FIG. 3. Aceticacid-ureagelelectrophoresis ofthe acidextractofGVnucleocapsidsandofnucleifrom
uninfected
P.interpunctella
larvae. For theisolationofnuclei,larvaewerehomogenizedand thecellswere
pelletedby centrifugationat5,500rpm(HB-4rotor).
The cells were incubated in 0.15 MNaCl-0.01 M sodium citrate,pH8.0, containing1% Triton X-100 and 0.05M sodiumbisulfite for 10min at40C and
then centrifugedat5,500rpm (HB-4rotor). The
nu-clearpellet was suspended in distilled water and extractedwith 0.4 Msulfuricacidasdescribed in the
legend to Fig. 2. (A) Calf thymus histones; (B) acid
extract ofGV
nucleocapsids; (,
D, E,andF)50, 100,150, and200
Pil,
respectively,
oftheacidextractfrom
uninfected
larvalnuclei.that the
arginine-rich
protein
was notanormal
constituent of the host cell
protein
composition.
On
the otherhand,
apolypeptide
with anelec-trophoretic mobility
inacetic acid-urea
gels
characteristic of VP12 was present in acid
ex-tracts
of virus-infected
cellsand nuclei
frominfected cells (data not shown). These results
suggest that VP12 is coded for
by
the viralgenome. It is also possible that VP12is a
host-contributed polypeptide that is induced and syn-thesized during virus infection.
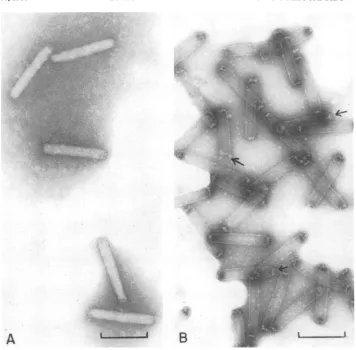
Isolation of capsids and comparison of
their
polypeptide composition with that of
nucleocapsids.
Because of itsbasicity,
it was speculated that VP12 was associated with theviral
DNAand, thus,would be
aninternalcom-ponent of the nucleocapsids. To determine
whether
this was the case,capsids,
devoid ofDNA andany core
proteins,
wereprepared.
Thiswas accomplished by treating purified
nucleo-capsids with 1 M NaCl-0.01 M EDTA in 0.01 M
Tris-hydrochloride, pH
8.5, followedby velocitysedimentationin cesium chloride gradients. The
bandofcapsids was recovered from the gradients
and examinedby electron microscopy(Fig. 4B).
The
tubular structures nolonger
took up theuranyl
acetatestain as did intactnucleocapsids(Fig. 4A),
indicating
that they had lost theirDNA core. In
addition,
whencapsids
wereiso-lated from
[3H]thymidine-labeled
nucleocapsids,no
radioactivity
wasassociated with theresult-ing
capsid
preparations, demonstrating
that the viral DNA had been removed from them (data notshown).
The
polypeptide composition
ofthe
capsids
was
then
compared
with that ofnucleocapsids
by
SDS-polyacrylamide
gel electrophoresis (Fig.
5). Present in the
nucleocapsids, (Fig.
5, lane A)were the
eight
polypeptides described
earlier,with VP31 and VP12
being
thepredominant
components.
Electrophoresed
inlane
B was theprotein extracted from the
nucleocapsids
by thesulfuric
acid treatment. It consisted of one ofthe
major nucleocapsid
structuralpolypeptides,
VP12. Lane C contained the
nucleocapsid
pro-teins that were insoluble in the acid. All of the
nucleocapsid
proteins, except VP12, wereob-served. The
SDS-polyacrylamide
gel profile ofthe
capsids
ofGV
(Fig. 5, lane D) revealed that itclosely resembled
theprofile of the acid-insol-ublenucleocapsid
proteins(lane C).
Present inthecapsid preparations were all ofthe
nucleo-capsid proteins except the basic protein, VP12,
and VP44. The absence of VP12from thecapsids
suggests that it is an internal or core protein
that was removed from the capsidsalong with
the viral DNA. The fact that VP44, an
acid-insoluble
polypeptide,
was not present in thecapsids may indicate that it also is a core protein
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.514.95.226.349.520.2]~~~~~~~~~I
e
?"-43,
>
".
9
A
.J4.
B
FIG. 4. Electronmicrographs of P.interpunctella GVnucleocapsids and capsids. Capsids were isolated by
incubating nucleocapsids in 2% Nonidet P-40-0.01 M EDTA-1 MNaCIin 0.01 MTris-hydrochloride,pH8.5,
for12hat37°C. The preparation was then centrifuged on apreformedcesiumchloride gradient (1.20 to 1.50
g/cm3in 0.01 MTris-hydrochloride, pH 8.5) at 34,000 rpm (SW41 rotor) for 2 h. The band of capsids recovered
fromthe gradient and nucleocapsids were mounted on
Formnvar-coated
grids andnegatively stained with 2%uranylacetate.Bar=200nm. (A)Nucleocapsids; (B) capsids.
or
that it
comprises
thestructures
located at theends
ofthe
capsids.
Thesestructural
compo-nents appear to
have
been "blown out" orre-moved
during
theisolation
of thecapsids (Fig.4B,
arrows)such
that the capsids look likehol-low
cylinders.
Acid extraction of
other
baculovirus
nu-cleocapsids.
It
wasof interest to
determine
whether abasicprotein similar to that
obtained
from the
nucleocapsids
of P.interpunctella
GVwas
also
astructuralcomponent
ofotherbacu-loviruses. Toinvestigate this
possibility,
wepre-pared nucleocapsids
from theGVs
of P. rapaeand S.
frugiperda
and from the NPVs of A.californica and S.
frugiperda.
Eachnucleocap-sid
preparation
wasextracted
with 0.25 Msul-furic
acid,
and theresulting acid-soluble
frac-tions were
analyzed
onacetic
acid-ureagels.
Theresults of
two suchelectrophoretic
analyses
(lanes
A toC and lanes
D toG)
are shown inFig.
6.Electrophoresed
inbothgels
asreference
proteins
werecalf thymus histones
(lanes
AandG)
and the basic
protein, VP12,
isolated
from P.interpunctella GV
(lanes
Band
F).
Anacid-extractable
protein having
afast
electrophoretic
mobility in acetic acid-urea
gels
wasobtained
from
all
ofthebaculoviruses examined. The acidextractofA.
californica
NPVnucleocapsids
pre-pared
fromnonoccluded
enveloped
nucleocap-sids consisted
ofasingle
polypeptide
(lane
C)
migrating
slightly
behind
VP12. Asimilar
pro-tein
wasacidextracted
fromnucleocapsids
iso-lated from A.
californica
polyhedra (data
notshown).
Theproteins
acidextracted
fromnu-cleocapsids
of the NPV of S.frugiperda
and
theGV of
P. rapae are shown in lanes D andE,
871
Wew
Z.
.1.
11
,i
.:I:j
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.514.78.434.68.418.2]TWEETEN,
A
B
C
D
VP44 _
VP31 _
I
A
B
C
D
E
F
G
[image:7.514.78.249.57.254.2]VP12
-JFIG. 5. SDS-polyacrylamide gel electrophoresis of
nucleocapsids, capsids, and the acid-soluble and
-insolublefractions from nucleocapsids ofP.
inter-punctella GV. Acid-solubleand -insoluble
nucleocap-sidproteinsandcapsidswerepreparedasdescribed
in thelegendstoFig.2 and4, respectively. Samples were boiled in 2%o SDS-5%
2-mercaptoethanol-1O0o
glycerolandelectrophoresedon15%polyacrylamide
slab gels. (A)Nucleocapsids; (B)acid-soluble
nucleo-capsidproteins; (C)acid-insolublenucleocapsid
pro-teins;(D) capsids.
respectively. Once
again,
themigration
of theseproteins
inthe acid-urea
gels
wascharacteristicof
arginine-rich
proteins.
Themultiple
bands ofprotein
observed in the acid extracts of these twobaculoviruses
maybe dueto amodificationof
the
basicproteins
by
phosphorylation
oracet-ylation since
SDS-polyacrylamide gel
electro-phoresis
of eachof
thesepreparations resolved
only
asingle
polypeptide species (data described
below). When
theacid-soluble
fraction obtained fromnucleocapsids
of theGV of S.
frugiperda
was
analyzed
onacetic
acid-ureagels,
italso
wasfound
tohaveanelectrophoretic mobility
simi-lartothat of
VP12(data
notshown).
SDS-polyacrylamide
gel
analysis of
bac-ulovirus
nucleocapsids
andacid-soluble
ex-tracts.
Nucleocapsids
isolated from the GV ofP. rapae and the NPV of A.
californica
andtheir acid extracts were
subjected
toelectropho-resison
SDS-polyacrylamide
gels. This was doneto
determine
the molecular weights of the basic proteins derived from these viruses so thatcom-parisons
could be made with the P.interpunc-tella GV basic protein, VP12. As shown in Fig.
7,the basic proteins extracted from P. rapae GV
(lane
B) and from A. californica NPV (lane E) were bothlow-molecular-weight
polypeptides,having
molecular weights of 12,400 and 13,000, respectively. These polypeptides, as in the caseFIG. 6. Acetic acid-ureagel electrophoresis of acid
extractsof nucleocapsids from variousbaculoviruses.
Acid extraction of nucleocapsids andelectrophoresis
wereconductedasdescribed in the legend to Fig. 2.
This figure is a composite of two separategels: Ato
C and D to G. (A and G) Calf thymus histones; (B
andF) acidextractfromnucleocapsids of P.
inter-punctellaGV; (C, D, and E) acidextractsfrom
nu-cleocapsids of A. californica NPV, S. frugiperda
NPV, and P. rapae GV, respectively.
A
B
C
D
E
qu
4
I-_I~
4.
FIG. 7. SDS-polyacrylamidegelelectrophoresis of
nucleocapsids, capsids, and nucleocapsid acid
ex-tractsfrom P. rapae GV and A. californica NPV. Acid-soluble nucleocapsid proteins and capsids were
isolated as described in the legends to Fig. 2 and
4,
respectively. This figure is a composite of two
sepa-rategels (A to B and C to F),prepared as described
in thelegend to Fig. 5. (A) P. rapaeGVnucleocapsids;
(B) P. rapae GV nucleocapsid acid extract; (C, D,
and E) nucleocapsids, capsids, and nucleocapsid
acidextract,respectively, from the NPV ofA.
califor-nica.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.514.278.464.62.233.2] [image:7.514.277.465.351.547.2]of
VP12,
weremajor
componentsof the
nucleo-capsids
(P.
rapaeGV,
lane A; A.
californica
NPV,lane C) of these viruses.
Capsids also
wereprepared from
nucleocapsids
of thesebaculovi-ruses, and their protein composition was
deter-mined. Like
P.interpunctella GV capsids, they
lacked
thearginine-rich
protein (A.
californica
NPV,
lane
E; P. rapae,data
notshown). These
results
suggestthat
alow-molecular-weight,
ex-tremely basic
coreprotein is characteristic of the
baculoviruses.
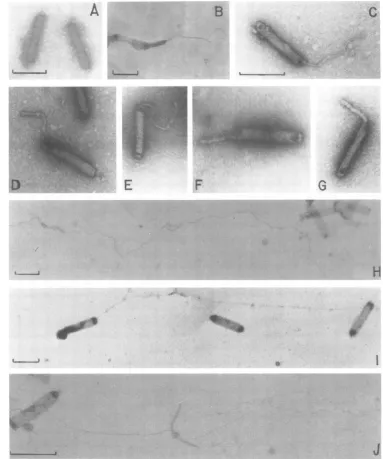
Visualization of GV
nucleoprotein
com-plex
by electron
microscopy.
Preliminary
ev-idence that the
arginine-rich
protein
isolated
from
GV
nucleocapsids
wasassociated with the
viral
DNA inthe
formof
aDNA-protein
com-plex
wasobtained
by electron
microscopy
of
dissociated
nucleocapsids.
Nucleocapsids
from
P.
interpunctella
GV
wereexposed
briefly
toachelator
(0.01
MEDTA)
andareducing
agent(0.005
Mdithiothreitol).
These
agentshave been
found
toefficiently
dissociatepolyoma virions
tocapsomeres
and
aDNA-protein
complex (4, 5).
After
treatment,the
disrupted
nucleocapsids
werestained with
uranyl
acetateand examined
by electron microscopy
(Fig.
8B toG).
Obser-vations revealed that subterminal
openings had
been
generated
inthe
nucleocapsids through
whichacompact,
rod-shaped
structure was seentoemerge.
The
compactness andstaining
prop-erties of these
structureswerecharacteristic of
a
DNA-protein
complex
(16, 17).
Exposure
of
the
nucleocapsids,
onthe other
hand,
to 1.0MNaCl resulted in the release of naked viral
DNAwhich was
visualized
aslong,
thin strands
(Fig.
8H to
J).
Itappeared
thatthe salt
treatment notonly
disrupted the capsids but also removed the
proteins associated with the viral
DNA.DISCUSSION
The results
presented in this
paperdemon-stratethatoneof the
major
structural
polypep-tides
of
nucleocapsids from
P.interpunctella
GV, VP12, is
anextremely
basic
protein.
The
basic
natureof
VP12 wasinitially
revealed
by
its acid
solubility,
a propertycharacteristic
ofbasic
proteins
such as histones orprotamines
(14).
WhenGV
nucleocapsids
weretreated with
diluted,
strongacid,
VP12wasreadily
andselec-tively extracted
from theviral
preparation.
Also indicative of the
basicity
of thenucleo-capsid
polypeptide
wereits fastelectrophoretic
mobility
inacetic acid-ureagels,
high
isoelectricpoint, and
amino acidcomposition.
Whenana-lyzed
on ureagels,
VP12migrated
to aposition
intermediatetothatof calf
thymus
histones andprotamine
sulfate,
averyarginine-rich
protein.
Because themolecular
weight
of VP12wassim-ilar
tothat of the histones, its greater cathodicmobility suggested that it was considerably more basic than the histones. This was confirmed by amino acid analysis of isolated VP12, which indicated that 27% of the residues were arginine
ascompared with 14% for thearginine-rich
his-tones (14). Also contributing to the basicity of
VP12 was histidine, which accounted for 12% of the amino acid residues.
The chemical composition of VP12 was found
to be unique. The high arginine content and
apparent
lack
of lysine are characteristic ofprot-amines, suggesting that VP12 is more prota-mine-like than histone-like. Its lack of aspartic acid and glutamic acid, along with its relatively
simple amino
acid composition, also suggests amoreprotamine-like
character.
Inaddition,ser-ine, valser-ine, and isoleucine were present in higher
proportions than are
typically
found inverte-brate and
invertebrate
histones (35). VP12,how-ever, was
distinguishable
fromboth mammalianand
insect histones
and mostprotamines
by itsunusuallyhigh histidine content and low glycine
and alanine contents.
The basic
polypeptide
appears to be astruc-turalcomponent
characteristic
of thebaculovi-ruses.
Nucleocapsids
of all of the NPVs and GVsexamined inthe present study contained an
acid-extractable
polypeptide
thatexhibited fastelec-trophoretic mobility
in acetic acid-urea gels.Electrophoretic
analysis onSDS-polyacryl-amide gels revealed that, like VP12, the basic
proteins
extractedfrom the
other baculoviruseswere of low molecular weight, ranging from
12,400
for
P. rapaeGV
to13,000 for A.califor-nica NPV. In
addition,
the basicpolypeptides
accounted for
asubstantial
amount ofthe
pro-teinassociated
with thenucleocapsids from
thevarious baculoviruses
examined, being
themajorconstituent
ofnucleocapsids from
theGVs of
P.interpunctella
and P. rapae andthe
NPV of A.californica.
The basicproteins
present in theGV and NPV of S.
frugiperda
appear to beexceptions.
Proteinsdemonstrating
electropho-retic
mobilities similar
to that of VP12 wereobserved
inacetic
acid-ureagels
of the acidextracts from
nucleocapsids
of these viruses.However,
whenthesepreparations
wereelectro-phoresed
onSDS-polyacrylamide
gels,
noCoo-massie brilliant blue-stained bands of
protein
wereobserved. One
explanation
maybe that thearginine-rich
proteins
whicharecomponents ofthe baculoviruses that infect S.
frugiperda
areof
such low molecularweight
thatthey
elutedfrom the
gels
along
with thetracking
dye.
It is
interesting
tonotethatmostof the NPVand GV
enveloped nucleocapsids
whosestruc-tural
polypeptide
compositions
have beendeter-mined contain a
low-molecular-weight
on November 10, 2019 by guest
http://jvi.asm.org/
A
B
L
. . ...4
C
t.f6D .4
E
F
G
TV
.
H
[image:9.514.74.457.82.541.2]._
AJ
FIG. 8. Electron micrographs of intact and disrupted P. interpunctella GVnucleocapsids. (A) Intact
nucleocapsids; (BtoG)nucleocapsids air driedonFormvar-coatedgrids and then incubated indroplets of
0.01M EDTA-0.005 Mdithiothreitol in 0.01 MTris-hydrochloride,pH7.0,for30min;(HtoJ) nucleocapsids
incubated in0.01MEDTA-1 MNaClin 0.01MTris-hydrochloride,pH 8.5,for30min. All specimenswere
stained with2ouranyl acetate. Bar=200 nm.
peptide
astheirmajor
component. In thenon-occluded baculovirus of
Oryctes rhinoceros
(25)and the
enveloped
nucleocapsids
of theGV
ofPieris brassicae
(6)
and of theNPVs ofRachi-plusia
ou,Trichoplusia
ni,
andGalleriamello-nella
(8,
21),
thepredominant
polypeptide
hasamolecular
weight
of 12,000 to 12,600. For theNPVs of
Spodoptera littoralis, Tipula
palu-dosa,
andLymantria
dispar,
thisprotein
is of aslightly higher
molecularweight, ranging
from14,000to16,000
(11,
12,21).
Basedontheobser-vations made in the present
study,
it islikely
.;,
40011,
I
O.-...
on November 10, 2019 by guest
http://jvi.asm.org/
that these low-molecular-weight polypeptides
are
also arginine-rich, core-associated proteins.
Because they comprise such a significant
amount of the protein found in the
nucleocap-sids,
these proteins probably areimportant
structural
andfunctional
components of thebac-uloviruses.
Histones and
histone-like
proteins have beenisolated from a number of animalviruses.
Asso-ciated with
the DNA of papovaviruses such assimian
virus 40 (7), polyoma virus (5, 20),and
human
papillomaviruses
(9) arecellular
his-tones.
Another
DNAvirus,
adenovirus, containsfour basic proteins
inits
corewhich
resemble
arginine-rich histones. One of these adenovirus
proteins,
polypeptide
IL,
closely resembles
prot-amine
inits
arginine
contentand
extremeelec-trophoretic
mobility
(15). However,unlike the
GV-derived basic protein, it is
presentonly
as aminor
componentof
thevirion.
Histones and
protamines
areusually found
associated with DNA in the
form
ofDNA-pro-tein
complexes. From
afunctional
point of
view,
all
of the
supercoiling
present inthe
covalently
closed
DNAof
polyoma virus and simain virus
40is
accounted for
by the binding of the cellular
histones (10). The arginine-rich
histones H3 and
H4
particularly play fundamental roles in
nu-cleosome
formation and condensation of the
viral
DNA (2, 34). Thefactors
responsible for
the
supercoiling
of thehigh-molecular-weight
DNAof the
baculoviruses
are notknown.Most
likely, these
samefactors
play
a part inthe
condensation
ofthe
DNA necessaryfor
pack-aging of
the genomewithin the GV and
NPVcapsids. Protamines and other arginine-rich
pro-teins have been
observed
toreplace histones
onDNA
during late
stagesof
spermatogenesis (3)
in
anumber
of vertebrate and invertebrate
spe-cies. These
proteins bind strongly
toDNA and
have been
implicated
incondensing
DNAand
inrendering it
transcriptionally
inactive (14).
It ispossible
that thearginine-rich
proteins isolated
fromthe
baculovirus
nucleocapsids
perform
sim-ilar
functions during viral maturation.
Thefol-lowing
twolines of evidence
supporting this
the-ory were
obtained.
(i) Experiments
designed
tolocalize the basic
protein
inthe
nucleocapsid
structure
revealed
that,
although
it
wasthema-jor
constituent of thenucleocapsids,
itwasab-sent from
capsids.
This observation suggeststhat the
arginine-rich polypeptides
areinternalor core
proteins. (ii)
Electronmicroscopic
obser-vations
provided evidence
that the coreof GV
nucleocapsids
consists ofanucleoprotein
com-plex. Rupture
of the ends of thecapsids
withchelating
agentsresulted
in the releaseof
athick
fiber from within the
capsid.
Fibrillarstructuressimilar in diametertothosepresent in P.
inter-punctella GV
nucleocapsids alsohave beenre-leased
from baculoviruses exposed toalkaline
carbonate or thioglycolate (22, 26). Compact but less stable structures have been demonstratedafter disruption of NPV or GV enveloped
nu-cleocapsids by
thermal
shock (24, 27). Strandsranging in diameter from 25 to 30
nm
to a sizecharacteristic of naked duplex DNA (2.5 nm)
were observed, probably representing various
stages of DNA decondensation. Inthese earlier
studies, the factors responsible forthe
aggrega-tion
of the viral DNA were notdetermined. Thesensitivity
of the compactstructure found in P.interpunctella
GV nucleocapsidsto salt iscon-sistent
with the speculation that protein isbound
to the DNA. If this indeed isthe case, thedata
presented in this report strongly suggestthat
the protein bound to baculovirus DNA isthe extremely basic nucleocapsid polypeptide.
Experiments
arecurrently
inprogress toiso-late the
nucleoprotein complex
in anintact form
sothat its
biochemical
properties
and associatedproteins
canbe
identified and characterized.
As theproperties of the complex
areinvestigated,
insight should be
gained into the function of the
arginine-rich nucleocapsid protein.
Forexample,
is its
appearance ininfected
cells correlated with
the
condensing and
packing of the viral
DNA intocapsids?
Doesit interact with the viral
genome
only during
assembly,
ordoes
it remainassociated
with the DNAafter
uncoating
of the
nucleocapsids
and act toregulate
transcription
of the
viral DNA?
Answers to thesequestions
not
only will lead
toa betterunderstanding
of theGV infection
process, butalso
will
provide
information
onwhat appearstobeaveryunique
group
of
proteins.
ACKNOWLEDGMENTS
Thisworkwassupported byPublic Health Service grant ES02036fromthe National Institute ofEnvironmental Health Services. K.A.T. wassupported byaresearchassociateship
from the U.S. Grain MarketingResearchLaboratory, U.S. DepartmentofAgriculture,Science and Education Adminis-tration, andfromthe KansasAgricultural ExperimentStation. WethankKimberlyOsborne,DennisK.Anderson,Diane Potts, and Viola Hill for their excellent technicalassistance.
LITERATURE CITED
1. Beaton, C.D., and B. K. Filshie. 1976.Comparative ultrastructuralstudies of insectgranulosisand nuclear polyhedrosisviruses. J. Gen. Virol. 31:151-161. 2. Bina-Stein, M.,and R. T.Simpson.1977.Specific
fold-ing andcontraction of DNAbyhistones H3 and H4. Cell11:609-618.
3. Bloch,D. P. 1969. Acatalogofsperm histones. Genetics
61(Suppl.):93-111.
4. Brady,J.N.,V. D.Winston,and R. A.Consigli. 1977. Dissociation ofpolyomavirusbythe chelation of cal-cium ions found associated with purified virions. J. Virol.23:717-724.
5. Brady,J.N.,V. D.Winston,and R. A.Consigli.1978. Characterization ofaDNA-proteincomplex and
on November 10, 2019 by guest
http://jvi.asm.org/
somere subunitsderived frompolyomavirusby treat-ment withethyleneglycol-bis-N,N'-tetraaceticacid and dithiothreitol. J. Virol.27:193-204.
6. Brown, D. H., H. M. Bud, and D. C. Kelly. 1977. Biophysical propertiesof the structuralcomponentsof agranulosis virusisolated from thecabbagewhite but-terfly, (Pierisbrassicae).Virology81:317-327. 7. Christiansen, G.,T.Landers,J.Griffith,and P.Berg.
1977. Characterizationofcomponentsreleasedbyalkali disruptionofsimianvirus 40. J. Virol. 21:1079-1084. 8. Cibulsky, R.J.,J. D.Harper,and R. T. Gudauskas.
1977.Biochemicalcomparisonof virionproteinsfrom fivenuclearpolyhedrosisvirusesinfecting plusiine lar-vae(Lepidoptera:Noctuidae).J. Invertebr. Pathol. 30: 303-313.
9. Favre,M., F.Breitburd,0.Croissant,and G. Orth. 1977.Chromatin-likestructuresobtained after alkaline disruption of bovine and humanpapillomaviruses. J. Virol.21:1205-1209.
10.Germond,J.E.,B.Hirt,P.Oudet,M.Gross-Bellard,
and P.Chambon. 1975.Foldingof the DNA double helix in chromatin-likestructuresfrom simian virus 40. Proc.Natl. Acad. Sci.U.S.A. 72:1843-1847.
11. Guelpa, B., M. Bergoin, and G. Crozier. 1977. La proteined'inclusionetlesproteinesdu virion du Bacu-lovirus du diptere Tipulapaludosa (Meigen). C.R. Acad.Sci. 284:779-782.
12. Harrap,K.A.,C. C.Payne,and J. S. Robertson. 1977. The properties of threebaculoviruses fromclosely re-lated hosts.Virology79:14-31.
13. Hirs,C. H. W.1967.Determination ofcystineascysteic
acid. MethodsEnzymol.2:59-62.
14. Hnilica,L.S.1972.Chemicalpropertiesofhistones,p. 3-45.In R. C. Weast(ed.),Thestructureandbiological
functionof histones.CRCPress,Cleveland,Ohio. 15. Hosokawa, K., andM. T. Sung. 1976. Isolation and
characterization ofan extremely basic protein from adenovirus type5.J. Virol.17:924-934.
16. Kierszenbaum,A.L.,and L. L. Tres.1975.Structural and transcriptional features of the mousespermatid genome. J.Cell Biol. 65:258-270.
17. Kleinschmidt, A.,and R. K. Zahn.1960.Morphologie
geloster Deoxyribonucleinsaure-Praparate und einige ihrerEigenschafter in Oberflachen-Mischfilmen. Proc. 4th Int. Congr.Electron Micros. (Berlin), p. 115-118. 18. Laemmli, U. K. 1970. Cleavage ofstructuralproteins during the assembly ofthehead ofbacteriophageT4. Nature(London) 227:680-685.
19. Mackay,R.L., andR.A.Consigli.1976.Earlyevents in polyoma virus infection: attachment, penetration, and nuclear entry.J.Virol.19:620-636.
20. McMillen,J., and R. A.Consigli. 1974. Characterization ofpolyoma DNA-protein complexes. I. Electrophoretic identification of the proteins in a nucleoprotein complex isolated frompolyoma-infected cells. J. Virol. 14:1326-1336.
21. Merdan, A.,L.Crozier,J.C.Veyrunes, and G.
Cro-zier.1977.Etudecomparee des proteines des polyedres etdesvirions de trois isolats de Baculovirusde Spodop-teralittoralis.Entomophaga 22:413-420.
22. Monsarrat,P.,B.Revet, and I. Gourevitch. 1975. Mise enevidencestabilisation et purification d'une structure nucleoproteique intracapsidaire chez le Baculovirus d'Oryctes rhinoceros L. C.R. Acad. Sci. 281:1439-1442. 23. Panyim, S., and R. Chalkley. 1969. High resolution acrylamide gel electrophoresis of histones. Arch. Bio-chem.Biophys. 130:337-346.
24. Pawar, V. M. 1975. Structure, development and physi-cochemicalproperties of the nuclear polyhedrosis virus ofSpodoptera litura (Fabricius) and associated bio-chemicalchanges in the hemolymph of the host. Ento-mol.Newsl. 5:14-15.
25. Payne, C. C., D. Compson, and S. M. DeLooze. 1977. Properties of thenucleocapsids of a virus isolated from Oryctesrhinoceros. Virology 77:269-280.
26. Ponsen, M. B. 1965.Electron microscopy of DNA-cores innuclear polyhedral viruses. Neth. J. Plant Pathol.71: 54-56.
27. Shvedchikova, N. G.,V.P.Ulanov, and L. M. Tar-asevich. 1969.Structure of thegranulosisvirus of Si-berian silkwormDendrolimus sibiricus Tschetw. Mol. Biol. 3:283-287.
28. Summers, M. D., and D. L. Anderson. 1973. Character-ization ofnuclearpolyhedrosis virus DNAs. J. Virol. 12:1336-1346.
29. Summers, M. D., and G. E. Smith. 1978. Baculovirus structuralpolypeptides. Virology 84:390-402. 30. Tweeten, K. A., L. A. Bulla, Jr., and R. A. Consigli.
1977. Isolation and purification of a granulosis virus frominfected larvae of the Indian meal moth,Plodia interpunctella. Appl. Environ. Microbiol. 34:320-327. 31. Tweeten, K. A., L. A. Bulla, Jr., and R. A. Consigli. 1977.Supercoiled circular DNA of an insect granulosis virus.Proc. Natl. Acad. Sci. U.S.A.74:3574-3578. 32. Tweeten, K. A., L. A. Bulla, Jr., and R. A. Consigli.
1978.Characterization of an alkaline protease associ-ated withagranulosis virus ofPlodiainterpunctella. J. Virol.26:702-711.
32a.Tweeten, K. A., L. A. Bulla, Jr., and R. A. Consigli. 1980.Structuralpolypeptides of the granulosisvirusof Plodiainterpunctella. J. Virol. 33:877-886.
33. Weber, K., and M. Osborne. 1969. Thereliability of molecular weightdeterminations by dodecyl sulfate-polyacrylamide gel electrophoresis. J. Biol. Chem. 244: 4406-4412.
34. Weihe, A., C. VonMickwitz, K. Grade, and R. Lindig-keit.1978.Complexes of DNA with arginine-rich and slightly lysine-rich histones.Transcription and electron microscopy. Biochim. Biophys. Acta 518:172-176. 35. Yoshida, M., K. Yokotsuka, and K. Shimura. 1966.
Amino acidcomposition and heterogeneity of histones fromposterior silkgland. J.Biochem. (Tokyo) 60:586-588.