0022-538X/82/090925-10$02.00/0
CopyrightC 1982,AmericanSocietyfor Microbiology
Vol.43,No. 3
Characterization
of
c-lil,
a
Chicken Cellular Sequence
Associated with a Stock of B77 Avian Sarcoma Virus
M.BOCCARA,t N. PLUQUET, J. COLL, C. ROMMENS, AND D. STEHELIN* Laboratoire d'Oncologie Moleculaire, InstitutPasteur, 59Lille, France
Received 22 March 1982/Accepted 27 May 1982
Using biochemical methods, we have shown that a new specific sequence,
v-lil,
is associated with a given stock of B77 avian sarcoma virus (clone 9). We prepared
aDNAcomplementary tov-lilsequences, using substractive hybridizations, and
investigated the properties of this sequence. v-lilhas a genetic complexity of ca. 2,000 nucleotides and is not present in various stocks of avian sarcoma virus,
avian leukosis virus, or defective leukemia virus.v-lilis not associated with B77
avian sarcoma virus isolated from the original tumor and thus has been acquired
byinvitropassage of the virus on chicken embryo fibroblasts. A search for the
origin ofthev-lil sequence among the DNAs of different avian species has shown
that asimilarsequence,c-lil,is present in normal chicken DNA (1 to 2 copies per
haploid genome). c-lil is not highly conserved but is present in the DNA of all
chickens fromthe genus Gallus. Thec-lil sequence is transcribed at a low level (1 to3copiesper cell) in normal chicken embryo fibroblasts. The biological function, if any, of v-lil or its cellular equivalent has yet to be determined.
The genome of avian leukosis viruses¢ALVs)
hasbeen shown to contain three genes
implicat-edin the replicative functions of the virus: gag
(internal group-specific antigen); pol (reverse transcriptase); and env (envelope glycoprotein) (4). The avian sarcoma viruses (ASVs) possess
either an additional gene, src (Rous viruses
[RSVs]),
or distinct specific sequences (oncgenes) involved in the fibroblastic
transforma-tion ofhost cells (8, 16, 18, 20). Similarly, the
defective leukemia viruses (DLVs) contain
dif-ferent onc genes,accordingtotheir
pathogenici-ty (5), insertedin a deleted ALV genome (13).
Hybridization techniques have allowed stud-iesoftheoriginand evolution ofviralgenesand
their homologous cellular counterparts. It has been shownthat sequences relatedtothe
repli-cative genes of ALV are present in normalchickencellular DNA(3, 4,8, 12).These
endog-enous viral (ev) sequences, which have been
called evi to evl3 (1), have been
acquired
re-cently by chicken
cells;
indeed,
these sequenceswere shown tobeabsentinexotic chickens(4),
andAstrinetal. havebreda roosterwhich lacks
endogenous ALV (1). In contrast, the cellular
homologs of the src sequence from RSV and
onc sequences from DLV have been shown to
be highly conserved
throughout
thephylogeny
ofhighervertebrates(12,
15,
17, 20).
Until now, the only recombinant viruses
de-tPresentaddress:Departmentof ViralOncology,
Memori-alSloan-KetteringCancerCenter,NewYork,NY 10021.
scribed in the chicken were obtained by
recom-bination between exogenous viral genes and
their cellular endogenous counterparts (7,
23-25) orbetween ALV-related viral sequences and
phylogenetically stable cellular genes, which generated a highly oncogenic virus (6, 12, 13, 15, 16, 18, 22). We present here the finding that
stableassociation between aretrovirusgenome
and acellular sequence which does not appear to
behighly conserved inevolution can occur.
This sequence,calledv-lil, has been foundby
using biochemical methods(20).Weprepareda
src probe from the stock ofB77ASV clone 9.
Afterselection, we analyzedthe characteristics
ofthis cDNAsrc. We found that this probe hy-bridizedtoPragueC strain(PrC)-RSVRNAtoa
plateau value of50% and to aplateau value of
90% with the template B77 RNA. This was
unexpectedsince both strains have been
report-ed to be of similar genetic structure. A likely
explanation was that the RNA from acellular
sequence associated with the RNA from our
stock of B77 ASVwas reverse transcribed and
copurified with the cDNAsrc.
According
totheprocedure we used for
selecting
the cDNAsrcandtothekinetics we
obtained,
this shouldnotbeaheterogeneous
population
of cellularRNAs.Therefore, wedecidedto
purify
thecDNAspe-cific for this sequence. Here we describe the
purification of this probe, called cDNAtil, and themeasurementof its
complexity.
Thehybrid-ization of
cDNAj,,
withdifferent cellular DNAselucidated the
origin
of this sequence.925
on November 10, 2019 by guest
http://jvi.asm.org/
MATERIALS ANDMETHODS
Cells and viruses. ViruseswerepropagatedonC/E chicken fibroblasts (Brown Leghorn, Institute
Gus-taveRoussy, Villejuif, France).Ahigh-titerisolateof Bratislava 77 ASV (subgroup C) was originally ob-tained from P. Vogt and has been cultured after cellularrecloning (B77clone9)forseveralyears inthe laboratoryof M. Bishop (SanFrancisco, Calif.). After repeatedpassage,thisvirus consistedmainlyof trans-formation-defective (td) variants (90%o of particles; unpublished data) but retained ahigh titer of trans-forming virus (107 focus-forming units perml). This virus stockwas used for thepreparation ofcDNA,j, and willbe called B77 ASV(lit).
Weprepared another B77 ASVfromanearly pas-sage (gift of J. Huppert, Villejuif, France) of the original tumor. Other B77 ASV cultures have been providedbyH. Temin(Madison, Wis.),C.Moscovici (Gainesville, Fla.),R.Friis(Giessen,WestGermany), L. H. Wang (New York, N.Y.), and J. A. Wyke (London, England).
The other viruses were obtained as follows: the Prague B (PrB) strain of RSV from R. Junghans (Pasadena, Calif.); the Schmidt-Ruppin strain,
sub-groupD, ofRSV andreplication-defective(rd) Bryan RSV from H. Rubin(Berkeley, Calif.); PrCRSVfrom University Laboratories (Bethesda, Md.); the Carr-Zilber strain of RSV, Rous-associated virus (RAV)-0 (subgroup E), RAV-2 (B), RAV-49 (C), RAV-50(D), td B77 ASV, and td PrC RSV from P. Vogt (Los Angeles, Calif.) through M. Bishop (San Francisco, Calif.).
The following DLVs were provided by T. Graf (Heidelberg, West Germany):the avian erythroblasto-sisvirus, the fourmyelocytomatosis viruses (MC29, OKIO,MH2,andCM2),andtheavianmyeloblastosis viruses (AMV and E26).
Preparation of viral RNA.Supernatant from infected cellswasharvested,andafterclarification (9,000 x g
for10 minat40C), virions werepelleted(19,000rpm
for90minat4°CinaSpinco L19 rotor) and suspended
inSTE buffer (0.1 M NaCl, 0.001 M EDTA, 0.02 M
Tris-hydrochloride, pH 7.4)with 200 ,ug of proteinase K(Boehringer Mannheim Corp.)perml and1% sodi-umdodecyl sulfate. After digestion for10minat370C, theviralRNAwasextracted twice withSTE-saturated phenol,andtheaqueousphasewasprecipitated
over-night at -20°C with 2 volumes of ethanol in the
presenceof0.2Msodiumacetateand 50,ugofyeast RNApermlascarrier.
For quantitative analysis, the viral RNA was
dis-solved in hybridization buffer (0.6 M NaCl, 0.02 M Tris-hydrochloride [pH 7.4], 500 ,ug ofcalfthymus DNAperml).
Toprepare70SRNA, the viralpelletwasdissolved
in STE. The 70S RNA was separated from
low-molecular-weight RNAs by sedimentation through a
15 to30%o sucrose-STEgradientinanSW41rotor at
40,000rpmfor3.5 hat40C. The70S RNA peakwas
detected byUV spectralanalysis(ISCO,modelUA5), pooled, and ethanol precipitated.
Preparationof cellular DNAs. CellularDNAs from
various avianspecieswereobtainedfrom11-to15-day embryos, tissue culture cells, or erythrocytes. The
detailed procedure has been describedbyFrisbyetal.
(4).
Preparation of cellular RNA. Tissue culture cells
(>50% confluent 100-mm dishes) were suspended in
STEcontaining200,ugofproteinaseKpermland 1%
sodiumdodecylsulfate. Afterincubation for 10minat
37°C,theRNAwasextractedtwice with
STE-saturat-ed phenol and ethanol precipitated. The DNA was
spooled outand discarded. After centrifugation, the RNA pellet was dissolved in STE to a final RNA concentration of 10mg/ml.Theabsorbance at 260 nm/ absorbanceat280nmratiowasfound to be:1.95, and DNAcontamination was measured by the diphenyl-amine testto be less than 10%. The integrityof the RNAwasshownbycentrifugation througha15 to30% STE-sucrosegradientunder conditions which detect-ed therRNAprofile.
Preparation ofvirus-specific single-strandedcDNAs.
[3H]cDNA,il and [32P]cDNA,,p were synthesized by
using purifiedAMVreversetranscriptaseinareaction
containing70SB77(lil)RNAas atemplatetomakea
cDNAj,,,ortdPrB RNAtomakeacDNArep (comple-mentary to all replicative genes of td PrB RNA).
Oligomers of calfthymusDNAwereusedasprimers
(21).Thecompletereactionmixturewasessentiallyas
describedbyShanketal.(14):50 mMTris(pH 8.1);2
mMdithiothreitol; 3mM MgCl2;50 mM KCI;100 ,ug
ofactinomycinDperml;0.1 mM eachdGTP, dATP,
anddCTP; 2mg of calfthymusDNA primerperml;
280 U of AMV polymerase per ml (J. Beard, Life Science Inc.); and 20to40jigof70S templateRNA per ml inafinal volume of 500 ±lfor[3H]cDNAand 125 ,ul for [32P]cDNA. We used 3 mCi of[3H]dTTP
(0.8 mM, 60Ci/mmol, ICN Pharmaceuticals) and 0.5
to 1 mCi of [32P]dTTP (6 ,uM, 610 Ci/mmol, New
EnglandNuclearCorp.).After 90 min ofincubationat
41°C, the reaction was stopped by the addition of
sodiumdodecyl sulfate (1%finalconcentration). The mixturewasincubated for 1 hat37°C with 100 ,ugof proteinase K per ml, extracted with STE-saturated
phenol inthe presence of 200,ugofyeastRNA,and
ethanol precipitated. Thespecific activitieswere 2 x
10' cpm/,ug for [3H]cDNA and 108 cpm/p.g for
[32P]cDNA. These specific cDNAs were then
sedi-mented througha5 to20%alkaline sucrosegradient
(0.9M NaOH, 1 M NaCl,0.01 M EDTA)for 24 hat
110,000xgat240C. The cDNAs ranged in size from4
to 8S, and a size poolof 5 to 7S was used for the selection ofspecific probes. The detailed procedure for[3H]cDNAsrc preparationhas been described
previ-ously(19).
Nucleicacidhybridization.Thestandard solution for hybridization contained 0.6 M NaCl, 0.02 M Tris-hydrochloride(pH 7.4), 0.01 M EDTA, 500 p.g of calf
thymusDNA perml,2,000 cpm (0.04 ng) of[3H]cDNA
or2,000cpm(0.005 ng)of[32P]cDNA,and the appro-priate RNA or DNA in at least 10- to 1,000-fold excess. Incubationswere carried out at 68°C in glass capillaries (Brandt). The extent of annealing was de-terminedafterS1 nuclease digestion (9).
Chromatography onHAP. Chromatography of nu-cleic acids on hydroxylapatite (HAP) columns was carriedout aspreviouslydescribed (20), using 1 mg of packedHAP(Bio-GelHTP, Bio-RadLaboratories) for 200,ugofnucleicacid. For selection probes, two types ofprocedureswereused.
(i) Negative selection (recovery of single-stranded cDNA). To stabilize the hybrids, the column was maintained in a water bath at 50°C. Samples were
on November 10, 2019 by guest
http://jvi.asm.org/
c-lil, A CHICKEN CELLULAR SEQUENCE 927
loadedin 10 mMphosphate buffer (pH 6.8) containing 1.5 M NaCl. Single-stranded cDNAs were eluted by usingalineargradient of phosphate buffer, pH 6.8(0.1 to0.2 M in1.5 MNaCI).The temperature was shifted to 60'C, and the column was washed with 0.15 M phosphate buffer to elute partial hybrids. Double-stranded nucleic acids were recovered with 0.4 M phosphate buffer.
(li) Positive selection (recovery of cDNA hybridized with homologous RNA). Chromatography was con-ducted under conditions which eliminated partial hy-brids before the elution of double-stranded nucleic acids. The column was maintained at 60°C, and sam-pleswereloaded in 10 mM phosphate buffer without NaCl. Single-stranded cDNA and partial hybrids were eluted by 0.14 M phosphate buffer. In both proce-dures, we measured theradioactivity of a sample of eachfraction, tested it forS1nucleaseresistance, and then determined residual hybridization of the recov-ered single-stranded cDNA with the selective RNA.
Determination of geneticcomplexity.Thecomplexity ofcDNA,,,wasdetermined by a method described by Young et al. (26) and adapted by Saule et al. (13), based on the fact that the complexity of a given radioactive cDNA can be defined experimentally by
the Cotia ofthe hybridization kinetics between this
cDNAin excess and thehomologous nonradioactive RNA. To meet this condition, the concentration of B77(li)RNA waspreciselydetermined by a classical Crt curve with
[32P]cDNA,P.
The concentration of RNA must belower than the concentration ofcDNA,j,
tobe inDNA-driving condition, but greater than the concentration of
[32P]cDNA,,p
tobeinCrtcondition. Hybridizationswereconducted at68°Cin 0.6 MNaCl from 150 s to 32 h with 0.2 ng of[3HjcDNA,,,(10,000 cpm), 0.005 ng of[32P]cDNA,,p
(2,000 cpm),and the required amounts of viral RNA in a final volume of 1 ,ul perpoint.Determination of thermal denaturation. The prepara-tion of3H-labeled unique-sequencechicken DNA has been described by Frisby et al. (4). The DNA of normalchickencells washybridizedwith[3H]cDNA,wj
or with 3H-labeled unique-sequence chicken DNA (2,000 cpm per point) to a Cot value of 20,000
mol-sliter-1 at afinal DNA concentration of 0.3 M NaCl. A total of 20,000 cpm of [32P]cDNAr,p was
annealed to normal chicken DNA and added as a control, and thesamplewasdivided into 10aliquots, each inasealedcapillary. Thecapillarieswere incu-bated in a water bath equilibrated to the required temperature for 10min,and the percentage of cDNA
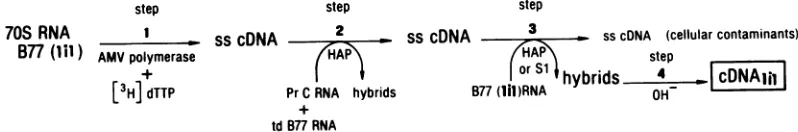
step step
70S RNA I1 sscDNA 2
B77 (lil) AMVpolymerase HA
[3H]dTTP PrC RNA hybrids
tdB77 RNA
remaining annealed to the cellular DNA at each de-fined temperature was determined by S1 nuclease digestion (9).
RESULTS
Preparation of cDNAw. DNAspecific forv-lil
was prepared by transcribing the70S RNAfrom
our stock of B77 ASV (lit) grown on chicken
embryo fibroblasts (CEFs). We then selected
[3H]cDNAwhich couldnothybridizetothe70S
RNA from PrC RSV and td B77-C ASV. The
viralstrains used for the selection werecongenic
and of the same subgroup, to eliminate possible
contamination of cDNA15, by cDNA from viral
mutatedsequences. The strategyofpreparation
ofcDNA111 was based on subtraction
hybridiza-tion (Fig. 1). After synthesis, the cDNA was
sedimented through an alkalinesucrosegradient
toeliminate cDNA smaller than 5S or largerthan
7S which would not be appropriate for correct hybridization.
(i)Negative selection. In this step, we selected
thecDNA which was notcomplementary to PrC
and tdB77-C RNAs. Single-strandedcDNA (2
,ug) was hybridized to an excess of PrC (10 ,ug)
and td B77-C RNAs (5 ,ug) under nonstringent
conditions (1.5 M NaCl at 60°C) to reach aC.tof
10
mol'sliter-1
and aCot
of 2mol's
liter-1
andthenwasseparated from hybrids by
chromatog-raphy on HAP. The single-stranded cDNA was
elutedby a lineargradient of sodium phosphate
buffer (0.1 to 0.2 M) containing 1.5 MNaCl. It
was 2% resistant to hydrolysis by S1 nuclease
and did nothybridize with PrC or td B77RNAs
(<5%S1 resistance at a Crtof20mol
s-liter-1),
but it did hybridize to B77
(lit)
RNA with aplateau of 50% hybridization, indicating the
presence of DNAcomplementary to sequences
other thanv-lilin thesingle-strandedDNA,orof
sequences too damaged to hybridize to this
RNA.
(ii) Positive selection. In this step,
single-stranded cDNA was
hybridized
to 70S RNAfrom B77 ASV
(lit)
at alow Crt (2mol-sliter-1)
understringentconditions(0.6MNaClat
68°C)
toavoid thehybridizationofnonspecificcDNA
step
ss cDNA 3 sscDNA (cellular contaminants)
HSAP
stepds
__|________t1
or
Sihybrids
4 ,cDNAjj1i
I
B77(II)RNA
OH-FIG. 1. PreparationofcDNAj,,. Single-strandedcDNA was prepared by reverse transcription of 70S RNA
fromnondefectiveB77ASV(lit)andcentrifugationthroughanalkaline sucrose gradient to select 5 to 7S cDNA (step1).Instep2,single-strandedcDNAwasannealed toanexcessof PrC and td B77 RNAs and separated from hybrids by HAP chromatography. In step 3, this single-stranded cDNA, annealed to the template RNA to eliminatecellularcontaminants,wasrecoveredashybridsafterasecond HAP chromatography (or
S1
digestion).cDNA,,,wasobtainedafteralkali treatment of thehybrids (step 4).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.58.457.553.619.2]928 BOCCARA ET AL.
withcontaminatingcellularRNA presentin the
viral RNApreparation. The
cDNA,jj
was elutedasdouble-strandhybridonHAP and after
dena-turation wasfully sensitivetoSi nuclease
diges-tion. At this step, the cDNA,,1 obtained after
hybridizationwith B77(lil)RNAunderstringent
conditions (0.6 M NaClat68°C) followedbySi
nuclease treatmentinstead of elutionon aHAP
column had the same properties. Recoveries
were similar to those described previously for
cDNAsrc (19), and
cDNAj,,
represented 7% oftheinitial total probe.
The specificity of cDNA,,1 was tested by
hy-bridization with PrC RNA, td B77 RNA, and B77 (lil) RNA (Fig. 2).
[32P]cDNArep
and[3H]cDNAs,7
were included in the reactions asinternal standards for hybridization. Only the
parental RNAhybridizedwith
cDNAji,,
whereasall three RNAs fully hybridized with cDNArep.
As expected, B77 (lit) RNA and PrC RNA
hybridized to cDNAsrc.
GeneticcomplexityofcDNAw.
According
toamodified technique of Youngetal. (13,26), the
complexity of [3H]cDNA111 can be defined by
using the
Cotj12
drawn from the hybridizationkinetics between this cDNA in excess and the
homologous nonradioactive RNA. This method
presented two advantages over the method
which used
32P-labeled
RNA (19): it was notnecessary touse a purified RNA specific to the
cDNA nor to know the ratio between helper
virusRNAand specific RNA.
Wedeterminedprecisely theconcentrationof
specific RNA and chose RNA concentrations corresponding to 20 and 40% hybridization with
[3H]cDNAwjj.
Inadditionto[3H]cDNA,j1
and B77(lil) RNA, each sample contained
[32P]cDNArep
as an internal control which would allow us to
construct a Ct curve and to make a precise
determination of the RNA concentration in the
hybridization reaction. If the RNA were in
excess over the two probes, the Crtl/2 and the
Cotj12
would beidentical,and theapparentcom-plexity would be smaller than expected. If the
[3H]cDNA,11 were in excess, the
Cot1/2
shouldbeconstant, and the hybridization plateau value
should be a function of the amount of RNA in
the reaction mixture (Fig. 3A and B). We found
a
COtj12
value of 4.5 x10-3
mols4liter-1. Since theCrt1/2
of PrB RNAwas2 x 10-2mol-sliter-1and corresponded to a complexity of 10,000
nucleotides, we determined that
cDNA1j,
had acomplexityof 2,250 nucleotides ± 10%.
Specificity
of cDNAw. We searched for thepresence of thelid sequence in RNA from ALV,
DLV, and strains of RSV otherthan the ones
used to prepare
cDNA1j,. [3H]cDNA1j,
washy-bridizedat aCrt of 5 molsliter-lunder stringent
conditions to different viral RNAs (Table 1). It is striking to see that cDNA,,1 hybridized only with
the B77 RNAused to prepare the probe.
Irre-spective of the group (RSV, ALV, DLV) orthe
subgroup (B, C, D, E) tested, the percentage of S1 resistance was no more than 8%. We
elimi-nated the possibility that these RNAs were
degraded, since
[32P]cDNArep
used in the sameexperiments extensively hybridized with all
test-edRNAs.
v-illsequence becomes associated with B77ASV
afterrepeatedpassage inCEFs.v-lil was present
FIG. 2. Hybridization of viral RNAs with
cDNA,j,.
Inparallel experiments,[3H]cDNAj,,
and[32P]cDNA,,,
or[3H]cDNAsrcand
[3cP]cDNArP
were hybridized understringent conditions (0.6 MNaCI, 68°C) todilutions of various RNAs in a volume of 7,ulfor 16 h. The extent of annealing was measured by Si nucleasedigestion.Symbols: 0,
[3H]cDNA,,,; U, [3H]cDNAsrc;
+, [32P]cDNArep. Crt(Mxsx1-l)
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.501.109.395.452.634.2]c-lil, A CHICKEN CELLULAR SEQUENCE 929
50j_ I
/#
/I
/
'I
/
...,
...a A a !..-2
4_-. L-A . Cot
10U' iloU 1o-, (Mxs x1J')
'"'I"II' 't11CCe
lo-, 2 10- 10o
FIG. 3. Geneticcomplexity ofcDNA,11. ToconstructtheCotcurve,cDNA,11 inexcess washybridizedtoB77
(lil) RNA. TheCrt curvewas determined with the concentration ofthe RNA in excesswith regard to the
concentration of the
[32P]cDNAep.
Hybridizationwasconductedat68°Cin0.6 MNaCl from150sto32 h with0.2ngof
[3H]cDNAj,i
(10,000cpm), 0.005ngof[32P]cDNA,,p
(2,000cpm),and0.1ng(A)or0.05ng(B)ofviralRNA ina1-il finalvolumeperpoint. Theextentofhybridizationwasmeasured by resistancetoS1 nuclease.
[image:5.501.51.453.61.249.2]Symbols: 0,[3HJcDNA,il; +, [32P]cDNArp.
TABLE 1. Hybridizationsof viral 70S RNA to
cDNA,j/'
b%Hybridizationto:
Virus RNA(subgroup) CDNA
cDNA,il
ALV RAV-0 (E) 74 5
PrBtd 95 4
RAV-2(B) 92 7
RAV49(C) 89 6
RAV-50(D) 91 9
B77td(C) 100 4
ASV SR D 74 8
nd B77(C) 95 100
nd PrB 100 4
CZ 91 3
rd BRY 77 8
DLV AEV(RAV-2) 84 6
MC29(RAV-2) 76 7
MH2(RAV-3) 70 12
OK10(RAV-2) 78 7
CM2 (CMAV) 75 9
AMV 61 3
E26(RAV-2) 59 6
a
Hybridization
reactions were performed understringentconditionsat afinalCrtof 5 mol-sliter'.The values werenormalizedtothefinal extentof
hybrid-ization withanhomologous RNA: 85% forcDNA,,p
with tdPrB RNA and80%oforcDNA,,,with B77(1il) RNA. The valueof 12%hybridizationwith MH2 RNA hasbeen shownbykineticanalysisof the reactionto beinsignificant.
bSRD, Schmid-Ruppin RSV,groupD;nd,
nonde-fective; CZ, Carr-Zilber RSV; BRY, Bryan RSV; AEV,avianerythroblastosisvirus.
inB77 ASVbutwasnotfoundtobeassociated
with any other tested viruses. To determine
whetherv-lilwaspresentinthe B77 ASV
isolat-ed from the original tumor or if it became
associated with B77 ASV after repeated
pas-sages in tissue culture, we prepared B77 ASV
from an early passage of the original tumor.
Tumor extracts wereinoculated into 1-day-old
chickens(C/E), and after 3 weeks,the
second-ary tumor was removed. CEFs were infected
with a filtrate from this tumor, and the viral
RNA which was produced was analyzed by
hybridization with
[3H]cDNA,jj
and[32P]cDNArep.
cDNA,,, failed to hybridize withthe viralRNA isolated from the original tumor
(Fig. 4). This result is in goodagreement with
therecentassociationof thec-lilsequencewith
one given stock of B77 ASV and was further
documented by analyzing other stocks of B77
ASV provided by different laboratories (listed
above). It was found that all viral RNAs that
were tested failed to hybridize with
cDNA,j,
(datanotshown).
ThecellularIU (c-il)sequence.v-lil couldbeof cellularorigin,since thehistoryof the B77 ASV
(lil) stockinvolvedrepeated passagesin CEFs.
To testthishypothesis, we hybridized cDNAti,
withDNAextractedfrom normal chickencells.
t32P]cDNArep
which hybridized with endoge-nous viral sequences was used as an internal standard. Inaparallel experiment,wemeasuredthereassociation kinetics ofchickenunique
se-quencesbyhybridizationof3H-labeled chicken
unique-sequence DNA with chicken DNA,
us-U
£6n
e
0
w __p
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.58.253.358.579.2]lus. Recent data fromFrisbyet al. (4) show that
,-+
evsequences are notpresentinthe DNA from allspecies
of the genus Gallus. Because we,'1+
foundc-lilin the DNAfrom the domestic chick-en,weinvestigated other DNAsfrom the genusGallus. We
hybridized
cellular DNAs to/
[3H]cDNA1il
under stringent conditions,includ-ing
[32P]cDNArep
in all the kineticanalysesasa measureofhomology withev sequences (Table' 3). As already shown(4), [32P]cDNArep
hybrid--/ ized to the DNA from a close ancestor of the
I' domestic chicken(redjungle fowl)but notto the
-
.,~'
DNA oftwootherspecies
of the genus Gallus,/'/(Sonnerat jungle/ fowland green jungle fowl). On
V ,' theother
hand,
cDNA1il
hybridized
totheDNA,
,+_, from these twospecies
tothesame extentas to0-3
10-2
10-1 10o domestic chicken DNA. From this result, weCrt(Mxs xl-1) concluded
specific
thatc-lil
was a cellular sequencefor
species
ofthegenusGallus.4. Hybridization of
cDNAil
with RNA from Relationshipofc-ilandv-il. Therelationship (isolated from the original tumor.[3H]cDNA,i,
of the DNA sequences homologous to cDNA,1zIcDNArep
were hybridized to dilutions of B77 in the chicken was analyzed by denaturing du-der stringent conditions. The percentage of plexes formed between cDNA11 and normalwasmeasuredafter
Sl
digestion. Symbols:
chicken DNA. In addition, we denatureddu-A,11;
+,
[32PIcDNArep-
plexes between3H-labeled
unique-sequencechicken DNA and chicken DNA to obtain the
same internal standard. Both tritiated Tm value of chicken unique-sequence
dissocia-hybridized to cellular DNA with the tion. As described above, in separate
experi-netics (Fig. 5) (we standardizedwith the ments [32P]cDNArep was annealed to chicken
yve obtained after hybridization with DNA and thenwasaddedtothe DNA which had
cDNAe p; data not shown). This result
showedthatc-lilwaspresentin normalchicken
cellular DNA and behaved likeunique-sequence
DNA. The
Cotj12
value was 1,300 molsliter-1,
corresponding to about 1 copy of the c-lil
se-quence perhaploidgenome.
Hybridization of cDNAil to various avian
DNAs. Studieson the evolution ofc-srcand ev sequences have shown that these sequences
have evolved differently (8). We first analyzed
the distribution of c-lil in various avian species
by hybridization ofcDNA111 with cellular DNA
understringent conditions(0.6 MNaClat68°C,
Sl analysis) and at
Cot
values greater than 104mol-s
liter-1
(10 times higher than theC0t1/2
ofunique-sequence reassociation). cDNA,,1
hy-bridized to chicken DNA but not to the DNA
from differentrepresentatives of the family
Pha-sianidae (pheasant, quail, partridge, guinea
fowl, grouse) or from other families (turkey, emu), nor to DNA from XC cells (rat cells
transformedby RSV) or calf thymus (Table 2).
Tocompare the distribution ofc-liltothat of ev sequences, we hybridized
[32P]cDNArep
to the sameDNAs, since it was already shown (4) that ev sequences were extensively present only innormal chicken DNA. In XCcells, we detected
the integratedprovirus with cDNArep
Comparative study of the origin of
RAV-0-relatedsequences andc-lilwithin the genus
Gal-c
_ U,
a)
50+
a)
a,
0
10' 103
Cot[Mxsxl'-1 1O4
FIG. 5. v-lil in normal chicken DNA. Denatured
normal chicken DNA (200 1Lg) was hybridized with
2,000 cpm of DNA per point ([3H]cDNA,i, or
3H-labeled unique-sequence chicken DNA). In both
ex-periments, [32P]cDNArep was added as an internal
standard (curves not represented). Hybridizations
wereconductedunder stringent conditionsat increas-ing Cot values to afinal Cot of 20,000 mols-liter-l.
Symbols: 0, [3H]cDNA,,,; A, 3H-labeled chicken
unique-sequenceDNA.
50
cc
.eCO)
cc
C,
FIG. 4 B77ASV and[32P] RNA un
annealing
[3H]cDN
ing the probes
samekii
Cqt
cur930 BOCCARA ET AL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.59.231.60.267.2] [image:6.501.257.446.398.588.2]c-lil, A CHICKEN CELLULAR SEQUENCE 931
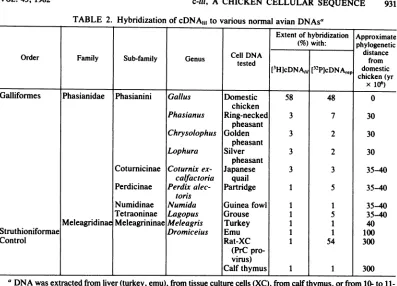
TABLE 2. Hybridization of
cDNAjj,
to various normal avianDNAsaExtent ofhybridization Approximate (%)with: phylogenetic
CellDNA
~~~~~~distance
Order Family Sub-family Genus
tested
from[3HJcDNAj,,
[32P]cDNArep
domestic chicken(yrX
106)
Galliformes Phasianidae Phasianini Gallus Domestic 58 48 0
chicken
Phasianus Ring-necked 3 7 30
pheasant
Chrysolophus Golden 3 2 30
pheasant
Lophura Silver 3 2 30
pheasant
Coturnicinae Coturnixex- Japanese 3 3 35-40
calfactoria quail
Perdicinae Perdix alec- Partridge 1 5 35-40 toris
Numidinae Numida Guinea fowl 1 1 35-40
Tetraoninae Lagopus Grouse 1 5 35-40
MeleagridinaeMeleagrininae Meleagris Turkey 1 1 40
Struthioniformae Dromiceius Emu 1 1 100
Control Rat-XC 1 54 300
(PrC pro-virus)
Calfthymus 1 1 300
aDNA wasextractedfrom liver (turkey, emu), from tissue culture cells (XC), from calf thymus, or from
10-
to 11-day-oldembryos (for all other considered species). Reaction mixtures containing denatured DNA (10 mg/ml),[3HJcDNA,1j, and
[32P]cDNA,,p
were incubated at 68°C, and the percentage of annealing was assayed by S1nucleasedigestion.The approximate phylogenetic distance from the domestic chicken was estimated by Prager etal. (10) on thebasis of the immunological relationship of transferrin and albumin.
been annealed to cDNAIil or to 3H-labeled
unique-sequence chicken DNA. Duplexes be-tween
cDNAjIt
and chicken DNA had a Tm of88.5 ± PC, which is similartothe Tm value of
unique-sequence chickenDNA dissociation (89
± 1°C).
Expression of c-il innormal CEFs. Because
c-idhas been showntobeachickensequence,we
analyzed its level of expression in CEFs. Total
cellular RNA was hybridized to
[3H]cDNAIil,
andas a control to
[32P]cDNA,,p.
We found aCrtlj2
of 500 mols Iiter-1 for the expression ofRAV-0-related sequences, which corresponds
to 80 copies per cell (the cells were chf+)
with a conventional value of
Crtj2
= 4 x l04mol*s*liter-1 for1 copy percell. For c-lil
expres-TABLE 3. Distributionofc-lilamong DNAsfrom the genus Gallus'
Extentofhybridization (%)with: Phylogenetic distance
Genus Species Cell DNA from
[3H]cDNA,i
[(32PJcDNA,P
domestic chicken(yrx 106)
Gallus Gallus Domestic chicken 58 48 0
(SPAFAS)
Redjungle fowl 54 45 0
Varius Greenjunglefowl 56 3 5
Sonnerati Sonneratjungle 57 1 0
fowl
Control Japanesequail 3 3 30
Calfthymus 1 1 300
a DNAwasextractedfromerythrocytes of exotic chickens, fromembryos(11 to15days old)of domestic chickens, orfrom calfthymus. Hybridizations wereperformed under stringentconditions atamaximumCot valueof20,000mol-s'liter-l.Theextentofannealingwasmeasured afterS1nucleasedigestion.Theapproximate phylogeneticdistance from the domestic chickenwasestimatedby Prageretal.(10).
VOL.43, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.501.60.458.510.640.2]932 BOCCARA ET AL.
80H
70O
60-U)
Z50 =40
W-U)S
_
10_
80 85 90
Temperature(OC)
FIG. 6. Thermal denaturation profiles. Normal chicken DNA wasannealedto[3H]cDNA,,,orto 3H-labeledunique-sequencechickenDNA, and Tm values were determined asdescribed in the text. A duplex performed between [32P]cDNArepand normal chicken DNA was included in each Tm determination as an internal standard. Symbols: A,
[3H]cDNAj,,;
0, 3H-labeled chicken unique-sequence DNA; +,[32P]cDNA,,p.
Numbers on the curves representthe experimental values ofTm.sion, wefound a
Crt1/2
of2 x 104 mol-sliter-1whichcorrespondsto1 to3copiesper cell(Fig.
7). Inaparallel experiment, usingtotal cellular
RNA from td B77 ALV-infected CEFs, we found the same Crt value forc-lilaswithnormal CEF cellularRNA, suggestingthat a retrovirus
10' 102 103 104
CrtMsx.I-l
FIG. 7. Transcriptionofc-lilinnormalCEFs.
To-talRNAwasextractedfromnormal CEFs and
hybrid-ized under stringent conditions to [3H]cDNA,i, and
[32P]cDNA,,p
toafinalC,t of 20,000mols-fliter' The extentofhybridizationwasmeasured by Si nucleasedigestion.Symbols: 0,[3H1cDNA,j,;+,[32P]cDNArep. ACrtl2 of4 x 104was arbitrarily taken for1 RNA
copyper averagecell.
I
B
[image:8.501.211.443.64.392.2]Crt[M.sJ-'J
FIG. 8. Analysis of viral RNA and cellular DNA after infection ofquail cells by B77 ASV (lil).Japanese quail fibroblasts were infected at low multiplicity of infection by B77 ASV (lit). The cellswere passaged three timesand were fully infected and transformed before nucleic acid extraction. (A) High-molecular-weight DNA was prepared as describedpreviously(3). The DNA was sonicated tofragments approximately 500nucleotideslong,asestimatedbyelectrophoresis in1% agarose gel withaHindllldigestionofpolyoma DNAas amarker. The DNA wasfully denaturedby boilingfor 5 min before annealingwith cDNA. The DNA (finalconcentration, 10 mg/ml) was hybridized to
[3H]cDNAj,,
(0) and[32P]cDNA,,p
(+) toreach a maximumCotvalue of 2 x 104 mol sliter-1at68°C in 0.6 M NaCl at various times in glass capillaries (Brandt). The extent of annealing was measured by S1 nucleasedigestion. (B) Thesupernatant was collected, andviral RNA was extracted as described previously (19, 21). The RNA pellet dissolved in 0.6 M NaCl buffer was further diluted and hybridized to [3H]cDNA,1,(0) and[32P]cDNA,rp
(+) at 68°Cfor 24 h in7,ul of the NaCl buffer. The extent of hybridization was measured byresistance of the cDNA to hydrolysis byS1 nuclease.lacking thev-lil sequencesisnot able toinduce
the cellular c-lil sequences in infected CEFs
(data not shown).
v-il is a transmissible sequence. Our
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.501.57.248.467.602.2]c-lil, A CHICKEN CELLULAR SEQUENCE 933
mentsfavoredthehypothesis that the B77 ASV
(lit) contained a transmissible new sequence but could not exclude the specific induction of the c-fil sequences at each infection cycle, which was
followed by the packaging of such transcripts.
We thus infected with the B77 ASV (lit) quail
fibroblasts that did not contain the c-lil
se-quences in their genome (see Table 2). The
results are shown in Fig. 8. Indeed, the v-lil
sequences were retrotranscribed, found (Fig.
8A) in the DNA (about 1 copy per haploid
genome), and transcribed as efficiently as the
other viral sequences (Fig. 8B). Thus, the
tro-pismof thev-lil sequences extends to host cells
that do not contain the homologous cellular sequences in their genomes.
DISCUSSION
Inthis paper, we have shown that a
chicken-specificcellular sequence
called
c-lilhasbecomeassociatedwith a given stock of B77 ASV. From
our stock of B77 ASV which had been
main-tained in vitro, we prepared a cDNA111 by a previously described selection procedure (19).
Its genetic complexity was measured by the
Cot11
ofhybridization between an excess of 3H-labeledcDNA111andits homologousnonradioac-tiveRNA;the
cDNA1j1
wasestimated to contain2,250 nucleotides ± 10%.
The v-lil sequence was shown to be present
onlyin our stockof B77 ASV, not in other avian
retroviruses, irrespective of their group or
sub-group: we did not find v-lil associated with
RSVs, ALVs, orDLVs, regardless of whether
they were from subgroup B, C, D, or E. We havetriedtoelucidatethe naturalhistoryof this
stockof virus. v-lilwas notassociated with the
B77 ASVisolated fromtheoriginaltumor.
Like-wise, various B77 ASV stocks from different
laboratories did not containv-lil. Wethus
con-clude that this sequence has been
acquired
re-cently by cell culture
passaging
ofaparticular
stock ofB77 ASV. Thisspecific
acquisition
isprobably a rare eventsinceitwas notobserved
in any of the other viral strains
tested,
butrecombinations ofother sequences with
retro-viralgenomesmighthaveoccurredin other viral
stocks.
Because our stock was passaged in chicken
cells, we determined whether
cDNA,j,
hybrid-ized with a chicken cellular sequence. We
showed thatourprobe
hybridized
toDNAfromuninfecteddomesticchickens. This sequenceis
absent from the DNA of various membersof the
family Phasianidae
(pheasant,
quail,
partridge,
guineafowl, andgrouse)
and from the DNAofother families
(turkey, emu)
aswell
as frommammalianDNAs(rat
XC,
calfthymus);
there-fore, c-lil is not a
highly
conserved sequence.This resultdifferentiates c-lil from c-src and all
other avianc-oncsequences sofar characterized
(12, 15-17).
To define the origin ofc-lil in chickens, we
hybridized cDNA,l to the DNA from various exotic chickens belonging to the genus Gallus.
Unlike ev
sequences,
which are found only insomespecies (4), c-lil is present in the DNA of
all tested chickens from the genus Gallus, and
thus appears to be characteristic of this genus.
The v-lil associated with B77 ASV has
changed little from its cellular counterpart,since
theTmvalue for duplexes between
cDNAIil
andchicken DNA was similar to the Tm value of
chickenunique-sequencedissociation. This
sug-gests that the percentage of mutation of v-lil
during the course ofpropagation of B77 ASV
(lit) must have been low.
What mechanism can we propose for the
appearance of c-lil during the in vitro passageof
B77 ASV (clone 9)? This might have occurred
through packaging of a major
sp;ries
of RNApresent in tissue culture cells as virus-like 30S
RNAin mouse cells (2). However, c-lil is
tran-scribed at a very low level in normal chicken
fibroblasts (1 to 3 copies per cell), and there is no increase of c-lil transcription in CEFs infected
with td B77 ALV. Moreover,c-lilacquisition by
simple packaging seems unlikely, as the B77
ASV(lit) stock has been passaged many times at
low multiplicity of infection on CEFs or even in
quailfibroblasts (that do not contain the
homolo-gousc-lil sequences in their DNA) and has still
conserved v-lil. It is also unlikely that v-lil
sequences represent an autonomously
replicat-ing virus, since their complexity of ca. 2
kilo-basesis quite small for a complete viral genome.
This suggests that v-lil is transmissible because
it is covalently linked to viral RNA from B77
ASV or its td B77 accompanying mutant.
In-deed, preliminary evidence suggests that the lil
sequences arecovalently inserted next to the src
gene in the recombinant virus (manuscript in
preparation). So far, we have no clues to the
function, ifany, conferred to our virus bythe
presence of this additional piece of nucleotide
sequences. This will require cloning these
se-quences deprived of the src transforming
se-quences and studying the biological properties
of thisvirus.
ACKNOWLEDGMENTS
We thank theresearchersalreadymentioned in the text for
providing biological materials to us. We also thank Simon Saule and Edward Stavnezer for helpful discussions, and ChristianLagroufor excellent technical assistance.
This workwassupported by grantsfromInstitutNational de la Sant6etde la RechercheM6dicale(CRL 802036, CRL
812043), Centre National de la Recherche Scientifique (AI 033695),D61egation Generaleala Recherche Scientifique et
Technique(81L0725),and the Pasteur Institute of Lille.
VOL. 1982
on November 10, 2019 by guest
http://jvi.asm.org/
934 BOCCARA ET AL.
LITERATURECITED
1. Astrln, S. M., E. G. Buss, and W. S. Hayward. 1979. Endogenous viralgenesarenon-essentialin thechicken. Nature(London)232:339-341.
2. Besner,P.,U.Obbevsky,D.Baltimore,D.Dolberg, and H. Fan.1979. Virus-like30S RNA inmousecells. J.Virol. 29:1168-1176.
3. Bishop,J. M.1978. Retroviruses. Annu. Rev. Biochem. 47:35-88.
4. Frisby, D. P., R. A.Weiss, M. Roussel,and D.Stehelin. 1979. Thedistribution of endogenous chicken retrovirus
sequencesin theDNA of Galliform birds doesnot coin-cidewith avianphylogenetic relationships. Cell 17:623-634.
5. Graf, T., and H. Beug. 1978. Avian leukemia viruses.
Interaction with theirtarget cells in vivo and in vitro. Biochim.Biophys. Acta 516:269-299.
6. Hanafusa, H. 1977. Cell transformationby RNA tumor
viruses, p. 401-458. In H. Fraenkel-Conrat and R. R. Wagner (ed.), Comprehensive virology: regulation and
genetic viral gene expression and integration, vol. 10. PlenumPublishing Corp., New York.
7. Ha1f1s, T., H. Hanas, and T. Mlyamoto. 1970.
Recoveryofanewvirus fromapparently normal chick cells by infection with avian tumorviruses. Proc. NatI. Acad.Sci.U.S.A. 67:1797-1803.
8. Hughes, S. H., F.Payvar,D.Spector, R.T.Schlmke,H. L.
Robson, G. S.Payne,J.M.Bishop, and H. E. Varmus. 1979. Heterogeneityofgenetic loci in chickens:analysis ofendogenous viral and nonviralgenes by cleavage of DNAwith restriction endonucleases. Cell 18:347-359. 9. Leong,J.-A.,A.-C.Garapln, N.Jaclkon,L.Fanshler, W.
Levnson,andJ.M.Bisbop. 1972.Virus-specific ribonu-cleic acid in cells producing Roussarcomavirus: detec-tion and characterizadetec-tion.J.Virol.9:891-902.
10. Prager, E. M., A. H. Brush, R.A.Nolan,M.Nakanishi,
andA.C. Wilson. 1974. Slow evolution of transferrinand
albumin in birdsaccordingtomicrocomplementfixation
analysis. J. Mol. Evol. 3:243-262.
11. Robinson, H. L. 1978. Inheritance and expression of chickengenesthatarerelatedtoavian leukosissarcoma
virusgenes.Curr. Top. Microbiol. Immunol.83:1-37.
12. Rousel, M., S. Saule, C. IAgrou, C.Rommuns, H. Beug, T.Graf, and D. Stebheln. 1979. Threenewtypesof viral
oncogeneof cellularorigin specific forhaematopoietic cell transformation. Nature(London)281:452-459. 13. Saule, S., M. Rosel, C.Lagrou,and D.Stehelin.1981.
Characterization of theoncogene(erb) of avian erythro-blastosis virus and its cellular progenitor. J. Virol. 38:409-419.
14. Shank, P. R., S. H. Hughes, H.J.Kung,J.E.Majors, N.
Quintrell, R. V. Guntaka, J. M. Bishop, and H. E.
J.VIROL. Varmus.1978.Mapping unintegratedaviansarcomavirus DNA:termini of linear DNA bear 300nucleotidespresent once or twice in two species of circular DNA. Cell
15:1383-1395.
15. Sheiness, D.,andJ.M.BIsbop.1979.DNA and RNA from uninfected vertebrate cells contain nucleotide sequences relatedtotheputativetransforminggeneof avian myelo-cytomatosisvirus.J. Virol.31:514-521.
16. Shlbuya,M.,T. Hanafusa, H.Hanafusa, and J. R.
Ste-phenson.1980.Homologyexists among thetransforming
sequences ofavian and feline sarcoma viruses. Proc. Natl.Acad. Sci. U.S.A.77:6536-6540.
17. Spector,D. H.,H. E.Varmus,andJ.M.Bishop. 1978.
Nucleotidesequencesrelated tothetransforminggeneof avian sarcomavirus arepresent in DNA of uninfected vertebrates. Proc. Natl. Acad. Sci. U.S.A. 75:4102-4106. 18. Stavuezer,E., D. S. Gerhard, R. C. Binari, and I.Balm.
1981. Generationoftransformingviruses in cultures of chicken fibroblasts infected with an avianleukosisvirus. J.Virol.39:920-934.
19. SteheUn, D.,R. V. Guntaka,H. E. Varmus, and J. M.
Bishop. 1976. Purification of DNA complementary to
nucleotide sequences required for neoplast
transforma-tion offibroblastsbyavian sarcomaviruses.J.Mol. Biol.
101:349-365.
20. StebeUn, D.,H. E.Varmus, J.M.Bisbhop,andP.K.Vogt.
1976. DNA relatedtothetransforming gene(s)ofavian
sarcomaviruses is present in normal avian DNA. Nature
(London)260:170-178.
21. Taylor, J. M., R. ilMmensee, and J. Summers. 1976. Efficient transcription of RNA into DNA by avian
sarco-maviruspolymerase. Biochim. Biophys. Acta 442:324-330.
22. Vlgne,R.,M. L.Breltman, C. Moscovici, and P. K.Vogt.
1979. Restitutionoffibroblast-transforming ability insrc
deletion mutants of avian sarcoma virus during animal passage.Virology93:413-426.
23. Vogt,P.K., and R. R. Frils. 1971. An avian leukosis virus related to RSV(0): properties and evidence for helper
activity. Virology 43:223-234.
24. Wang, L. H., C. C. Halpern, M. Nadel, and H. Hanafusa. 1978. Recombination between viral and cellular se-quences generates transforming sarcoma virus. Proc. NatI. Acad. Sci. U.S.A. 75:5812-5816.
25. Weiss,R.A., W. S. Mason, and P. K. Vogt. 1973.Genetic
recombinants and heterozygotes derived from endoge-nousandexogenousavian RNA tumor viruses. Virology S2:535-552.
26. Young,B. D., P. R. Harrison, R. S.Gilmour,G. D.Blrnie, A. Hell, S.Humphries,and J. Paul. 1974. Kinetic studies ofgenefrequency.II.Complexityofglobin complemen-taryDNA and its hybridization characteristics.J. Mol. Biol.84:555-568.
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG. [3H]cDNA,,,; U, [3H]cDNAsrc; RNAs RNAsSymbols: andvarious[3H]cDNAsrc +, experiments, in a volume of 7 were ,ul for hybridized 16 h](https://thumb-us.123doks.com/thumbv2/123dok_us/1454856.98150/4.501.109.395.452.634/cdnasrc-rnas-rnassymbols-andvarious-cdnasrc-experiments-volume-hybridized.webp)


![FIG. 8.Thewasbyafter0.6quailinbeforeandbufferinfectionthree(19,weight500[3H]cDNA,1,inDNAboilingto(Brandt).nucleaseDNAmaximum 7 1% [3H]cDNAj,, Analysis of viral RNA and cellular DNA infection of quail cells by B77 ASV (lil)](https://thumb-us.123doks.com/thumbv2/123dok_us/1454856.98150/8.501.57.248.467.602/thewasbyafter-quailinbeforeandbufferinfectionthree-indnaboilingto-brandt-nucleasednamaximum-analysis-cellular-infection.webp)
