Herpes Simplex Virus 1 Infection of Tree Shrews Differs from That of
Mice in the Severity of Acute Infection and Viral Transcription in the
Peripheral Nervous System
Lihong Li,aZhuoran Li,aErlin Wang,aRui Yang,aYu Xiao,aHongbo Han,bFengchao Lang,aXin Li,aYujie Xia,c,dFeng Gao,eQihan Li,f Nigel W. Fraser,gJumin Zhoua
Key Laboratory of Animal Models and Human Disease Mechanisms, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming, Yunnan, Chinaa; School of Life Sciences, Panzhihua College, Panzhihua, Sichuan, Chinab; Kunming Primate Research Center, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming, Yunnan, Chinac; Center for Drug Safety Evaluation, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming, Yunnan, Chinad; Key Laboratory of Zoonosis, Ministry of Education, College of Veterinary Medicine, Jilin University, Changchun, Chinae; Department of Viral Immunology, Institute of Medical Biology, Chinese Academy of Medicine Science, Peking Union Medical College, Kunming, Kunming, Chinaf; Department of Microbiology, Perelman School of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania, USAg
ABSTRACT
Studies of herpes simplex virus (HSV) infections of humans are limited by the use of rodent models such as mice, rabbits, and guinea pigs. Tree shrews (Tupaia belangeri chinensis) are small mammals indigenous to southwest Asia. At behavioral, anatomi-cal, genomic, and evolutionary levels, tree shrews are much closer to primates than rodents are, and tree shrews are susceptible to HSV infection. Thus, we have studied herpes simplex virus 1 (HSV-1) infection in the tree shrew trigeminal ganglion (TG) following ocular inoculation.In situhybridization, PCR, and quantitative reverse transcription-PCR (qRT-PCR) analyses con-firm that HSV-1 latently infects neurons of the TG. When explant cocultivation of trigeminal ganglia was performed, the virus was recovered after 5 days of cocultivation with high efficiency. Swabbing the corneas of latently infected tree shrews revealed that tree shrews shed virus spontaneously at low frequencies. However, tree shrews differ significantly from mice in the expres-sion of key HSV-1 genes, including ICP0, ICP4, and latency-associated transcript (LAT). In acutely infected tree shrew TGs, no level of ICP4 was observed, suggesting the absence of infection or a very weak, acute infection compared to that of the mouse. Immunofluorescence staining with ICP4 monoclonal antibody, and immunohistochemistry detection by HSV-1 polyclonal anti-bodies, showed a lack of viral proteins in tree shrew TGs during both acute and latent phases of infection. Cultivation of super-natant from homogenized, acutely infected TGs with RS1 cells also exhibited an absence of infectious HSV-1 from tree shrew TGs. We conclude that the tree shrew has an undetectable, or a much weaker, acute infection in the TGs. Interestingly, compared to mice, tree shrew TGs express high levels of ICP0 transcript in addition to LAT during latency. However, the ICP0 transcript remained nuclear, and no ICP0 protein could be seen during the course of mouse and tree shrew TG infections. Taken together, these observations suggest that the tree shrew TG infection differs significantly from the existing rodent models.
IMPORTANCE
Herpes simplex viruses (HSVs) establish lifelong infection in more than 80% of the human population, and their reactivation leads to oral and genital herpes. Currently, rodent models are the preferred models for latency studies. Rodents are distant from primates and may not fully represent human latency. The tree shrew is a small mammal, a prosimian primate, indigenous to southwest Asia. In an attempt to further develop the tree shrew as a useful model to study herpesvirus infection, we studied the establishment of latency and reactivation of HSV-1 in tree shrews following ocular inoculation. We found that the latent virus, which resides in the sensory neurons of the trigeminal ganglion, could be stress reactivated to produce infectious virus, follow-ing explant cocultivation and that spontaneous reactivation could be detected by cell culture of tears. Interestfollow-ingly, the tree shrew model is quite different from the mouse model of HSV infection, in that the virus exhibited only a mild acute infection following inoculation with no detectable infectious virus from the sensory neurons. The mild infection may be more similar to human infection in that the sensory neurons continue to function after herpes reactivation and the affected skin tissue does not lose sensation. Our findings suggest that the tree shrew is a viable model to study HSV latency.
T
he herpes simplex viruses (HSVs) (herpes simplex virus 1 [HSV-1] and 2 [HSV-2]) infect close to 80% of the population and establish lifelong latent infection in most infected individuals. The reactivation of these viruses causes symptoms ranging from skin lesions and keratitis to mostly fatal herpes simplex encepha-litis (1). Although a great amount of detail about HSV-1 primary infection or lytic infection is known, how the virus establishes, maintains, and is reactivated from its latency in neuronal cells under the surveillance of the immune system is not totally under-stood.Current animal models used include mice and rabbits for HSV-1 infection and guinea pigs for HSV-2 infection (2–5). Fol-lowing infection at the periphery of these animals, the virus repli-cates in epithelial cells and spreads to axons of the peripheral ner-vous system neurons innervating the site of infection. From here, it is transported to the nucleus of the sensory neuronal cell where it establishes latency (6). A common model for human infections involves eye inoculation followed by latent infection in sensory neurons of the trigeminal ganglion (TG) of mice or rabbits. Dur-ing mouse latency, most genes are silent except for a long,
on November 7, 2019 by guest
http://jvi.asm.org/
coding RNA called the latency-associated transcript (LAT) (7–9). This gene has an antiapoptotic effect and codes for microRNAs (miRNAs) that are important to maintain latency in the infected neuron (10, 11). Although the virion DNA is a linear double-stranded molecule, the latent viral DNA is endless (probably cir-cular) and episomal, and it adopts a chromatin structure similar to that of silent host chromatin (12–17).
Many of the approximately 80 genes in the HSV-1 genome are involved in modulating virus-host interactions, i.e., fending off or modifying the host intrinsic antiviral response, the host native immune response, the apoptotic pathway, and other host stress responses (18,19). However, due to differences in immunity be-tween rodents and humans, many of the HSV-1 genes that modify certain aspects of host immunity may not be performing normally and may result in aberrant infections in these animal models (4). Indeed, in mice, HSV-1 initiates productive infection in trigemi-nal ganglion sensory neurons following primary infection in the eye, which often leads to apoptosis of affected neurons (10). In humans, herpes outbursts do not result in the loss of sensation in the skin at and around the lesions, a fact that the current rodent models cannot explain (6,20).
The tree shrew is a small animal (a prosimian primate) indig-enous to southwest Asia and has been used to study myopia and depression (21–24). The recently published complete genome and transcriptome of the tree shrew suggest that it is closely related to primates (25,26). This, and the fact that tree shrews are known to be susceptible to HSV-1 and HSV-2 infections, opens the possi-bility to use the tree shrew as a surrogate for primate models of HSV pathogenesis (27,28). Here, we have investigated and char-acterized HSV-1 infection in the TGs of the tree shrew and con-firmed that HSV-1 establishes latency in the TG after ocular inoc-ulation. The latent viral DNA could be detected by PCR, and the LAT transcript could be detected byin situhybridization in the TG. Explant cocultivation of latently infected TG caused reactiva-tion of the virus. The infected tree shrew also shed live virus spon-taneously in tears. However, tree shrews differ from mice in the acute phase of HSV-1 infection in that tree shrew TGs lack de-tected amounts of HSV-1 replication and infectious HSV-1 virus, while mouse TGs contained large amount of infectious HSV-1 virus and express high levels of viral proteins. Consistent with this, we found that the expression profile of key HSV-1 genes from infected tree shrew TGs differs significantly from that of infected mice. Our evidence suggests that tree shrews fail to mount, or develop very mild, acute infections in the TG following HSV-1 primary infections in the eye.
MATERIALS AND METHODS
Animals.Adult tree shrews (Tupaia belangeri chinensis), 6-month-old females, were obtained from the animal breeding facility at the Kunming Institute of Zoology (KIZ). They were housed, and no field permit is needed. BALB/c mice were used as experimental controls and were kept in the specific-pathogen-free (SPF) facility at the KIZ.
Ethics statement.All tree shrew and mouse experiments were per-formed according to the procedures and policies of theGuide for the Care and Use of Laboratory Animals(29). The study protocol of ocular scarifi-cation and ocular HSV-1 infection was reviewed and approved by the Institutional Animal Care and Use Committee of the Kunming Institute of Zoology of the Chinese Academy of Sciences. The identification num-ber is SYDW20121201001.
Handling of animals.Animals were anesthetized with an intramuscu-lar injection with ketamine (100 mg/kg of body weight) prior to corneal scarification with the edge of a 24-gauge syringe needle (10 vertical strokes followed by 10 horizontal strokes) and dropping 106PFU of HSV-1 17⫹
virus inoculum on each eye. Mock-infected animals were subjected to corneal scarification, and a phosphate-buffered saline (PBS) solution at a volume equal to that of the virus inoculum solution was added to each eye of mock-infected animals. When the McKrae strain of HSV-1 was used, no ocular scarification was performed. PBS solution was directly applied to the eyes in mock-infected animals. Animals were returned to cages following recovery from the anesthesia. They were monitored every 12 h during the first week and daily during the second week following infection for signs of disease. The eyes of the animals were examined for herpes keratitis daily after the first week. Tears were sampled every other day to test for HSV shedding.
Viruses.The pathogenic HSV-1 strain 17⫹and HSV-1 strain McKrae were used in our experiments. All viruses were grown on Vero cells, and the titers of the viruses on RS1 cells were determined.
Cell lines and antibodies.All cells were obtained from Conservation Genetics Chinese Academy of Science (CAS) Kunming Cell Bank and grown in T75 flasks (catalog no. 708001; NEST Biotechnology) with Dul-becco’s modified Eagle medium (catalog no. 12800-017; Gibco) and 5% (vol/vol) fetal bovine serum (catalog no. 1101-500; Pufei). These cells were used to prepare the viruses and determine virus titer. Rabbit skin cells (RS1), cultured in the same conditions, were used for TG explant cocultivation experiments. ICP0 monoclonal antibody (MAb) (anti-ICP0 MAb 11060 [its epitope recognizes the essential residues between amino acids 20 and 105]) was originally obtained from Roger Everett’s labora-tory (30), and ICP4 monoclonal antibody was obtained from Gerd Maul’s laboratory at the Wistar Institute. HSV-1 polyclonal antibodies were from American Qualex Antibodies (catalog no. RR1190P).
DNA extraction from cells and tissue.Infected cells were lysed by 0.25% Triton X-100, 10 mM EDTA, and 10 mM Tris hydrochloride (pH 7.9) and were incubated at room temperature (RT) for 10 min with gentle mixing. NaCl was then added to a final concentration of 0.2 M, and the mixture was centrifuged at 100⫻gat 4°C for 10 min. The supernatant was incubated with 100g of proteinase K per ml and 0.2% sodium dodecyl sulfate at 37°C for 2 h, and DNA was extracted with phenol, phenol-chloroform, and phenol-chloroform, followed by ethanol precipitation. The amount of DNA was measured byA260. DNA was extracted from tissue by
homogenization prior to the procedures described above.
PCR.Normal PCR was done in an S1000 thermal cycler manufactured by Bio-Rad using recombinant Taq (rTaq) polymerase (catalog no. R001Q; TaKaRa). Quantitative PCR and quantitative reverse transcrip-tion-PCR (qRT-PCR) were done in a 7900HT instrument from Applied Biosystems using a FastStart Universal SYBR green master (ROX) kit (Roche).
Detection of latent HSV-1 genomes in infected tree shrews.At 4 weeks postinfection (p.i.), tree shrews were euthanized, and TGs were dissected for PCR and explant cocultivation. For PCR detection of the viral genome, TG total DNA was extracted using TIANamp genomic DNA kit (catalog no. DP304-02), followed by PCR detection of HSV-1-specific Received6 September 2015Accepted19 October 2015
Accepted manuscript posted online28 October 2015
CitationLi L, Li Z, Wang E, Yang R, Xiao Y, Han H, Lang F, Li X, Xia Y, Gao F, Li Q, Fraser NW, Zhou J. 2016. Herpes simplex virus 1 infection of tree shrews differs from that of mice in the severity of acute infection and viral transcription in the peripheral nervous system. J Virol 90:790 – 804.doi:10.1128/JVI.02258-15.
Editor:K. Frueh
Address correspondence to Jumin Zhou, [email protected].
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
on November 7, 2019 by guest
http://jvi.asm.org/
sequences using primers (see the table inFig. 1F). The PCR product was sequenced, and a Basic Alignment Search Tool (BLAST) search was per-formed to verify that the sequence was HSV.
Quantitative PCR detection of HSV DNA.Mice and tree shrews were infected as described above, and DNA was extracted from TGs by using Genomic DNA purification kit (catalog no. DP304-03; Tiangen). Real-time PCR was run in triplicate with 40 ng mouse TG genomic DNA or 50 ng tree shrew genomic DNA using FastStart Universal SYBR green master kit (catalog no. 04913914001; Roche) and ABI7900HT. Viral DNA was quantified by using HSV-1 UL30 gene-specific primers 5=-TGTTTCGCG TGTGGGACATA and 5=-TTGTCCTTCAGGACGGCTTC. Host-actin was used as a cellular control. Mouse-actin was quantified by using primers 5=-GGCTGTATTCCCCTCCATCG and 5=-CCAGTTGGTAACA ATGCCATGT. Tree shrew-actin was quantified by using primers 5=-C
CAGGGCTTACCTGTACACTG and 5=-CATTTTTAGGTGTGCACTTT
TATTG. Viral DNA levels at each time point were quantified relative to the sample at 13 days postinfection (dpi) by the⌬CTmethod. To deter-mine the relative DNA at various times, the averageCTvalues for-actin were subtracted from the average threshold cycle (CT) values for UL30 genes. The-actinCTvalue was subtracted from the calibrator value (HSV sample at 13 dpi). To obtain the⌬⌬CTvalue, theCTvalue was subtracted by theCTvalue of the input time point as follows:⌬⌬CT⫽ (CTtest⫺CTreference)⫺(CT13 dpi⫺CT13 dpi-actin) where test, reference, 13
dpi, and 13 dpi-actin refer to the samples (e.g.,CTtestis theCTof the test sample).The fold enrichment value is 2⫺⌬⌬
CT .
Reactivation test.Explant cocultivation was performed as previously described (31). At 4 weeks p.i., the animals were euthanized, TGs were removed from the animals, cut into smaller sections using a surgical knife, and placed onto monolayers of rabbit skin cells (RS1). The monolayers were inspected daily for signs of cytopathic effect (CPE) until reactivation occurred. If after 10 days, no CPE was detected, the TG was scored nega-tive for reactivation. The samples that showed no CPE were incubated for up to a month to ensure that there was indeed no CPE developing. If virus CPE was noted, the cell medium was removed and saved for DNA extrac-tion and testing (PCR or sequencing) for HSV-1. To distinguish persistent infections from latent infections, portions of the TGs were also homoge-nized, and the supernatant was incubated with RS1 cells to detect infec-tious virus. To assay for the presence of infecinfec-tious virus in tree shrew or mouse TGs during the acute stage of infection, portions of the TGs were homogenized, and the supernatant was incubated with RS1 cells to detect infectious virus. Viral titers were determined by the method of Harland et al. (32).
In situhybridization.RNA probes corresponding to the latency-as-sociated transcript (LAT) region probe position of the HSV-1 LAT intron (nucleotide [nt] 120384 to 121418) were labeled with digoxigenin-11-UPT (DigU) (DIG RNA labeling mix [catalog no. 11 277 073 910; Roche]). Dissected tree shrew TGs were embedded in paraffin and cut into sections 5m thick on a microtome (Leica RM2245). The sections were deparaffinized in xylene and a gradient concentration of ethanol. RNAin situ hybridization was performed as described on the Goodrich lab website (http://goodrich.med.harvard.edu/uploads/3/7/7/1/37718659/in _situ.pdf).
Briefly, slides were fixed in 4% paraformaldehyde (PFA) at room tem-perature before protease K treatment and a second fixation in 4% PFA. After fixation, slides were acetylated in a solution of 0.25% acetic anhy-dride (catalog no. 320102; Sigma) and 0.1 M triethanolamine (catalog no. 90279; Sigma). The slides were then treated in prehybridization solution for 2 h at 60°C and subsequently hybridized in hybridization buffer with LAT probe overnight at 60°C (prehybridization solution and hybridiza-tion buffer from the RNAin situhybridization protocol on the Goodrich lab website). After the slides were washed in a series of SSC buffers of different concentrations, the antidigoxigenin antibody conjugated to al-kaline phosphatase (Anti-Digoxigenin-AP, Fab fragments; catalog no. 11 093 274 910; Roche) is incubated in MABT buffer (see the RNAin situ hybridization protocol on the Goodrich lab website) overnight at 4°C.
The slides were then developed in alkaline phosphatase buffer with 5-bro-mo-4-chloro-3-indolylphosphate (BCIP)/nitroblue tetrazolium (NBT) and hematoxylin & eosin counterstaining. Finally, the sections were in-spected using a Nikon 80i fluorescence microscope for the presence of LAT.
Detection of spontaneous shedding of HSV-1.Tree shrews were se-dated by injecting them with ketamine. Thirty microliters of precooled PBS per eye was applied to one corner of the eye, followed by gentle rubbing of the eyelids (about 10 times each eye). The PBS was collected from the opposite corner of the eye by using a pipette, transferred to a 1.5-ml Eppendorf tube, and stored at⫺20°C for, at most, a week. Samples were also analyzed by cell plaque assay on RS1 cells to detect HSV-1.
Immunohistochemistry detection of HSV-1 proteins.Tree shrew TGs were embedded in paraffin and sectioned 5m thick on a microtome (Leica RM2245 microtome). The slides were treated at 60°C for 30 to 60 min and then deparaffinized in xylene and a gradient concentration of ethanol. Deparaffinized slides were rinsed with water, and antigen was retrieved in boiling citric acid buffer (pH 6.0) for 24 min (boiled in a microwave). After the slides were allowed to cool naturally, they were washed sufficiently with water, immersed in 3% H2O2for 10 min, and
permeabilized with 0.5% Triton X-100 at RT for 10 min. The slides were blocked in 5% bovine serum albumin (BSA) at RT for 30 min and incu-bated overnight with diluted primary antibody (rabbit anti-HSV1 [cata-log no. RR1190P; American Qualex Antibodies]) at 4°C, and then diluted secondary antibody was added (horseradish peroxidase [HRP]-labeled anti-rabbit IgG antibody [catalog no. 7074S; Cell Signaling Technology]) at room temperature for 60 min. After development with diaminobenzi-dine (DAB) (catalog no. DAB-0031; Fuzhou Maixin, China) for 3 min, the slides were washed in water and counterstained with hematoxylin. The slides were dehydrated in a gradient concentration of ethanol and dehy-drated with xylene for 3 to 5 min each. Finally, the slides were mounted with neutral balsam, and signal was detected with a Nikon 80i fluores-cence microscope.
Immunofluorescence detection of ICP4 proteins.Tree shrew TGs were embedded in paraffin and sectioned 5m thick on a microtome (Leica RM2245). The slides were treated at 60°C for 30 to 60 min and then deparaffinized in xylene and a gradient concentration of ethanol. The deparaffinized slides were rinsed with water, and antigen was retrieved in boiling citric acid buffer (pH 6.0) for 24 min (boiled in a microwave). After the slides were allowed to cool naturally, they were washed suffi-ciently with water and permeabilized with 0.5% Triton X-100 at RT for 10 min. The slides were blocked in 5% BSA at RT for 30 min and then incubated with diluted primary antibody (monoclonal antibody against ICP4 obtained from Gerd Maul’s laboratory) at 4°C overnight, and then diluted secondary antibody (Alexa Fluor 594-labeled goat anti-mouse IgG [H⫹L] from Life Technologies) was added at RT for 60 min (added in the dark, because of light sensitivity). To visualize the cell nucleus, all samples were double stained with Hoechst. Finally, the slides were mounted with mounting medium and examined for signal with a Nikon 80i microscope.
RESULTS
To establish latently infected tree shrews, an ocular route of infec-tion was utilized. To inoculate tree shrews with HSV strain 17⫹, the animal’s cornea was first scarified. This method of infection is commonly used in mouse and rabbit models. Tree shrews infected with either strain of the virus (17⫹or McKrae) showed ruffling of fur, weight loss, and lack of activity. About 10% of the tree shrews showed severe nervous system disease symptoms similar to hu-man encephalitis, such as ataxia, astasia, torticollis, looking up at the sky (or star gazing), and other abnormal behaviors. These symptoms suggest that HSV-1 infected the central nervous system (CNS) of the tree shrew during the acute stage of infection. Most of the animals displaying these symptoms perished within the first 2 weeks of the experiment. Unlike strain 17⫹, which requires
on November 7, 2019 by guest
http://jvi.asm.org/
FIG 1PCR for HSV-1 DNA in tree shrew trigeminal ganglia 4 weeks after eye infection. For PCR detection of the viral genome, TG total DNA was extracted as described in Materials and Methods, followed by PCR detection of HSV-1-specific sequences using primers (see table in panel F). (A) PCR results from primer pair 1. Lanes 1 and 2, TGs from mock-infected tree shrews; lanes 3 through 10, infected tree shrew (TS) TGs; lane 11, HSV-infected Vero cells at 48 h postinfection (hpi) in culture; lane 12, double-distilled (DD) water as a PCR template control. Primer pair 1 gives a PCR product of 185 bp. (B and C) PCR products from the same set of templates as panel A but using primer sets 2 and 3, respectively. The PCR product was sequenced, and a Basic Alignment Search Tool (BLAST) search was performed to verify that the sequence was HSV. (D) Mouse (BALB/c) control. Lanes 1 and 2, mock-infected mouse TGs subjected to PCR using primer pair 1; lanes 3 to 6, PCR on TGs from four different mice infected with HSV-1 17⫹strain; lane 7, PCR of Vero cells infected with HSV-1 17⫹at 48 hpi; lane 8, PCR using DD water as a control. (E) Similar to panel D, except that primer pair 3 was used. (F) Table of the three pairs of primers used in the experiments. The orientation of the primer is indicated at the end of the primer name as follows: F, forward; R, reverse.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.99.483.64.587.2]ocular scarification, the McKrae strain can be applied directly to initiate the symptoms of keratitis. As a result, all McKrae infec-tions were done without ocular scarification in this study.
HSV-1 DNA can be detected by PCR in infected tree shrew TGs 4 weeks p.i.At 4 weeks postinfection (p.i.), tree shrews were euthanized, and TGs were removed for PCR. To determine whether the infected tree shrews become latently infected, we dis-sected the TGs from eight tree shrews infected with HSV-1 17⫹ and from four BALB/c mice as a positive control. DNAs from these TGs were extracted and subjected to PCR using three pairs of primers designed against the UL36 gene of the HSV-1 genome. All three pairs gave consistent PCR products from the HSV-1-in-fected Vero cells; however, primer pair 1 provided the most robust signal (Fig. 1). Using the first pair of primers, all eight tree shrew TGs yield a PCR fragment at the predicted size of 185 bp (Fig. 1A, lanes 3 to 10). The second and third pairs of primers also produced PCR fragments at the predicted 334 and 345 bp, respectively (Fig. 1BandC, lanes 3 to 10). In the mouse positive-control experi-ment, using just primer pairs 1 and 3, all four latently infected mice yielded PCR products at the predicted sizes (Fig. 1DandE, lanes 3 to 6). In mock-infected tree shrews or mice, no PCR
prod-ucts were seen (lanes 1 and 2 fromFig. 1AtoE). The PCR product from infected tree shrew TGs were recovered and sequenced, fol-lowed by analysis using Basic Local Alignment Search Tool (BLAST), which verified that the sequence was HSV and that the results matched the expected regions of the HSV-1 genome.
Stress-induced reactivation by explant cocultivation.To de-termine whether the detected HSV-1 DNA in 4-week-old tree shrew TGs was from a latent infection, an explant cocultivation test was performed as described in Materials and Methods. Briefly, at 4 weeks p.i., animals were euthanized, and TGs were removed and incubated with monolayers of rabbit skin cells (RS1). The monolayers were inspected daily for signs of CPE until reactiva-tion occurred. If no CPE was detected after 10 days, the TG was scored negative for reactivation. If virus-induced CPE was noted, the cell medium was removed, saved for DNA extraction, and tested for HSV-1 (PCR and sequencing).
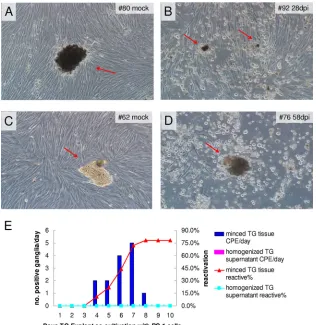
Figure 2Ashows a mock-infected tree shrew (animal 80) TG tissue (arrow) incubated with RS1 cells, which after 10 days of incubation exhibited no signs of CPE, while inFig. 2B, TG from a tree shrew 4 weeks p.i. (animal 92) showed clear CPE, where cells are mostly rounded up near the TG tissues (arrows). Similarly, a FIG 2Explant cocultivation of tree shrew TG on RS1 cells. At 4 weeks or 60 days postinfection (p.i.), the animals (animal 80, 92, 62, and 76) were euthanized, TGs were removed from the animals and incubated with monolayers of rabbit skin cells (RS1). For a control, we also homogenized part of each TG and collected the supernatant after spinning down the tissue debris and incubated the supernatant with a monolayer of RS1 cells. The monolayers were inspected daily for signs of CPE until reactivation occurred. (A) Mock-infected tree shrew TG showed no sign of CPE during the entire time of incubation. (B) HSV-1 17⫹-infected tree shrew TG displayed CPE around the tree shrew tissue. (C) Mock-infected tree shrew TG underwent explant cocultivation, and no CPE was observed. (D) TGs from infected tree shrew at 60 days p.i. were cultured with RS1 cells. CPE is seen near TG tissue debris. (E) Statistical analysis of reactivation from latently infected tree shrews. There was 78% total reactivation. dpi, days postinfection.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.135.451.66.391.2]latently infected tree shrew 8 weeks p.i. also yielded CPE-produc-ing TG (Fig. 2CandD). We summarized the number of reactivat-ing tree shrews as a function of the number of days of cocultivation before they produce CPE inFig. 2E. As can be seen here, the ma-jority of latent TGs become activated around day 6 or day 7, and the total amount of TGs showing CPE was 78%, which is similar to that seen in mouse experiments.
To distinguish between persistent infections and latent infec-tions, part of each TG was homogenized, and the supernatant was incubated with RS1 cells for 10 days. No CPE was seen (Fig. 2E), indicating that infectious virus was not present or was below the level of detection by this method. We therefore tested the sensitiv-ity of our assay by plating different dilutions of HSV-1 17⫹on monolayers of RS1 cells in the presence of TG tissue (cut into several pieces) from mock-infected tree shrews. We could detect virus from an input viral titer of 10 PFU (Table 1).
In situhybridization detected LAT intron signal during la-tency.The LAT is the only transcript consistently expressed dur-ing HSV latency. Its stable intron is a hallmark of latency. To detect the LAT intron in the infected tree shrew TG, RNA probes corresponding to the HSV-1 LAT intron region (probe positions, 120384 to 121418 nt) were labeled with DigU. Dissected tree shrew TGs were embedded in paraffin, sectioned with a micro-tome, and then deparaffinized in xylene and a gradient concentra-tion of ethanol before being subjected to RNAin situ hybridiza-tion as described in Materials and Methods. Although no positive signal could be seen in mock-infected tree shrew TG sections (Fig. 3A), LAT signal could be detected as early as 7 days p.i. (see the arrow inFig. 3B) in TG from infected animals. This signal ap-peared at lower levels, and fewer large neuronal cells, at 7 days p.i. than at later times p.i. At 4 weeks p.i., a time when latency is considered to be established, we detected robust LAT signals pres-ent in more neuronal cells than at 7 days p.i. (Fig. 3C, arrows), and at 2 months p.i., strong LAT signals were still present (Fig. 3D, arrows). We noted that these LAT signals are nuclear, consistent with its distribution in human and other animal models of HSV latency and its role as a noncoding RNA. Since the probe was made against the intron region of LAT, the strength of the signal also supports high stability of the intron in tree shrews, as previously reported in mice (33,34).
Spontaneous reactivation detected by virus shedding in tears. To determine whether HSV-1 in latently infected tree shrews can spontaneously reactivate, we collected tears from in-fected animals. Tree shrews were sedated, and 30l of precooled PBS per eye was applied to one corner of the eye, followed by gentle rubbing of the eye lids and collection of the PBS wash from
[image:6.585.41.544.80.160.2]the opposite corner of the eye with a pipette (as described in Ma-terials and Methods). The collected liquids were analyzed by plaque assay to observe CPE.Table 2shows the animal identifiers (IDs) and summarizes the results of the tear HSV shedding plaque assay during the course of the experiment. To minimize stress to the animals, tears were collected every other day. To improve the survival of the animal through the acute stage of infection, tears were collected from only a subgroup of 5 animals during the first and second weeks p.i. The remainder of the experimental group was not tested during this time (a total of six infected animals). All tears collected from 4 mock-infected tree shrews were negative by the plaque assay (Table 2), while 5 of the 11 infected tree shrews tested during the first 2 weeks (acute phase) all showed positive results during the first week, but the number of tree shrews with positive results dropped during the second week. During the third week, no animals were positive. After the third week, some posi-tive tears were detected. However, the number of animals with positive shedding was small (Table 2). Positive tears were detected TABLE 1Sensitivity assay for explant cocultivation experimenta
Sampling time
CPEbwith the following inoculum (PFU):
105 104 103 102 50 10 1
2 dpi CPE⫹⫹⫹⫹ CPE⫹⫹⫹ CPE⫹⫹ CPE⫹ CPE⫹/⫺ No CPE No CPE
3 dpi Cells fell off Cells fell off Cells fell off Cells fell off CPE⫹⫹⫹⫹; 30% cells fell off
CPE⫹⫹⫹ No CPE
4 dpi Cells fell off Cells fell off No CPE
7 dpi No CPE
aHSV-1 17⫹strain was plated onto single layers of RS1 cells in the presence of cut-up tree shrew TG tissue to closely mimic experimental conditions of explant cocultivations. The appearance of CPE is scored depending on the size and number of viral plaques in culture dishes and recorded as⫹⫹⫹⫹,⫹⫹⫹,⫹⫹,⫹,⫹/⫺, or no CPE. After the appearance of CPE, cells usually become detached from plate and fall off.
b
Symbols:⫹⫹⫹⫹, 80% rounded cells;⫹⫹⫹, 60% rounded cells;⫹⫹, 40% rounded cells;⫹, 20% rounded cells;⫹/⫺,⬍10% rounded cells.
FIG 3In situhybridization detection of LAT signal in HSV-1-infected TGs. RNA probes corresponding to the LAT intron region (positions 120384 to 121418) of HSV-1 were labeled with DigU. Dissected tree shrew TGs were embedded in paraffin and cut into sections 5m thick on a microtome (Leica RM2245 microtome). The sections were deparaffinized in xylene and a gradi-ent concgradi-entration of ethanol. RNAin situhybridization was performed as described in Materials and Methods. Each panel shows representative staining from groups of between three to six animals. (A) Mock-infected tree shrew TG showed no LAT signal. (B) The LAT signal was first detected by day 7 p.i. (yellow arrow). (C) By day 28, the LATin situsignal was readily detected. (D) By 58 days postinfection (dpi), LAT signal persisted and remained strong.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.299.544.447.621.2]in four animals, one of which shed on 2 separate days. Both eyes from a single animal never shed.
Viral plaques were recovered, and PCR was performed using HSV-1 LAT intron-specific primers (forward primer 5=-CGGCG GTCTCTAAGAAGC-3=[positions 6326 to 6343 and 120031 to 120048] and reverse primer 5=-CAGCCAGAGAATCACAGGA G-3=[positions 6234 to 6253 and 120121 to 120140]). The recov-ered PCR fragments were confirmed by sequencing to be HSV-1 DNA. From this experiment, HSV-1-positive tears were detected in 100% of the animals tested during the acute stage of infection, while during latency, 22% of the animals tested positive for HSV-1 at some time between 3 weeks and 2 months p.i. This result sug-gests that latent HSV-1 in tree shrew TG can spontaneously reac-tivate at a low frequency.
HSV-1 antigens in tree shrew TGs are below the level of de-tection during the acute stage of infection.HSV-1 infection in BALB/c mice results in lytic infection in the TGs, which includes expression of lytic genes, viral replication, production of HSV-1 virions, and frequent destruction of infected neurons (6). To char-acterize the tree shrew infection, we performed
immunohisto-chemistry to detect HSV-1 antigens during the course of infection. Surprisingly, we failed to detect any HSV antigen signal in tree shrew TGs infected with HSV-1 17⫹. As shown inFig. 4A(mock infected),Fig. 4B(3 dpi),Fig. 4C(5 dpi),Fig. 4D(7 dpi),Fig. 4E
(27 dpi), andFig. 4F(53 dpi), all showed negative signals, while control experiments in mouse TGs were positive for HSV-1 anti-gen at 5 dpi (Fig. 4GandH). In contrast to the TG results, HSV-1 antigen-positive signals were detected in infected tree shrew CNS around 7 dpi (Fig. 5), indicating that these tree shrews were in-fected. Taken together, these results suggest that acute infection was either absent or below the point of detection by immunohis-tochemistry in the tree shrew sensory neurons.
[image:7.585.42.541.78.429.2]Key HSV-1 genes showed a differential expression profile from mice during the course of HSV-1 infection.To further in-vestigate the characteristics in HSV-1 acute infection in tree shrew TGs, we monitored the expression of ICP0, ICP4, LAT intron, and UL36 (encoding large tegument protein 1-2 [35], which is trun-cated in strain McKrae [36]) in tree shrew TGs and CNS tissues by qRT-PCR. First, we used gene-specific primers for these genes to detect the RNA levels by qRT-PCR in BALB/c mice infected with TABLE 2Detection of spontaneous shedding of HSV-1 in tearsa
aTree shrews were sedated, and tears were collected as described in Materials and Methods. Tears from each animal were incubated with RS-1 cells to observe plaque. The animal
identification numbers for animals 1 to 59 are shown across the top of the table. To the left of the table, the leftmost column shows whether the animal was mock infected or infected with HSV-1 17⫹strain. The next column shows the date postinfection when tears were collected (days postinfection) and whether the tears were collected from the right (R) or left (L) eye. During the acute stage of infection, tears were collected every other day, as the procedure inflicted some stress, which may complicate the course of disease progression.
on November 7, 2019 by guest
http://jvi.asm.org/
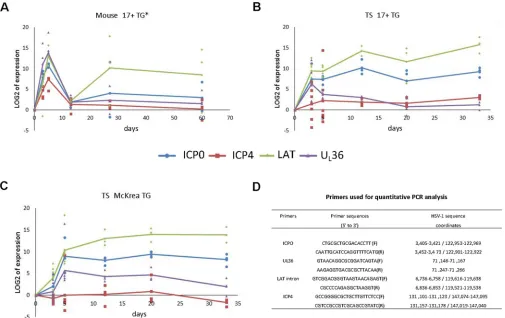
HSV-1 17⫹strain. As can be seen inFig. 6A, all four genes express transcript at high levels at 3 dpi and peak at 5 dpi. At 2 weeks, or day 13, the levels of these gene transcripts dropped to a low basal level. However, at 4 weeks and again at 2 months, the level of LAT increased and remained high, while the expression of other gene transcripts remained low. This pattern is consistent with pub-lished results and suggests that these mice were going through acute infection, which resolved by the end of the second week (37). After 2 weeks, LAT remained expressed, a sign typical of latency.
When tree shrews were similarly infected, we detected a quite different expression pattern. First, the typical high expression of ICP4 transcript, a key virus-specific transcription activator, was absent during the entire course of infection (Fig. 6B). The remain-ing three tested gene transcripts were expressed, with LAT ex-pressed at a level comparable to that in mice (compare the LAT levels inFig. 6AandB) and ICP0 transcript was lagging at 3 dpi but reached a level similar to that in mice by 5 dpi (see ICP0 inFig. 6A
andB). UL36 transcript, on the other hand, was expressed at lower levels than that in mice throughout the acute phase of infection
(compare UL36 inFig. 6AandB). These results suggest that tree shrew TGs developed a much weaker acute infection.
Another key difference from mice is that by 2 weeks p.i., the expression of ICP0 transcript and LAT in tree shrew TG both remained high, with LAT and ICP0 transcript at levels similar to those in mice during acute infection (Fig. 6AandB). The trend of high levels of both LAT transcripts in tree shrew TGs continued at 20 and 33 days p.i., suggesting that, at this stage, HSV-1 in tree shrew TG exhibited latent infection. The robust levels of ICP0 transcript, however, is in contrast to HSV-1 latency seen in mice and rabbits, where ICP0 transcript is very low (38). Considering that we did not detect any HSV-1 antigens in tree shrew TGs (Fig. 4BtoF), we speculate that the high level of ICP0 transcript was not translated and/or degraded quickly in the cytoplasm. We subse-quently conducted immunofluorescence staining with ICP0 monoclonal antibodies, but did not detect any ICP0 protein in acutely infected TGs either in mice or in tree shrews (data not shown). This result is supported by the earlier study of Chen et al., in which they observed ICP0 transcripts but no ICP0 protein in primary neuron cultures (39) and the studies of Mailett et al., who studied ICP0 expression in an oro-ocular model of HSV infection and noted ICP0 transcripts in the nuclei of some neuronal cells during latency (38). The level of ICP0 transcript will be further investigated later in this study.
We also examined the McKrae strain of HSV-1 in the tree shrew TG. As shown inFig. 6C, the expression profile of all four viral genes is similar to that of the 17⫹strain shown inFig. 6B, with the ICP0 transcript and UL36 transcript expressed at slightly higher levels in McKrae-infected TGs.
To confirm the lack of ICP4 transcript expression shown inFig. 6, we performed immunofluorescence staining of tree shrew TGs using an ICP4 monoclonal antibody to detect ICP4 protein. Fig-ure 7AtoDshow mouse TGs during the acute phase of strain 17 HSV infection (positive control). Here we could see that in mock-infected mice or at 3 days p.i., there was no ICP4 signal (arrows point to sensory neuron nuclei inFig. 7AandB). By day 5 p.i., however, clear ICP4 staining could be seen in the nuclei of sensory neurons (white arrows inFig. 7CandD). Occasionally, nonneu-ronal cells near the infected neuron also showed ICP4 staining (see yellow arrows inFig. 7D). In infected tree shrew TGs, however, no positive ICP4 protein staining was seen in mock-infected tree shrews or at 3, 5, or 7 day p.i. (Fig. 7EtoH). This result is consis-tent with the results of the qRT-PCR experiment described above and suggests a diminished acute infection in infected tree shrews compared to infected mice.
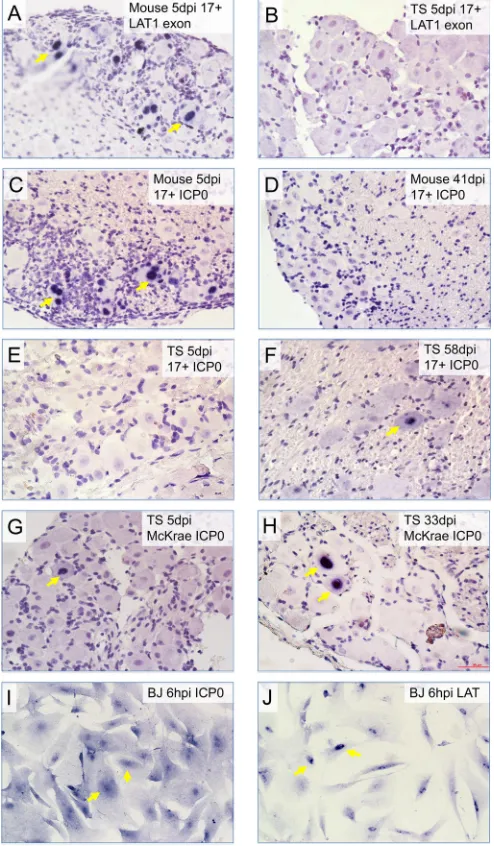
The LAT contains a stable 2.0-kb intron that accumulates at high levels during latency, while the remainder of the LAT (the exon material) is detected only during acute infection (7–9). We examined the expression of LAT exon material, using an exon probe (exon 1, position 124,277 to 124927 nt) andin situ hybrid-ization to distinguish it from the stable intron level during the acute stage of infection. In control experiments, HSV-1-infected mouse TGs showed robust LAT exon expression at 5 days p.i.; however, tree shrew TGs showed no detected LAT exon expres-sion (compareFig. 8AandB). These data are consistent with the lack of, or a low, not detected level of productive infection in tree shrew TGs following eye inoculation of HSV-1.
Because the LAT gene and the ICP0 gene are overlapping and transcribed on opposite strands, qRT-PCR detection is unable to distinguish between these two in the overlapping regions. There-FIG 4Immunohistochemistry using antibody against HSV-1 protein showed
negative signals in tree shrew TGs during the acute phase of infection. (A) Mock-infected tree shrew (TS) TG subjected to the immunohistochemistry (IHC) procedure was negative for HSV-1 proteins. (B to D) HSV-1 17⫹ -infected TS TGs display no HSV-1 antigen signals at 3, 5, and 7 days postin-fection (dpi), respectively. (E and F) During the latent inpostin-fection period, 4 and 8 weeks p.i. (27 and 53 days postinfection, respectively), tree shrew TG IHC was negative for HSV antigens. (G) Six BALB/c mice infected with the 17⫹ strain of HSV-1 showed no detected amount of HSV-1 antigen at 3 dpi. (H) Six BALB/c mice infected with HSV-1 17⫹strain showed significant amounts of HSV-1 antigen at 5 dpi. The black arrows indicate large sensory neurons, the yellow arrows indicate smaller nonneuronal cells, and the red arrows show that the majority of HSV-1 antigen is located in the cytoplasm.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.62.268.65.370.2]fore, we performedin situhybridization to detect ICP0 gene ex-pression during the acute stage and latency. The qRT-PCR result inFig. 6shows that in mouse TGs, ICP0 was expressed at high levels during the acute stage but at very low levels during latency. In contrast, tree shrew TGs showed a moderate ICP0 during the acute phase, but the robust level of ICP0 persists during latency. To confirm whether this was indeed due to ICP0 transcription and not a stable region of LAT overlapping with ICP0, we performed in situ hybridization using an ICP0 transcript-specific probe (probe positions, 122,624 to 123,335 nt) in mouse and tree shrew TGs infected with HSV-1. The result is shown as representative stainings inFig. 8CtoH. Here ICP0 transcript is readily seen in acutely infected mouse TGs (Fig. 8C) but is hardly detected in latent mouse TGs (Fig. 8D). In contrast, TGs of tree shrews in-fected with the 17⫹strain showed no ICP0 transcript during the acute stage (Fig. 8E) but exhibited readily detected ICP0 signal during latency (Fig. 8F). The McKrae strain-infected tree shrew TGs displayed detected but infrequent ICP0 transcript during the acute stage (Fig. 8G) and robust ICP0 during latency (Fig. 8H). Although we could hardly detect ICP0 transcript in mouse latently infected TGs, ICP0 transcript has been detected during latency in mice by others (38); nonetheless, bothin situhybridization results discussed here and qRT-PCR results shown inFig. 6strongly sug-gest that latently infected tree shrew TGs express much higher levels of ICP0 transcript than mouse TGs during HSV-1 latency. As ICP0 transcript is probably not translated into ICP0 protein (discussed earlier and inFig. 4; also data not shown), we examined the distribution of ICP0 RNA in TG neurons and noticed that ICP0 RNA is exclusively nuclear (see arrows inFig. 8FtoH). In contrast, in HSV-1-infected HeLa cells (which express ICP0 pro-tein), ICP0 transcript showed both nuclear and cytoplasmic ICP0 in situsignals (Fig. 8I; also compareFig. 8Iwith the nuclear
local-ized, untranslated LAT inFig. 8J). The exclusive nuclear localiza-tion of the ICP0 transcript supports the observalocaliza-tion that ICP0 is not translated in neuronal cells in mouse and tree shrew TGs. The role of nuclear ICP0 transcript expression during latency is not yet known and is being further investigated.
Active HSV-1 virus was not detected in tree shrew TGs dur-ing the acute stage of infection.As the immunohistochemistry with HSV-1 antibodies, immunofluorescence staining with ICP4 antibody, and qRT-PCR results suggested that there is a weak, or an absence of detected, productive infection of HSV-1 in tree shrew TGs during the acute stage of infection. To directly test that a productive infection is absent or not detected, we dissected and homogenized the TGs from mock-infected animals or infected animals at 3, 5, and 7 days postinfection and then cocultured the supernatant with RS1 cells to detect live HSV-1 virus. At 20 h after incubation with supernatant from TGs of infected mice, we could readily observe CPE due to HSV-1 lytic infection in RS1 cells (Fig. 9A). However, no CPE was seen in RS1 cells cultured with super-natant from infected tree shrew TGs even after extended incuba-tion of more than 10 days (Fig. 9B). We measured the titer of the viral yield of the mouse and tree shrew TGs from groups of three animals (6TGs) for each time point (mock infected and at day 3, 5, and 7 p.i.), and the results were presented inFig. 9C. The amounts of infectious HSV-1 virus in mouse TGs were similar to previously published data, which peaked at day 5 and became weaker at day 7 (40). The tree shrew TGs, however, produced no infectious virus from all tested animals. The results of this experiment strongly argue that there is an absence, or an undetectable level (smaller than 10 PFU [Table 1]), of lytic infection in tree shrew TGs during the acute phase of HSV-1 infection. This result also contrasts the mouse experiments where productive infection is readily detected by all methods employed during the course of our investigation. FIG 5Immunohistochemistry detection of HSV-1 antigens in infected mouse and tree shrew CNS. The corneas of mice and tree shrews were inoculated with the McKrae strain of HSV-1 without ocular scarification. CNS tissues were dissected, sectioned, and processed for immunohistochemistry with anti-HSV-1 polyclonal antibodies. (A) Mock-infected mouse control. (B) Mouse CNS. The arrows point to clusters of infected neurons positive for HSV-1 antigens. (C) Tree shrew CNS mock-infected control. (D) HSV-1-infected tree shrew CNS. The arrows indicate neurons positive for HSV-1 antigens.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.137.452.67.301.2]The lack of detected productive infection during the acute stage prompted us to determine the viral genome load in infected tree shrew TGs (Fig. 9D). We found that, while mouse TGs con-tained much higher levels of viral genomic DNA during the acute stage, tree shrew TGs generally had lower levels of HSV genome during this time. However, acutely infected tree shrew TGs (from day 3 to day 10 p.i.) still showed higher viral genome content than at day 13 p.i., a time when the acute stage ends and during latency. This result suggests that HSV-1 underwent productive infection in tree shrew TGs, although the live virus was below the threshold of detection.
DISCUSSION
The mechanism of latent herpes infection remains poorly defined due to the need to study peripheral nervous system tissue and the lack of appropriate animal models to closely mimic HSV latency in humans. Mouse and rabbit models have been used to charac-terize the HSV-1 infection, and guinea pigs have been favored for HSV-2 infection, at a pathological level, whilein vitrotissue cul-ture and primary cell culcul-ture models have been used to study molecular pathways of latency in more detail (2,41,42). Although we have learned a great deal from the mouse and rabbit models of human HSV-1 infection, there are many questions that could not be answered with these models. Rodent models are seldom used
for chronic stress paradigms of human disease, whereas tree shrews are (43–45). Furthermore, tree shrews are finding a place in many models of human brain pathogenesis from mapping of neuropeptide distribution (46), models for opiate addiction (47) to drug target mining (48) and specific diseases such as Parkin-son’s disease (49). HSV infection was first reported in the tree shrew in 1978 by Darai et al., who studied the pathogenesis fol-lowing intravenous, intraperitoneal, and subcutaneous infection (27). They later studied infection by intraherpetic transfection of HSV DNA (50) and viral latency in this system (28). An important goal in our study of tree shrews was to characterize tree shrew herpes simplex infection following eye inoculation and discover potential differences between tree shrew and rodent models in the hope of refining our knowledge of human HSV latency and reac-tivation.
We have studied the HSV-1-infected tree shrew trigeminal ganglion. Using PCR, qRT-PCR,in situ hybridization, explant cocultivation, immunohistochemistry, and immunofluorescence, we have confirmed that the tree shrew does indeed establish latent infection in trigeminal ganglia with both 17⫹and McKrae strains of HSV-1 following eye infection. The kinetics and efficiency of formation of latency and reactivation following explant cocultiva-tion are similar to those of mice. Unlike the mouse model (but like rabbit and human infection), tree shrews appear to spontaneously FIG 6qRT-PCR detection of HSV-1 transcripts in infected animal TGs shows different expression profiles in mice and tree shrews. Mice and tree shrews infected with 17⫹or McKrae strain of HSV-1 were euthanized and TGs were removed from the animals. RNAs from these samples were extracted for qRT-PCR analyses. (A) Six BALB/c mice were infected with the 17⫹strain of HSV-1, and TGs were isolated at various days postinfection. qRT-PCR results for ICP0, ICP4, LAT, and UL36 were plotted. (B) Similar to panel A, except TGs from tree shrews infected with HSV-1 17⫹strain were analyzed. (C) Similar to panel B except that the McKrae strain was used. (D) Primers used in the qRT-PCR detection of viral transcripts.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:10.585.41.546.66.384.2]shed virus from their eyes during latency. The frequency of spon-taneous reactivation is less than that reported for the rabbit, and this may be due to the less invasive technique of washing solution across the eye, rather than the more invasive swabbing technique commonly used on rabbit eyes. However, at the molecular level, tree shrew latency exhibits a different expression profile for key regulatory viral genes from the mouse model during both acute and latent stages of HSV-1 infection.
When monitoring acute infection in the tree shrew TGs follow-ing inoculation of 17⫹or McKrae strain of HSV-1, we failed to detect HSV-1 antigen in the tree shrew at 3, 5, or 7 days p.i., although in the control experiment, HSV-1 antigen was readily detected in tree shrew CNS, and in both the TGs and CNSs of infected mice (Fig. 4and5). This observation suggests that acute infection in tree shrew TG was either very weak or simply absent.
When we measured ICP0 and ICP4 transcript expression in the tree shrew TG during the acute phase of infection by qRT-PCR, we found that ICP0 is expressed at a level slightly lower than that in the mouse TG. However, ICP4 expression is nearly undetected (Fig. 6BandC). Since ICP4 is necessary for most viral early and late gene expression, the lack of ICP4 explains the absence of HSV-1 antigens in the TGs of infected tree shrews. This result strongly contrasts with that of the infected mouse TG where ICP4 FIG 7Immunofluorescence staining of ICP4 in mouse and tree shrew TGs.
BALB/c mice and tree shrews were infected with the 17⫹strain of HSV-1, and at 3 dpi (mouse and tree shrew), 5 dpi (mouse and tree shrew), and 7 dpi (tree shrew), the animals were euthanized and dissected to obtain TGs, which were paraffin embedded and sectioned on a microtome before immunofluores-cence staining with ICP4 antibody. (A) Mock-infected BALB/c mouse TG. No positive signal was detected. Arrows indicate neuronal nucleus. (B) HSV-1 17⫹-infected mouse at 3 days postinfection showed no ICP4 signal. (C) At 5 dpi, the ICP4 signal was readily detected in large neuronal cells in the TG. The arrows point to large neuronal nuclei, where ICP4 signal could be seen. (D) At 5 dpi, ICP4 stained not only the neuronal nucleus (white arrows) but also smaller nonneuronal cells (yellow arrows). (E) TG section in a mock-infected tree shrew. Arrows point to neuronal cell nuclei. (F) Infected TS TG at 3 dpi. No ICP4 signal was seen. (G) Infected TS TG at 5 dpi. No ICP4 signal was detected. (H) HSV-1 17⫹-infected tree shrew TG section at 7 dpi. No ICP4 signal was detected.
FIG 8In situhybridization detection of ICP0 signals in mouse and tree shrew TGs. (A) During the acute stage of infection (5 dpi using HSV-1 17⫹), LAT transcript is readily detected using RNA probe against the first exon of LAT. (B) Compared to mouse TG, LAT exon is undetected at the same time point in infected tree shrew TGs. (C) ICP0 is expressed in HSV-1 17⫹-infected mouse TG at 5 dpi. (D) The ICP0 transcript is undetected in most HSV-1 17⫹ -infected mouse TGs during latency. (E) In acutely HSV-1 17⫹-infected tree shrew TG, no ICP0 was detected at 5 dpi. (F) In latently HSV-1 17⫹-infected tree shrew TG, ICP0 could be detected at day 58 postinfection. (G) Unlike strain 17⫹, the McKrae strain-infected tree shrew TGs show ICP0 expression at 5 dpi. (H) In latent TGs infected with McKrae strain, ICP0 is readily de-tected. (I)In situhybridization of ICP0 in HSV-1-infected BJ cells. (J)In situ hybridization of LAT exon in HSV-1-infected BJ cells.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:11.585.62.266.62.383.2] [image:11.585.298.545.71.494.2]expression is very high during acute infection (Fig. 6A) (51). The UL36 gene indeed showed a moderate level of expression during the acute stage of infection. These results are consistent with a weaker acute infection in general observed in tree shrews. Indeed,
supernatant obtained from homogenized TGs of acutely infected tree shrews produced no CPE when cocultured with RS1 cells, while the mouse control yielded consistent CPE (Fig. 9AandB). When viral genome load was monitored in infected tree shrew FIG 9HSV-1 virus is not detected from tree shrew TGs during the acute stage of infection. TGs were dissected from mock-infected mice and tree shrews and infected mice and tree shrews. The TGs from infected animals were dissected 3, 5, or 7 days after inoculating the eyes of the animal with the 17⫹strain of HSV-1, and the TGs were homogenized. The supernatant was incubated with RS1 cells to observe CPE and also incubated with RS cells on 96-well plates to calculate viral titer. (A) Mock-infected mouse TG control. A CPE was detected after 3 days. Day 5 showed stronger CPE than day 7 postinfection. (B) No CPE was detected with tree shrew TG supernatants. (C) Titers of HSV-1 loads were measured for infected mouse and tree shrew TGs. Each point represents the geometric mean titer determined from four individual ganglia at the indicated time (days) postinfection. The titers are plotted on a logarithmic scale as the number of PFU per trigeminal ganglia (PFU/TG). (D) Relative viral genome copy number in mouse and tree shrew TGs were detected by real-time PCR. Mice or tree shrews were infected as described above. Viral DNA levels at each time point were quantified relative to the 13 dpi sample by the⌬⌬CTmethod as follows:⌬⌬CT⫽(CTtest⫺ CTreference)⫺(CT13 dpi⫺CT13 dpi-actin) where test, reference, 13 dpi, and 13 dpi-actin refer to the samples (e.g.,CTtestis theCTof the test sample). The fold
enrichment value is 2⫺⌬⌬CT. The values are means⫾standard errors (SE) (error bar) ofⱖ3 samples per data point.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:12.585.40.536.65.564.2]TGs throughout the course of infection, we noted that there was an increase of viral DNA during acute infection, although it was much lower compared to that in infected mouse TGs (Fig. 9C). Taken together, these analyses suggest that HSV-1 infection in tree shrew TGs is much weaker than in mouse TGs.
The absence of a detectable productive infection in tree shrew TG suggests that the infected neurons may survive the acute/reac-tivation stage of infection and remain viable for the lifetime of the animal. This may shed some light on the long-standing puzzling fact that following frequent herpes recurrences in humans, the tissue in affected areas does not lose sensation, suggesting that there is no death of neurons innervating the affected area. It is likely, as the tree shrew experiment suggests that in tree shrews and in primates, acute infection in the TG neurons, or reactivation from latently infected neurons, generally does not lead to high level of HSV-1 lytic gene expression, and thus permits the survival of infected neurons.
Another notable difference between tree shrew and mouse la-tency is that the tree shrew expressed ICP0 transcript at high levels by qRT-PCR (Fig. 6BandC) andin situhybridization using an ICP0 antisense probe (Fig. 8CtoH). ICP0 is an immediate early gene, its protein product is a promiscuous activator of both cell and virus gene expression. Its presence ensures efficient lytic in-fection and reactivation from latency (52). ICP0 expression is also known to trigger apoptosis (53). Although there is much available data on transcript levels for tissue culture infections, there is little available data for animal model infections (54–56). Differing lev-els of HSV-1 transcripts in rabbits and mice have been noted be-fore (57); however, ICP0 represents a special case, i.e., although ICP0 RNA was detected, ICP0 protein was not detected in tree shrew TGs using HSV-1 polyclonal antibodies (Fig. 4) and ICP0 monoclonal antibody (data not shown). The lack of ICP0 protein during acute infection was also seen in mouse TGs (data not shown). The lack of ICP0 protein in TG neurons was supported by the observation that ICP0 mRNA is exclusively nuclear (Fig. 8Fto
J). These results are consistent with the results of Chen et al. (39) and Maillet et al. (38). Chen et al. (39) found that ICP0 protein was absent from cultured neuronal cells; thus, neuronal tissue-specific mechanisms exist to prevent the RNA from being translated or entering the cytoplasm or the RNA is degraded quickly in the cytoplasm. Maillet et al. noted nuclear accumulation of ICP0 in mouse TG neurons during latency (38). Indeed, a recent study suggests that in mouse neuronal cells, miR138 specifically targets ICP0 transcript, prevents its translation, and increases its degra-dation (58). A similar mechanism might be operating in tree shrew TG neurons as well. This neuron-specific, uneven nuclear versus cytoplasmic distribution of viral products is not limited to ICP0 mRNA, the VP16 protein and the host HCF1 factor are pre-vented from entering the nucleus, a potential cause of suppression of HSV-1 lytic infection in neurons (59).
Taken together, HSV-1 infection in tree shrew TGs differs sig-nificantly from that in mouse TGs, especially the lack of ICP4 expression during the acute stage and the high level of the ICP0 transcript (although not translated into ICP0 protein) during la-tency in tree shrew TGs. The biological consequences and mech-anisms leading to the differing expression of HSV-1 genes, espe-cially ICP4 and ICP0 in tree shrew and mouse TGs are unknown at this point and merit additional investigation. Whether tree shrews could become a popular organism with which to study HSV-1 latency will need further characterization, and established tree
shrew strains would have to be available before meaningful im-munological study could be conducted. However, the fact that HSV-1 does indeed establish latency in tree shrew TGs, the lack of detectable productive infection in TG sensory neurons during the acute phase of infection, the interesting differences in gene expres-sion profiles in tree shrews and mice, and the fact that tree shrews are a less expensive model for HSV-1 infections than rabbits and more closely related to primates are compelling enough to warrant a closer examination of this creature.
ACKNOWLEDGMENTS
ICP4 monoclonal antibody was originally obtained from Gerd Maul’s laboratory. ICP0 monoclonal antibody was originally from Roger Ever-ett’s laboratory. We thank Guijun Chen and Liping Yang for their excel-lent technical assistance.
FUNDING INFORMATION
National Science Foundation of China provided funding to Jumin Zhou under grant number NSFC81471966. A Common Project of Panzhihua, Science and Technology Bureau of China provided funding to Hongbo Han under grant number 2012CY-S-22(9). A Visiting Professorship for Senior International Scientists from CAS provided funding to Nigel W. Fraser under grant number 2012T1S0001. Kunming Institute of Zoology, Chinese Academy of Sciences provided funding to Jumin Zhou under grant number Y102421081. Yunnan Provincial Government provided funding to Jumin Zhou under grant numbers 2011HA005 and 2013FA051. Chinese Academy of Sciences (CAS) provided funding to Jumin Zhou under grant number KSCXZ-EW-BR-6.
This work was partially supported by the 863 Project (2012AA021801, 2012AA022402) from the Ministry of Science and Technology of China and by a one-three-five strategic planning project from the CAS.
REFERENCES
1.Steiner I, Benninger F.2013. Update on herpes virus infections of the nervous system. Curr Neurol Neurosci Rep13:414.http://dx.doi.org/10 .1007/s11910-013-0414-8.
2.Webre JM, Hill JM, Nolan NM, Clement C, McFerrin HE, Bhattacha-rjee PS, Hsia V, Neumann DM, Foster TP, Lukiw WJ, Thompson HW. 2012. Rabbit and mouse models of HSV-1 latency, reactivation, and re-current eye diseases. J Biomed Biotechnol2012:612316.http://dx.doi.org /10.1155/2012/612316.
3.Stuart PM, Keadle TL.2012. Recurrent herpetic stromal keratitis in mice: a model for studying human HSK. Clin Dev Immunol2012:728480.http: //dx.doi.org/10.1155/2012/728480.
4.Dasgupta G, BenMohamed L. 2011. Of mice and not humans: how reliable are animal models for evaluation of herpes CD8⫹-T cell-epitopes-based immunotherapeutic vaccine candidates? Vaccine 29:5824 –5836. http://dx.doi.org/10.1016/j.vaccine.2011.06.083.
5.Bernstein DI, Stanberry LR, Harrison CJ, Kappes JC, Myers MG.1986. Antibody response, recurrence patterns and subsequent herpes simplex virus type 2 (HSV-2) reinfection following initial HSV-2 infection of guinea pigs: effects of acyclovir. J Gen Virol67:1601–1612.http://dx.doi .org/10.1099/0022-1317-67-8-1601.
6.Roizman B, Whitley RJ.2013. An inquiry into the molecular basis of HSV latency and reactivation. Annu Rev Microbiol67:355–374.http://dx.doi .org/10.1146/annurev-micro-092412-155654.
7.Stevens JG, Wagner EK, Devi-Rao GB, Cook ML, Feldman LT.1987. RNA complementary to a herpes virus gene mRNA is prominent in la-tently infected neurons. Science 235:1056 –1059. http://dx.doi.org/10 .1126/science.2434993.
8.Spivack JG, Fraser NW.1987. Detection of herpes simplex virus type 1 transcripts during latent infection in mice. J Virol61:3841–3847. 9.Rock DL, Nesburn AB, Ghiasi H, Ong J, Lewis TL, Lokensgard JR,
Wechsler SL.1987. Detection of latency-related viral RNAs in trigeminal ganglia of rabbits latently infected with herpes simplex virus type 1. J Virol 61:3820 –3826.
10. Umbach JL, Kramer MF, Jurak I, Karnowski HW, Coen DM, Cullen
on November 7, 2019 by guest
http://jvi.asm.org/
BR.2008. MicroRNAs expressed by herpes simplex virus 1 during latent infection regulate viral mRNAs. Nature454:780 –783.http://dx.doi.org /10.1038/nature07103.
11. Perng GC, Jones C, Ciacci-Zanella J, Stone M, Henderson G, Yukht A, Slanina SM, Hofman FM, Ghiasi H, Nesburn AB, Wechsler SL.2000. Virus-induced neuronal apoptosis blocked by the herpes simplex virus latency-associated transcript. Science287:1500 –1503. http://dx.doi.org /10.1126/science.287.5457.1500.
12. Mellerick DM, Fraser NW.1987. Physical state of the latent herpes sim-plex virus genome in a mouse model system: evidence suggesting an epi-somal state. Virology158:265–275.http://dx.doi.org/10.1016/0042-6822 (87)90198-X.
13. Rock DL, Fraser NW.1985. Latent herpes simplex virus type 1 DNA contains two copies of the virion DNA joint region. J Virol55:849 – 852. 14. Rock DL, Fraser NW.1983. Detection of HSV-1 genome in the central
nervous system of latently infected mice. Nature302:523–525.http://dx .doi.org/10.1038/302523a0.
15. Bloom DC, Giordani NV, Kwiatkowski DL.2010. Epigenetic regulation of latent HSV-1 gene expression. Biochim Biophys Acta1799:246 –256. http://dx.doi.org/10.1016/j.bbagrm.2009.12.001.
16. Deshmane SL, Fraser NW.1989. During latency, herpes simplex virus type 1 DNA is associated with nucleosomes in a chromatin structure. J Virol63:943–947.
17. Cliffe AR, Coen DM, Knipe DM.2013. Kinetics of facultative hetero-chromatin and polycomb group protein association with the herpes sim-plex viral genome during establishment of latent infection. mBio 4:e00590-12.http://dx.doi.org/10.1128/mBio.00590-12.
18. Mohr I.2004. Neutralizing innate host defenses to control viral transla-tion in HSV-1 infected cells. Int Rev Immunol23:199 –220.http://dx.doi .org/10.1080/08830180490265600.
19. Goodkin ML, Morton ER, Blaho JA.2004. Herpes simplex virus infec-tion and apoptosis. Int Rev Immunol23:141–172.http://dx.doi.org/10 .1080/08830180490265574.
20. Ren H, Wilson G.1996. Apoptosis in the corneal epithelium. Invest Ophthalmol Vis Sci37:1017–1025.
21. He L, Frost MR, Siegwart JT, Jr, Norton TT.2014. Gene expression signatures in tree shrew choroid during lens-induced myopia and recov-ery. Exp Eye Res123:56 –71.http://dx.doi.org/10.1016/j.exer.2014.04.005. 22. He L, Frost MR, Siegwart JT, Jr, Norton TT.2014. Gene expression signatures in tree shrew choroid in response to three myopiagenic condi-tions. Vision Res102:52– 63. http://dx.doi.org/10.1016/j.visres.2014.07 .005.
23. Guo L, Frost MR, He L, Siegwart JT, Jr, Norton TT. 2013. Gene expression signatures in tree shrew sclera in response to three myopiagenic conditions. Invest Ophthalmol Vis Sci54:6806 – 6819.http://dx.doi.org /10.1167/iovs.13-12551.
24. Ohl F, Oitzl MS, Fuchs E.1998. Assessing cognitive functions in tree shrews: visuo-spatial and spatial learning in the home cage. J Neurosci Methods81:35– 40.http://dx.doi.org/10.1016/S0165-0270(98)00011-9. 25. Fan Y, Huang ZY, Cao CC, Chen CS, Chen YX, Fan DD, He J, Hou HL,
Hu L, Hu XT, Jiang XT, Lai R, Lang YS, Liang B, Liao SG, Mu D, Ma YY, Niu YY, Sun XQ, Xia JQ, Xiao J, Xiong ZQ, Xu L, Yang L, Zhang Y, Zhao W, Zhao XD, Zheng YT, Zhou JM, Zhu YB, Zhang GJ, Wang J, Yao YG.2013. Genome of the Chinese tree shrew. Nat Commun4:1426. http://dx.doi.org/10.1038/ncomms2416.
26. Xu L, Fan Y, Jiang XL, Yao YG. 2013. Molecular evidence on the phylogenetic position of tree shrews. Dongwuxue Yanjiu34:70 –76. (In Chinese.)http://dx.doi.org/10.3724/SP.J.1141.2013.02070.
27. Darai G, Schwaier A, Komitowski D, Munk K. 1978. Experimental infection of Tupaia belangeri (tree shrews) with herpes simplex virus types 1 and 2. J Infect Dis137:221–227.http://dx.doi.org/10.1093/infdis/137.3 .221.
28. Rosenwolff A, Scholz J, Darai G.1989. Organotropism of latent herpes simplex virus type 1 is correlated to the presence of a 1.5 kb RNA transcript mapped within the BamHI DNA fragment B (0.738 to 0.809 map units). Virus Res12:43–52.http://dx.doi.org/10.1016/0168-1702(89)90052-X. 29. National Research Council.1996. Guide for the care and use of laboratory
animals. National Academy Press, Washington, DC.
30. Everett RD, Cross A, Orr A.1993. A truncated form of herpes simplex virus type 1 immediate-early protein Vmw110 is expressed in a cell type dependent manner. Virology197:751–756. http://dx.doi.org/10.1006 /viro.1993.1651.
31. Steiner I, Spivack JG, Lirette RP, Brown SM, MacLean AR,
Subak-Sharpe JH, Fraser NW.1989. Herpes simplex virus type 1 latency-associated transcripts are evidently not essential for latent infection. EMBO J8:505–511.
32. Harland J, Brown SM.1998. HSV growth, preparation, and assay. Meth-ods Mol Med10:1– 8.
33. Farrell MJ, Dobson AT, Feldman LT.1991. Herpes simplex virus laten-cy-associated transcript is a stable intron. Proc Natl Acad Sci U S A88: 790 –794.http://dx.doi.org/10.1073/pnas.88.3.790.
34. Thomas DL, Lock M, Zabolotny JM, Mohan BR, Fraser NW.2002. The 2-kilobase intron of the herpes simplex virus type 1 latency-associated transcript has a half-life of approximately 24 hours in SY5Y and COS-1 cells. J Virol76:532–540.http://dx.doi.org/10.1128/JVI.76.2.532-540 .2002.
35. Caduco M, Comin A, Toffoletto M, Munegato D, Sartori E, Celestino M, Salata C, Parolin C, Palu G, Calistri A.2013. Tsg101 interacts with herpes simplex virus 1 VP1/2 and is a substrate of VP1/2 ubiquitin-specific protease domain activity. J Virol87:692– 696.http://dx.doi.org/10.1128 /JVI.01969-12.
36. Watson G, Xu W, Reed A, Babra B, Putman T, Wick E, Wechsler SL, Rohrmann GF, Jin L.2012. Sequence and comparative analysis of the genome of HSV-1 strain McKrae. Virology433:528 –537.http://dx.doi .org/10.1016/j.virol.2012.08.043.
37. Spivack JG, Fraser NW.1988. Expression of herpes simplex virus type 1 latency-associated transcripts in the trigeminal ganglia of mice during acute infection and reactivation of latent infection. J Virol62:1479 –1485. 38. Maillet S, Naas T, Crepin S, Roque-Afonso AM, Lafay F, Efstathiou S, Labetoulle M.2006. Herpes simplex virus type 1 latently infected neurons differentially express latency-associated and ICP0 transcripts. J Virol80: 9310 –9321.http://dx.doi.org/10.1128/JVI.02615-05.
39. Chen X, Li J, Mata M, Goss J, Wolfe D, Glorioso JC, Fink DJ.2000. Herpes simplex virus type 1 ICP0 protein does not accumulate in the nucleus of primary neurons in culture. J Virol74:10132–10141.http://dx .doi.org/10.1128/JVI.74.21.10132-10141.2000.
40. Steiner I, Spivack JG, Deshmane SL, Ace CI, Preston CM, Fraser NW. 1990. A herpes simplex virus type 1 mutant containing a non-transinducing Vmw65 protein establishes latent infection in vivo in the absence of viral replication and reactivates efficiently from explanted tri-geminal ganglia. J Virol64:1630 –1638.
41. Hill JM, Nolan NM, McFerrin HE, Clement C, Foster TP, Halford WP, Kousoulas KG, Lukiw WJ, Thompson HW, Stern EM, Bhattacharjee PS.2012. HSV-1 latent rabbits shed viral DNA into their saliva. Virol J 9:221.http://dx.doi.org/10.1186/1743-422X-9-221.
42. Kim JY, Shiflett LA, Linderman JA, Mohr I, Wilson AC.2014. Using homogeneous primary neuron cultures to study fundamental aspects of HSV-1 latency and reactivation. Methods Mol Biol1144:167–179.http: //dx.doi.org/10.1007/978-1-4939-0428-0_11.
43. Fuchs E, Czeh B, Flugge G. 2004. Examining novel concepts of the pathophysiology of depression in the chronic psychosocial stress para-digm in tree shrews. Behav Pharmacol15:315–325.http://dx.doi.org/10 .1097/00008877-200409000-00003.
44. Wang J, Chai A, Zhou Q, Lv L, Wang L, Yang Y, Xu L.2013. Chronic clomipramine treatment reverses core symptom of depression in subor-dinate tree shrews. PLoS One8:e80980.http://dx.doi.org/10.1371/journal .pone.0080980.
45. Cao J, Yang EB, Su JJ, Li Y, Chow P.2003. The tree shrews: adjuncts and alternatives to primates as models for biomedical research. J Med Primatol 32:123–130.http://dx.doi.org/10.1034/j.1600-0684.2003.00022.x. 46. Ni RJ, Shu YM, Luo PH, Fang H, Wang Y, Yao L, Zhou JN.2014.
Immunohistochemical mapping of neuropeptide Y in the tree shrew brain. J Comp Neurol523:495–529.http://dx.doi.org/10.1002/cne.23696. 47. Shen F, Duan Y, Jin S, Sui N.2014. Varied behavioral responses induced by morphine in the tree shrew: a possible model for human opiate addic-tion. Front Behav Neurosci8:333.http://dx.doi.org/10.3389/fnbeh.2014 .00333.
48. Zhao F, Guo X, Wang Y, Liu J, Lee WH, Zhang Y.2014. Drug target mining and analysis of the Chinese tree shrew for pharmacological testing. PLoS One9:e104191.http://dx.doi.org/10.1371/journal.pone .0104191.
49. Ma KL, Gao JH, Huang ZQ, Zhang Y, Kuang DX, Jiang QF, Han YY, Li C, Wang WG, Huang XY, Xu J, Tong PF, Yin XX, Dai JJ.2013. Motor function in MPTP-treated tree shrews (Tupaia belangeri chinen-sis). Neurochem Res 38:1935–1940. http://dx.doi.org/10.1007/s11064 -013-1099-8.