0022-538X/09/$08.00⫹0 doi:10.1128/JVI.00908-09
Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Site-Specific Phosphorylation Regulates Human T-Cell Leukemia
Virus Type 2 Rex Function In Vivo
䌤
Matthew Kesic,
1,2Michael Ward,
5O. John Semmes,
5and Patrick L. Green
1,2,3,4*
Center for Retrovirus Research,1Departments of Veterinary Biosciences2and Molecular Virology, Immunology, and Medical Genetics,3
and Comprehensive Cancer Center and Solove Research Institute,4The Ohio State University, Columbus, Ohio 43210,
and Department of Microbiology and Molecular Cell Biology and Center for Biomedical Proteomics, Eastern Virginia Medical School, Norfolk, Virginia 235075
Received 6 May 2009/Accepted 12 June 2009
Human T-cell leukemia virus type 2 (HTLV-2) Rex is a transacting regulatory protein required for efficient cytoplasmic expression of the unspliced and incompletely spliced viral mRNA transcripts encoding the struc-tural and enzymatic proteins. Previously, it was demonstrated that phosphorylation of Rex-2, predominantly on serine residues, is correlated with an altered conformation, as observed by a gel mobility shift and the detection of two related protein species (p24Rexand p26Rex). Rex-2 phosphorylation is required for specific binding to its viral-mRNA target sequence and inhibition of mRNA splicing and may be linked to subcellular compartmentalization. Thus, the phosphorylation-induced structural state of Rex in the infected cell may be a switch that determines whether HTLV exists in a latent or productive state. We conducted a phosphoryl and functional mapping of both structural forms of mammalian-cell-expressed Rex 2 using affinity purification, liquid chromatography-tandem mass spectrometry, and site-directed substitutional mutational analysis. We identified two phosphorylation sites in p24Rexat Ser-117 and Thr-164. We also identified six phosphorylation sites in p26Rex at Thr-19, Ser-117, Ser-125, Ser-151, Ser-153, and Thr-164. We evaluated the functional significance of these phosphorylation events and found that phosphorylation on Thr-164, Ser-151, and Ser-153 is critical for Rex-2 function in vivo and that phosphorylation of Ser-151 is correlated with nuclear/nucleolar subcellular localization. Overall, this work is the first to completely map the phosphorylation sites in Rex-2 and provides important insight into the phosphorylation continuum that tightly regulates Rex-2 structure, cellular localization, and function.
Human T-cell leukemia virus types 1 and 2 (HTLV-1 and HTLV-2) are closely related complex retroviruses that have the ability to transform primary human T cells in culture and are associated with leukemia and a variety of human diseases (57). HTLV-1 is causally associated with adult T-cell leukemia, an aggressive CD4⫹T-cell malignancy, and a chronic neuro-degenerative disorder, HTLV-associated myelopathy/tropical spastic paraparesis (14, 50). Disease association with HTLV-2 is less clear, with only a few cases of leukemia and neurological diseases reported (20, 39, 40). The difference in pathogenesis between the two related viruses has yet to be elucidated but likely results from the activities of the regulatory and accessory proteins, as well as their distinct Env cell surface molecule interaction profiles (11, 22, 23, 53).
In addition to the archetypal structural and enzymatic ret-roviral genesgag,pol, andenv, HTLV carries twotrans -regu-latory genes,taxandrex, which are essential for efficient viral replication/transformation, as well as several accessory genes important for viral infection and persistence in vivo (27). The viral oncoprotein Tax acts totrans-activate transcription from the viral long terminal repeat (7, 10) and modulates the tran-scription/functions of numerous cellular genes involved in cell growth, cell cycle control, DNA repair, and cell differentiation (6, 30, 35, 36, 43). The pleiotropic affects of Tax on these
numerous cellular processes are critical for its ability to induce cellular transformation and oncogenesis (11, 13, 38, 41).
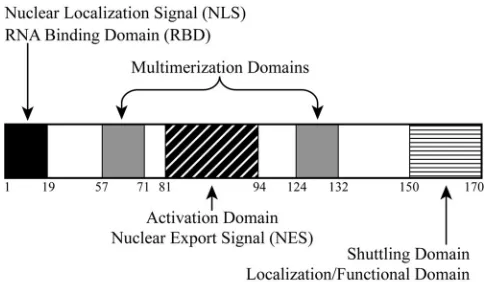
HTLV Rex is a positivetrans-acting regulator of viral rep-lication. Rex is a nuclear-localizing and shuttling phosphopro-tein that acts posttranscriptionally by binding and selectively exporting the unspliced and incompletely spliced viral mRNAs from the nucleus to the cytoplasm, thus controlling the expres-sion of the viral gene products that are essential for viral progeny production (19, 26). Therefore, it has been proposed that Rex is critical for the transition from the early, latent phase to the late, productive phase of HTLV infection. Rex binds viral mRNAs via acis-acting viral RNA structure termed the Rex response element (RxRE) located in the R region of the viral long terminal repeat (3, 33, 48). Previous analysis of HTLV-1 Rex (Rex-1), HTLV-2 Rex (Rex-2), and their RxREs revealed structural homologies and showed that they are func-tionally interchangeable (55, 56). Rex mutational analysis has defined several domains that are critical for its functions, in-cluding an arginine-rich N-terminal sequence that functions as an RNA binding domain (RBD), which also overlaps with a nuclear localization signal, a leucine-rich central core activa-tion domain that contains a nuclear export signal, and two flanking Rex-Rex multimerization domains (4, 5, 18, 21, 32, 34, 37, 45, 51). More recently, it has been reported that Rex-2 contains a novel C-terminal inhibitory/stability functional do-main that is positively regulated through phosphorylation (31, 32, 54).
Phosphorylation is a well-documented regulatory event with a pivotal role in controlling the activities/functions of proteins
* Corresponding author. Mailing address: The Ohio State Univer-sity, 1925 Coffey Rd., Columbus, OH 43210. Phone: (614) 688-4899. Fax: (614) 292-6473. E-mail: [email protected].
䌤Published ahead of print on 24 June 2009.
8859
on November 8, 2019 by guest
http://jvi.asm.org/
(47). Earlier studies demonstrated that both Rex-1 and Rex-2 are phosphoproteins and that this modification is critical to their function (1, 2, 17). In HTLV-2-infected T cells and cell lines transfected with Rex-2 expression plasmids, two species of Rex-2 (p24Rexand p26Rex) are detected. Both Rex-2 species
have the same amino acid backbone and differ by a charge-induced conformational change due to a serine/threonine phosphorylation (15, 16, 31). This altered gel mobility is unique to Rex-2, since Rex-1 presents as a single 27-kDa phosphopro-tein. We reported the identification of important Rex-2 phos-phorylation sites using targeted serine- or threonine-to-alanine substitution mutational analysis. That study identified at least one critical phosphorylation site at serine 151 (31). A serine-to-alanine substitution at this residue (S151A) resulted in re-duced phosphorylation, impaired RNA binding capability, dif-fused cytoplasmic localization, and decreased functional activity. In contrast, replacement of the serine with an aspartic acid (S151D) resulted in detection of only the p26Rexspecies in
cells. The S151D mutant was found to be more active than wild-type (wt) Rex-2 and displayed enhanced RNA binding capacity and intense localization to the nucleus (31, 32).
Our earlier studies suggested a linear relationship between phosphorylation, conformational changes, and Rex-2 function, implicating phosphorylation as a key regulatory event within the productive life cycle of the virus (31). To understand the regulation of Rex-2 function by phosphorylation, we per-formed complete phosphorylation mapping of both p24Rexand
p26Rex. In this study, we combined liquid
chromatography-tandem mass spectrometry (LC-MS/MS) analysis (28) of affin-ity-purified Rex-2 protein with substitution mutational analysis to identify and functionally characterize key phosphorylation sites. We achieved 100% coverage of both p24Rexand p26Rex.
We identified two phosphorylation sites in p24Rexand six total
phosphorylation sites in p26Rex. We also determined three
specific amino acid phosphorylation events that were found to be critical for Rex-2 function in vivo (Ser-151, Ser-153, and Thr-164). Phosphorylation at Ser-151 and Ser-153 is the major switch event for the conformational change between p24Rex
and p26Rex. Phosphorylation of one of these sites (Ser-151)
was critical for nuclear subcellular localization of the Rex-2 protein. Taken together, our data support a model in which a phosphorylation continuum of Rex-2 at the carboxy terminus regulates its biological properties and, ultimately, viral repli-cation.
MATERIALS AND METHODS
Cells.293T and HeLa-Tat cells were maintained in Dulbecco’s modified Ea-gle’s medium at 37°C in a humidified atmosphere of 5% CO2in air. The medium was supplemented with 10% fetal bovine serum, 2 mM glutamine, penicillin (100 U/ml), and streptomycin (100g/ml).
Mammalian expression plasmid.The Rex-2 expression vector BC20.2, which contains the HTLV-2tax/rexcDNA expressed from the cytomegalovirus (CMV) immediate-early gene promoter, was described previously (16, 55). The S-tagged expression vector S-Rex-2 was constructed by inserting the HTLV-2rexopen reading frame from BC20.2 into pTriEx4-Neo (Novagen, Madison, WI) in frame with the amino-terminal His tag and S tag via SmaI and BamHI. All generated rexexpression vectors contained a previously described mutation in the overlap-pingtaxreading frame (F4Term), which had no effect on the Rex amino acid sequence but severely truncated Tax, completely knocking out its expression and function (42). The variousrex-2targeted mutations were generated using the QuikChange site-directed mutagenesis kit (Stratagene, La Jolla, CA). All muta-tions were confirmed by DNA sequence analysis, and vector expression was
verified by transfection and Western blot analysis. The human immunodeficiency virus type 1 (HIV-1) Tat expression vector, pcTat; Rex-2 reporter plasmid (pCgagRxRE-II); and CMV-luciferase (firefly) (CMV-luc) were described pre-viously (31).
Rex functional reporter assay.The Rex-2 functional assay was performed as described previously with slight modifications (31). Briefly, 0.1g Rex-2 cDNA expression plasmids were cotransfected into 293T cells with 0.05g of CMV-luc, 0.25g of pcTat, and 0.5g of Rex reporter plasmid pCgag-RxRE-II using Lipofectamine reagent (Invitrogen, Carlsbad, CA). Cell lysates were prepared at 24 h posttransfection in Passive Lysis Buffer (Promega, Madison, WI) with a protease inhibitor mixture (Roche Applied Science, Indianapolis, IN) on ice for 30 min. Luciferase activity was determined and used as a transfection efficiency control. HIV-1 p24 Gag levels in the cellular lysates were determined by enzyme-linked immunosorbent assay (ZeptoMetrix, Buffalo, NY). All transfection ex-periments were performed in triplicate in three independent exex-periments.
Immunoblot and immunofluorescence analyses.Cell lysates were prepared at 24 h posttransfection in Passive Lysis Buffer (Promega, Madison, WI) with a protease inhibitor mixture (Roche Applied Science, Indianapolis, IN) on ice for 30 min. After centrifugation, total protein concentrations were determined by Bradford protein assay (Bio-Rad). The cell lysates were subjected to 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose (Schleicher & Schuell Biosciences, Keene, NH). Western blots were performed as recommended by the manufacturer, and proteins were visu-alized using an enhanced-chemiluminescence–Western blot analysis system (Santa Cruz Biotechnology, Santa Cruz, CA). Rex-2 was detected using protein-specific rabbit polyclonal antisera. Subcellular localization of Rex-2 was per-formed as previously described (49) with slight modifications. HeLa-Tat cells were transfected with 2g of vector control plasmid, wt Rex-2, or various Rex-2 mutants. At 24 h posttransfection, the cells were washed and fixed in phosphate-buffered saline containing 2% paraformaldehyde and permeabilized in phos-phate-buffered saline containing 0.2% Triton X-100 and 0.5% fetal bovine serum for 15 min at 4°C. The cells were incubated in blocking buffer (0.5% fetal bovine serum and 2 mg/ml human immunoglobulin G) for 30 min at room temperature. Staining was conducted in blocking buffer with rabbit Rex-2-specific antisera, followed by incubation with secondary antibody conjugated to fluorescein iso-thiocyanate Alexa Fluor 488 (Molecular Probes, Eugene, OR). Nuclear staining was performed using 4⬘6-diamidino-2-phenylindole (DAPI) Slow Fade Gold (Invitrogen, Carlsbad, CA). Fluorescence was visualized on an epifluorescence microscope (Olympus, Melville, NY), and digital images were taken using an Optronics (Goleta, CA) imaging system.
Purification of Rex-2 protein.Protein purification was performed as described previously with slight modifications (9). Briefly, the cell lysate (1.5-ml volume) was incubated with a 75-l bed volume of S protein-agarose (Novagen, Gibbs-town, NJ) overnight at 4°C and washed twice with a high-salt modified RIPA buffer (0.05 M Tris-HCl, pH 8.0, 0.1% SDS, 1% Triton X-100, 1.0 M NaCl, 0.01 M EDTA) and twice with a low-salt modified RIPA buffer (0.05 M Tris-HCl, pH 8.0, 0.1% SDS, 1% Triton X-100, 150 mM NaCl). The S protein-agarose beads were resuspended in 100l SDS loading dye containing-mercaptoethanol and subsequently boiled for 2 min. Samples were electrophoresed in a 12% SDS one-dimensional polyacrylamide gel and visualized by Coomassie blue staining. Both the p24Rex
and p26Rex
bands were excised from the gel separately for further proteomic analysis.
MS analysis.LC-MS/MS analysis was performed as described previously with slight modifications (9). Briefly, p24Rexand p26Rexexcised gel slices were cut into 1- to 2-mm cubes, washed three times with 500l ultrapure water, and incubated in 100% acetonitrile for 45 min. Samples were reduced with 50 mM dithiothreitol at 56°C for 45 min and then alkylated with 55 mM iodoacetamide for 1 h at room temperature. The material was dried in a speed-vac, rehydrated in a 12.5-ng/l modified sequencing-grade trypsin solution (Promega, Madison, WI), and incu-bated in an ice bath for 40 to 45 min. The trypsin solution was removed and replaced with 40 to 50l of 50 mM ammonium bicarbonate and 10% acetonitrile (pH 8.0), and the mixture was incubated overnight at 37°C. Elastase digestions were performed as described for trypsin at an enzyme concentration of 15 ng/l in the absence of acetonitrile in the reaction buffer. Peptides were extracted two times with 25l 50% acetonitrile, 5% formic acid and dried in a speed-vac. The digests were resuspended in 20l buffer A (5% acetonitrile, 0.1% formic acid, 0.005% heptafluorobutyric acid), and 3 to 6l was loaded onto a 12-cm by 0.075-mm fused silica capillary column packed with 5m-diameter C18beads (The Nest Group, Southboro, MA) using an N2pressure vessel at 1,100 lb/in2. The peptides were eluted over 55 min by applying a 0 to 80% linear gradient of buffer B (95% acetonitrile, 0.1% formic acid, 0.005% heptafluorobutyric acid) at a flow rate of 150l/min with a precolumn flow splitter, resulting in a final flow rate of⬃200 nl/min directly into the source. In some cases, the gradient was
on November 8, 2019 by guest
http://jvi.asm.org/
extended to 150 min to acquire more MS/MS spectra. An LTQ Linear Ion Trap (ThermoFinnigan, San Jose, CA) was run in automated collection mode with an instrument method composed of a single segment and five data-dependent scan events with a full MS scan followed by four MS/MS scans of the highest-intensity ions. The normalized collision energy was set at 35, and activation quadrupole was 0.250 with minimum full scan signal intensity at 1⫻105
, with no minimum MS/MS intensity specified. Dynamic exclusion was turned on, utilizing a 3-minute repeat count of 2, with the mass width set at 1.0m/z. Sequence analysis was performed with TurboSEQUEST (ThermoFinnigan, San Jose, CA) or MASCOT (Matrix Sciences, London, United Kingdom) using an indexed human subset database of the nonredundant protein database from the National Center for Biotechnology Information website (http://www.ncbi.nlm.nih.gov).
RESULTS
Functional domains of HTLV-2 Rex.Two species of Rex-2 (p24Rexand p26Rex) are detected in virus-infected cells. The
two Rex-2 species have the same amino acid backbone and differ by a charge-induced conformational change due to a serine/threonine phosphorylation (15, 16, 31). The p24Rex
spe-cies is incapable of binding target mRNA and is found primar-ily in the cytoplasm, whereas the p26Rex species is localized
primarily to the nucleus, and its presence is correlated with Rex-2-specific RxRE target RNA binding (17, 56). Mutational analysis permitted the assignment of functional properties to distinct domains of the Rex-2 protein (Fig. 1). In addition to the nuclear localization signal/RBD, central core activation domain/nuclear export signal, and multimerization domains that were also shared with the functionally homologous HTLV-1 Rex (32), we also reported that Rex-2 contained a unique C-terminal inhibitory/stability domain that was found to be important for proper nucleocytoplasmic shuttling, sub-cellular localization, and function (32, 54). This domain also encompasses at least one phosphorylated serine residue (S151), identified by substitutional mutational and peptide phosphoamino acid analysis, that is critical for Rex-2 function in vivo (31, 32). Replacement of this serine with aspartic acid to mimic phosphorylation (S151D) resulted in detection of
only the p26Rexspecies in cells. The S151D mutant was more
functionally active than wt Rex-2 and was locked in a p26Rex
conformation, since in vitro phosphatase treatment failed to alter its mobility. Therefore, since phosphorylation of Rex-2 may be a key regulatory event, a complete analysis of Rex-2 phosphorylation is well justified.
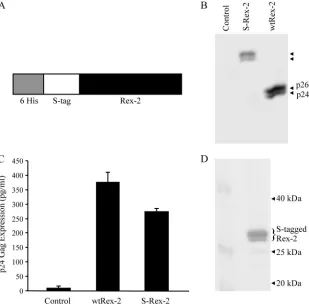
Expression and detection of biologically active affinity S-tagged Rex-2 protein.To determine the distinct phosphoryla-tion events present in both p24Rex and p26Rex that occur in
vivo, we used a tandem affinity-tagged Rex-2 construct that was expressed in and purified from mammalian cells. The Rex-2 vector (S-Rex-2) expresses full-length Rex-2 protein fused to amino-terminal His6 and S tags (Fig. 2A). Since the HTLV regulatory proteins Tax and Rex are translated from the same mRNA in partially overlapping reading frames, we utilized a cDNA mutant sequence containing an amino-terminal stop codon in the Tax reading frame, leaving the Rex-2 reading frame unchanged (41). This mutation abrogated Tax protein expression (data not shown). The S-Rex-2 and the wt Rex-2 expression constructs were transiently transfected into 293T cells, and protein expression was determined by Western blot analysis using rabbit polyclonal anti-Rex antisera (Fig. 2B). As expected, the wt-Rex-2-transfected cells produced both p24Rex
and p26Rex species. S-Rex-2-transfected cells also produced
two distinct species of increased molecular weight, consistent with the addition of the tags. We performed further analyses to confirm that the S-tagged Rex-2 species had properties similar to those of wt Rex-2 (p24Rex and p26Rex). Pro Q diamond
staining, a fluorescence method for detection of phosphopro-teins in polyacrylamide gels, as well as protein phosphatase treatment, indicated that the upper band was a phosphorylated form of the lower band, consistent with what had been re-ported previously for wt Rex-2 (reference 17 and data not shown). We then tested S-Rex-2 for its ability to function in our quantitative reporter bioassay, in which HIV-1 p24 Gag production was measured and used as a readout of Rex func-tional activity (31). As shown in Fig. 2C, S-Rex-2 displayed significant functional activity, although slightly lower than that of wt Rex-2. We hypothesize that this reduced activity is likely due to the proximity of the amino-terminal tag to the RBD. Taken together, these data demonstrate the proper and effi-cient expression of a functionally active S-tagged Rex-2 from mammalian cells.
Affinity purification of Rex-2 from mammalian cells. We successfully purified S-tagged Rex-2 protein from transfected 293T cells using S protein-agarose beads as described in Ma-terials and Methods. This purification procedure is based on the strong affinity between the 15-amino-acid S tag and the S protein that is immobilized on the agarose beads, both of which are derived from RNase S (25). The affinity-purified Rex-2 protein was resolved by SDS-PAGE, in which both spe-cies of S-tagged Rex-2, equivalent to wt p24Rex and p26Rex,
were detected by Coomassie blue staining (Fig. 2D). This pu-rification process produced adequate quantities of highly purified Rex-2 protein from mammalian cells and allowed the subsequent posttranslational modification analysis by LC-MS/MS.
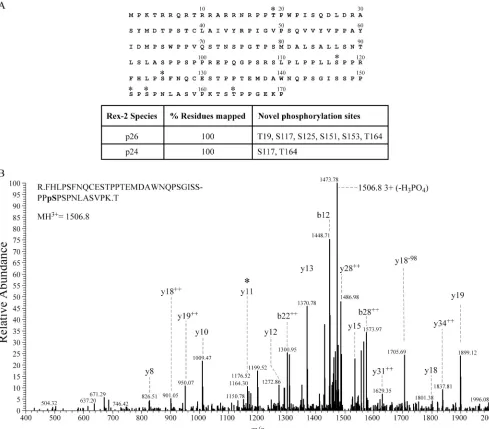
[image:3.585.41.283.68.209.2]Phosphopeptide mapping of Rex-2 using LC-MS/MS. Mul-tiple strategies were employed to identify the phosphorylation sites within Rex-2. The two species of S-tagged Rex-2 were excised and treated separately. This ensured that we could
FIG. 1. Functional domains of HTLV-2 Rex. The functional do-mains of the 170-amino-acid Rex-2 protein are depicted in shaded boxes. The nuclear localization signal and the RBD are positioned within the first 19 amino acids of the N terminus. The activation domain and the nuclear export signal are located between residues 81 and 94. This region is flanked by the two multimerization domains. The first lies between amino acids 57 and 71, whereas the second spans amino acids 124 to 132. The C-terminal domain that lies between amino acids 150 and 170 has been reported to be important for Rex-2 functions, such as nucleocytoplasmic shuttling, and the domain also plays a functional role in protein localization and function.
on November 8, 2019 by guest
http://jvi.asm.org/
determine the specific phosphorylation maps of each species independently in an effort to better understand how phosphor-ylation alters Rex-2 conformation and regulates its function. Initially, the proteins were subjected to trypsin enzymatic di-gestion. The tryptic peptides that were too large to detect were either further digested with elastase or independently digested with elastase. This combined analytical approach allowed us to obtain a detailed physical map covering 100% of the polypep-tide for both species of S-tagged Rex-2 (Fig. 3A). Our analysis identified two phosphorylation sites, Ser-117 and Thr-164, in the lower species (p24Rexequivalent). In addition to Ser-117
and Thr-164, we also identified four additional residues (Thr-19, Ser-125, Ser-151, and Ser-153) that are phosphorylated in the upper S-tagged species (p26Rex equivalent). Figure 3B
shows a representative MS/MS spectrum of the tryptic phos-phopeptide that identified phosphorylation at Ser-151. Fur-thermore, phosphorylation of Ser-151 was previously shown to be important for Rex-2 subcellular localization, RNA binding, and Rex-2 function in vivo (31).
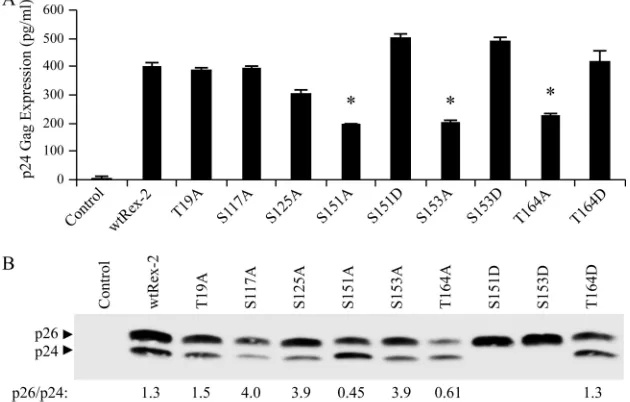
Substitution mutational analysis of the identified Rex-2 phosphorylation sites.To determine possible regulatory roles of the six identified phosphorylation sites, we generated single alanine amino acid substitutions and tested these Rex-2
mu-tants for the ability to function in our quantitative reporter bioassay. The Rex-2 mutants were transiently cotransfected into 293T cells with pcTat and pCgagRxRE-II, along with a CMV-luc transfection efficiency control. We identified three mutants, S151A, S153A, and T164A, within the C-terminal domain of Rex-2 that displayed significantly reduced function (Fig. 4A). Further mutational analysis of these three residues by converting them to aspartic acid (S151D, S153D, and T164D), which mimics a phosphoserine or threonine, restored functional activity to wt Rex-2 levels, indicating that phosphor-ylation plays a positive functional role (Fig. 4A). Aspartic acid substitution for Thr-19, Ser-117, or Ser-125 had no effect on protein function, consistent with the conclusion that phosphor-ylation of these three residues does not negatively regulate function but is silent (data not shown). The steady-state ex-pression levels of the wt and mutant Rex-2 proteins were determined by Western blot analysis using rabbit polyclonal anti-Rex antisera (Fig. 4B). All of the Rex-2 mutants were stably expressed. With the exception of S151D and S153D, detected only as p26Rex, all of the mutants displayed both the
p26Rex and p24Rex forms. We previously reported that the
p26Rex/p24Rex ratio of wt Rex in transfected cells is greater
than or equal to 1 (31). One exception was mutant S151A,
FIG. 2. Efficient expression and functional activity of Rex-2. (A) Illustration of the S-tagged Rex-2 (S-Rex-2) expression vector construct (not drawn to scale). (B) Western blot of S-Rex-2 protein expressed from 293T cells transiently transfected with Rex-2 cDNA plasmids. Proteins were detected using rabbit Rex-2-specific antisera. Wt p24Rexand p26Rexare indicated, and the arrowheads identify S-tagged Rex-2 proteins. (C) The
functional activity of S-Rex-2 was determined using the modified HIV p24 Gag reporter assay. 293T cells were transfected with 0.25g pcTat, 0.5 g pCgagRxRE-II, 0.05g CMV-luc, and 0.1g of wt Rex-2 or S-Rex-2 plasmids. Twenty-four hours posttransfection, cells were harvested and assayed for p24 Gag. The values represent actual p24 Gag production from a representative experiment performed in triplicate. The error bars indicate standard deviations. (D) Affinity purification of S-tagged Rex-2 from mammalian cells. 293T cells were transfected with S-Rex-2, purified with S protein-agarose beads, eluted and resolved by SDS-PAGE analysis, and detected by Coomassie blue staining.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:4.585.138.447.69.373.2]which showed a reduction in the amount of p26Rexrelative to
p24Rex, implying a role for phosphorylation of this residue in
altering the conformation/mobility of the protein. It is notable that T164A displayed a similar reduction in p26Rex, indicating
that phosphorylation of both Thr-164 and Ser-151 is correlated with p26Rex expression. Moreover, replacing threonine with
aspartic acid (T164D) resulted in an increase in p26Rexand the
restoration of function to wt levels (Fig. 4A), which also is consistent with the conclusion that p26Rexis the active form of
the protein. Taken together, our results help further define a critical functional domain that is regulated via phosphorylation of three key amino acids, Ser-151, Ser-153, and Thr-164. Since
p24Rexwas already found to contain phosphorylated Thr-164,
we hypothesize that the altered ratio of p24Rexto p26Rexfound
in the T164A mutant and previously reported for S151A is indicative of an important role for this residue in the Rex-2 phosphorylation continuum.
Subcellular localization of Rex-2 carboxy terminus mutants.
We next wanted to determine if the phosphorylation of key Rex-2 residues identified in the carboxy terminus regulates subcellular localization of the protein. It has been docu-mented that p26Rex is found primarily in the nucleus and
nucleolus, whereas the inactive p24Rexis diffusely dispersed
[image:5.585.45.534.69.498.2]throughout the cytoplasm and nucleus (32, 56). We utilized
FIG. 3. Mapping Rex-2 p24 and p26 phosphorylation sites by MS. (A) A compilation of the results obtained with LC-MS/MS analysis of Rex-2. The 170-amino-acid Rex-2 protein is depicted with phosphorylation sites identified (*). The table below shows percent total amino acid coverage from LC-MS/MS analysis for both p24Rexand p26Rex. (B) A representative MS/MS spectrum of the tryptic phosphopeptide R.FHLPSFNQCES
TPPTEMDAWNQPSGISSPPpSPSPNLASVPK.T (p denotes phosphorylation, and the period denotes tryptic cleavage sites), which identified phosphorylation at Ser-151, is shown. A collision-induced dissociation mass spectrum ofm/z1506.8 (3⫹) revealed a 41-amino-acid peptide with anMrof 4,517.3. B and Y ion designations marked with⫹⫹are doubly charged. The presence of the y11 ion (*) and the accompanying upstream
y series to S-158 mapped the phosphorylation to Ser-151. The MASCOT peptide score was 77, with an expected score of 0.00043.
on November 8, 2019 by guest
http://jvi.asm.org/
indirect immunofluorescence to assess whether the three phosphor acceptor residues (Ser-151, Ser-153, and Thr-164) that were critical for function in our bioreporter assay were important for nuclear localization. HeLa-Tat cells were transiently transfected with wt Rex-2 and each of the alanine or aspartic acid substitutional mutants. Wt Rex, as well as all three phosphomimetic mutants (S151D, S153D, and T164D), localized predominantly to the nucleus with the detection of very minimal cytoplasmic staining (Fig. 5).
[image:6.585.135.450.74.275.2]Mu-tant S151A was widely diffused throughout the cell with significant cytoplasmic staining (Fig. 5). Mutants S153A and T164A showed significant nuclear staining, similar to wt Rex, but also displayed moderate cytoplasmic staining. Tak-ing the data together, we conclude that although phosphor-ylation of Ser-151, Ser-153, and Thr-164 is pivotal for Rex-2 function, only the substitution of the Ser-151 phosphor ac-ceptor residue for alanine resulted in a significant change in subcellular localization to the cytoplasm.
FIG. 4. Mutational analysis of Rex-2 phosphospecific mutants. (A) The functional activities of either wt Rex-2 or Rex-2 mutants, as indicated, were determined using the modified HIV p24 Gag reporter assay. The specific amino acid substitution for each Rex-2 mutant is shown. 293T cells were transfected with 0.25g pcTat, 0.5g pCgagRxRE-II, 0.05g CMV-luc, and 0.1 g of wt Rex-2 or Rex-2 mutant plasmids. At 24 h posttransfection, cells were harvested and assayed for p24 Gag. The values represent actual p24 Gag production from a representative experiment performed in triplicate. The error bars indicate standard deviations. (B) Western blot analysis of wt and Rex-2 mutants. Whole-cell lysates normalized for transfection efficiency were subjected to Western blotting using rabbit Rex-2-specific antisera. p24Rexand p26Rexare indicated.
Densitometry was used to quantitate the amounts of p26Rexand p24Rex, and the numbers below the gel indicate the p26/p24 ratios.
FIG. 5. Subcellular localization of Rex-2 mutants. To determine the subcellular localization of the Rex-2 mutants, HeLa-Tat cells were transfected with 1g of a control plasmid, wt Rex-2, or various Rex-2 mutants. At 24 h posttransfection, the cells were stained with rabbit Rex-2-specific antisera (green). Nuclei were stained with DAPI (blue).␣-Rex, anti-Rex.
on November 8, 2019 by guest
http://jvi.asm.org/
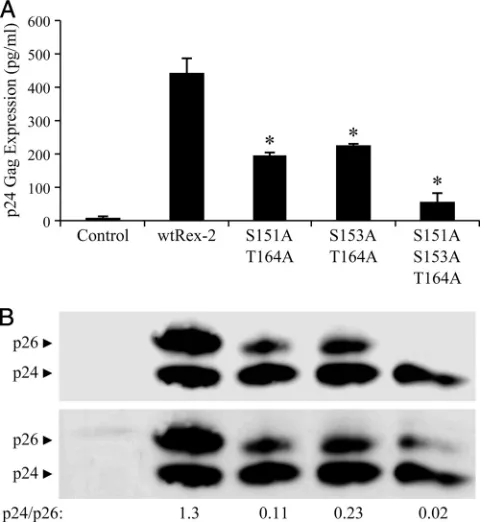
Protein expression and functional activity of Rex-2 combi-nation mutants. Combining the results from this study with previous published work suggests that phosphorylation of the carboxy terminus of Rex-2 triggers a protein conformation change that is correlated with nuclear localization, protein stability, and function. Since individual mutations at the car-boxy terminus still maintain partial function and p24Rexshows
Thr-164 phosphorylation, we hypothesized that phosphoryla-tion occurs on a continuum and that phosphorylaphosphoryla-tion of all three residues is required for optimal biological activity. To test this hypothesis and to determine if there is a functional relationship between Ser-151, Ser-153, and Thr-164, we gen-erated and characterized a combination of Rex-2 alanine mu-tants for function and protein species expression. As shown in Fig. 6, the two double mutants S151A T164A and S153A T164A display similar but significantly reduced functional ac-tivity compared to wt Rex-2. However, their functional acac-tivity was comparable to single-alanine-substitution mutants at ei-ther of those residues (compare Fig. 2A and Fig. 6). As seen with the S151A and T164A single mutants, Western blot anal-ysis of the double mutants revealed a reduction of p26Rex
relative to p24Rex, and the functional activity was directly
cor-related with the levels of p26Rexdetected. The function of the
S151A S153A T164A triple mutant was knocked down even further, but some activity over background levels was observed. Western blot analysis revealed a significant and nearly com-plete loss of the p26Rex species (Fig. 6). Some p26Rex was
detected on longer exposure, which was correlated with the residual functional activity. Taken together, these data allow us to formulate a new, more complex Rex-2 phosphorylation con-tinuum model that initiates with phosphorylation of Thr-164, followed by subsequent phosphorylation of 151 and Ser-153, thus tightly regulating Rex-2 function and, ultimately, virus production.
DISCUSSION
Phosphorylation is a well-known regulatory event that plays a key role in controlling the functions of cellular and viral proteins (1, 24, 29, 46, 47, 52). The modulation of protein function can occur by multiple mechanisms, including regula-tion of subcellular localizaregula-tion, protein-protein interacregula-tions, protein stability, and protein-nucleic acid binding. Previously, it was demonstrated that phosphorylation of HTLV-2 Rex, predominantly on serine residues, was correlated with an al-tered conformation, as observed by a gel mobility shift and the detection of p24Rexand p26Rexprotein species (16, 31). Rex-2
phosphorylation and the presence of the p26Rexspecies were
correlated with functional activity, including specific binding to its viral mRNA target sequence, cytoplasmic export, and trans-lation of the target mRNA (31, 32). Moreover, we recently reported that the carboxy terminus of Rex-2 contains a stabil-ity/inhibitory domain that is positively regulated through phos-phorylation (54). However, a complete map of site-specific phosphorylation of Rex-2 is lacking, and the precise role it plays in protein function remains unclear. The goal of this study was to use affinity purification and LC-MS/MS to identify all phosphorylation sites in both p24Rexand p26Rexto provide
insight into a possible phosphorylation continuum and the mechanism that controls Rex-2 function and ultimately the “on/off” switch that regulates productive viral replication. Con-sistent with previous reports, we confirmed that Rex-2 is phos-phorylated predominantly on serine residues and some threo-nine residues, and although the p26Rex species is heavily
phosphorylated, p24Rexcontains minimal phosphorylation. We
reported the phosphorylation of p24Rexat two sites, Ser-117
and Thr-164. We also reconfirmed phosphorylation of Ser-151 in p26Rex(31) and identified five additional phosphorylation
sites, Thr-19, Ser-117, Ser-125, Ser-153, and Thr-164. Using mutational analysis replacing the identified phosphor acceptor residues in Rex-2 with alanine or aspartic acid (which mimics phosphoserine or threonine), we showed that (i) the site-specific phosphorylations at Ser-151, Ser-153, and Thr-164 all positively contributed to Rex-2 function; (ii) the phosphor-ylation of Ser-151, Thr-164, and, to a lesser extent, Ser-153 dictate the p24Rex/p26Rexratio and thus Rex conformational
switching; and (iii) the phosphorylation of Ser-151 regulates the ability to efficiently localize to the nucleus. Both p24Rexand
p26Rexare phosphorylated at Ser-117 and Thr-164, but only
[image:7.585.43.285.70.331.2]phosphorylation at Thr-164 is functionally significant. Thus, since we do not have evidence that Ser-151 or Ser-153 is phos-phorylated without prior phosphorylation of Thr-164, a logical
FIG. 6. Functional analysis of Rex-2 S151A, S153A, and T164A combination mutants. (A) Functional activities of either wt Rex-2 or Rex-2 mutants, as indicated, were determined using the modified HIV p24 Gag reporter assay. 293T cells were transfected with 0.25 g pcTat, 0.5g pCgagRxRE-II, 0.05 g CMV-luc, and 0.1g of wt Rex-2 or Rex-2 plasmids. Twenty-four hours posttransfection, cells were harvested and assayed for p24 Gag. The values represent actual p24 Gag production from a representative experiment performed in triplicate. The error bars indicate standard deviations. (B) Western blot analysis of wt Rex-2 and Rex-2 mutants. Whole-cell lysates nor-malized for transfection efficiency were subjected to Western blotting using rabbit Rex-2-specific antisera. p24Rexand p26Rexare indicated.
Densitometry was used to quantitate the amounts of p26Rex and
p24Rex, and the numbers below the gel indicate the p26/p24 ratios.
on November 8, 2019 by guest
http://jvi.asm.org/
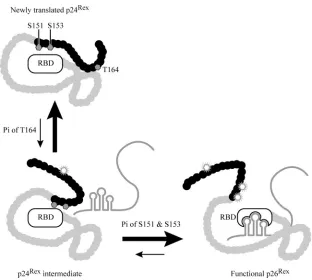
conclusion is that phosphorylation occurs on a continuum, with phosphorylation of Thr-164 an earlier initiating event that likely facilitates subsequent phosphorylation of Ser-151 and/or Ser-153 and Rex function. In Fig. 7, we present a model, consistent with the published literature and our data, showing how phosphorylation regulates Rex-2 function. The initial translation product of Rex-2, p24Rex, is located primarily in the
cytoplasm. It is an inactive species in a structural conformation in which the C terminus is configured in such a way that it prevents efficient phosphorylation of Ser-151 and Ser 153 and generation of an active p26Rex mRNA binding protein. We
propose that the phosphorylation continuum starts with the phosphorylation of Thr-164, which begins the rotation of the C-terminal inhibitory domain, exposing Ser-151 and/or Ser-153 and making them available to be phosphorylated. This is also consistent with the phenotype of a previously characterized Rex-2 truncation mutant (S158Term) in which the 13 carboxy-terminal amino acids, including the Thr-164 phosphor accep-tor, are deleted; Rex S158Term, although less stable, is func-tionally more active than the wt Rex-2 (54). However, since proteins are dynamic and not fixed in space, as long as one or more key phosphor acceptor sites are available within the C terminus, a hierarchical phosphorylation pattern could be side-stepped at a low frequency in mutant Rex proteins. This is consistent with the functional and Western blot results of our single (S151A or T164A) and double (S151A T164A or S153A T164A) alanine mutants, which maintain some function, and
the expression of the active p26Rex. However, the triple alanine
mutant (T164A S151A S153A), which contains none of the identified key phosphor acceptor residues, shows minimal functional activity and very little detection of the active p26Rex
species. We have yet to experimentally identify the cellular kinase(s) that phosphorylates Rex on these key functional res-idues. However, using bioinformatics analyses, we previously reported (31) that Ser-151 falls in a predicted consensus site for phosphorylation by casein kinase 1 (Sp/Tp-X2-3-S/T-X).
Ser-153 also shows high homology to the casein kinase 1 con-sensus sequence, and Thr-164 is predicted to be phosphory-lated by either GSK3or casein kinase 1 (data not shown).
Typically, stable protein conformational changes require bond rotations. Interestingly, using proteomic protein identi-fication (mudpit) analysis, we recently identified the associa-tion of the cellular protein Pin1 in the Rex-2 pulldown complex (data not shown). Pin1 is a peptidylprolinecis-transisomerase of Ser/Thr peptide bonds N-terminal to proline residues. Pin1 is known to induce conformational changes that can affect enzymatic activities, phosphorylation status, subcellular local-ization, protein stability, and protein/protein interactions (8, 12, 24, 29, 44, 46, 58). One hypothesis consistent with our data is that once Thr-164 is phosphorylated, Pin1 could bind and cause a structural conformational change, rotating the inhibi-tory C-terminal domain and allowing access of the cellular kinase to phosphorylate Ser-151. The single mutants alone only hinder the ability of the kinase to reach its target
consen-FIG. 7. Rex-2 phosphorylation continuum and function model. The newly Rex-2-translated product p24Rexis inactive due to the structural and
spatial inhibition of the carboxy-terminal domain (black ribbon) that masks the RBD and nuclear localization signal. Initial phosphorylation (Pi) of threonine 164 induces a conformational change that results in a p24Rexintermediate. This intermediate can be stabilized further by continued
phosphorylation (Pi) of serine 151 and 153, facilitating the dislocation of the inhibitory C-terminal domain, resulting in a detectable conformation change/gel mobility shift. The conformation change, which results in nuclear localization, allows the fully functional and stable p26Rexprotein, via
its exposed RBD, to interact with the viral target mRNA (depicted with RxRE secondary structure).
on November 8, 2019 by guest
http://jvi.asm.org/
[image:8.585.136.448.65.345.2]sus sequence. For example, S151A or S153A could still be phosphorylated at Thr-164, facilitating subsequent phosphory-lation of either Ser-153 or Ser-151, respectively, resulting in the formation of p26Rexand modest function. In the case of the
mutant T164A, the C-terminal domain is not efficiently dislo-cated, and since a biological protein is not static, a minimal amount of phosphorylation can occur on both Ser-151 and Ser-153. Whereas in the double mutant we not only inhibit Thr-164 phosphorylation, and thus the conformational change of the C terminus, but also block any phosphorylation of Ser-151, with this mutant we detected a greater increase in the amount of p24Rex, which correlates with the reduced function
of the mutant protein, with the only possible phosphorylation being Ser-153.
The question arises as to whether this C-terminal phosphor-ylation domain is unique to Rex-2 or if it is also contained in the closely related Rex-1. Although the carboxy termini of the two related proteins at the amino acid level are quite divergent with no apparent Ser-151, Ser-153, and Thr-164 counterpart in Rex-1 (31, 32), we have previously shown that, like that of Rex-2, the carboxy terminus of Rex-1 does contain a stability domain (54). Rex-1 is known to be phosphorylated (1, 2), but further studies are needed to determine whether Rex-1 site-specific phosphorylation has a significant functional role.
Taken together, the data presented in this study provide a wealth of knowledge about the cellular signaling controlling Rex-2 protein function, structure, and localization. Further studies will be aimed at identifying the cellular proteins, in-cluding the kinase(s), involved in the regulation of Rex-2 func-tion.
ACKNOWLEDGMENTS
We thank Tim Vojt for preparation of the figures and Kate Hayes-Ozello for editorial comments.
This work was supported by a grant from the National Institutes of Health (CA100730) to P.L.G.
REFERENCES
1.Adachi, Y., T. D. Copeland, C. Takahashi, T. Nosaka, A. Ahmed, S. Oros-zlan, and M. Hatanaka.1992. Phosphorylation of the Rex protein of human T-cell leukemia virus type I. J. Biol. Chem.267:21977–21981.
2.Adachi, Y., T. Nosaka, and M. Hatanaka.1990. Protein kinase inhibitor H-7 blocks accumulation of unspliced mRNA of human T-cell leukemia virus type I (HTLV-I). Biochem. Biophys. Res. Commun.169:469–475. 3.Askjaer, P., and J. Kjems.1998. Mapping of miltiple RNA binding sites of
the human T-cell lymphotropic virus type 1 Rex protein with 5⬘- and 3⬘-Rex response elements. J. Biol. Chem.273:11463–11471.
4.Bogerd, H., and W. C. Greene.1993. Dominant negative mutants of human T-cell leukemia virus type I Rex and human immunodeficiency virus type 1 Rev fail to multimerize in vivo. J. Virol.67:2496–2502.
5.Bogerd, H. P., G. L. Huckaby, Y. F. Ahmed, S. M. Hanly, and W. C. Greene. 1991. The type 1 human T-cell leukemia virus (HTLV-I) Rex trans-activator binds directly to the HTLV-I Rex and the type 1 human immunodeficiency virus Rev RNA response elements. Proc. Natl. Acad. Sci. USA88:5704– 5708.
6.Boxus, M., J. C. Twizere, S. Legros, J. F. Dewulf, R. Kettmann, and L. Willems.2008. The HTLV-1 Tax interactome. Retrovirology5:76. 7.Cann, A. J., J. D. Rosenblatt, W. Wachsman, N. P. Shah, and I. S. Y. Chen.
1985. Identification of the gene responsible for human T-cell leukemia virus transcriptional regulation. Nature318:571–574.
8.Crenshaw, D. G., J. Yang, A. R. Means, and S. Kornbluth.1998. The mitotic peptidyl-prolyl isomerase, Pin1, interacts with Cdc25 and Plx1. EMBO J. 17:1315–1327.
9.Durkin, S. S., M. D. Ward, K. A. Fryrear, and O. J. Semmes.2006. Site-specific phosphorylation differentiates active from inactive forms of the hu-man T-cell leukemia virus type 1 Tax oncoprotein. J. Biol. Chem.281:31705– 31712.
10.Felber, B. K., H. Paskalis, C. Kleinman-Ewing, F. Wong-Staal, and G. N.
Pavlakis.1985. The pX protein of HTLV-I is a transcriptional activator of its long terminal repeats. Science229:675–679.
11.Feuer, G., and P. L. Green.2005. Comparative biology of human T-cell lynphotropic virus type 1 (HTLV-1) and HTLV-2. Oncogene24:5996–6004. 12.Fujimori, F., K. Takahashi, C. Uchida, and T. Uchida.1999. Mice lacking Pin1 develop normally, but are defective in entering cell cycle from G0arrest. Biochem. Biophys. Res. Commun.265:658–663.
13.Gatza, M. L., J. C. Watt, and S. Marriott.2003. Cellular transformation by the HTLV-I Tax protein, a jack-of-all-trades. Oncogene22:5141–5149. 14.Grant, C., K. Barmak, T. Alefantis, J. Yao, S. Jacobson, and B. Wigdahl.
2002. Human T cell leukemia virus type I and neurologic disease: events in bone marrow, peripheral blood, and central nervous system during normal immune surveillance and neuroinflammation. J. Cell Physiol.190:133–159. 15.Green, P. L., and I. S. Y. Chen.2001. Human T-cell leukemia virus types 1
and 2, p. 1941–1969.InD. M. Knipe, P. Howley, D. Griffin, R. Lamb, M. Martin, and S. Straus (ed.), Virology, 4th ed. Lippincott Williams & Wilkins, Philadelphia, PA.
16.Green, P. L., Y. Xie, and I. S. Y. Chen.1991. The Rex proteins of human T-cell leukemia virus type II differ by serine phosphorylation. J. Virol.65: 546–550.
17.Green, P. L., M. T. Yip, Y. Xie, and I. S. Y. Chen.1992. Phosphorylation regulates RNA binding by the human T-cell leukemia virus Rex protein. J. Virol.66:4325–4330.
18.Hammes, S. R., and W. C. Green.1993. Multiple arginine residues within the basic domian of HTLV-1 Rex are required for specific RNA binding and function. Virology193:41–49.
19.Hidaka, M., J. Inoue, M. Yoshida, and M. Seiki.1988. Post transcriptional regulator (rex) of HTLV-I initiates expression of viral structural proteins but suppresses expression of regulatory proteins. EMBO J.7:519–523. 20.Hjelle, B., O. Appenzeller, R. Mills, S. Alexander, N. Torrez-Martinez, R.
Jahnke, and G. Ross.1992. Chronic neurodegenerative disease associated with HTLV-II infection. Lancet339:645–646.
21.Hope, T. J., B. L. Bond, B. McDonald, N. P. Klein, and T. G. Parslow.1991. Effector domains of human immunodeficiency virus type 1 Rev and human T-cell leukemia virus type I Rex are functionally interchangeable and share an essential peptide motif. J. Virol.65:6001–6007.
22.Jones, K. S., K. Fugo, C. Petrow-Sadowski, Y. Huang, D. C. Bertolette, I. Lisinski, S. W. Cushman, S. Jacobson, and F. W. Ruscetti.2006. Human T-cell leukemia virus type 1 (HTLV-1) and HTLV-2 use different receptor complexes to enter T cells. J. Virol.80:8291–8302.
23.Jones, K. S., C. Petrow-Sadowski, D. C. Bertolette, Y. Huang, and F. W. Ruscetti.2005. Heparan sulfate proteoglycans mediate attachment and entry of human T-cell leukemia virus type 1 virions into CD4⫹T cells. J. Virol. 79:12692–12702.
24.Joseph, J. D., E. S. Yeh, K. I. Swenson, A. R. Means, and Winkler.2003. The peptidyl-prolyl isomerase Pin1. Prog. Cell Cycle Res.5:477–487. 25.Kim, J. S., and R. T. Raines.1993. Ribonuclease S-peptide as a carrier in
fusion proteins. Protein Sci.2:348–356.
26.Kusuhara, K., M. Anderson, S. M. Pettiford, and P. L. Green.1999. Human T-cell leukemia virus type 2 Rex protein increases stability and promotes nuclear to cytoplasmic transport ofgag/polandenvRNAs. J. Virol.73:8112– 8119.
27.Lairmore, M., and G. Franchini.2007. Human T-cell leukemia virus types 1 and 2, p. 2071–2106.InB. Fields, D. Knipe, P. Howley, R. Chanock, T. Monath, J. Melnick, B. Roizman, and S. Straus (ed.), Fields virology,vol. 5. Lippincott Williams, and Wilkins, Philadelphia, PA.
28.Loyet, K. M., J. T. Stults, and D. Arnott.2005. Mass spectrometric contri-butions to the practice of phosphorylation site mapping through 2003: a literature review. Mol. Cell Proteomics4:235–245.
29.Lu, K. P., and X. Z. Zhou.2007. The prolyl isomerase PIN1: a pivotal new twist in phosphorylation signalling and disease. Nat. Rev. Mol. Cell Biol. 8:904–916.
30.Mulloy, J. C., T. Kislyakova, A. Cereseto, L. Casareto, A. LoMonico, J. Fullen, M. V. Lorenzi, A. Cara, C. Nicot, C. Giam, and G. Franchini.1998. Human T-cell lymphotropic/leukemia virus type 1 Tax abrogates p53-in-duced cell cycle arrest and apoptosis through its CREB/ATF functional domain. J. Virol.72:8852–8860.
31.Narayan, M., K. Kusuhara, and P. L. Green.2001. Phosphorylation of two serine residues regulates human T-cell leukemia virus type 2 Rex function. J. Virol.75:8440–8448.
32.Narayan, M., I. Younis, D. M. D’Agostino, and P. L. Green.2003. Functional domain structure of human T-cell leukemia virus type 2 Rex. J. Virol. 77:12829–12840.
33.Ohta, M., H. Nyunoya, H. Tanako, T. Okamoto, T. Akagi, and K. Shimo-tohno.1988. Identification of acis-regulatory element involved in accumu-lation of human T-cell leukemia virus type II genomic mRNA. J. Virol. 62:4445–4449.
34.Palmeri, D., and M. H. Malim.1996. The human T-cell leukemia virus type 1 posttranscriptional trans-activator Rex contains a nuclear export signal. J. Virol.70:6442–6445.
35.Ramadan, E., M. Ward, X. Guo, S. S. Durkin, A. Sawyer, M. Vilela, C. Osgood, A. Pothen, and O. J. Semmes.2008. Physical and in silico
on November 8, 2019 by guest
http://jvi.asm.org/
proaches identify DNA-PK in a Tax DNA-damage response interactome. Retrovirology5:92.
36.Ressler, S., G. F. Morris, and S. J. Marriott.1997. Human T-cell leukemia virus type 1 Tax transactivates the human proliferating cell nuclear antigen promoter. J. Virol.71:1181–1190.
37.Rimsky, L., M. Duc Dodon, E. P. Dixon, and W. C. Greene.1989.trans -dominant inactivation of HTLV-I and HIV-1 gene expression by mutation of the HTLV-I Rex transactivator. Nature341:453–456.
38.Robek, M. D., and L. Ratner.1999. Immortalization of CD4⫹and CD8⫹ T-lymphocytes by human T-cell leukemia virus type 1 Tax mutants expressed in a functional molecular clone. J. Virol.73:4856–4865.
39.Rosenblatt, J. D., J. C. Gasson, J. Glaspy, et al.1987. Relationship between HTLV-II and atypical hairy-cell leukemia: a serologic study of hairy-cell leukemia patients. Leukemia1:397–401.
40.Rosenblatt, J. D., D. W. Golde, W. Wachsman, A. Jacobs, G. Schmidt, S. Quan, J. C. Gasson, and I. S. Y. Chen.1986. A second HTLV-II isolate associated with atypical hairy-cell leukemia. N. Engl. J. Med.315:372–377. 41.Ross, T. M., M. Narayan, Z. Y. Fang, A. C. Minella, and P. L. Green.2000.
Tax transactivation of both NFB and CREB/ATF is essential for Human T-cell leukemia virus type 2-mediated transformation of primary human cells. J. Virol.74:2655–2662.
42.Ross, T. M., S. M. Pettiford, and P. L. Green.1996. Thetaxgene of human T-cell leukemia virus type 2 is essential for transformation of human T lymphocytes. J. Virol.70:5194–5202.
43.Schmitt, I., O. Rosin, P. Rohwer, M. Gossen, and R. Grassmann.1998. Stimulation of cyclin-dependent kinase activity and G1- to S-phase transition in human lymphocytes by the human T-cell leukemia/lymphotropic virus type 1 Tax protein. J. Virol.72:633–640.
44.Shen, M., P. T. Stukenberg, M. W. Kirschner, and K. P. Lu.1998. The essential mitotic peptidyl-prolyl isomerase Pin1 binds and regulates mitosis-specific phosphoproteins. Genes Dev.12:706–720.
45.Siomi, H., H. Shida, S. H. Nam, T. Nosaka, M. Maki, and M. Hatanaka. 1988. Sequence requirements for nucleolar localization of human T cell leukemia virus type I pX protein, which regulates viral RNA processing. Cell 55:197–209.
46.Stukenberg, P. T., and M. W. Kirschner.2001. Pin1 acts catalytically to promote a conformational change in Cdc25. Mol. Cell7:1071–1083. 47.Tootle, T. L., and I. Rebay.2005. Post-translational modifications influence
transcription factor activity: a view from the ETS superfamily. Bioessays 27:285–298.
48.Toyoshima, H., M. Itoh, J.-I. Inoue, M. Seiki, F. Takaku, and M. Yoshida. 1990. Secondary structure of the human T-cell leukemia virus type I Rex-responsive element is essential for Rex regulation of RNA processing and transport of unspliced RNAs. J. Virol.64:2825–2832.
49.Voss, O. H., S. Batra, S. J. Kolattukudy, M. E. Gonzalez-Mejia, J. B. Smith, and A. I. Doseff.2007. Binding of caspase-3 prodomain to heat shock protein 27 regulates monocyte apoptosis by inhibiting caspase-3 proteolytic activa-tion. J. Biol. Chem.282:25088–25099.
50.Watanabe, T.1997. HTLV-1-associated diseases. Int. J. Hematol.66:257–278. 51.Weichselbraun, I., G. K. Farrington, J. R. Rusche, E. Bohnlein, and J. Hauber.1992. Definition of human immunodeficiency virus type 1 Rev and human T-cell leukemia virus type 1 Rex protein activation domain by func-tional exchange. J. Virol.66:2583–2587.
52.Whitmarsh, A. J., and R. J. Davis.2000. Regulation of transcription factor function by phosphorylation. Cell Mol. Life Sci.57:1172–1183.
53.Xie, L., and P. L. Green.2005. Envelope is a major viral determinant of the distinct in vitro cellular transformation tropism of human T-cell leukemia virus type 1 (HTLV-1) and HTLV-2. J. Virol.79:14536–14545.
54.Xie, L., M. Kesic, B. Yamamoto, M. Li, I. Younis, M. D. Lairmore, and P. L. Green.2009. HTLV-2 Rex carboxy terminus is an inhibitory/stability domain that regulates Rex functional activity and viral replication. J. Virol.83:5232– 5243.
55.Ye, J., L. Xie, and P. L. Green.2003. Tax and overlapping Rex sequences do not confer the distinct transformation tropisms of HTLV-1 and HTLV-2. J. Virol.77:7728–7735.
56.Yip, M. T., W. S. Dynan, P. L. Green, A. C. Black, S. J. Arrigo, A. Torbati, S. Heaphy, C. Ruland, J. D. Rosenblatt, and I. S. Y. Chen.1991. HTLV-II Rex protein binds specifically to RNA sequences of the HTLV LTR but poorly to the HIV 1 RRE. J. Virol.65:2261–2272.
57.Yoshida, M.2005. Discovery of HTLV-1, the first human retrovirus, its unique regulatory mechanisms, and insights into pathogenesis. Oncogene 24:5931–5937.
58.You, H., H. Zheng, S. A. Murray, Q. Yu, T. Uchida, D. Fan, and Z. X. Xiao. 2002. IGF-1 induces Pin1 expression in promoting cell cycle S-phase entry. J. Cell Biochem.84:211–216.