0022-538X/06/$08.00
⫹
0
doi:10.1128/JVI.80.9.4388–4395.2006
Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Subunit Stoichiometry of Human Immunodeficiency Virus Type 1
Envelope Glycoprotein Trimers during Virus Entry into Host Cells
Xinzhen Yang,
1,2* Svetla Kurteva,
1Xinping Ren,
1,2Sandra Lee,
3,4and Joseph Sodroski
1,2,5Department of Cancer Immunology and AIDS
1and Department of Biostatistical Science,
3Dana-Farber Cancer Institute, and
Department of Pathology, Division of AIDS,
2Harvard Medical School, Boston, Massachusetts 02115, and Department of
Biostatistics
4and Department of Immunology and Infectious Diseases,
5Harvard School of Public Health,
Boston, Massachusetts 02115
Received 3 November 2005/Accepted 15 February 2006
The envelope glycoproteins of human immunodeficiency virus type 1 (HIV-1) function as a homotrimer of
gp120/gp41 heterodimers to support virus entry. During the process of virus entry, an individual HIV-1
envelope glycoprotein trimer binds the cellular receptors CD4 and CCR5/CXCR4 and mediates the fusion of
the viral and the target cellular membranes. By studying the function of heterotrimers between wild-type and
nonfunctional mutant envelope glycoproteins, we found that two wild-type subunits within an envelope
glyco-protein trimer are required to support virus entry. Complementation between HIV-1 envelope glycoglyco-protein
mutants defective in different functions to allow virus entry was not evident. These results assist our
under-standing of the mechanisms whereby the HIV-1 envelope glycoproteins mediate virus entry and membrane
fusion and guide attempts to inhibit these processes.
Human immunodeficiency virus type 1 (HIV-1) encodes a
gp160 envelope precursor glycoprotein. HIV-1 envelope
gly-coprotein spikes mature by trimerization, glycosylation, and
proteolytic cleavage of the gp160 precursor into gp120 and
gp41 glycoproteins. The glycoproteins are then loaded onto
virions as a homotrimer of gp120 and gp41 heterodimers (14,
45). Viral entry is initiated by binding of the gp120
glycopro-tein subunits to the cellular receptor CD4 (9, 23). The
inter-action with CD4 leads to structural changes within gp120 that
allow binding to the coreceptors, either CCR5 or CXCR4 (1, 4,
8, 10, 11, 15, 30). CD4 binding also induces conformational
changes in the gp41 ectodomain that result in the exposure of
a trimeric coiled coil (20, 37). Formation of the gp120/CD4/
CCR5 complex is thought to result in exposure of the fusion
peptide, a highly hydrophobic region at the gp41 N terminus
that contacts the target cell membrane (18). Further
rear-rangements within the gp41 ectodomain result in the formation
of a six-helix bundle structure and the fusion of opposing
membranes from the virion and the target cell (6, 7, 13, 41, 43).
Peptide or small-molecule inhibitors targeting CD4-induced
conformational changes in gp120, gp120-coreceptor binding,
or six-helix bundle formation in gp41 have been shown to block
HIV-1 infection, representing promising new directions for
developing the next generation of anti-HIV-1 therapeutics (27,
35, 44, 49). The number of subunits within an HIV-1 envelope
glycoprotein trimer that is required for virus entry is unknown.
This information specifies the number of subunits that have to
be bound and inhibited by candidate drugs to limit HIV-1
infection.
Cooperativity among the three individual subunits in an
envelope glycoprotein trimer may allow efficient function. The
existence of such cooperativity was suggested by a study of the
effects of fusion-inducing pH decreases on conformational
changes in heterotrimers composed of hemagglutinin
glyco-proteins from two different strains of influenza virus (5).
Complementation in a cell-cell fusion assay among HIV-1
en-velope glycoprotein mutants defective in receptor binding and
fusion peptide function has been reported (36). Such
comple-mentation implies subunit cooperativity within a trimer and
suggests that fewer than three functional subunits can achieve
membrane fusion. However, these studies did not provide a
precise estimate of the number of functional subunits that are
required for a trimer to support virus entry. Interestingly,
sol-uble envelope glycoprotein trimers of simian
immunodefi-ciency virus apparently bind only a single molecule of soluble
CD4, whereas three antibody molecules can be bound (22). In
contrast, more than one soluble CD4 molecule could bind to a
soluble envelope glycoprotein trimer of HIV-1
ADA(48). In this
study, we attempted to estimate the subunit stoichiometry
within an envelope glycoprotein trimer during virus entry by
testing the function of heterotrimers of HIV-1 envelope
gly-coproteins carrying one or two loss-of-function mutant
sub-units.
MATERIALS AND METHODS
Theoretical model building.Recently, by studying viruses carrying heterotri-mers between the wild-type HIV-1 envelope glycoproteins and closely related dominant-negative or neutralization escape mutants, we demonstrated that an individual HIV-1 envelope glycoprotein trimer functions independently to me-diate virus entry (46, 47). This stoichiometry has been designated T⫽1, where T denotes the minimum number of trimers that function as a unit to mediate virus entry. The logical arguments, model-building techniques, and experimental ap-proaches applied in the present study are similar to those used previously to investigate the stoichiometry of HIV-1 entry and neutralization.
The HIV-1 gp160 precursor trimerizes soon after its synthesis in the endo-plasmic reticulum (12). Subsequently, proteolytic cleavage produces the mature, functional trimer composed of three gp120 and three gp41 subunits (19, 39). When two different species of viral glycoproteins are coexpressed in a cell, heterotrimers can form. Heterotrimers between two species of closely related
* Corresponding author. Mailing address: Dana-Farber Cancer
In-stitute, Department of Cancer Immunology and AIDS, 44 Binney
Street, JFB 824, Boston, MA 02115. Phone: (617) 632-4359. Fax: (617)
632-3113. E-mail: [email protected].
4388
on November 8, 2019 by guest
http://jvi.asm.org/
viral glycoproteins can form with an efficiency close to that predicted by random mixing of two species of monomeric subunits (5, 38). The frequency of different species of envelope glycoprotein trimers formed by random mixing is dependent on the frequency of mutant monomers (fM) in the pool of available envelope
glycoprotein monomers. Then, the frequency of different trimers in a viral stock can be calculated as follows: the frequency of the wild-type homotrimer is (1⫺ fM)
3
; the frequency of the heterotrimer with two wild-type subunits and one mutant subunit is 3(1⫺fM)2fM; the frequency of the heterotrimer with one
wild-type and two mutant subunits is 3(1⫺fM)fM
2
; and the frequency of the mutant homotrimer isfM3.
Let S be the number of wild-type subunits within an envelope glycoprotein trimer that is required for trimer function. For the purpose of model building, we assume that an individual envelope glycoprotein trimer functions in an all-or-none fashion; thus, a trimer with S or more wild-type subunits is assumed to function like a wild-type homotrimer, and a trimer with fewer than S wild-type subunits is considered to be totally nonfunctional. If all three subunits in a trimer are required for function (S⫽3 model), only the wild-type homotrimers, which are present at a frequency of (1⫺fM)3, are potentially functional in a viral stock.
In the S⫽2 model, in which two wild-type subunits in a trimer are required for function, both the type homotrimers and the heterotrimers with two wild-type subunits and one mutant subunit are potentially functional. In this case, the frequency of functional trimers is (1⫺fM)3⫹3(1⫺fM)2fMor 1⫺3fM2⫹2fM3.
In the S⫽1 model, in which only one wild-type subunit in an envelope glyco-protein trimer is required for function, all of the envelope glycoglyco-proteins except the mutant homotrimers are potentially functional. Thus, functional envelope glycoproteins exhibit a frequency of 1⫺fM3in this case.
HIV-1 stocks contain a very high proportion of functionally defective virions (often over 99.9%); thus, the chance that one HIV-1 virion carries more than one infectious unit is statistically negligible (29, 32). Because each envelope glyco-protein trimer functions independently (T⫽1) (46, 47), the infectivity of an HIV-1 viral stock should directly reflect the frequency of virion-associated tri-mers that are potentially functional. The experimentally determined infectivity of a virus preparation containing a given proportion of mutant envelope glycopro-teins can be expressed as a relative infectivity, RI(%), which is a percentage of the infectivity observed for a viral stock with only the wild-type envelope glyco-proteins. The relationships between thefMand RI(%) values of viral stocks
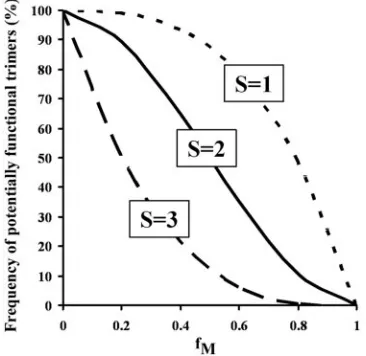
predicted by the three candidate models are shown in Fig. 1. Results consistent with the S⫽3 model can hypothetically be achieved in one of two ways: either by the requirement for three functional subunits in an envelope glycoprotein trimer or by the presence of a transdominant-negative mutant subunit. The latter
scenario has been observed previously with two transdominant-negative mutant envelope glycoproteins (47).
The overall relationship among S,fM, and RI(%) may be summarized as
RI(%)⫽{1⫺[fM3⫹3fM2(1⫺fM)(S⫺1)⫹3fM(1⫺fM)2(S⫺1)(S⫺2)/2⫺
3fM2(1⫺fM)(S⫺1)(S⫺2)/2]}⫻100, assuming that each envelope glycoprotein
trimer functions independently to mediate viral infection, i.e., T⫽1 (46, 47). Reduction of this formula by S⫽1, S⫽2, or S⫽3 would result in thefM/RI(%)
relationships for individual models, as described above.
Plasmids expressing HIV-1 envelope glycoproteins.The functionality of trim-ers composed of wild-type and mutant HIV-1 envelope glycoproteins was eval-uated using envelope glycoproteins derived from the HIV-1YU2isolate. HIV-1YU2is a primary isolate, uses CCR5 as a coreceptor, and is very resistant to neutralizing antibodies. The wild-type HIV-1YU2gp160 glycoproteins were ex-pressed from the pSVIIIenv vector (40). The glycoprotein mutants were created by the PCR-based QuikChange protocol (Stratagene). The D368R, R315G/ L317S, and L520E mutants disrupt CD4 binding, CCR5 binding, and membrane fusion, respectively (17, 34, 42). The residue numbers correspond to those of the prototypic HXBc2 envelope glycoproteins, according to current convention (25). The presence of the desired mutations and the absence of unintended coding changes were confirmed by DNA sequencing of the entireenvreading frame. All plasmids expressing the wild-type and mutant envelope glycoproteins were pre-pared using a QIAFilter kit (QIAGEN), quantified, stored, and used as a set.
Analysis of HIV-1 envelope glycoprotein variants.To examine the expression, processing, and gp120-gp41 association of the mutant HIV-1 envelope glycopro-teins, 9g of the pSVIIIenv plasmid was cotransfected with 1g of a plasmid expressing the HIV-1 Tat protein into 293T cells by using Lipofectamine reagent (Invitrogen) according to the manufacturer’s recommendations. Beginning at approximately 16 to 24 h after transfection, the cells were labeled with 200Ci each of [35
S]methionine and [35
S]cysteine in methionine- and cysteine-free me-dium for 16 to 24 h. The secreted gp120 glycoproteins were harvested in the culture medium after a brief centrifugation to remove cell debris. The labeled cells were washed with 5 ml phosphate-buffered saline (PBS) and stripped from the plates with 10 mM EDTA-PBS. The harvested cells were lysed in 1 ml lysis buffer (20 mM Tris-HCl, pH 7.4, 0.5 M NaCl, 0.5% Nonidet P-40, and 1⫻ protease inhibitor cocktail [Pharmacia]) for 30 min at 4°C. Cell lysates were collected after clearing the cell debris by centrifugation at 14,000 rpm in an Eppendorf microfuge for 30 min at 4°C. For immunoprecipitations, 400l of medium or cell lysate was incubated overnight at 4°C with 3l of pooled sera from HIV-1-infected individuals and 50l of protein A-Sepharose (10% in PBS) (Pharmacia) that had been preincubated with 5% bovine serum albumin in PBS. After three washes with 1 ml lysis buffer, the beads were boiled for 5 min in 1⫻
sodium dodecyl sulfate (SDS) sample buffer with 1.5%-mercaptoethanol. The protein samples were then analyzed on 8% SDS-polyacrylamide gels (Invitrogen) and visualized by autoradiography.
Reporter virus stocks and single-round infectivity assay.Recombinant HIV-1 encoding firefly luciferase and pseudotyped with the wild-type or mutant enve-lope glycoproteins was produced as previously described (21, 47). Briefly, 293T cells in 100-mm tissue culture dishes were cotransfected by the Lipofectamine reagent with 2g of the pSVIIIenv plasmid expressing the HIV-1 envelope glycoprotein variants, 2g of the pCMV⌬P1⌬envpA plasmid, and 6g of the pHIV-1Luc plasmid. The pCMV⌬P1⌬envpA plasmid encodes the packaging components (Gag/Pol proteins) and the Tat protein of HIV-1. The pHIV-1Luc plasmid encodes a packageable HIV-1 vector that is defective in all HIV-1 genes excepttatand that expresses the luciferase reporter gene. The viral stocks, which are capable of a single round of infection, were harvested 2 days later, aliquoted, and stored at⫺80°C. The infectivity of recombinant viruses was measured by incubation of the viruses with Cf2Th-CD4/CCR5 cells. Target cells (6⫻103
cells per well) were seeded into 96-well tissue culture isoplates (EG&G Wallac) and cultured for 16 to 24 h. Viral stocks and the tissue culture medium used for the dilution were prewarmed to 37°C, and serial dilutions of the viral stocks were made. After thorough removal of the media from the target cells, 100l of the diluted virus suspension was added to each well. After 48 h, viral infectivity was quantified by measuring the luciferase activity in the target cells with a luciferase detection kit (Pharmingen) and an automated luminometer (Microlumat Plus; EG&G Berthod). For each sample of diluted virus, multiple (four or six) wells of target cells were infected in parallel and the mean value of luciferase activity obtained. All three mutant envelope glycoproteins described above (D368R, R315G/L317S, and L520E) were defective in the ability to support virus entry by at least 4 orders of magnitude relative to the wild-type HIV-1YU2envelope glycoproteins (see Fig. 4).
To produce viral stocks with differentfMvalues, 293T cells were transfected as
[image:2.585.72.255.72.250.2]described above except that the pSVIIIenv plasmids expressing the wild-type HIV-1YU2envelope glycoproteins and one derivative mutant were mixed at a
FIG. 1. Theoretical models for the subunit stoichiometry of the
HIV-1 envelope glycoprotein trimer. S is defined as the number of
wild-type subunits within one HIV-1 envelope glycoprotein trimer that
is required for the trimer to support virus entry.
f
Mis defined as the
frequency of mutant monomers in the total pool of HIV-1 envelope
glycoproteins. The frequency of potentially functional trimers was
cal-culated using the theoretical assumptions described in Materials and
Methods. The frequency of potentially functional trimers in a viral
stock is predictive of the infectivity relative to that of the wild-type
viruses [RI(%)].
on November 8, 2019 by guest
http://jvi.asm.org/
given ratio, keeping the total amount of plasmid DNA at 2g. To minimize the potential for pipetting errors in creating viruses with very low or highfMvalues,
the pSVIIIenv plasmids were first diluted so that all pipetting could be per-formed in the 4- to 20-l range. All viral stocks associated with one series of differentfMvalues were prepared and subsequently tested as a set. Recombinant
viral production was measured with a standard reverse transcriptase (RT) assay, and all viral stocks had RT levels within 20% of each other (data not shown). The infectivities of recombinant viruses with different ratios (fMvalues) of wild-type
and mutant envelope glycoproteins were determined. In pilot experiments, we compared variation in infectivity of viral stocks with the wild-type glycoproteins by normalization either to the volume of input virus or to the RT activity. Due to higher variance of infectivity of different viral stocks normalized by RT activity (data not shown), normalization by viral stock volume was employed in all subsequent experiments. The infectivities of the viral stocks associated with a given series offMvalues were expressed as a percentage of the infectivity of the
wild-type virus to generate percentages of relative infectivity [RI(%)]. The rela-tionship betweenfM and RI(%) values was then compared with the curves
predicted by the candidate theoretical models of subunit stoichiometry. Quan-titative data fitting was conducted to fit the observed RI(%) data to the values expected by candidate models based on individual mutants and on aggregation of all five mutants.
Data fitting.The construction of theoretical models and their predictions were explained in “Theoretical model building” above. Four parallel measurements of RI(%) for viruses withfMvalues of 0.2, 0.4, 0.6, and 0.8 were collected for
individual envelope glycoprotein mutants, and the means of the four primary data points were generated. To analyze the model fitness for each individual mutant, the means of the RI(%) data were used for analysis due to the ease of data handling; the RI(%) values offM⫽0 andfM⫽1 were excluded from the
analysis because they were predetermined as 0% and 100%, respectively, thus contributing no variance to the assessment of fitness. To examine the deviation of the observed RI(%) data [RI(%)observed
] from the RI(%) values predicted by a given model [RI(%)expected], total modeling variance of a given mutant was calculated by⌺[RI(%)observed⫺RI(%)expected]2/RI(%)expectedforf
Mvalues of
0.2 to 0.8. For each mutant, the total modeling variances were separately gen-erated for models of S⫽1, S⫽2, or S⫽3; the candidate model with the smallest total modeling variance was considered to best describe the observed data.
Simultaneous estimation of S and T values.We use T to designate the number of HIV-1 envelope glycoprotein trimers required for virus entry. Two indepen-dent approaches indicated that each HIV-1 envelope glycoprotein trimer func-tions independently to mediate virus entry, i.e., T⫽1 (46, 47). We attempted to simultaneously estimate the T and S values by using the data generated from the loss-of-function mutants in this study without any preset assumptions of possible S and T values. To estimate the S and T values based on the aggregated data from all five envelope glycoprotein mutants, a mixed nonlinear regression model using all four primary RI(%) data points from each of five mutants was devel-oped, thus including 20 independent data points at each level offMfrom 0 to 1.0 (2).
Let RI(%)ij⫽{1⫺[fM3⫹3fM2(1⫺fM)(S⫺1)⫹3fM(1⫺fM)2(S⫺1)(S⫺
2)/2⫺3fM2(1⫺fM)(S⫺1)(S⫺2)/2]}T⫻100⫹eij⫹ui, wherei⫽1, 2, . . . 20
andj⫽1, 2, . . . 6. The S and T values are the fixed-effect parameter to be computed,uiis the random-effects parameter for theith set, andeijis the residual
error for theith set at thejthfM level. Theiparameter designated the 20
individual RI(%) data points at each level offM; thejparameter represented six
levels of thefMvalues. We assumed thatuiandeijwere independent and both
followed a normal distribution. This nonlinear regression model was solved for S and T by use of an SAS procedure, PROC NLMIXED (2). The Newton-Rhap-son method was used as an optimization technique in the computation (36a).
Complementation of HIV-1 envelope glycoprotein mutants for function in virus entry and cell-to-cell fusion.To test whether functional complementation between envelope glycoprotein mutants with defects in different functional mo-dalities can occur during viral entry, 1g each of two of the three plasmids expressing D368R, L317S/R315G, and L520E mutant envelope glycoproteins of HIV-1YU2gp160, in pairwise combinations, was cotransfected with 2g of the pCMV⌬P1⌬envpA plasmid and 6g of the pHIV-1Luc plasmid to make viral stocks, as described above. The infectivities of these heterotrimer viral stocks were measured in parallel with those of viruses with singular wild-type or mutant envelope glycoproteins made under the same conditions. The means and ranges of variation of luciferase signals from four wells of parallel infections were compared.
To test whether three different HIV-1 envelope glycoprotein mutants could complement each other for function in a three-way combination by forming heterotrimers with three different mutant subunits, we made luciferase reporter viruses by cotransfecting 0.66g each of the three plasmids expressing the D368R, L317S/R315G, and L520E mutant HIV-1YU2gp160s with 2g of the
pCMV⌬P1⌬envpA plasmid and 6g of the pHIV-1Luc plasmid. In one set of experiments, these viruses were compared with viruses carrying the wild-type or D368R, L317S/R315G, or L520E mutant envelope glycoproteins of HIV-1YU2 gp160 individually and those carrying the pairwise combinations of the three mutant glycoproteins, as described above.
To examine the potential of these mutant envelope glycoproteins to comple-ment each other in inducing cell-cell fusion or syncytium formation, 293T cells (the “effector cells”) in six-well plates were transiently transfected with 0.2g of the plasmid expressing HIV-1 Tat and 0.9g each of the plasmid DNAs ex-pressing two of the three mutant HIV-1YU2envelope glycoproteins by the stan-dard Lipofectamine procedure. For comparison, cells were transfected with the plasmids expressing wild-type and mutant HIV-1 gp160 glycoproteins individu-ally and with the pcDNA3.1 vector as a negative control. Approximately 3⫻103 Cf2Th-CD4/CCR5 cells were seeded in the 96-well plates to serve as the target cells. After 24 h of cocultivation of the effector cells (0.1% of cells transfected in six-well plates) and the target cells in six parallel wells, the level of syncytium formation was quantified by counting giant cells with five or more nuclei under a light microscope. The average numbers and ranges of variation of syncytia in the six parallel wells were compared.
RESULTS
HIV-1 envelope glycoprotein mutants.
Our strategy for
in-vestigating the subunit stoichiometry associated with HIV-1
entry involves the functional analysis of viruses containing
mix-tures of wild-type and defective envelope glycoproteins. Based
on existing information on functional motifs within the HIV-1
envelope glycoproteins, we designed mutant HIV-1
YU2enve-lope glycoproteins defective in one of the following functions:
CD4 binding, CCR5 binding, or fusion peptide activity.
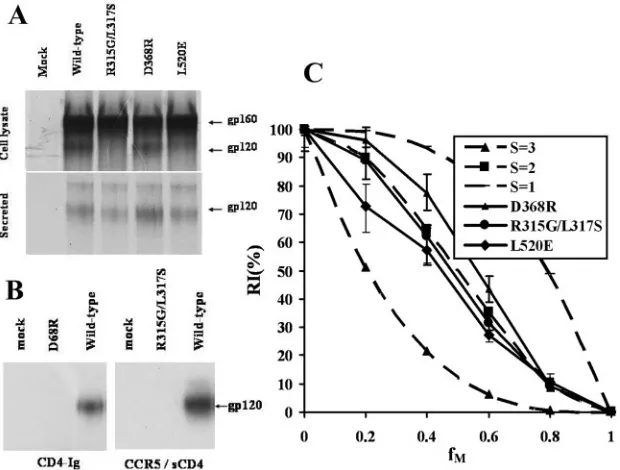
The D368R mutant is altered in aspartic acid 368, which
forms a key salt bridge with CD4, and is markedly defective in
CD4 binding (26, 42). The D368R HIV-1
YU2envelope
glyco-proteins were expressed and processed comparably to the
wild-type HIV-1
YU2envelope glycoproteins (Fig. 2A). Although
similar amounts of the D368R mutant and wild-type gp120
envelope glycoprotein were shed into the medium (Fig. 2A),
the precipitation of the shed gp120 by soluble
CD4-immuno-globulin (Ig) was undetectable for the D368R mutant (Fig. 2B,
left panel). The soluble CD4-Ig molecule contains the four
N-terminal domains of CD4 fused to the Fc domain of mouse
IgG1. These results are consistent with the poor ability of the
D368R mutant to interact with CD4.
The L317S mutant of the HIV-1
YU2envelope glycoproteins
has an alteration in the gp120 V3 loop that decreases the
efficiency of CCR5 binding (34). The L317S mutant retained
some ability to support HIV-1 entry when pseudotyped onto a
recombinant luciferase-expressing HIV-1 vector (data not
shown). To eliminate this residual function, a second residue
change (R315G) was introduced into the V3 loop; the resulting
mutant, R315G/L317S, was efficiently expressed but exhibited
decreases in proteolytic processing of the gp160 glycoprotein
precursor (Fig. 2A). Nonetheless, substantial amounts of
gp120 were present in the supernatants of cells expressing the
R315G/L317S envelope glycoproteins. The wild-type and
R315G/L317S gp120 glycoproteins in cell supernatants were
concentrated 10-fold with a Centriprep YM30 filter (Amicon).
The concentrated gp120 proteins were incubated with 20
g/ml
soluble CD4 and 3
⫻
10
6Cf2Th-CCR5 cells, which express
CCR5. After extensive washing, the cell-bound gp120 was
de-tected by immunoprecipitation. In contrast to the wild-type
HIV-1
YU2gp120 glycoprotein, the R315G/L317S mutant
on November 8, 2019 by guest
http://jvi.asm.org/
gp120 did not detectably bind CCR5 on the cell surface (Fig.
2B, right panel).
The L520E mutant contains a charged residue in the
nor-mally hydrophobic fusion peptide at the gp41 N terminus (17).
The L520E mutant envelope glycoproteins were efficiently
ex-pressed and processed and exhibited a level of gp120-gp41
association that was comparable to that of the wild-type
enve-lope glycoproteins (Fig. 2A).
Subunit stoichiometry of the HIV-1 envelope glycoproteins.
The infectivity of recombinant HIV-1 viruses containing
dif-ferent ratios (
f
Mvalues) of wild-type and mutant envelope
glycoproteins was determined by using Cf2Th-CD4/CCR5
tar-get cells. The relationship between RI(%) and
f
Mfor these
viruses is shown in Fig. 2C. For the three mutant envelope
glycoproteins, the observed curves were consistent with the
S
⫽
2 theoretical curve.
The subunit stoichiometry of the HIV-1 envelope
glycopro-teins was examined for another virus strain, HIV-1
HXBc2. The
HIV-1
HXBc2envelope glycoproteins differ substantially from
those of HIV-1
YU2; HIV-1
HXBc2is a well-characterized T-cell
line-adapted isolate, uses CXCR4 as a coreceptor, and is very
sensitive to neutralizing antibodies (16). The D368R mutant of
the HIV-1
HXBc2envelope glycoproteins has been shown to be
defective for CD4 binding (42); the D368R mutant was
effi-ciently expressed and processed compared to the wild-type
HIV-1
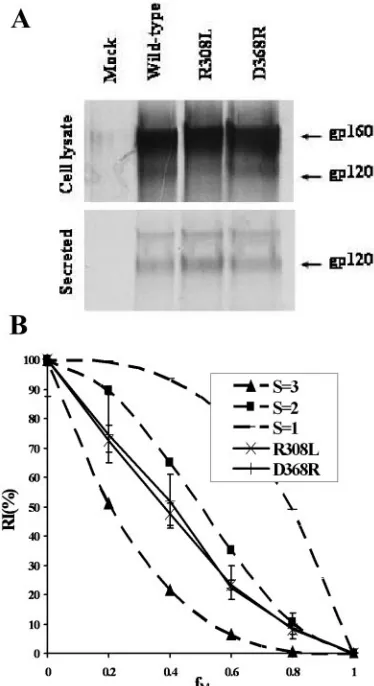
HXBc2envelope glycoproteins (Fig. 3A). We screened
several existing candidates for an HIV-1
HXBc2envelope
glyco-protein mutant that is defective for CXCR4 binding and is
suitable for our purpose (3). The R308L mutant was incapable
of supporting virus entry (data not shown) but was expressed
and processed relatively efficiently (Fig. 3A). Several attempts
to identify candidate HIV-1
HXBc2mutants that are defective
for fusion peptide function and suitable for our experimental
design were not successful, due to a low level of residual
function in supporting virus entry (F522Y and F522L),
abnor-mal protein processing (L520E), or significant shedding of the
gp120 subunits (F522Y/L523S) (data not shown).
[image:4.585.138.448.70.305.2]Viral stocks pseudotyped with the wild-type, D368R mutant,
or R308L mutant HIV-1
HXBc2envelope glycoproteins were
produced as described before. The infectivity of these viral
FIG. 2. HIV-1
YU2envelope glycoproteins use an S
⫽
2 subunit stoichiometry to support viral infection. (A) Expression and processing of the
HIV-1
YU2envelope glycoprotein variants. Transfected 293T cells expressing the wild-type and mutant envelope glycoproteins were radiolabeled
with [
35S]methionine and [
35S]cysteine for 24 h. The secreted glycoproteins were harvested from the culture medium, and the cell-associated
glycoproteins were harvested from lysed cells. The radiolabeled glycoproteins were precipitated with pooled sera from HIV-1-infected individuals
and analyzed on 8% SDS-polyacrylamide gels. The gp160 envelope glycoprotein precursors and the mature gp120 glycoproteins are indicated.
(B) The receptor-binding abilities of the wild-type and mutant gp120 glycoproteins were assessed as in our previously published studies (24, 28).
To test the glycoproteins’ ability to bind CD4 in solution, CD4-Ig was incubated with protein A-agarose and culture medium containing
35S-labeled
gp120 at room temperature for 3 h. After being washed with the cell lysis buffer, the precipitated glycoproteins were resolved on 8%
SDS-polyacrylamide gels (left panel). To measure CCR5 binding,
35S-labeled culture media were concentrated 10-fold by using a Centriprep YM30 filter
(Amicon) and then incubated with 3
⫻
10
6Cf2Th-CCR5 cells and 2
g of soluble CD4 at 37°C for 1 h. After removal of the unbound envelope
glycoproteins by three washes with PBS, the target cells were lysed in the cell lysis buffer. The cell-bound glycoproteins in the lysates were then
detected by immunoprecipitation with pooled sera from HIV-1-infected persons and resolved by SDS-polyacrylamide gel electrophoresis (right
panel). (C) Relative infectivities of viruses with mixtures of the wild-type and mutant HIV-1
YU2envelope glycoproteins. Recombinant
luciferase-expressing HIV-1 with various ratios (
f
M) of wild-type and mutant envelope glycoproteins (D368R, L317S/R315G, or L520E) was incubated with
Cf2Th-CD4/CCR5 cells. The luciferase activity in the target cells was measured, and the relative infectivity of a given viral stock was calculated
by normalization to the infectivity of the viral stock carrying only the wild-type HIV-1
YU2envelope glycoproteins. The means and the ranges of
variation from four or six parallel wells of infection are shown (solid lines). All experiments were performed at least twice with two independent
sets of viral stocks in one series of
f
Mvalues, and results from a single typical experiment are shown. Also shown are the curves expected for the
theoretical models of S
⫽
1, S
⫽
2, and S
⫽
3 (dashed lines).
on November 8, 2019 by guest
http://jvi.asm.org/
stocks was measured on Cf2Th-CD4/CXCR4 target cells, as
described above. The two mutant HIV-1
HXBc2gp160 envelope
glycoproteins exhibited decreases of at least 4 orders of
mag-nitude in the ability to support virus entry (data not shown).
Viral stocks with a series of
f
Mvalues were produced by
coex-pressing the wild-type and either the D368R or the R308L
mutant HIV-1
HXBc2envelope glycoproteins in individual sets.
The relationships between RI(%) and
f
Mobserved for the
D368R and R308L mutants were close to that of the S
⫽
2
model curve (Fig. 3B). Thus, the S
⫽
2 subunit stoichiometry
appears to be applicable to the envelope glycoproteins of two
phenotypically distinct HIV-1 isolates.
Quantitative model fitting to estimate S values based on
individual HIV-1 envelope glycoprotein mutants.
We applied
statistical methods to evaluate the fitness of the model
predic-tions for the relapredic-tionships between the
f
Mvalues and the
ob-served RI(%) values. In the theoretical models, the RI(%)
values for individual HIV-1 envelope glycoprotein mutants are
predicted by the formula (1
⫺
f
M)
3, 1
⫺
3
f
M2⫹
2
f
M3, or 1
⫺
f
M3for models of S
⫽
1, S
⫽
2, or S
⫽
3, respectively (see Materials
and Methods). Using these formulae, the expected RI(%)
val-ues [RI(%)
expected] for viral stocks with
f
M
values of 0.2, 0.4,
0.6, or 0.8 were generated (Table 1). For each HIV-1
enve-lope glycoprotein mutant, the model fitness was judged
based on the cumulative deviations of the observed RI(%)
[RI(%)
observed] data from the RI(%)
expectedvalues for the four
viral stocks with
f
Mvalues of 0.2, 0.4, 0.6, or 0.8. The viruses
with
f
Mvalues of 0 or 1.0, i.e., the viruses with pure mutant or
wild-type envelope glycoproteins, had RI(%) values of 0% or
100% by definition; thus, they did not contribute to the
vari-ance for the purpose of modeling. The total modeling varivari-ance
represents the cumulative deviation of the RI(%)
observedval-ues from the model predictions. The total modeling variance
for each envelope glycoprotein mutant was calculated by
⌺
[RI(%)
observed⫺
RI(%)
expected]
2/RI(%)
expectedfor the
f
M
values of 0.2, 0.4, 0.6, and 0.8. For all five mutant envelope
glycoproteins of HIV-1
YU2and HIV-1
HXBc2, the total
model-ing variances were the smallest for the S
⫽
2 model by very large
margins (Table 2).
Simultaneous estimation of S and T values by use of
aggre-gated data from all five HIV-1 envelope glycoprotein mutants.
Our previous work indicates that the minimal functional unit
of HIV-1 envelope glycoproteins in mediating virus entry is
composed of a single trimer, i.e., T
⫽
1. This conclusion was
supported by studying the target size of anti-HIV-1 Env
neu-tralizing antibodies (46) and by using dominant-negative
mu-tants of HIV-1 envelope glycoproteins (47). Furthermore, a
T
⫽
1 stoichiometry is observed with the envelope glycoproteins
of two other viruses, Friend murine leukemia virus and avian
sarcoma/leukosis virus type A. Thus, a significant amount of
evidence supports the T
⫽
1 model for at least some
retrovi-ruses. We also wished to estimate the S and T values by using
a parametrical statistical method to analyze the data generated
from the above-described study of the five envelope
glycopro-tein mutants. Unlike the above-described analysis of individual
HIV-1 envelope glycoprotein mutants, the multivariate
analy-sis was conducted without any assumption about the S and T
values. Because of the limited statistical power when only the
five means were used for modeling, four primary RI(%) data
points from each of the five mutants with
f
Mvalues of 0 to 1.0,
i.e., 20 independent RI% values at each of the six levels of
f
M,
were included to enhance the modeling power. According to
the general relationships among RI(%),
f
M, and S (see
Mate-rials and Methods), let RI(%)
ij⫽
{1
⫺
[
f
M3
⫹
3
f
M
2
(1
⫺
f
M
)(S
⫺
1)
⫹
3
f
M(1
⫺
f
M)
2(S
⫺
1)(S
⫺
2)/2
⫺
3
f
M2(1
⫺
f
M)(S
⫺
1)(S
⫺
2)/2]}
T⫻
100
⫹
e
ij
⫹
u
i, where
i
⫽
1, 2, . . . 20 and
j
⫽
1,
TABLE 1. RI(%)
expectedvalues for models with different S values
Model
RI(%)expectedvalue atf
Mvalue of:
0.2 0.4 0.6 0.8
S
⫽
1
99.2
93.6
78.4
48.8
S
⫽
2
89.6
64.8
35.2
10.4
[image:5.585.68.255.73.416.2]S
⫽
3
51.2
21.6
6.4
0.8
FIG. 3. S
⫽
2 subunit stoichiometry of the HIV-1
HXBc2envelope
glycoproteins in virus entry. (A) Expression and processing of the
HIV-1
HXBc2envelope glycoprotein variants. The wild-type and mutant
HIV-1
HXBc2envelope glycoproteins were detected in the culture media
and cell lysates as described in the legend for Fig. 2A. (B) Relative
infectivities of viruses with mixtures of wild-type and mutant HIV-1
HXBc2envelope glycoproteins. The relative infectivity of recombinant
lucif-erase-expressing HIV-1 with various ratios (
f
M) of wild-type and
mu-tant HIV-1
HXBc2glycoproteins (R308L or D368R) was measured using
Cf2Th-CD4/CXCR4 cells and reported as described in the legend for
Fig. 2C.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:5.585.301.542.662.727.2]2, . . . 5. A mixed nonlinear regression model was developed to
solve for the S and T values by use of an SAS procedure,
PROC NLMIXED (2). When all 20 primary RI(%) data
points at all six values of
f
Mwere included, the model
con-verged nicely on an estimated S value of 1.97 (95% confidence
interval, 1.70 to 2.25) and an estimated T value of 0.78 (95%
confidence interval, 0.54 to 1.01). This estimation of T is
con-sistent with our previous studies (46, 47), providing further
support for the conclusion that individual HIV-1 envelope
glycoprotein trimers mediate virus entry independently. The
estimation of S is consistent with that derived by model fitting
of the data obtained from the studies of individual envelope
glycoprotein mutants. Thus, the results support a model in
which two wild-type or functional subunits within an HIV-1
envelope glycoprotein trimer are required for the trimer to
mediate virus entry.
Absence of complementation between mutants with defects
in different functional modalities.
It has been reported that
envelope glycoprotein mutants that are defective in different
functional modalities could complement cell-cell fusion activity
when coexpressed in the same cells by use of vaccinia virus
vectors (36). In this context, two subunits of the trimer cannot
possess wild-type function to directly fulfill the S
⫽
2 subunit
stoichiometry. For example, if a CD4-binding-defective mutant
is coexpressed with a CCR5-binding-defective mutant, the two
kinds of homotrimers will be nonfunctional, and two species of
heterotrimers could be formed; one kind of heterotrimer has
one wild-type CD4-binding motif and two wild-type
CCR5-binding motifs, and the other kind of heterotrimer has two
wild-type CD4-binding motifs but one CCR5-binding motif.
Thus, neither of the two species of heterotrimer would be
comprised of two subunits functional in each of the two
mo-dalities, i.e., CD4 binding and CCR5 binding. To fulfill the
requirement of the S
⫽
2 subunit stoichiometry, cross talk
among subunits of the trimer would be required.
To test whether complementation between envelope
glyco-protein mutants with defects in different functional modalities
can occur during viral entry, the D368R, R315G/L317S, and
L520E mutant HIV
YU2envelope glycoproteins were
coex-pressed pairwise at a 1:1 ratio to produce viral stocks
contain-ing heterotrimers. The infectivities of these
heterotrimer-con-taining viral stocks were compared with those of viruses with
homotrimeric wild-type or mutant envelope glycoproteins. The
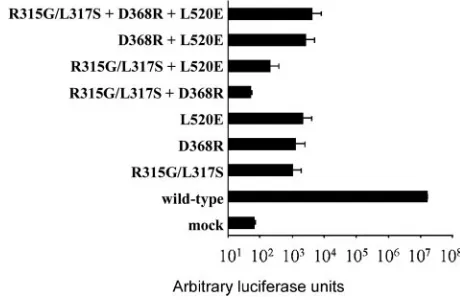
three viruses with homotrimeric mutant envelope
glycopro-teins exhibited infectivities that were approximately 4 logs
lower than that of the wild-type virus (Fig. 4). Viruses with the
three pairwise combinations of mutant envelope glycoproteins
did not demonstrate infectivities beyond those observed for
viruses with the individual mutant envelope glycoproteins (Fig.
4). Similarly, no functional complementation during virus entry
was observed with the D368R and R308L mutants of the
HIV-1
HXBc2envelope glycoproteins (data not shown). Thus, these
envelope glycoprotein mutants did not appreciably
comple-ment one another to support virus entry.
[image:6.585.44.283.88.187.2]When the three mutants (D368R, R315G/L317S, and
L520E) of the HIV-1
YU2envelope glycoproteins are all
coex-pressed in equal quantities in cells, a unique kind of
hetero-trimer that contains one subunit consisting of each of these
three mutant glycoproteins could be formed. Theoretically, this
specific heterotrimer could comprise up to 22% of the total
envelope glycoprotein trimers (3/3
⫻
2/3
⫻
1/3, i.e., 6/27, or
22%). This type of heterotrimer would theoretically retain two
subunits that can negotiate each of the three functions of the
HIV-1 envelope glycoproteins. Thus, there is a remote
possi-bility that these heterotrimers will satisfy the requirement of
the S
⫽
2 subunit stoichiometry. Hypothetically, if the
hetero-trimers with three subunits defective in three different
func-tions are functional, such a viral stock should exhibit a level of
infectivity close to 22% of that of the wild-type viruses and
more than 3 orders of magnitude over those of the viruses with
individual mutant envelope glycoproteins. To explore this, we
made luciferase reporter viruses by cotransfecting three
plas-mids expressing the D368R, L317S/R315G, and L520E mutant
HIV-1
YU2envelope glycoproteins in equal quantities. The
in-fectivity of such viral stocks was not significantly greater than
FIG. 4. Lack of genetic complementation between envelope
glyco-protein mutants with defects in different functional modalities. The
D368R, R315G/L317S, and L520E mutant HIV-1
YU2envelope
glyco-proteins are defective in CD4 binding, CCR5 binding, and membrane
fusion, respectively. Equal amounts of plasmids encoding the indicated
mutant HIV-1
YU2envelope glycoproteins were cotransfected in 293T
[image:6.585.303.533.466.617.2]cells to produce luciferase reporter viruses. The infectivity of the
re-sulting viral stocks was measured on Cf2Th-CD4/CCR5 target cells.
The infectivities of viruses with single wild-type or mutant envelope
glycoproteins were measured in parallel. The means and the ranges of
variation in luciferase counts from four parallel wells of infection are
shown on a logarithmic scale.
TABLE 2. Analysis of variance of RI(%)
observedvalues
afrom
RI(%)
expectedvalues
HIV-1
strain Mutant
RI(%)observedvalue atf
M
value of:
Total modeling varianceb
0.2 0.4 0.6 0.8 S⫽1 S⫽2 S⫽3
YU2
D368R
96.1 77.6 43.4
9.3
50.4
5.0
488.7
R315G/L317S
89.1 62.2 31.5
9.2
71.8
0.6
290.9
L520E
70.5 55.6 26.4 10.1
88.8
7.6
232.3
HXBc2 D368R
74.4 51.2 22.4
8.7
98.0
10.2
169.7
R308L
72.4 47.8 23.2
8.4 102.2
12.3
155.8
aThe mean RI(%)observedvalues were calculated from four independent wells of infections, as described in Materials and Methods. The RI(%)expectedvalues are given in Table 1.
bFor each HIV-1 envelope glycoprotein mutant, the total modeling
vari-ance was calculated using the formula⌺关RI(%)observed⫺RI(%)expected兴2/ RI(%)expectedfor the data points associated withf
Mvalues of 0.2, 0.4, 0.6, and
0.8. The lowest modeling variance for each mutant is highlighted in bold.
on November 8, 2019 by guest
http://jvi.asm.org/
that of the viruses with single mutant envelope glycoproteins
(Fig. 4). Thus, the existence of two functional motifs in each of
the subunits of an HIV-1 envelope glycoprotein trimer was
insufficient to allow viral entry, suggesting the lack of efficient
intersubunit communication within a trimer.
We also examined the potential of these mutant envelope
glycoproteins to complement each other to induce cell-cell
fusion or syncytium formation. In these experiments, 293T
cells were transiently transfected with pairwise combinations of
plasmid DNAs expressing two of the three mutant HIV-1
YU2envelope glycoproteins to create effector cells. These effector
cells were cocultivated with Cf2Th-CD4/CCR5 target cells in
96-well plates, and syncytia were counted 24 h later. 293T cells
expressing single wild-type or mutant envelope glycoproteins
were used as controls. In all of the cells expressing pairwise
combinations of mutant HIV-1 envelope glycoproteins,
syncy-tium formation was not appreciably higher than in the cells
expressing the individual mutants (data not shown). Similarly,
cotransfecting three different mutant envelope glycoproteins
did not result in significant syncytium formation (data not
shown). Thus, functional complementation among the mutant
envelope glycoproteins in mediating cell-cell fusion was not
detected in our experimental setting.
DISCUSSION
Here we provide evidence that at least two of the subunits of
an HIV-1 envelope glycoprotein trimer must function to result
in detectable virus entry. We conclude that S
⫽
2 from fitting
the observed entry data of viruses with mixtures of wild-type
and mutant HIV-1 envelope glycoproteins to the curves
pre-dicted by theoretical models. The theoretical models for this
study were generated using some basic assumptions about
HIV-1 envelope glycoprotein and its function. These
assump-tions underlie the construction of these theoretical models and
allow us to reach conclusions using this approach. These
as-sumptions are as follows: (i) the wild-type and mutant
enve-lope glycoproteins are expressed in the same cells after
co-transfection, the relative levels of the paired envelope
glycoproteins are proportional to the ratios of the transfected
plasmids, and the monomers of the paired Env proteins form
trimers by random association; (ii) each HIV-1 virion particle
contains no more than one infectious unit, or, in other words,
the infectious units in HIV-1 viral stocks function
indepen-dently; and (iii) all heterotrimers containing mutant subunits
function in an all-or-none fashion. Detailed justifications for
the first two assumptions, i.e., random mixing and
indepen-dence of infectious units, have been published previously (47)
and will not be repeated here. Violation of the third
assump-tion would imply that some heterotrimers exhibit partial
func-tion. This would result in our underestimation of S. Thus, we
considered the possibility that S is actually 3 and that some
heterotrimers retain partial function. For the observed data to
be explained by the S
⫽
3 theoretical model, the heterotrimers
containing mutant subunits would need to exhibit significant
levels of partial function. This is particularly the case for the
data points where the
f
Mvalues are high and the observed
RI(%) values are readily measurable. For the five viral stocks
with
f
Mvalues of 0.8, the observed RI(%) values were 9.3, 9.2,
and 10.4 for D368R, L317S/R315G, and L520E of HIV-1
YU2,
respectively, and 8.7 and 8.3 for D368R and R308L of
HIV-1
HXBc2, respectively; these values are at least 10-fold higher
than the RI(%) value of 0.8% predicted by the S
⫽
3 model.
Thus, very high levels of heterotrimer function, approaching
that predicted by the S
⫽
2 model, are required to explain the
data. Finally, a fourth assumption, i.e., that T
⫽
1, was used to
fit the data from the study of individual HIV-1 envelope
glyco-protein mutants. However, we also analyzed the data derived
from the study of all five mutants by a mixed nonlinear
regres-sion model without making any assumptions about T. The
results from the latter analysis support our conclusion that S
equals 2 and are consistent with a T value of 1.
The S
⫽
2 subunit stoichiometry of envelope glycoprotein
trimers in HIV-1 entry specifies the minimum requirement of
subunits in support of trimer function. During the normal
HIV-1 entry process, all three subunits in an envelope
glyco-protein trimer may uniformly contribute to function. We
ob-served no evidence of functional complementation between
HIV-1 envelope glycoprotein mutants defective in different
entry-related functions. Thus, minimal cross-communication
occurs among the trimeric subunits with respect to
transmis-sion of receptor-induced conformational changes triggering
membrane fusion.
Several different types of envelope glycoprotein inhibitors
have been developed as potential therapeutics against HIV-1
infection (27, 35, 44, 49). Neutralizing antibodies block HIV-1
envelope glycoprotein spikes using a 1:1 stoichiometry (46).
Such a neutralizing stoichiometry is not necessarily in conflict with
the S
⫽
2 subunit stoichiometry reported here, given that
neutral-izing antibodies can function by steric hindrance through their
relative bulkiness (33) or by interference with overall structural
changes in the trimer complex (31). It is of interest to define the
stoichiometry of peptide blockers, e.g., T20, that interfere with the
conformational changes in the trimer required for overall
enve-lope glycoprotein function (7). As an inference from the S
⫽
2
subunit stoichiometry, molecular blockers that compete with CD4
or CCR5 for binding to individual gp120 subunits may need to
bind two or more subunits of the targeted trimer to effectively
inhibit its function. Understanding the requirements for
inhibi-tion of HIV-1 entry may assist the development of effective
ther-apies or prophylactic approaches.
ACKNOWLEDGMENTS
We thank Yvette McLaughlin and Sheri Farnum for manuscript
preparation.
This work was supported by grants from the NIH (AI24755,
AI31782, AI46725, and a Center for AIDS Research award), the
In-ternational AIDS Vaccine Initiative, the Bristol-Myers Squibb
Foun-dation, and the William A. Haseltine Foundation for the Arts and
Sciences and a gift from William F. McCarty-Cooper.
REFERENCES
1.Alkhatib, G., C. Combadiere, C. C. Broder, Y. Feng, P. E. Kennedy, P. M. Murphy, and E. A. Berger. 1996. CC CKR5: a RANTES, MIP-1alpha, MIP-1beta receptor as a fusion cofactor for macrophage-tropic HIV-1. Sci-ence272:1955–1958.
2.Bard, J.1974. Nonlinear parameter estimation. Academic Press, New York, N.Y.
3.Basmaciogullari, S., G. J. Babcock, D. Van Ryk, W. Wojtowicz, and J. Sodroski.2002. Identification of conserved and variable structures in the human immunodeficiency virus gp120 glycoprotein of importance for CXCR4 binding. J. Virol.76:10791–10800.
4.Bleul, C. C., M. Farzan, H. Choe, C. Parolin, I. Clark-Lewis, J. Sodroski, and T. A. Springer.1996. The lymphocyte chemoattractant SDF-1 is a ligand for LESTR/fusin and blocks HIV-1 entry. Nature382:829–833.
on November 8, 2019 by guest
http://jvi.asm.org/
5.Boulay, F., R. W. Doms, R. G. Webster, and A. Helenius.1988. Posttransla-tional oligomerization and cooperative acid activation of fixed influenza hemagglutinin trimers. J. Cell Biol.106:629–639.
6.Chan, D. C., D. Fass, J. M. Berger, and P. S. Kim.1997. Core structure of gp41 from the HIV envelope glycoprotein. Cell89:263–273.
7.Chan, D. C., and P. S. Kim.1998. HIV entry and its inhibition. Cell93:681–684. 8.Choe, H., M. Farzan, Y. Sun, N. Sullivan, B. Rollins, P. D. Ponath, L. Wu, C. R. Mackay, G. LaRosa, W. Newman, N. Gerard, C. Gerard, and J. Sodroski.1996. The beta-chemokine receptors CCR3 and CCR5 facilitate infection by primary HIV-1 isolates. Cell85:1135–1148.
9.Dalgleish, A. G., P. C. Beverley, P. R. Clapham, D. H. Crawford, M. F. Greaves, and R. A. Weiss. 1984. The CD4 (T4) antigen is an essential component of the receptor for the AIDS retrovirus. Nature312:763–767. 10.Deng, H., R. Liu, W. Ellmeier, S. Choe, D. Unutmaz, M. Burkhart, P. Di
Marzio, S. Marmon, R. E. Sutton, C. M. Hill, C. B. Davis, S. C. Peiper, T. J. Schall, D. R. Littman, and N. R. Landau.1996. Identification of a major co-receptor for primary isolates of HIV-1. Nature381:661–666.
11.Doranz, B. J., J. Rucker, Y. Yi, R. J. Smyth, M. Samson, S. C. Peiper, M. Parmentier, R. G. Collman, and R. W. Doms.1996. A dual-tropic primary HIV-1 isolate that uses fusin and the beta-chemokine receptors CKR-5, CKR-3, and CKR-2b as fusion cofactors. Cell85:1149–1158.
12.Earl, P. L., B. Moss, and R. W. Doms. 1991. Folding, interaction with GRP78-BiP, assembly, and transport of the human immunodeficiency virus type 1 envelope protein. J. Virol.65:2047–2055.
13.Earp, L. J., S. E. Delos, H. E. Park, and J. M. White.2005. The many mechanisms of viral membrane fusion proteins. Curr. Top. Microbiol. Im-munol.285:25–66.
14.Eckert, D. M., and P. S. Kim.2001. Mechanisms of viral membrane fusion and its inhibition. Annu. Rev. Biochem.70:777–810.
15.Feng, Y., C. C. Broder, P. E. Kennedy, and E. A. Berger.1996. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science272:872–877.
16.Fisher, A. G., E. Collalti, L. Ratner, R. C. Gallo, and F. Wong-Staal.1985. A molecular clone of HTLV-III with biological activity. Nature316:262–265. 17.Freed, E. O., E. L. Delwart, G. L. Buchschacher, Jr., and A. T. Panganiban.
1992. A mutation in the human immunodeficiency virus type 1 transmem-brane glycoprotein gp41 dominantly interferes with fusion and infectivity. Proc. Natl. Acad. Sci. USA89:70–74.
18.Freed, E. O., D. J. Myers, and R. Risser.1990. Characterization of the fusion domain of the human immunodeficiency virus type 1 envelope glycoprotein gp41. Proc. Natl. Acad. Sci. USA87:4650–4654.
19.Freed, E. O., D. J. Myers, and R. Risser.1989. Mutational analysis of the cleavage sequence of the human immunodeficiency virus type 1 envelope glycoprotein precursor gp160. J. Virol.63:4670–4675.
20.He, Y., R. Vassell, M. Zaitseva, N. Nguyen, Z. Yang, Y. Weng, and C. D. Weiss.2003. Peptides trap the human immunodeficiency virus type 1 enve-lope glycoprotein fusion intermediate at two sites. J. Virol.77:1666–1671. 21.Hofmann, W., D. Schubert, J. LaBonte, L. Munson, S. Gibson, J. Scammell,
P. Ferrigno, and J. Sodroski.1999. Species-specific, postentry barriers to primate immunodeficiency virus infection. J. Virol.73:10020–10028. 22.Kim, M., B. Chen, R. E. Hussey, Y. Chishti, D. Montefiori, J. A. Hoxie, O.
Byron, G. Campbell, S. C. Harrison, and E. L. Reinherz.2001. The stoichi-ometry of trimeric SIV glycoprotein interaction with CD4 differs from that of anti-envelope antibody Fab fragments. J. Biol. Chem.276:42667–42676. 23.Klatzmann, D., E. Champagne, S. Chamaret, J. Gruest, D. Guetard, T.
Her-cend, J. C. Gluckman, and L. Montagnier.1984. T-lymphocyte T4 molecule behaves as the receptor for human retrovirus LAV. Nature312:767–768. 24.Kolchinsky, P., E. Kiprilov, P. Bartley, R. Rubinstein, and J. Sodroski.2001.
Loss of a single N-linked glycan allows CD4-independent human immuno-deficiency virus type 1 infection by altering the position of the gp120 V1/V2 variable loops. J. Virol.75:3435–3443.
25.Korber, B. F., C. Kuiken, B. Foley, S. Pillai, and J. Sodroski.1998. Num-bering positions in HIV relative to HXBc2, p. II-A-54–II-A-69.InB. K. Korber, C. Foley, F. Hahn, B. McCutchan, F. Mellor, and J. Sodroski (ed.), Human retroviruses and AIDS. Los Alamos National Laboratories, Los Alamos, N.Mex.
26.Kwong, P. D., R. Wyatt, J. Robinson, R. W. Sweet, J. Sodroski, and W. A. Hendrickson.1998. Structure of an HIV gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody. Nature
393:648–659.
27.Lin, P. F., W. Blair, T. Wang, T. Spicer, Q. Guo, N. Zhou, Y. F. Gong, H. G. Wang, R. Rose, G. Yamanaka, B. Robinson, C. B. Li, R. Fridell, C. Deminie, G. Demers, Z. Yang, L. Zadjura, N. Meanwell, and R. Colonno.2003. A small molecule HIV-1 inhibitor that targets the HIV-1 envelope and inhibits CD4 receptor binding. Proc. Natl. Acad. Sci. USA100:11013–11018. 28.Mirzabekov, T., H. Kontos, M. Farzan, W. Marasco, and J. Sodroski.2000.
Paramagnetic proteoliposomes containing a pure, native, and oriented sev-en-transmembrane segment protein, CCR5. Nat. Biotechnol.18:649–654. 29.Nara, P. L., and P. J. Fischinger.1988. Quantitative infectivity assay for
HIV-1 and -2. Nature332:469–470.
30.Oberlin, E., A. Amara, F. Bachelerie, C. Bessia, J. L. Virelizier, F. Arenzana-Seisdedos, O. Schwartz, J. M. Heard, I. Clark-Lewis, D. F. Legler, M. Loetscher, M. Baggiolini, and B. Moser.1996. The CXC chemokine SDF-1 is the ligand for LESTR/fusin and prevents infection by T-cell-line-adapted HIV-1. Nature382:833–835.
31.Parren, P. W., and D. R. Burton.2001. The antiviral activity of antibodies in vitro and in vivo. Adv. Immunol.77:195–262.
32.Piatak, M., Jr., M. S. Saag, L. C. Yang, S. J. Clark, J. C. Kappes, K. C. Luk, B. H. Hahn, G. M. Shaw, and J. D. Lifson.1993. High levels of HIV-1 in plasma during all stages of infection determined by competitive PCR. Sci-ence259:1749–1754.
33.Ren, X., J. Sodroski, and X. Yang.2005. An unrelated monoclonal antibody neutralizes human immunodeficiency virus type 1 by binding to an artificial epitope engineered in a functionally neutral region of the viral envelope glycoproteins. J. Virol.79:5616–5624.
34.Rizzuto, C. D., R. Wyatt, N. Hernandez-Ramos, Y. Sun, P. D. Kwong, W. A. Hendrickson, and J. Sodroski.1998. A conserved HIV gp120 glycoprotein structure involved in chemokine receptor binding. Science280:1949–1953. 35.Root, M. J., M. S. Kay, and P. S. Kim.2001. Protein design of an HIV-1 entry
inhibitor. Science291:884–888.
36.Salzwedel, K., and E. A. Berger.2000. Cooperative subunit interactions within the oligomeric envelope glycoprotein of HIV-1: functional comple-mentation of specific defects in gp120 and gp41. Proc. Natl. Acad. Sci. USA
97:12794–12799.
36a.SAS Institute.1999. SAS/STAT user’s guide, version 8, vol. 2. SAS Institute, Cary, N.C.
37.Si, Z., N. Madani, J. M. Cox, J. J. Chruma, J. C. Klein, A. Schon, N. Phan, L. Wang, A. C. Biorn, S. Cocklin, I. Chaiken, E. Freire, A. B. Smith III, and J. G. Sodroski.2004. Small-molecule inhibitors of HIV-1 entry block recep-tor-induced conformational changes in the viral envelope glycoproteins. Proc. Natl. Acad. Sci. USA101:5036–5041.
38.Sklyanskaya, E. I., M. Shie, Y. S. Komarov, S. S. Yamnikova, and N. V. Kaverin.1988. Formation of mixed hemagglutinin trimers in the course of double infection with influenza viruses belonging to different subtypes. Virus Res.10:153–165.
39.Stein, B. S., and E. G. Engleman.1990. Intracellular processing of the gp160 HIV-1 envelope precursor. Endoproteolytic cleavage occurs in a cis or me-dial compartment of the Golgi complex. J. Biol. Chem.265:2640–2649. 40.Sullivan, N., Y. Sun, J. Li, W. Hofmann, and J. Sodroski.1995. Replicative
function and neutralization sensitivity of envelope glycoproteins from pri-mary and T-cell line-passaged human immunodeficiency virus type 1 isolates. J. Virol.69:4413–4422.
41.Tan, K., J. Liu, J. Wang, S. Shen, and M. Lu.1997. Atomic structure of a thermostable subdomain of HIV-1 gp41. Proc. Natl. Acad. Sci. USA94:
12303–12308.
42.Thali, M., U. Olshevsky, C. Furman, D. Gabuzda, J. Li, and J. Sodroski.
1991. Effects of changes in gp120-CD4 binding affinity on human immuno-deficiency virus type 1 envelope glycoprotein function and soluble CD4 sensitivity. J. Virol.65:5007–5012.
43.Weissenhorn, W., A. Dessen, S. C. Harrison, J. J. Skehel, and D. C. Wiley.1997. Atomic structure of the ectodomain from HIV-1 gp41. Nature387:426–430. 44.Wild, C. T., D. C. Shugars, T. K. Greenwell, C. B. McDanal, and T. J.
Matthews.1994. Peptides corresponding to a predictive alpha-helical do-main of human immunodeficiency virus type 1 gp41 are potent inhibitors of virus infection. Proc. Natl. Acad. Sci. USA91:9770–9774.
45.Wyatt, R., and J. Sodroski.1998. The HIV-1 envelope glycoproteins: fuso-gens, antifuso-gens, and immunogens. Science280:1884–1888.
46.Yang, X., S. Kurteva, S. Lee, and J. Sodroski.2005. Stoichiometry of anti-body neutralization of human immunodeficiency virus type 1. J. Virol.79:
3500–3508.
47.Yang, X., S. Kurteva, X. Ren, S. Lee, and J. Sodroski.2005. Stoichiometry of envelope glycoprotein trimers in the entry of human immunodeficiency virus type 1. J. Virol.79:12132–12147.
48.Zhang, C. W., Y. Chishti, R. E. Hussey, and E. L. Reinherz.2001. Expression, purification, and characterization of recombinant HIV gp140. The gp41 ectodomain of HIV or simian immunodeficiency virus is sufficient to main-tain the retroviral envelope glycoprotein as a trimer. J. Biol. Chem.276:
39577–39585.
49.Zhao, Q., L. Ma, S. Jiang, H. Lu, S. Liu, Y. He, N. Strick, N. Neamati, and A. K. Debnath.2005. Identification of N-phenyl-N⬘ -(2,2,6,6-tetramethyl-pi-peridin-4-yl)-oxalamides as a new class of HIV-1 entry inhibitors that pre-vent gp120 binding to CD4. Virology339:213–225.
on November 8, 2019 by guest
http://jvi.asm.org/