JOURNAL OFVIROLOGY, Sept. 1986, p. 605-618 Vol.59, No. 3 0022-538X/86/090605-14$02.00/0
Copyright© 1986, AmericanSocietyforMicrobiology
Functional
Domains
within
the
a
Sequence
Involved in the
Cleavage-Packaging of Herpes Simplex Virus DNA
LOUIS P. DEISS, JOANY CHOU, AND NIZA FRENKEL*
DepartmentofMolecular Genetics andCellBiology, The University of Chicago, Chicago, Illinois60637 Received24February 1986/Accepted 16May 1986
Newlyreplicated herpes simplexvirus(HSV)DNAconsists of head-to-tailconcatemerswhicharecleavedto generate unit-length genomes bounded by the terminally reiterated a sequence. Constructed defective HSV vectors(amplicons)containing aviral DNAreplicationoriginand the asequencearesimilarly replicatedinto largeconcatemers whicharecleavedat asequencespunctuatingthejunctionsbetweenadjacent repeat units, concurrentwith thepackaging of viralDNAintonucleocapsids.Inthepresentstudywetested theabilityof seed amplicons containing specific deletions in the a sequence to become cleaved and packaged and hence be propagated in virus stocks. These studies revealed that two separate signals, located within the Ub and Uc elements ofthe a sequence, were essential foramplicon propagation. No derivative defective genomeswere recovered from seedconstructs which lackedtheUc signal.Incontrast,propagation ofseed constructslacking the Ubsignal resultedin theselectionof defective genomeswithnoveljunctions, containing specific insertions of a sequences derived from the helper virus DNA. Comparison of published sequences of concatemeric junctions of several herpesviruses supported a uniform mechanism for the cleavage-packaging process, involving the measurement from two highly conserved blocks of sequences (pac-l and pac-2) which were homologous tothe required Uc and Ub sequences. These resultsform the basis for general models for the mechanism ofcleavage-packaging of herpesvirusDNA.
The 150-kilobase (kb) DNA genomes of standard herpes
simplex viruses1and 2(HSV-1andHSV-2)arecomposed of two components, L and S, each consisting of unique se-quencesbracketedby invertedrepeats,aband b'a' flanking UL, andacand c'a' flankingUs. The L and S components invert relative to each other, yielding four genomic isomers
(reviewed in reference 25). The L-S junctions and the L termini of all four isomers contain a variable number of tandemly reiterated a sequences. In contrast, eachoftheS termini contains only a single copy ofthe a sequence (15,
38). Thestandard virus genome can thus berepresented by anb-UL-b'a'mc'-Us-ca, with n and m varying from 1 to greaterthan 10.
The linear viral DNA molecules circularize soon after
infection (22), and newly replicated viral DNA molecules
consist of large head-to-tail concatemers,arisingmost prob-ably by rolling-circle replication (2, 3, 12). The large concatemers are subsequently cleaved within a sequences situatedatthejunctions between adjacentviral genomes (6,
21, 25). Studies with temperature-sensitive mutants
sug-gestedthat thecleavageprocess maybe linked topackaging
(13, 14, 23; S. Bachenheimer, personal communication). Defective virus genomes present inseriallypassaged virus stocks are typically composed of head-to-tail repeat units,
eachcontainingaDNAreplication originand the a sequence
(reviewed in reference 8). In the presence of helper virus,
multimeric defective genomes can also be generated from seed plasmids (amplicons) containing a DNA replication
origin and the a sequence, which is required for con-catemericcleavageandpackaging(7, 28, 29, 31, 32, 36). Any
pairofa sequencesalong the concatemeric molecule could become cleaved, resulting in the packaging of monomers,
dimers, and higher multimeric forms of defective genomes into nuclear capsids. However, only full-length
(approxi-* Correspondingauthor.
mately 150kb) defective genomes were found to be present in cytoplasmic virions (37). Like the L-Sjunctions in stan-dard viral DNA, thejunctions separatingthe repeatunits of defective genomes contain a variable number oftandemly
reiterated a sequences, which are cleaved to generate a variable numberof a sequences at one terminus and
predom-inantly asingle a sequence atthe other terminus(7). TheasequencesofHSV-1 and HSV-2strainsrangeinsize
from 250 to 500 basepairs (bp)andcontainbothunique(U) anddirectly repeated (DR)elements(6,18, 19, 35). The size differences predominantly reflect variability in the copy number of the DR elements. For example, the 501-bp a sequenceofHSV-1 (F) is represented by
DR1-Ub-(DR2)19-(DR4)3-Uc-DR1, whereas the 264-bp a sequence of the
Justinstrain containsonlytwocopiesof DR2, asinglecopy
of DR4, and two copies of another reiteration designated
DR3.5 (18, 19). In adouble a (baac)junction the tandemly reiterated a sequences share the intervening DR1 (20 bp) element (6, 19, 21). Furthermore, in the F strain, the L
terminus (ba) contained 18bp plusone 3' overhang base of the DR1 sequence, whereas the S terminus (ca) carried the
remaining 1 1/2 bp. These findings led Mocarski and Roiz-man(21) to propose that the double ajunctions formedthe substrate for cleavage. In this regard, the cleavage ofviral DNA concatemers represented a puzzle inasmuch as,
al-though the number of a sequences at the junctions is variable, the predominantjunctions in both standard and defective viral genomes containa single a sequence (7, 15, 38). We haverecently shown that thejunctions containinga singleasequencecanindeedserve asthe targetfor
cleavage-packaging. Furthermore, thecleavage process itself appears to be accompanied by the amplification ofan a sequence, inasmuch as molecules carrying junctions of the structure xayarecleaved togenerate xaand ya termini (7).
The presentinvestigationwasdesigned to further mapthe signals within the a sequence which are involved in the 605
on November 10, 2019 by guest
http://jvi.asm.org/
606 DEISS ET AL. A.
E PE Ba
)oril' I|test
ahelper virus test DNA construct
I
CotranstectI
Propagate virusPassage 2
DNA 4Restriction
enzymes
E1 CM helper
+ ?detective
B.
an L
L. S
lb
3UL ;bamc ca
....~~~~~~~~~~~~~~~~~~...
Construct Ub Uc
DR1 Ub (DR2)22 (DR4)3 Uc DR1
.F---.---
*, *,A-2 + + SA DBA S S S B BSA E(DR2),
-D_
{I
A
-e_-B -e_-B
.s*
S S
b-S S
A
...XB.
A A
A A
S
Oa
B I K:
A-i + +
propagatior
H-i +A
B-i +
-C-i +
-E-i +
-I-l
A-C- D-1
L
+ [+helper]S
G-1 - + t-helper]
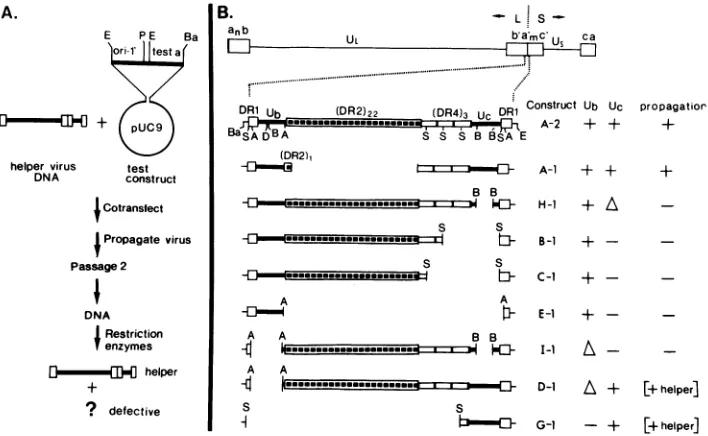
FIG. 1. Propagationtest and structureoftest constructs. (A) Scheme of the propagation test (28). (B) Structure of the test a sequences. Top line, schematic representation of the standard viral genome and the location of the a sequence in the L-S junction. Second line, arrangement of the directly repeated (DR) and unique (U) elements in the a sequence ofHSV-1 F DNA as described by Mocarski and Roizman (19). Shownarethe sites forBamHI (Ba),SstII (S),ApaI(A),DraI (D),BssHII (B), andEcoRI(E). The test a sequences (4) are contained between theBamHIandtheEcoRI sitesin thegeneralconstructshown inpanelA.Symbols:+or-,presence or absence of intact UborUcelements;A,deletionswithinthe Ub or Ucelements.The resultsofthepropagation tests are described as efficient propagation(+), lack of propagation (-),andpropagationatlowefficiency coupledtoselectionof genomeswith insertedhelpera sequence (+ helper).
cleavage-packaging process. Analyses ofthe products
re-sulting from the propagation ofseed amplicons containing
various deleted versions of the a sequence revealed the presence of two regions within the a sequence that are
essential for efficient propagation. The results also revealed aspects of the mechanism by which single a-containing junctions become cleaved.
MATERIALS AND METHODS
Cells and viruses. HEp-2 cells and Vero cells were
ob-tained from American Type Culture Collection, Rockville, Md., rabbit skin cells and HSV-1 strain F were obtained
fromB. Roizman, University of Chicago,and HSV-1 strain Justin was obtained from A. Sabin, who was then at the
University of
Cincinnati,
Cincinnati, Ohio.Derivation of plasmids. Cloning was done as previously described (28) with Escherichia coli DH1 (gift of D.
Hanahan, Cold Spring Harbor Laboratories, Cold Spring
Harbor,
N.Y.). Restriction enzymes and T4 DNA ligasewere purchased from New England BioLabs and Bethesda Research Laboratories. Calf alkaline phosphatase was pur-chasedfromBoehringerMannheimBiochemicals,and
poly-nucleotide kinase was purchased from P-L Biochemicals,
Inc. All enzymes wereusedasrecommendedbythe suppli-ers. The test constructs contained the sets of deleted a sequenceswhichwereconstructedby Chou and Roizman (4) as described in detail by them, starting from a 537-bp a sequencefromHSV-1 strain F. Briefly, theconstructs were made inpUC9 and contained3 bpof b sequences and 12bp
ofc sequences in addition to the a sequences. A series of deleted a subclones were made (Fig. 1B). In the present
study,a 1.90-kb EcoRIfragment, containingtheHSV DNA
replicationorigin
ori-i'
(oris;
20,31,36)wasinserted between theEcoRI sites of the deleted a clones derived by Chou andRoizman (4), yieldingthegeneral structure in Fig. 1A. The inserted EcoRIfragment contained a0.26-kb EcoRI-to-PvuII segmentofpKC7(24)and the1.64-kbori-' PvuII-to-EcoRI segment, derived from map coordinates 0.856 to 0.867 of HSV-1 F DNA. This ori-i' segment was derived originally frompRB373(20) andwastransferred topKC7 as described in our previous study (7). The construct A-2 contained the intact a sequence(537 bp) andwasderived by the insertion of ori-i' into pRB3119; clone A-1 was picked as a colony during thepreparation of the A-2construct.H-1wasderived frompRB3613 which containedaBssHIIdeletioninUc; B-1 wasderived from pRB3384 whichcontained a SstII deletion fusingthe second DR4 elementto theDR1element; C-1was derived from pRB3385 which contained a SstII deletion fusing the first DR4 element to DR1; E-1was derived from pRB3387 which contained an ApaI deletion fusing the Ub element to DR1; I-1 was derived from pRB3614 which contained a BssHII deletion in Uc as well as an Apal deletion in Ub; D-1 was derived from pRB3386 which contained an ApaI deletion in Ub; G-1 was derived from pRB3389 which contained a SstII deletion fusing the DR1 element to the third DR4 element. Plasmids pRB3386, pRB3613, and pRB3614 underwent intermediate recloning
intopUC9 before theori-i' segmentwas inserted,to obtain a uniform orientation of the deleted a segments in the plasmids. Asisterset ofplasmids was constructed with the EcoRI origin fragment in the opposite orientation. The constructs G-2, G-3, and G-4 were rescued from
[image:2.612.135.489.73.291.2]G-1-propagated defective genomes, as described in the text. Constructs G-5 and G-6wereconstructed (to eliminate Dral restriction sites) by insertion of a PvuII fragment which contained ori-i' and the G-deleted a sequence into a DraI-cleaved pBR322 plasmid. The PvuII fragment was taken from the sister clone ofG-1, which contained the EcoRI ori-i' segment in reverse orientation to that in G-1. The
...
l
*----.---
...s
J. VIROL.Bar
on November 10, 2019 by guest
http://jvi.asm.org/
HERPES SIMPLEX VIRUS CLEAVAGE-PACKAGING 607
- A-I B-I C-1 Etl H-l I-I D-1 G-1 .A-iG-l
(Jusi.aaa ba ba ba ba a b a b a a
i v l * - § helper j
~ ~ ~ v
a -
-ti
0-0323
0297
orX90.
1 2
8.a
0-.112C*a -_aOta303 a 04&o 278 *a.
-***or;90 *t
5 8 11 I5
17 18
FIG. 2. Analyses of DNA from transfection-derived virus
stocks.EcoRI digests of32P-labeled DNAfrom cells infected with passage2ofatransfection receivinghelperHSV-1 Justin alone (lane
1)orwithpassage2of duplicatecotransfections(aand b) receiving helpervirus along withthe test constructs shown(lanes 2 through
16). Thegroupof fragments visible in lane 1 correspond tohelper
virusDNAfragments. Shownare bandscontaining the replication
origin (ori), the intact single and double ajunctions (a and a-a,
respectively), thetestasequences (a*), and the chimeric junctions
(a*-a).Fragments arisingfrom digestion ofthegenerateddefective genome are marked by their sizes in kilobases. Lanes 17 and 18
show a shorter autoradiographic exposure of lanes 2 and 15,
respectively.
PvuII-DraI fusion resulted in the loss of both restriction sites. G-5 and G-6 differed in the orientation ofthe PvuII insert.
Cotransfectionandpropagationof virus stocks. Cotransfec-tion of rabbit skin cells with helper virus DNA and test
constructs was done as previously described (28) and as outlined in Fig. 1A.
Analysis of infectedcell DNA. Vero cell cultures (25 cm2)
were infected withpassage 2 virus and labeled with
32Pi
aspreviously described(28). At 20 h postinfection, DNA was
extracted from theinfected cells by lysis with amixture of sodium dodecyl sulfate and proteinase K which was fol-lowed byphenol-chloroform extraction and ethanol precip-itationaspreviously described(28). Following digestionwith the appropriate restriction enzyme, the DNA was
electro-phoresed in 0.5% agarose gels; 1/50 of the DNA produced
froman infected 25-cm2culture wasused perlane.
Nucleotidesequenceanalysis. For thenucleotide sequence
analysestheconstructsG-2, G-3, and G-4wereeach cleaved
with either EcoRI-PstI or BamHI-SstI. The appropriate
fragments containing theasequencewereelectroeluted, and theEcoRI orBamHI sites wereend labeled with polynucle-otide kinase. The nucleotide sequence was determined by theMaxam-Gilbert technique (17).
RESULTS
Propagation of amplicons containing deletions in the a
sequence.As reviewedabove, thea sequenceofHSV DNA
wasfoundto be essential forthe cleavage and packaging of viral DNA and therefore for the propagation of defective
genomes in serially passaged virus stocks. To map the
cleavage-packaging signal(s)within theasequencewetested
the ability of test constructs containing specific deletions in the a sequence to becomepropagated invirusstocks. Figure
1Asummarizes the general structure of these test constructs and the features of the propagation assay. All of the test constructs contained the replication origin
ori-i'.
In addi-tion, they contained either an intact a sequence derived from HSV-1 F or various deleted a segments derived from it (Fig.1B). The intact a as well as the deleted a segments were derived from plasmids constructed previously by Chou and Roizman (4), starting from the cloned a sequence ofHSV-1
F DNA. All constructs used in the present study were designated according to the a sequence which they con-tained; the lettering system of the original study of Chou and Roizman (4) was used. The constructA-1, which was used as a control for this set of propagation tests, contained an a sequence of 285 bp, i.e., close to the average size of the deleted test a constructs (range 86 to 497 bp). The 285-bp variant a sequence contained only a single DR2 element and in that respect resembled the shorter type a sequences found in some HSV-1 strains (6, 18, 35).
In the propagation tests, duplicate cell cultures (a and b) were cotransfected with helper HSV-1 Justin DNA and the control or test constructs. Virus stocks derived from these transfections were serially passaged, and
32P-labeled
DNAsfrom
cells infected with passage 2 of the resultant series were analyzed by digestion with EcoRI (Fig. 2) and additional enzymes (not shown) to test for the presence of defective genomes generated from the seed constructs. As seen in the maps shown in Fig. 3 (e.g., for the D-1 andG-1constructs),
two fragments were expected to arise byEcoRI digestion of concatemers generated by replication of the seed constructs: (i) a fragment of 1.90 kb, which contains the replication origin and was common to all constructs, and (ii) a variable-sized fragment containing the respective test a sequences.
The analyses revealed the following. (i) As expected, the control construct A-1 gave rise to abundant defective genomes composed of head-to-tail repeats of identical size to that of the input seed construct (represented by the 1.90- and 2.97-kb fragments; Fig. 2, lane 2). A minor proportion of the repeat units in the generated defective genomes contained tandemly reiterated a sequences, apparent as the "ladder bands" with increments of ca. 260 bp, extending from the
2.97-kb EcoRI band. (ii) Virus stocks derived from transfections receiving the constructs B-1, C-1, E-1, H-1, and I-1 did not contain authentically propagated defective genomes (Fig. 2, lanes 3 to 12). All of these constructs
containeddeletions affecting the Uc element, with the small-est deletion present in the construct H-1 (40-bp deletion
boundedbythe BssHII sites in Uc). (iii) Virus stocks derived fromcotransfections receiving D-1 and G-1 contained defec-tive genomes with repeat units identical to the input seed
constructs (yielding the 1.90-kb origin fragment and the fragments designated a* in Fig. 2, lanes 13 through 16).
However,the abundance of generated defective genomes in the population was relatively low compared with their
coun-terparts generated from the control construct A-1 (compare the relative intensities of defective and helper virus frag-ments in the short autoradiographicexposures showninFig. 2, lanes 17 and 18). In addition, the defective genomes contained significantproportions of repeats that were ca. 240 bplarger than the input seed plasmids (the 3.37-kb fragment in lanes 13 and 14 and the 3.03-kb fragment in lanes 15 and 16). As shown below, the increase in size resulted from the
insertion of an intact a sequence, derived from the helper virus DNA, within the repeat unit junctions. Consequently, these bands have been designated a*-a in Fig. 2.
VOL.59, 1986
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.65.284.72.275.2]C
I
E
coR
Ia*
Oral
Dra
Ia-a*
E
coRI
EB
Bp c
D- D
G1 D
E
L 312 1.90 3.12 1.90
2.78 1.90 2.78 1.90
E B E E B E EB
DDD D DDC D DOD
_
69 4A31 69 4.31
I
.69iI~
3.98 .69I 3.9869 2.97 i 162 69 2.97 i162
E0B 69i 297 }125 0691 297 1255
E
DB.69
E.
E©B E E®BD D D E DD
ED
D DD_a
a t + t"a+ a-a+--a--33 11.90 3.36 1.90
3 0 3 1.90 3.03 1.90
D-1 G-1
D-1 G-1
D-1 G-1
0-1 G-1
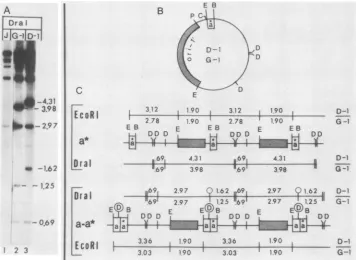
FIG. 3. Structureof chimeric junctions. (A)Dral-digested32P-labeledDNAs from cells infected with passages 2 of transfections receiving HSV-1 (Justin)helper DNA alone (J) or the constructs G-1and D-1. The DNAs correspond to those shown in Fig. 2, lanes 1, 16, and 14, respectively.(B) structure ofthe two testconstructs. Shownare locations of theori-l'fragment (stippled area), the test asegment(a*) and the sites forEcoRI(E),BamHI (B),DraI (D),ClaI (C), andPvuII (P). (C) predicted structures of generated defective genomes with repeat unitscontainingthe original deleted a sequence (a*) or the chimeric repeats (a-a*) with inserted helper virus a sequence. Sizes of generated EcoRIandDraI fragmentsaregiven inkilobases forthe D-1-generated (above the line) and G-1-generated (below the line) concatemers. The DraI sitepresent in the Ub element of theinsertedhelper virus a sequence is circled.
Two conclusions can be reached on the basis of these
observations. First, the failure of the constructs lacking the Ucelement (Uc-) to be propagatedin virus stocks revealed that the sequences bounded by the BssHII sites within the Uc element were essential for cleavage-packaging. These
boundaries are based on the 40-bp deletion in the H-1 construct. Second, the low-efficiency propagation of D-1
(containing a 64-bp deletion in Ub) suggested that the sequences bounded by the ApaI sites in Ub affected the efficiency of amplicon seed propagation. In addition to
testingthe constructs above, we tested (in duplicate sets) the
corresponding a deletions in plasmids which contained the EcoRI 1.90-kb fragment (carrying the replication origin) in
the opposite orientation to that in Fig. 1A. The results of
these studies(data not shown)yieldedthe sameconclusions asabove.
Structure of the larger repeat units derived fromthe
Ub--Uc+ constructs. The experiments described in this and the next sections revealed that the larger repeats found in the D-1- and G-1-derived defective genomes (Fig. 2, lanes 13 through 16; band a-a*) contained an insertion of an a sequence derived from the helpervirus DNA. Specifically,
because the increased size of the repeat units reflected
changesin thefragment carrying theasequence(Fig. 2; data notshown), the larger repeats could be derived by tandem reiterations of the deleted a sequence or, alternatively, by
insertion of an a sequence derived from the helper virus DNA. To test these alternatives we took advantage of the fact that the input test constructs did not contain the Ub sequence and therefore lacked the DraI site (TTTAAA) at position 31 of the a sequence. TheDral enzyme cleavesin three locations within the bacterial plasmid portion of G-1
andD-1,generatingcommonfragmentsof 19 and 692 bp, and the variable (a-containing) fragments of 3.98 and 4.31 kb
(Fig.3). Self-amplification ofthe test a sequences in G-1 and D-1was predicted toresult in increased size of the variable bands. In contrast, insertion of the a sequence from the
Justin helperDNA should result in cleavage ofthese
frag-ments, owingto the presence ofa DraI sitein the inserted Ub sequence. In this case, two new fragments would be
generated, with sizes dependent on the extract site of insertion. The DraI cleavagepatterns(Fig. 3) revealed that the novel repeats indeed containedan additional DraI site,
consistent with an insertion ofan intact (Ub+) a sequence derivedfrom thehelpervirus DNA. In addition,the results showed that thehelpervirusa sequencewasadjacenttothe test (Ub-deleted) a sequence on the side proximal to Uc, creatingachimericdoubleajunction.Specifically,defective genomesgeneratedfrom theconstructsD-1andG-1 yielded a common0.69-kb Dral fragment generated from the
plas-mid region, the a-containing Dral bands of sizes 4.31 and 3.98 kb representing theinputDNA (see map labeleda* in
Fig. 3), and two novel fragments not present in
the
input seed,of sizes 2.97 and 1.62 kbforD-1,and 2.97 and 1.25 kb for G-1. The sizes of these novelfragmentswereconsistent with the insertions of helper a (Ub+ and hence DraI con-taining) occurring on the side proximal to Uc (see map labeled a-a* in Fig. 3).Nucleotide sequence of the novel chimeric junction. To
further analyzetheeventswhich ledtotheformation of the chimericjunctions we shuttled the repeat units of the G-1-derived defective genomes back into bacteria and obtained the nucleotide sequence for three of the resultant clones.
Specifically, unlabeled passage 2 DNA from a
G-1-A_
A *- 2.97 DraI
1- -1.62
123
_ _
__-4.31
_0
3.9800
-2.97#& -1.62
-t.25
-0.69
X 2 3
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.129.485.75.335.2]HERPES SIMPLEX VIRUS CLEAVAGE-PACKAGING
Inserted a
-'4i1 J ---'I" ?/ F
DR cDRI1 Ub DR2/3 S/4 Uc DRI
Sathl
CZWAG CCCGGGCtGCg CCGCCGCgc
(RauHI)... CCG
(BRaHI)...CCG
CCGCGGGGGGCCCGGGCTGC cg CCGCCGC
DR11
-(CGCTCCTCCCCc)19
TTMAAGGGCCGCGCGCGACCCCCGGGGGGTGTGTrCGGGGGGGGCCCGTTTT
TTTAAAGGGCCGCGCGCGACCCCcGGGGGGTGTGTTTCGGGGGGGGCCCGTTTT
(c(;CTCCTCCCC
Sstll
GCTCCCGCCCCGCCCCC _ ACGCC)3 CGGCCCCGCCCCC a ACGCC CGGCCCCGCCCCC AACGCC
tggggtctggc (CGCTCCTCCCC )2 (gtctgtgggtgggGCTCCTCCCC)2GCTCCCGCGGCCCCGCCCCC[cACGCC
--DR2-- - DR4
- DR353
CGCCGCGCGCGCGCACGCCGCCCGGACCGCCGCCCGCCTTTrrTGCGCGCCGCCC CGCCGCGCGCGCGCACGCCGCCCGGACCGCCGCCCGCCTTTTTTGCGCGCCGCCC
CGCCGCGCGCGCGCACGCCGCCCGGACCGCCGCCCGCCTTTTTTGCGCGCCGCCC
CGCCGCGCGCGCGCACGCCGCCCGGACCGCCGCCCGCCTTTTTTGCGCGCCGCCC
-a-Deleted al
cgc CCGCGCCGGGCCCGGGCTGC cgc CCGCGGGGGGCCCGGGCTGC ...(REcoRI)
cgc CCGCGGGGGGCCCGGGCTGC g CCGCGGGGGGCCCGGGCTGC
- *~ DR1
I
Inserteda_-.MgCCGCCGC ic TTAAG CGCCGcCGCGACCCCCGGGGGGTGTGTTTCGGGGGGGGCCCGTTTT
Icg ICCGCCGCcgcTTTAAAGGGCCGCGCGCGACCCCCGGGGGGTGTGTTTCGGGGGGGGCCCGTTTT 1CgCCGCCGCI TrTAAAGGGCCGCGCGCGACCCCCGGGGGGTGTGTTTCGGGGGGGGCCCGTTTT
HSV-1(F) (CGCTCCTCCCCc)lg G-3/4 tggggtctggc (CGCTCCTCCCC )2 HSV-1(J) tggggtctggc (CGCTCCTCCCC )2
_ DR2 _
HSV-i(F) CGCCGCGCGCGCGCACGCCGCCCGGACC4 G-3/4 CGCCGCGCGCGCGCACGCCGCCCGGACC4 HSV-1(J) CGCCGCGCGCGCGCACGCCGCCCGGACC(
C.
HSV-1 (F) HSV-1 (J) HSV-1 (KOS) HISV-1 (17)
HSV-1 (USA-8)
chimera G-3/4
49/50 conserved
.T..TTTTGCGCGC
... TTTTTGCGCGC ... TTTTTGGCCGC
... ITT GOGCGC
... TITTTGCGCGC
... TTT GCGOGC
(cGCTCCTCCCC GCTCCCGCGGCCCCGCCCCC aACGCC)3
gtctgtgggtgggG... GCTCCCGCGGCCCCGCCCCCcACGCC
(gtctgtgggtgggGCTCCTCCCC)2GCTCCCGCGGCCCCGCCCCC cACGCC
__________ DR4
8
DR3.5.-CCGCCCGCCTTTTGCGCCGCCC cgc CCGCGGGGGGCCCGGGCTGC
GCCGCCCGCCrTTTTTGCGCGCCGCCC cgc CCGCGGGGGGCCCGGGCTGC ...(EcoRI)
CCGCCCGCCTTTT]rTGCGCGCCGCCCLg CCGCGGGGGGCCCGGGCTGC
Uc DR1
, 1a DRl-- I'I a -Ub
variable 19/20conserved variable 51/53conserved
cg5cccgc CCGCGGGGGGCCCGGGCTGC gccgccgcgc TTTMA ...
cgcCCg CCGCGGGGGGCCCGGGCTGC cgccgccgc TTTAAA
gcgcgcgc CCGCGGGGGGCCGGGCTGC ccgccgcccgcc TTTAAA ...
gcgcgcgc CCGCGGGGGGCCCGGGCTGC ccgccgccaccgc TTMA ...
cgccccgc CCGCGGGGGGCCCGGGCcGC ccgccgccgcgc TTTAAA ...
[image:5.612.136.483.71.455.2]cgccccgc CCGCGGGGGGCCCGGGCTGC cgccgccgccgc TTTAA
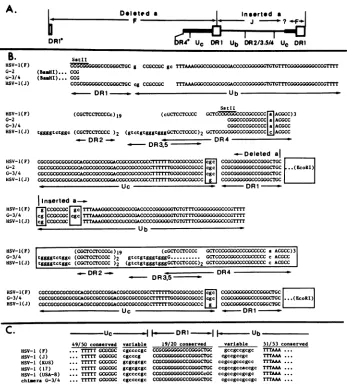
FIG. 4. Nucleotidesequencesof the chimeric junctions andsequenceconservation in the Ub and Uc elements of different HSV-1strains. (A) Generalstructureof the chimericjunctions in clones G-3 and G-4. The Ub--Uc+asequencefrom theconstruct(denotedasdeleteda)
and the insertedasequence arearranged in tandem. The heavy line denotessequencesoriginating from F; thinner line denotes sequences originating from strain Justin (J). DR1*and DR4* indicatepartial elements thatarefusedattheSstll siteasinthe seed G-1construct. ?,A
region conserved between F and Justin; therefore, the site of transition from Justin to F sequences in Uc is unknown. (B) Nucleotide sequencesof theasegmentsin the authentically sized (G-2), and larger (G-3 and G-4)repeatsandthesequences of doubleajunctions of
HSV-1strains F(19, 21) and Justin (18). Sequences which differ in the Justin and F strainsareboxed. The locations of the EcoRl and BamHl sites inthepolylinkerareshown. Thedotted line in the DR3.5repeatindicatesareaofsequenceuncertainty. (C) Sequence conservation in the Ub and Uc elementsin differentHSV-1 strains and in the G-3 and G-4 chimericrepeats.Theregions proximaltothe DR1arevariable,
whereas thosefartheraway areconserved. Nucleotidesequences arefrom Mocarski andRoizman (19, 21) for strain F, Mocarskietal. (18) forJustin(J), Varmuza and Smiley (35) for KOS, and Davison and Wilkie (6) for strains 17 and USA-8.
propagated virus serieswascleaved with ClaI which cleaves oncewithineachrepeatunit(site denotedby Cinmap [Fig.
3, top]). The cleaved fragments were resolved by gel
elec-trophoresis,andmonomericfragments correspondingtothe authentically sizedrepeatsand the larger (chimeric)repeats
wereelutedfrom the gel, circularized, and usedtotransform bacteria. We then determined the nucleotide sequences of
the a sequences in one of the authentically sized clones
(designated G-2) and in two ofthe larger chimeric clones (designated G-3 and G-4). The results are shown in Fig. 4
along with the corresponding DNA sequences of HSV-1
strain F (21), from which the deleted a construct was
derived, and HSV-1 strain Justin (18), which served as
helper virus. Theasequencein G-2wasfoundtobeidentical tothedeletedasequenceinthe seedHSV-1(F)-derived G-1
clone(4). Thus,itcontained the fused(attheSstIIsite) DR1
and DR4 elementsand the entire Ucelementfollowedby the DR1and linkersequences. Thenucleotide sequencesofthe chimericdouble ajunctionsin the twolarger cloned repeat units G-3 and G-4 were identical and contained several
interesting features. (i) Starting from the DR1-DR4 fusion site (SstII), the sequences were identical to those in the rescued, authentically sized clone (G-2) up to the DR1 junction. Thisincluded the DR4 and Uc elements in which the Justin strain differs from the F strain by 1 and 2 nucleotides, respectively (Fig. 4B, the boxed A and C in DR4 and theboxedCGC and G inUc). Thus,the insertionof the helper virus a sequence evidently occurred within the DR1 sequence. (ii) The inserted a sequence was generally
similar to the a sequence of the Justin helper virus DNA. Thus, the chimeric junctions contained the Justin-specific
sequence TGGGGTCTGGC between the Ub and DR2 ele-VOL. 59, 1986
A.
b
DRl
Dolsted a
- F
609
B.
HSV-1(F) G-2
U-3/4
HSV-1(J)
HSV-1(F)
G-2
G-3/4 HSV-1(J)
HSV-I( F)
G-2
G-3/4
HSV-1(J)
IHSV-1(F) G-3/4 HSV-1(J)
_--
-L
on November 10, 2019 by guest
http://jvi.asm.org/
A...,.EcORI EcoRI
Just in Helper F FHe1 er
a|b | a
|b
a|b Iab ab520[4hI
240480
24 1~~480
ON
1 2 3 5 7 9 11
B. EcCORI
FG
5i
G 5 A 2 1A 2 a b a b |
a.. -aa
a..
3 81 *
318
* * ~1.90
3 5
FIG. 5. Propagation of test constructs in the presence of Justin and F helper viruses and in transfection competition tests. (A) EcoRI digests of32P-labeledDNAsfrom duplicate transfections (lanes a and b) receiving the HSV-1 strains Justin (lanes 1 through 6) or F (lanes 8 through 11) helper virus DNA along with the indicated testconstructs. Lane 7 contains theEcoRI-digestedF helper DNA. The increments (in base pairs) between fragments containing single a and double a junctions are indicated. (B) Propagation competition tests. EcoRI digests of32P-labeledpassage 2 DNAs from transfections receiving helper virus DNA alone (lane 1) or with theG-5 construct (lanes 2 and 3), the A-2 construct(lane 6), or an equal mixture of both constructs (lane 4 and 5). Fragmentsderivedfrom thedefective genomes are indicated by size. a anda* denote fragmentscontaining the intact and Ub--Uc+ a sequences present in the seed A-2 and G-5 constructs, respectively. a* + aj denotes the fragment containing the chimeric junction with helper Justin a sequence; a + aFdenotes the double a junction arising by self-amplification of the intactasequencein A-2.
ments, two copies of the DR2 repeat, the Justin-specific DR3.5 repeat, and a single copy of the DR4 repeat (these
elementsareboxed inFig. 4B). (iii) two sequence alterations appearedinbothchimericrepeatclones.First, 10 bp into the
Ubelement,thechimericjunctions in G-3and G-4 contained
the triplet CGC (boxed in Fig. 4B) which was absent from the Justinhelper virusasequence.Second, the distal portion
ofthe Uc elementappearedtobederived fromF(construct) rather thanJustin (helper) DNA (Fig. 4A), inasmuch as at
the Uc-DR1transition, theinserted a sequences resembled thesequanceofthe F rather thanthe Justin strain(Fig. 4B,
boxed CGC and G).
Twopointsarenoteworthyconcerning thisresult. First,it
islikely that the sequence variations reflectfeatures ofthe
recombination events leading to the formation ofthe
chi-meric junctions, rather than sequence variation related to propagation in virus stocks. Thus, the nucleotide sequence
of the a sequence in the G-2 repeat (cloned following propagation) was identical to that in the input a construct
(G-1), whereas both chimeric a clones exhibited the same sequencealterations. Second, the sequence alterationswere ofinterest inthat, in the HSV-1 strains sequencedthusfar,
the sequences immediately adjacent to the DR1 (at the Uc-DR1-Ubtransitions)arequite variable,whereas the bulk
ofthe sequences making up the Uc and Ub elements are highlyconserved (Fig.4C). Itispossiblethat thevariability
in the Uc-DR1 andDR1-Ubtransitions within the chimeric
double a junction reflected inherent variability in these
regions. Apossible explanation for the source of sequence variations is proposed in the Discussion.
Essential role of Ub in cleavage-packaging. Insertions of
helper virusDNA sequencesinto defective genomes had not been detected inourprevious studies thatinvolvedthe use of amplicons with heterologous helper viruses (F strain-derived amplicons in combination with Justin helper, and
viceversa; see, e.g.,reference7). Apossible explanation for
theobserved insertions into the D-1- and G-1-derived defec-tive genomes was that the Ub deletion in these constructs affected the cleavage-packaging signal and therefore
chi-mericjunctionswhichrestored the deletedUbsequencesin an appropriate arrangement were selected forpropagation. Threepredictions of thishypothesis weretested below and
found to be correct. The first was that helper virus DNA insertions would not be detected in defective genomes
propagatedfromconstructscontaininganintactasequence. The secondwasthat incompetitiontests,theUb--Uc+ test plasmids willbe propagated with lowerefficiency than the
intacta constructs. Thethirdwasthat theUb--Uc+ repeat units will be propagated from passage to passage within heteroconcatemers, i.e., mixed concatemers containing (i)
repeats with the Ub- a sequence and (ii) repeats with the chimeric (helper a-containing) junctions. The cleavage-packaging functions of these heteroconcatemers could be
provided, at least in part,by thechimericjunctions. Transfer of the helper asequence into different seed con-structs.Totestthe firstpredictionweanalyzedthe patternof aamplificationin defective genomesderivedbypropagation
of the intactaconstructA-2and theUb-constructsD-1 and G-1 (Fig. 1B). Each of these constructs was used to cotransfect
duplicate
cultures in the presence ofHSV-1(F)
orHSV-1(Justin) helperDNAs(possessinga sequences of 501 and 264 bp, respectively) to determine whether the
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.140.477.73.321.2]HERPES SIMPLEX VIRUS CLEAVAGE-PACKAGING 611
A.
EcoRi Dral____
J G-5a
Jb
baG-6bG-5b
;ab;a6
a: b a bI1.49
8.88- - _I_
-5.87
4,88
3.80- -
-3.56- -W0_
B.
IR EOP
/t G-5
0 562
kID
E
0P
D E
, G-6
O.c 562kb
DP
*
C. BEE E BE *E BE wE BE BE BE wE
1.
_ 1-_-EcoRI _206 35o, '? _ 35ob 2^06 25c 206 356 206
Dra - Resistanl
B1E wE EOE LE BE *E BDE rE B E a,E
2.
EcoR 2.06 360 200 3.5o 2.06 380 206 356 206
5.r ? 49
DraFl
..I -w--- _--- ---_-_- _ _B D E E BC3 E E BE *E B DE
2.06 *. 3 --- or ' o
EcoRI 206 256 206 281
1 5 6 10 Oral _ 005 ,1657
FIG. 6. Structure of concatemers arising by propagation of the G-5 and G-6 test constructs. (A) EcoRI andDraI digests of 32P-labeled passage2DNAsfrom transfections receiving helper Justin DNA alone or with the G-5 or G-6 test constructs. Sizes are indicated in kilobases. The underlined sizes correspond to fragments which arise from the large inserts of helper virus DNA. The corresponding maps for the G-5-derived genomes areshown in panel C3 and accountfor the novel 8.88-kbEcoRI band and the 10.95 and 16.57-kb Dral bands. The constructG-6 also produces largerecombinant repeat units with expected 6.07-kb novelEcoRIfragment and 10.95- and 16.57-kb novel DraI fragments (maps not shown). (B) Structure of the constructs G-5 and G-6. The EcoRI-to-PvuIl segment contains ori-l'; a* denotes the Ub--Uc+asegment. The locationsof theEcoRI (E), BamHI (B), andthefusedDraI-PvuII (D/P) sites are shown. (C) Concatemeric structures for the G-5 construct.Cl,Homoconcatemerscontaining authentically sized (input) repeat units. i,DraI-PvuII fused site at the border of the ori-1' fragment. C2, Structure of heteroconcatemers containing interspersed chimeric and authentic junctions. Black rectangles denote the Ub--Uc+asequence (a*), and open rectangles denote the inserted helper virus a sequence. Cleavage of the heteroconcatemers with Dral isexpectedtoproducebands with increments ofauthentically sized repeats. The firsttwobands (5.87 and 11.49 kb) are shown both here and inpanel A, lanes 7 through 10. C3,Heteroconcatemers containing repeat units with an inserted helper DNA segment spanning the region from the a sequence to the replication origin ori-1'. The predicted sizes of fragments generated from EcoRI and Dral cleavages of the heteroconcatemers are shown for the construct G-5. The corresponding first two bands in the predicted ladder generated byDral cleavage (10.95and16.57kb) areunderlined in panel A, lanes7through 10, as is the novel 8.88-kbEcoRIfragment.
amplified a sequences were derived from the helper virus
DNA or from the seed test plasmid. Restriction enzyme
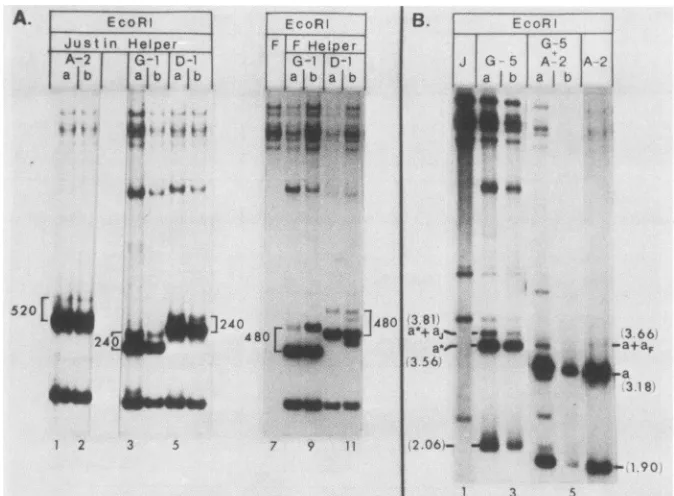
analyses ofpassage 2 DNAs from thesetransfections (Fig.
5A for EcoRI; additional data not shown) revealed the
following. (i)Theincrementbetween thebands representing
the double a and single ajunctions in defective genomes
propagated from A-2(intacta) in the presence of the Justin helper virus was ca. 520 bp (fig. SA, lanes 1, 2). Thus, the
double a junction was derived by self-amplification ofthe
537-bp a sequence ofthe seed construct(minus the shared
DR1 element) rather thanby insertion of the 264-bp helper
virus a sequence. (ii) In contrast, the increments between the double a and single ajunctions in defective genomes
propagated fromthe D-1 and G-1 constructs varied with the
helpervirus.Specifically, inthepresence of the Justin helper
(possessing a 264-bp a sequence) the increment
corre-spondedto ca. 240bp (Fig.5, lanes 3 through 6), whereas in thepresence ofthe Fhelper (501-bp a sequence) it was ca.
480 bp (lanes 8 through 11). These values were consistent
with insertions of the helper a sequences into the chimeric
junctions. Furthermore, bands arising from self-amplifica-tion of the Ub- a sequences (100 bp in G-1 and 470 bp in D-1) were notdetected.
The data support three conclusions. (i) The intact a sequence contains a complete set of cis-acting signals needed for the cleavage-packaging of viral DNA, as well as for the self-amplification of a sequences. (ii) Fordefective genomes containing intact asequence, the trans insertion of
a sequences from helper virus DNA is less efficient than
self-amplification of the a sequences. (iii) the Ub deletion affectsfunction(s) requiredfor aamplification, as well as for
defective genome propagation. Hence, defective genomes derivedfromthese constructsrequirethe presenceofat least some chimericjunctions (in the same concatemer, see
be-low) for their propagation.
Competition propagation tests. Studies described below
revealed that constructs containing the Ub--Uc+ a se-quences werepropagated invirusstocks less efficientlythan
theircounterparts containingthe intact a sequence. Specif-ically, duplicate cultures were cotransfected with helper
Justinvirus DNA and (i)the A-2intact a construct, (ii)the construct G-5, which contained a Ub--Uc+ a sequence
identical to that in G-1 (map in Fig. 6), or (iii) an equal mixture ofbothplasmids.Therelative propagationefficiency
ofA-2 and G-5 could be determined fromthe ratios ofthe
distinct restriction enzyme fragments which they were
ex-pectedto yield.
Figure SB shows the EcoRI digests of passage 2 DNAs from these transfections. Defective genomes with repeat units derived from the intact a construct were abundant in stocks derivedfromthe single(A-2; lane 6) and mixed (A-2 +G-5,lanes4and5) transfections(see bands 1.9 and 3.18 kb in these lanes). Defectivegenomes derived from the Ub- a construct were propagated in stocks derived from the transfectionsreceivingthe G-5plasmid singly(lanes 2 and 3; bands 2.06 and 3.56 kb). However, in stocks derived from VOL.59, 1986
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.91.530.78.301.2]themixed transfections, they were present only in very low abundance (lane 4) or were not detected at all (lane 5). (In lane 4, compare the intensities of the 1.90 and 3.18 kb bands
representingthe intact a construct with those of the 2.06 and 3.56 kbbands representing the Ub--Uc+ construct.)
These results demonstrated that the G-5 clone which
contained the Ub--Uc+ a sequence was propagated
ineffi-ciently, consistent with a defect in the cleavage-packaging
signal(s). In addition, the increments observed between
fragments containing the double a and single a junctions in the A-2- and G-5-derived repeats were consistent with the
results discussed earlier. Thus, A-2 repeats contained a
self-amplifieda sequence (a + aF; Fig. SB, lane 6), whereas
G-5-derivedrepeats containedaninserted Justin a sequence
(a* +aj; lanes 2 and 3).
Structure of concatemeric DNA molecules. As discussed
above, test constructs lacking the Ub element appeared to contain a nonfunctional cleavage-packaging signal. At the same time, authentically sized repeat units carrying the
Ub--Uc+junctions were recovered in propagated defective genomes (along with chimeric junctions containing helper a
sequence).Infact,they represented the major repeat units in
the serially passaged virus populations. The studies
de-scribed below provide an explanation for this puzzling
observationin thattheyrevealed that repeat units containing the
Ub--Uc'
junctions were propagated as constituents ofheteroconcatemers containing interspersed chimeric
junc-tions. Specifically, in these studiestheUb--Uc+ asequence was first introduced into plasmids lacking Dral sites. Since the DraI site in the Ub element could serve as the hallmark
for the inserted helper virus a sequences, the degree of
interspersion of chimeric and authentic repeat units within the same defective genome molecules could be determined
by analyzing the distribution ofDral sites along defective genome concatemers.
For the construction of the test clones lacking DraI, a PvuII fragment containing the replication origin and the
Ub--Uc+ a sequence was introduced into a Dral-cleaved
pBR322, resulting in the inactivation of both sites and
yieldingtheplasmidsG-5and G-6,differingin the orientation
ofthe PvuII insert(Fig. 6B, top). ThepredictedEcoRI and
Dral cleavages of homoconcatemers and (the simplest)
heteroconcatemers are shown forthe construct G-5 in Fig.
6C,
panels 1and 2. Usingthe G-5 and G-6testconstructsasinput
seed clones, we expectedthat homoconcatemerscon-taining solely the Ub- a sequences would be resistant to DraI(Fig. 6C, panel 1). In contrast, randominterspersionof the chimericjunctions (each containing a Dral site in Ub) within heteroconcatemers containing the authentic repeat units(devoid oftheDral site)wouldcreate aladderof Dral
fragments
with increments of single, authentically sized repeats. Because the authentically sized repeats were 5.62 kb and because the repeats withchimericdouble a were 5.87kb,we predictthatDralcleavage of the heteroconcatemers will generate a ladder described by 5.87 kb + n (5.62 kb), wheren isan integer, i.e., 5.87, 11.49, 17.11 kb, etc. (Fig.
6C;
panel 2).TheEcoRI andDral digestionpatternsof passage2 DNAs fromduplicatetransfectionsreceiving HSV-1 (Justin) helper
alongwith the G-5 and G-6 constructsareshown inFig. 6A. The DraI cleavage pattern of the G-5 and G-6 defective genomes contained bands of sizes predicted from random
interspersionof chimeric junctions within the concatemeric defective genomes (Fig. 6A). Interestingly, the DraI and EcoRI patterns revealed the presence of an additional type of chimeric repeat unit within the same defective genome
concatemers. Thefragments resulting fromcleavageofthese interspersed novel repeats are underlined in Fig. 6A and their structure is depicted in Fig. 6C, panel 3, for the construct G-5. In these novel repeats, the construct se-quences between the deleted a and the replication origin
havebeenreplaced by helper virus DNA sequences extend-ing from the a sequence to the replication origin in the S component. The presenceofthese repeatunits in the serially
passaged virus stocks was surprising in that, as already
noted above, we did not encounter chimeric repeat units containing inserted helper virus DNA sequences during propagation ofamplicons containing afunctional replication origin and anintact a sequence. Mostlikely, thesechimeric
repeats were selected for propagation and might in fact represent intermediate structures in the formation of the
chimeric junctions serving to propagate the
Ub--Uc+-containing seeds. Models for the evolution of these large repeats are considered in the Discussion.
DISCUSSION
Distinct roles of Uc and Ub elements in the cleavage-packaging process. We used amplicon propagation tests (28) to mapthe signals required for the cleavage and packagingof
HSV DNA. The constructs tested in these studies fell into three distinct categories. Constructs in the first category (A-1 and A-2) contained an intact a sequence and yielded defective genomes which did not contain additional se-quences derived from the helper virus genome. Constructsin the second category (D-1, G-1, G-5, and G-6) contained deletions in the Ub element but retained the Uc element. Defective genomes with authentically sized repeats were recovered from these constructs. However, propagation resulted in the selection of chimeric repeat units contain-ing intact a sequences derived from helper virus DNA. Moreover, in transfection competition tests the Ub--Uc+ defective genomes did not compete efficiently with their counterparts containing an intact a sequence. Finally, test constructs in the third category (B-1, C-1, E-1, and H-1) contained deletions in the Uc element and could not be propagated into defective genomes. Taken together, these data revealed that the Ub and Uc elements played critical yet distinct roles in the cleavage-packaging process. The in-volvement of the DR2 and DR4 elements in the cleavage-packaging process is at present unknown.
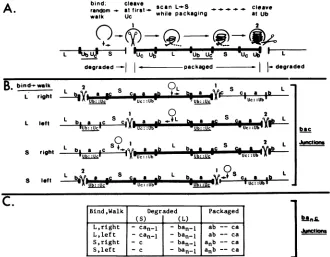
Directional packaging modelfor cleavage-packaging of viral DNA. In this and the next section we propose two alternative models for the cleavage-packaging of viral DNA. Both models are designed to explain the data summarized in this paper and to account for previous observations related to the cleavage-packaging process. The first model (the directional cleavage model) modifies the "theft" model proposed re-cently by Varmuza and Smiley (35) to include polarity inthe cleavage-packaging process. The model is schematically diagrammed in Fig. 7A for standard virus DNA concatemers and includes the following steps. (i) A packaging complex encounters the HSV DNA anywhere in the L or S compo-nent and traverses along the concatemeric DNA in either direction (random walk; Fig. 7A) until a junction containing a Uc signal is found. (ii) Cleavage (producing 3' single-base overhangs) occurs at the DR1 element proximal to the first Uc signal encountered (cleavage 1; Fig. 7). If the complex traverses into a
ba,c
type junction starting from the L component (ca. 80% of the cases, based on the relative sizes of L and S, and assuming random initial binding) this first cleavage will produce ba andca,
-1termini (Fig. 7B andC).on November 10, 2019 by guest
http://jvi.asm.org/
HERPES SIMPLEX VIRUS CLEAVAGE-PACKAGING 613
A.
bind; cleaverandom -. atfirst scan I-eS C___cleave
walk Uc while packaging at Ub
[image:9.612.136.466.76.333.2]~~~''Uc UIL UbU, SU LII L
degraded -| packaged b |-degraded
B. bind- walk 2
OLSS
LleftI
SUCm
b
.b a Cs ab1
L right a
EUbI
Uc:CIvUc: :UbY
I*L0 2
L left bi au 1)
P
a
Cb.b* - * E bac
ih
L ba a Mc+IX>
a
b bm a scU I^b
Lmj
U,Lb:S{ Uc S YPU@M b bUb: vUc -6
u-Bind,Walk Degraded Packaged
(S) (L)
L,right - c.an- - ban.I ab -- ca
L,left - can-I - ban_I ab -- ca S,right - c - ban.I anb -- ca
S,left - c - ban-I anb -- ca
b]n.L
JuctonFIG. 7. Directional cleavage model.(A)Basicfeatures ofthemodel.Thepackaging complex(open circle)bindsto thestandardviral DNA concatemer at arandomlocation andtraverses the concatemer in a random walk(rightorleftinthefigure).Thefirstcleavage[1] occurs at aconstantdistancefromthefirstUcsignalencountered, leavinga3'single-baseoverhang. Inthe standard viralgenomethiscleavage occurs inthe DR1 element.PackagingbeginsatthegeneratedLterminus(whichcontainsatleast one a sequence),whereas thegenerated S terminus (which is devoidofa sequences) is rapidly degraded. While packaging in the L toS direction, theDNA isscannedfor the nextdirectly repeated junction, skipping the invertedjunction (underlined).Asecondcleavage[2]occurs at aconstantdistancefrom the first Ubsequence encountered in the secondjunction, leaving 3' single-base overhangs and producingapackaged molecule (heavy line) which carries a ca
terminus, and anunpackaged terminus which isdegraded. (B)Diagram of thecleavagewithinsingleajunctions inastandard virus DNA concatemer,featuringthefourpossible permutations of initial binding and random walk. Assuming randombinding, ca. 80%oftheinitial bindingeventsshouldoccurinL(lines1and2) andca. 20% shouldoccurinS(lines3and4). (C) Termini produced in packaged and free moleculesas aconsequenceof thecleavage-packaging processstarting from each ofthefourinitial binding permutations.nrepresentsthe number ofasequencesinthejunctions whicharebeingcleaved.
If the process began within the S component, the first cleavage will generate ban and c termini. In both cases
directionalpackaging will begin fromthegeneratedLtermini (baorban), whereas the generated S termini(ca, - 1orc) will
be rapidly degraded. (iii) Packaging will continue by
scan-ningin theL-to-S direction,untiladirectly repeatedjunction
is encountered. (iv) A second cleavage (also yielding 3' single-base overhangs) will thenoccur proximal to the first Ubsignal encounteredwithin thatjunction.This cleavageis expectedtoproducea caterminusonthepackagedmolecule andafree(ban,)- terminuswhichmightberapidly degraded
orform the substrate for a newround ofpackaging (intoa
separate capsid).
The directional cleavage model can account for several previously unexplained observations. First, the proposed scanning for a second directly repeated junction (step 3)
accountsfortheobserved cleavageatalternate(i.e., directly repeated) junctions within standard virus concatemers. Fur-thermore, the polarity requirement for the two
prospec-tive termini has also been demonstrated in studies with constructed defective genomes, supporting a scanning
mechanism (9; L. P. Deiss and N. Frenkel, manuscript in preparation). Second, the proposed specificities ofjunction cleavages and termini degradation account for the produc-tion of two a-containing termini (with 3' overhangs [20]) from single a-containing junctions (7). Furthermore, the modelaccountsfor thepresence ofa variable numberofa
sequencesat the Lterminus, andasingle a sequenceat the S terminus. Third, the predicted generation of ba termini from
ba,c
junctions in the majority(thoseresulting with first encounterin L[ca.80%]) of first cleavages mightaccountfor theobservedpredominance ofLterminicarrying asingle a sequence(15). Moreover, itmight constitutethemechanism forthediminutionof
a sequencesfrom multiple a-containing junctions arisingas consequenceofgenome circularization, following virus entry into the cells (21, 22). Finally, theproposed roles for the Ub and Uc elements in cleavage-packaging might account for the distinct propagation pat-terns ofamplicon constructs tested in the present study. Specifically (Fig. 8 a through e), the model predicts that
defective genome concatemers arising by replication of Ub--Uc+ constructs (Fig. 8a) could nevertheless undergo
thefirstcleavageatthe Ucsignal, unlike theirUc- counter-parts. The resultant terminus, which carries the Ub- a sequence, might undergo an aberrant fusion with the S terminus of the helper virus DNA (Fig. 8b), yielding
chi-mericjunctions, afterrounds ofhomologousrecombinations
and replication (Fig. 8 c through e). Finally, the resultant concatemers mightserve as substrates for thenormal cleav-agepackagingprocess, with the first(Ucproximal)cleavage
occurring within a Ub--Uc+
junction,
followedby the sec-ond (Ub proximal) cleavage within a chimericjunction
present in the sameheteroconcatemer.
While the directional packaging model may account for VOL. 59,1986
on November 10, 2019 by guest
http://jvi.asm.org/
614 DEISS ET AL.
a. _ lori I lUcI lori I lucI Iori I IUcI ,
c
icleave b. Ior' lucIUc Ior I I Ub U
detectivegenome termi,ius e v helper S terminus
f
tuse
C _
A2jLLJY.ori
| | I .I ori I UcLUb Uc4 recombine-replicate
d. ri I lucI or I ori luc
:... ...t
recombine c. *j orilI lUcI Iori I IUcIUb Uci Ijori luc
4 2ndcleavage 1stcleavage
cleave/Package
L-Sjunction
a a
Ub
....---
cU Uci or l helperUb---~.~.---
.*-recombine
... ...ile detecti,
iori UcI oriI cUcTl or'i iuci
g. _--..uclori IUcUi
...t recombine h. IoritI 'UctUb
Uc
1junction 2
loriI Uc II IOri alitiction
I
aamplificationIUc I
i.eori UcIU- ijunction 1
|I I ___- - junction 2
Iresolution
J. I orII lUclUb Uc' I ori I Uc lUb Uci
cleave 4 cleave
cleave/package
FIG. 8. Generation and cleavage oftheheteroconcatemers from Ub--Uc+constructs. (a)through(e)Chimeric junctionsare gener-ated and cleaved by the directional cleavage mechanism. Concatem-ersarising by replication of the Ub--Uc+ seedconstructs(thin line inpanela) arecleavedattheDRI proximaltotheUcsignal by the firstcleavagestep.The generatedterminus, carryingUc,fuses with the S terminus of standard virus DNA (thick line in panel b). Homologousrecombinationatthe ori-1 region followed by rolling-circle replication of the resulting circular repeat generates heteroconcatemers (panel d). Homologous recombination in theUc elements within a large chimeric repeat and an authentic repeat along the same concatemer results in the formation of chimeric junctions (panele)containing the inserted helperasequence, with the last portions of the Uc sequence derived from the seed con-struct.Cleavage of theheteroconcatemers occursby the directional cleavage mechanism, thefirstcleavageoccurring withinthedeleted ajunction (atthe Ucsignal) and the secondcleavage occurringinthe chimeric junction (at the Ub signal). The model predicts that concatemersderived byreplication of theUc-constructswillnotbe cleaved instep (a). (f) through(j) Generation and cleavage of the chimeric repeats by the double-stranded break-gap repairmodel. Homologous double crossover recombination occurs between helpervirus DNAwith a double a L-Sjunction (heavyline) anda
defective viralgenome (thinline) whichwasgenerated by replica-tion oftheinput Ub--Uc+construct(panel f).Thecrossoverpoints are in the homologous Uc and ori-l' regions, yielding the heteroconcatemer shown (panel g). Achimericjunctionarises by homologous recombination betweenthe Uc sequences in an authen-tic repeat and the Uc segment intheinserted helpersegment. The chimericjunction contains the deleted a andthe intact inserted a carrying aUcelement which isderived in partfromthe construct (panel h). The heteroconcatemer is a substrate for the cleavage-packaging process, occurring by the double-stranded break-gap repairmechanismdepictedinFig.9. In the aamplificationprocess the Ub--Uc+ junctions is used as junction 1 and the chimeric junction isused asjunction2. ThesequencevariabilityinUbarises during the resolution ofthe Holliday (11) structures. This model
many of the previous observations, it suffers from two drawbacks. First, as recognized by Varmuza and Smiley (35), the modelpredicts the formation of termini devoid of a sequences. Such termini have not been observed in total nuclear DNA (7, 20). Therefore, it must be assumed that
they are rapidly degraded. Second, although the model explains some of the structural features of the chimeric
junctions, it fails to explain the observed variations in the
Ub-DR1 portion of the inserted a sequence. It appears unlikely that such variations could result from the fusion of the defective genome Uc terminus with the S terminus of standardvirusDNA.
Double-strand break and gap repair model for a amplifica-tion.The secondmodel proposed for the cleavage-packaging process is based on the double-strand break and gap repair
mechanism proposed by Szostak et al. (33) to explain
recombination events resulting in gene conversion. Specifi-cally, the model involves the interaction of two directly
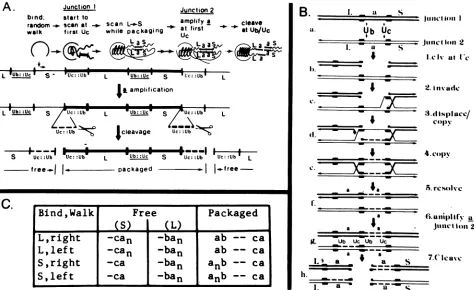
repeatedjunctions (Fig. 9; junctions 1 and 2), resulting in a
amplification by a gene conversion-like mechanism. The resultant junctions are then cleaved to yield the genomic
termini. Cleavage-packaging thus includes the following steps.(i)Asin thedirectionalcleavage model, the packaging
complex traverses along concatemeric viral DNA, starting
from anypoint in the Sor L components, until the first Uc
signal is encountered injunction1 (Fig. 9A). (ii) Scanning-packaging begins, proceeding in the L-to-S direction, until thedirectly repeatedjunction2, along thesame concatemer,
is encountered. The twojunctions are juxtaposed for the nextseries ofevents(depictedschematically in Fig. 9B). (iii) AUcsignal-directed double-strandedcleavage occurs within the DR1 elementof either junction,e.g., injunction 1,(Fig.
9B, step b). (iv) The resultant 3'-terminated strand invades
the homologous sequence injunction 2 and is extended by
copyingthe a sequenceinthatjunction,whiledisplacingthe
equivalent strand (steps c and d). (v) The displaced strand
fromjunction2 serves asthetemplatefor repair synthesis of
the second strand of junction 1 (step e). (vi) The process
terminates by the resolution ofthe two Holliday structures (11);this is predictedtooccur at or neartheDR1sequences (stepf). (vii)Ifjunction2containedasingleasequence(with Uc+ element), it too will become a recipient for an a sequence fromjunction 1 in asimilarsequence ofevents as
above.Theentireprocess thus results in theamplification of
the a sequence within each of the twojunctions (step g). Each ofthedoubleajunctionsnowcontainsaDR1 sequence flanked by the Uc and Ub signals. (viii) Following the a amplification, both double ajunctions arecleaved
(generat-ing a 3' single-base overhang) between the newly inserted
and the original a sequence (step h). This cleavage may
require a structure in whichthe DR1 element is flanked by
Ub and Uc
signals,
anarrangementwhichis presentonly
in the double ajunctions. The model predicts that ca.80%
of thepackagedgenomes (initialencounter inL)will carry ba and ca termini, regardless ofthe number ofa sequences in the cleaved junctions, whereas the remaining packagedgenomes (initial bindingin S) will be terminated withcaand
bansequences
(Fig.
9C). The termini ofthegenerated
free molecules(predictedtobeofthe typebanand ca, or ca;Fig.
9C)might eitherbecomedegradedor serveastargetfornew packaging cycles.
predicts that constructs containing the Uc+ repeats will form heteroconcatemers which willnotbecleavedandpackagedowingto
thelackof Uc element injunction1.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.612.67.296.71.392.2]HERPES SIMPLEX VIRUS CLEAVAGE-PACKAGING 615
Junction i Junction 2
bind; start to amplify c
random + scan at + scan L-a.S at fi a cleave walk first Uc while packaging Uc atUb/Uc
Las L aL a
L gUb:cI 'S- W--c IUw 'UbUcI UC: :Ub" ' L
I
I amplification~~p
L~~~
a apL*Ub::Uc S /UC::Ub L
Uc:: Ub -6
--I
Uc::bS Uc: :Ub Uc: :Ub L
'Ub: :Uc S /Uc: :Ubw L
Icleavage
Uc: :UbUbUb::.Uc-
=-Uc:
:UC 5 Uc: :UbI
UJc:-f
:UbbI Lfree-I
|. packaged , |_treeC.
B.
I._u
asa. Ub Uc
,4 ,_
-~~~~--- 11111 1i<112
_. _ -a s
4 l.dvU%t U*c
4
X
I-
W.c
rDc_ixz
'.____________4
4. copy
C.ZD
_-a _
_a
1
a - 6.u7iiillf
a a . jUiticIti
_Ik
I. Ub Uc Ub Uc
1.*a -a
1.-l,== -
e-11. _ _ _
II.
I a
-FIG. 9. Thedouble-stranded break-gap repair model fora amplification. (A) Basic features of the modelare shown fortwo singlea
junctions. Following randombinding and random walk, packaging beginsat the first Uc sequence encountered, proceeding in anLto S
direction, untiladirectly repeated junction is encountered. Thetwojunctions interact bythedouble-strandedbreak-gap repair mechanism (detailed in panel B), resulting inreciprocal copying ofanasequencefromonejunctiontotheother, theainsertionsoccurringatathe first Uc signal encountered in both junctions. Thegenerated doubleajunctionsarecleavedwithintheDR1elements flankedbythe Uc and Ub
signals.Unpackaged moleculesareeitherdegradedorutilized inanewroundofpackaging.(B) Proposedstepsinthe double-strandbreak-gap repair model foraamplification, featured after thegeneconversionmodel of Szostaketal.(33). (a)Twodirectlyrepeatedsingleajunctions
(junction 1, heavy line; junction 2, thin line)arejuxtaposed. (b) A double-strandbreakoccursattheDR1eIenient proximaltothefirst Uc
signal. (c)The resultant 3' terminus ofjunction1invadesjunction 2,displacingthehomologousstrand.(d) The invading3' terminus has been
extended(dotted line) bycopyingthecomplementary junction 2 strand;thedisplacedstrandpairsin theDR1regionwith thesecond cleaved
strand ofjunction 1, forming atemplate for extension of this strand (dotted line). (e) The newly synthesized portions join the original corresponding strands, forming the Hollidaystructures(11). (f)Resolution has occurred within theDR1elementsoradjacentsequences.(g)
Asimilar series ofsteps results in theamplificationof the asequence injunction 2. (h)Cleavage producing3' single-base overhangs has
occurred within theDR1 elements, separatingthenewandoriginalasequencesin eachjunction.Thiscleavagecould be directedbythe Ub andUcsignals which flank the cleavage site. Smallarrowsindicate the 3' end of each strand.(C)Terminiproducedinpackaged and free
moleculeas consequence ofcleavage-packaging starting from each of the four initialbinding permutations. nrepresents the number ofa
sequencesin thejunctions whicharebeing cleaved.
Unlike the directional packaging model, the double-strandedgaprepairmodel doesnotpredictthegenerationof termini devoid of a sequences. Furthermore, multiple-a-containing S termini have been observed in stocks contain-ingveryhigh ratios of defectivetohelpervirusgenomes (7), supporting the hypothesis that they represent intermediary products of the packaging process. Inaddition, the double-strandedbreak-gap repairmodelaccountsfor thesamesetof observations explained by the directional cleavage model, including the cleavage of alternate junctions in standard virus DNAand thegeneration of packaged DNA molecules terminated with a single a sequence at the S end and with predominantly singlea(but also multiple a)sequencesatthe Lends.Finallyitalsoprovidesaschemeforthepropagation
of the Ub--Uc+ construct and explains the structural fea-tures of the resultant chimeric repeats (Fig. 8f through j). Briefly, this scheme involves homologous recombination between helper virus DNA and the replicated
Ub--Uc+-defective genomes (Fig.
8f),
followed by intramolecular recombination between the created recombinant junction and an adjacent deleted junction in the same concatemer (Fig. 8g).This lastrecombinationcreateschimericjunctions with the inserted Justin a sequence carryinga Uc elementderived in part from the F construct (Fig. 8h). The heteroconcatemer is then cleaved-packaged by the double-stranded break-gap repair mechanism (Fig. 8i), with the
sequence variation in Ub arising as a result of slippage duringthe resolutionof the Holliday structures (Fig. 8i and j). In contrast, heteroconcatemers, which are predicted to ariseby similarrecombinational events withUb+-Uc-
con-structs, would notbecome cleaved and packaged owingto
the lack of Uc element in junction 1. Thus, unlike the directional cleavage model, thepresent model requiresthat bothjunctions (which are cleaved to generate the termini) containthe Uc signal.
pac-l and pac-2 homologies. The proposed role of the
A.
1iCti(ltioIl I
2.Invadc
3d( ISI)lIuC/
3cdislacc
Bind,Walk
Free
Packaged
(S)
h
(L)
L,right
-can
-ban
ab
--
ca
L,
lef
t-can
-ban
ab
--ca
S,right
-ca
-ban
anb
-- caS,left
-ca
j
-ban
anb
-- caI.resolve
u
.n2
VOL.59, 1986
1-ip--S.- s
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.612.71.545.73.363.2]a a
Ub
---Uc
DR1
Ub
---Uc
---
pac-2*
v
*pac-1---B.pac-2
ICGmotif
F
Justin
KOS 17 USA-8
HG52
vzv
EBV HVS 11
BHV-l
.... CGCCG;CG
.... CGCCGCC .... CGCCGCC .... CGCCGCC .... CGCCGCC .... CGCCGC( .... CGCCcC( ....gGCCGCC
. CcCCGaC
....CGCCGC(
HSV-1 (F) HSV-1 (Justin) HSV-1 (KOS) HSV-1 (17) HSV-1 (USA-8) HSV-2 (HG52)
VzV
EBV/Namalwa HVS (11)
D.
S S S
S
S S S
s
BssHII
1--
Bt9HII
T.
--I I ---I
cgcgcgcacgccgcccggaccgccgcccgcc gcgcgccgccc ; cgcgcgcacgccgcccggaccgccgcccgcc =.TTA gcgcgccgccc cgcgcgcgcgccgcccggaccgccgcccgcc gcqcgcgcgcc ; cgcgcgcacgccgcccggaccgccgcccgcc TTTTP gcgcgcgcgcc
cgcgcgcgcgccgcccggaccgccgcccgcc TTTT gcgcgccgcc B ggctgccttccgcggcgcccccgcgcggc =TLT cccgcgcccg ; caaacgcgcggggaggtggggtcgc ¶TTTTTPTT ctctctcgagc B ggggccagccacgccccctccac TTT ccaggaatgcc B ctcgctctagccacgcccaggaca T¶TT ccagctgccci
B cagagaaaa N=1T ctccgcgcccc
--- S terminal region -Cn-ri----II---
I
DraI SmaI
gcgccgccgcgctttaaagggccgcgcgcga CCCCC GGGGGG
gcgqccgccgctttaaagggccgcgcgcga CCCCC GGGGGG
geccgccgcccgcctttaaagggccgcgcgcga CCCCC GGGGGG
jgcccgccgccaccgctttaaagggccgcgcgcoga CCCCC GGGGGG
gEcccccgccgcgctttaaagggccgcccgcga CCCCC GGGGGG
ctcccgccccgcctcttttcccccggggagtcaa CCCCC GGGGGGGG
cctctcccqgggtccgccgggcgcccagaaa CC GGGGGGG ccccccatgccctctccaacaccccccgaag CCCCCC GGGGG
ccccttccccctctttqcctaccaaqttat CCCCC GGGGGG
L Terminal region ---I end SiaI
ccgcccgcggggggcccgggetgc L
cgccgcggggggcccgggct3c L
gcgcccgcggggggcccgggctgc L ;cgcccgcggggggcccgggctgcc L
ccgcccgcggqgggcccggqccgEc L cccccgcgcggcaggacggggactcccgccc L
ggqgccgcgagagggctggcctcc L gcggcggcccatgctccccccatgc
igcqcccactgcttggggcccc
ccccgcgqggggqctgggcc L
IT motifi I
---GnI---Apal
TGTGTTTC! GGGGGGGG cccgt ....
TGTGTTTC GGGGGGGG cccgt ....
TGTGTr rGGGGGGGG cccgt ..
TGTGTTTT GGGGGGGG cccgt ....
TGTGTTTT GGGGGGGG cccgt
TTTTTT GGGGGGGGG cgcga ....
TTATTTTC GGGGGGGGG tccga TCTTTCCT GGGGG ccttt ....
AAAATCAGT GGGGG ctgca
E.
summary:
~~~~~~~~~~~~~DRi
DR1 Ub ,
(DR2)22
(D~R4)3
UcDR
[ III
---IIN
I {1A A
[1
It- -- -
---
Construct D-1Construct H-l
pac-1
I 100 bp
pac-2
Homology
FIG. 10. pac homologies in different herpesviruses. (A) Alignment ofthe pac-J and pac-2 sequences flanking the cleavage site. (B)
Conservedpac-2 (Uchomologous) sequences(capital letters) foundatconcatemeric junctions of variousherpesviruses. References for the
nucleotide sequences arelisted in panel D. Cleavage sites reportedinthe literature areunderlinedatthe end of thesequences shown.The Epstein-Barr virussequence shown is that of the integratedgenomeinthe Namalwa cell line (16). Hence nocleavage (underlined) site is
indicated. (C)Conserved pac-J (Ub homologous) sequencesfound at concatemericjunctions of various herpesviruses. (D) Summary of
distances fromthe Tmotifinpac-I (arrowinpanelA)andT,, element in pac-2 (arrowinpanel B)tothe respectivegenomic termini, and the
percentG+Cresidues inthe Sand L terminalregions.TheaverageG+Ccontentof therespectiveviralgenomes aretaken from reference
26.(E)Deletionsdefinedbythe D-1(Ub)andH-1(Uc)constructsoverlapthe conservedpac-J andpac-2sequences.TheApaI (A)andBssHII
(B) sites which define thedeletions areindicated.
616
A.
S terminus/pac-1 L Terminus/pac-2
Virus Reference Distance %GC Ster Distance %GC Lter Ave %GC end + T region T + end region qenome
F Mocarski and Roizian, 1982 41-42 83 33-34 97 67
Justin Mocarski et al., 1985 40-41 83 31-32 97 67
KOS Varmuza and Smiley, 1985 43-44 84 33-34 97 67
17 Davison and Wilkie, 1981 44-46 80 33-35 97 67 USA-8 Davison and Wilkie, 1981 42-44 84 33-35 100 67
HG52 Davison and Wilkie, 1981 44-47 79 30-41 90 69
VZV Davison, 1984 41-42 83 33-34 76 46
EBV Matsuo et al., 1984 ? 79 ? 74 59
HVS 11 Bankier et al., 1985 40-41 66 31-32 78 46
BHV-1 Haamerschaidt etal., 1986 - - 30 93 72
B B
-2 I
ommmmmmmmmmmMME IL