0022-538X/89/094101-06$02.00/0
Copyright ©
1989, AmericanSociety
for MicrobiologyComplete Nucleotide Sequence of the Gene Encoding
VP4
of
a
Human Rotavirus (Strain
K8)
Which
Has
Unique
VP4
Neutralization Epitopes
KOKI
TANIGUCHI,'t*
KAZUO NISHIKAWA,' TOMOKO URASAWA,' SHOZOURASAWA,'KAREN MIDTHUN,3 ALBERT Z. KAPIKIAN,' AND MARIOGORZIGLIA'
Laboratory of Infectious
Diseases, National Institlute of Allergy and Infectious Diseases, Bethesda, Maryland 208921;Department
of
Hygiene,Sapporo Medical College,
Sapporo 060,Japan2;
and Department of International Health,Schoolof Hygiene and Public Health, Johns Hopkins University, Baltimore, Maryland212053 Received 21 February 1989/Accepted 5 June 1989
Inourprevious study (K. Taniguchi, Y. Morita, T. Urasawa, and S. Urasawa, J. Virol. 62:2421-2426, 1987) inwhich the cross-reactive neutralization epitopesonVP4of human rotaviruseswereanalyzed,onestrain, K8, was
found
tobear unique VP4 neutralization epitopes. This strain, which belongstosubgroupIIandserotype1, wasnotneutralized by anyof six anti-VP4 neutralizing monoclonal antibodies which reacted with human rotavirus strains ofserotypes 1, 3, and 4 or serotypes 1 through 4. We determined the complete nucleotide
sequenceof thegene
encoding
VP4of
strainK8
by primerextension.
TheVP4geneis2,359
basepairs in length,with
5'
and3'
noncoding regionsof 9 and 25 nucleotides, respectively. Thegenecontainsalongopenreadingframe of
2,325 bases capable of coding foraproteinof 775 amino acids. When compared with those of other human rotaviruses, VP4of
strain K8 hadaninsertionofoneaminoacidafter
residue 135,asfound in simianrotavirus
strains, and in addition, it hadadeletionofoneamino acid(residue 575). The amino acid homologyof VP4 of strain K8 and those of other virulent human rotaviruseswas
only
60to70%. Thiswasunusual,since over90% VP4homology
hasbeen found
amongthe othervirulent
humanrotavirus strains. Incontrast, the VP7amino acidsequenceof
theK8
strainwasquite similar (over98%
homology)tothoseof
otherserotype1 human rotaviruses. Thus, theK8
strainappearstohaveaunique VP4 genepreviously notdescribed.The
rotavirus virion contains
1idouble-stranded
RNA segmentsenclosed within the inner shell of its double-shelled
capsid (14). Two capsid proteins,
VP4 andVP7,
areconstit-uents
of the
outercapsid, and they
areindependent
neutral-ization antigens (9,
14,22).
VP7encoded by
RNAsegment8
or9 definesserotype
specificity (8, 12).
Theproduct of
RNA segment4, VP4, is responsible for growth restriction of
fastidious rotavirus strains
incell culture
andfor
protease-enhancedplaque
formation(2, 3, 11).
VP4 is also associatedwith rotavirus virulence. In
amurine model, the
geneencoding
VP4determined the dose of SA-11 and NCDV
rotavirusesrequired
to induceexperimental gastroenteritis
(23). Also, in humans,
adimorphism of VP4
sequenceis
found between virulent and
asymptomatic
strains(5-7).
Moreover,
VP4 appears to beresponsible
for somecross-neutralization
orcross-protection
among serotypes(15, 27,
28, 36-38).
We
previously prepared
six anti-VP4neutralizing
mono-clonal antibodies
(N-MAbs)
reactive with human rotavirus(HRV)
strains ofserotypes1through
4orserotypes1, 3,
and 4(28, 30). By sequencing
the VP4geneofantigenic
mutants resistanttoeachof
the sixN-MAbs,
weidentified the aminoacid residues involved in cross-neutralization
(27).
How-ever,one HRVstrain
(K8)
didnot react withanyof the sixanti-VP4
N-MAbs, although
it was neutralizedby
each offive
anti-VP7
serotype1-specific N-MAbs directed against
differentepitopes (21, 26).
These resultsimplied
that strain K8 has aunique VP4, prompting
usto characterize its VP4gene
by
sequenceanalysis.
In this report, we present the* Correspondingauthor.
tPresent address: Department of Hygiene, Sapporo Medical
College, South-1, West-17, Chuo-ku. Sapporo 060, Japan.
complete
nucleotide sequence of thegeneencoding
VP4 of strain K8 andcompared
it with those of other human and simian rotavirus strains.Strain K8wasderived fromastool
sample
fromaschool-boy
of 14yearsofage,who became illduring
anoutbreak ofgastroenteritis
which occurred inKitami, Hokkaido, Japan,
in 1977. The outbreak affected over53people,
of whom 39(73.6%)
wereover6yearsofage.The viruswassuccessfully
cultivated in cell culture
(34). Subsequently,
the virus wasgrown in MA-104 cells in the presenceof
trypsin (1 ,ug/ml)
and
purified by
fluorocarbon treatment and CsClgradient
centrifugation.
Viral mRNAwassynthesized
in vitroby
theendogenous transcriptase
presentinviral cores obtainedby
treatment with EDTA.
Synthetic oligonucleotide primers
were
used
to sequence mRNAby
reversetranscriptase
inthe presence of
dideoxynucleotide
as describedpreviously
(7).
The 3'-terminal 200 nucleotides of the viral RNA weredetermined
by using
denatureddouble-stranded virion RNA. Inourprevious
studies(28, 33-35),
strain K8 wassero-logically
characterized asbelonging
tosubgroup
IIby
the use of anti-VP6subgroup-specific
MAbs and to serotype 1by
neutralization andanenzyme-linked
immunosorbent as-say(ELISA), using polyclonal
antibodies orN-MAbs,
re-spectively. However,
it did not reactby
neutralization orELISA with six anti-VP4 cross-reactive N-MAbs that
rec-ognize
VP4s of serotype 1 strains(28).
Figure
1 shows thecomplete
nucleotide and the deduced amino acidsequence of the K8VP4gene.Thesequencehasa
single long
openreading
frameof2,325 bases,
whichbegins
with ATG atposition
10 to 12 and terminates with TAG atposition
2335 to 2337. The first ATG conforms to the consensus sequence for strong translation initiation(AN
NATGG) (16). By
direct amino acid sequenceanalysis
of 4101on November 10, 2019 by guest
http://jvi.asm.org/
12 24 36 48 60 72 84 96 108 120
GGC TAT AAA ATGGCT TCt TTA ATT TAT AGA CAGTTA TTA TCA AACTCA TATGTT ACAMC ATC TCT GAC GMGTT MT GMATT GGAACTAAA AMACA ACT MCGTT ACT GTTAATCCA
KET Ala Ser LeulieTyr Arg GinLeu Leu Ser Asn Ser TyrVat ThrAsn lieSer AspGiuVat AsnGtulIeGly Thr Lys Lys Thr Thr AsnVat ThrV&l AsnPro
132 144 156 168 180 192 204 216 228 240
GGG CCA TTC GCA CAA ACG GGA TAT GCG CCT GTT GAC TGG GGA CAT GGT GM TTGCCT GAT TCTACA TtGGTGCAG CCA ACT CTTGAT GGT CCATACCAA CCC ACC ICACTCAACTTACCA
GlyProPheAiaGin ThrGtyTyrAlaProVat Asp TrpGlyHisGtyGiuLeu Pro Asp Ser Thr LeuVat GinPro Thr LeuAspGtyPro Tyr GinPro ThrSerLeu AsnLeu Pro
252 264 276 288 300 312 324 336 348 360
GTT GATTAT TGGATG TTA ATT GCG CCT ACT AGA GM GGA AAA GTT GCT GAAGGT ACAAAT ACG ACT GACAGA TGGTTCGCT TGTGTACTA GTT GMCCAMTGTGCM AAT ACACAAAGA VatAspTyr Trp KET Leu lIe AlaPro Thr ArgGtuGlyLysVai AiaGtuGly ThrAsn Thr Thr Asp Arg TrpPheAla CysVai LeuVat GtuProAsn Vat GinAsn ThrGinArg
372 384 396 408 420 432 444 456 468 480
CAATAC GTA TTAGATGGG CMMTGTCCM TTG CATGTC TCAAAC GAT TCAAGT TCG TGG AM TTT ATA TTA TTC ATT AMA TIGACGCCT TATGGAACG TACACT CAATAT TCA ACA GinTyr VatLeu AspGlyGinAsnVat GinLeuHisVat SerAsn Asp Ser SerT Ser IrpLysPhe lte Leu Phe lteLys Leu Thr ProTyrGlyThr Tyr ThrGinTyr SerThr
492 504 516 528 540 552 564 576 588 600
TTA TCA ACG CCG CATMG TTGTGC GCG TGGATGAAA AGA GAT AAC AGA GTG TAC TGG TAT CAAGGA GCGACACCGMCGCA TCAGAG AGCTAT TACTTG ACC ATAMCAAT GAT AAC AGC Leu Ser ThrProHis LysLeuCysAla TrpMET Lys Arg Asp Asn Arg Vat TyrTrp Tyr GinGtyAla Thr Pro AsnAlaSerGtuSer Tyr Tyr LeuThrlieAsn Asn AspAsn Ser
612 624 636 648 660 672 684 696 708 720
MC GTt TCAAGTGACGCT GMTTt TAT TTAATACCGCAA TCG CAGACC GCT ATGTGT ACA CMTATATA AACMTGGT TTACCA CCA ATT CAGMT ACCAGGAAT AtT GTACCAGTAAAT AsnVatSer Ser AspAlaGtuPhaTyrLeuliePro GinSerGinThr AlaMETCys Thr GinTyrlIeAsnAsnGlyLeu Pro Pro Ite GinAsn Thr Arg AsnIteVat ProVat Asn
7321 744 4 756 768 780 792 804 816 828 840
AlT ACATCT AGACAGAtT AMA GAC GTA AGAGCT CAGATGMT AACGACAlAGIAATA TCA AAA ACCTCGTTA TGGAAACAA ATG CAA TATAAT AGAGAT ATAATCATTAGATTTAAA TTT lieThrSerArgGinIle LysAsp VaL ArgAla Gin MET AsnGtuAsp IteVat IteSer Lys Thr SerLeuTrpLysGlu MET GinTyr Asn Arg Asp lte IleIteArgPheLysPhe
852 864 876 888 900 912 924 936 948 960
GCTAATTCA ATA ATCAM tCAGOT GGA CTA GGT TAT MAG TGG TCGGAA ATA TCATTTMG CCCATGMC TAT CAA TAT ACGTAT ACGAGAGAT GhA GMAGAM GTAACAGCA CATACA ACA AlaAsnSer Ilt lIeLys SerGtyGlyLeuGly TyrLysTrpSerGtu lte SerPheLys ProMET AsnTyrGinTyr Thr Tyr ThrArgAsp GluGCuGtu Vat ThrAlt His Thr Thr
972 984 996 1008 1020 1032 1044 1056 1068 1080
TGT TCA GTT GGT GOT GTCMtGAT TTT MTTAT AATGGAGGt ACG TtG CCT ACT GAT TTT GCA AtA TCGAGA TTT GAGGTC ATAMOGM Mt TCT TAT GTA TAT GTA GAT TAT TGG GAT
Cys SerVat GlyGlyVatAsnAspPheAsnTyr AsnGiy Giy Thr LeuPro ThrAspPh.Ata IteSerArgPhe Gtu Val lteLysGtuAsn Ser TyrVat Tyr Vat Asp TyrTrpAsp
1092 1104 1116 1128 1140 1152 1164 1176 1188 1200
GAT TCACAAGCA TTT AGAMTATG GTATAT CAAGOT ACG TtAOCTGCAAAT ITGAAT GATGtAGTA TGCAGT GGA GOTtCt TACAOT TTT GCG TtA CCTGTA GGCAAT CAT CCGGtGATA
Asp 5er CGnAta PheArg AenMET Vat TyrGiuGty Thr LeuAlt AteAsnLeuAenAepVat Vet CysSerGty GtySer Tyr SerPheAteLeu ProVet GiyAsnHNI ProVet lte
1212 1224 1236 1248 1260 1272 1284 1296 1308 1320
GATGOT GOT GCAGTGACC TTAACA TCTGCT GGC GTAACACTATCA ACCCAGTAT ACAGAT TAT GTA TCATTAMTtCATT1 CMTTCAGATtCAGA TtGGCGGtGAGCCAACCATCA TTt AspGtyGlyAteVet ThrLeuThrSerAteGtyVat Thr Leu SerThr GinTyrThrAspTyr Vat SerLeuAsnSer LeuGinPheArgPheArg LeuAti Vat SerGtuProSer Phe
1332 1344 1356 1368 1380 1392 1404 1416 1428 1440
tCC ATCTCG CGAACTAGAAtGAOtGGCAtATAT GGA TTA CCA GCTGtAAAtCCA AATAATAGC GCACAATAT TAT GAGATA GCT GGtAGATTCTCA CTC ATATCA CTAGTACCAACAMT Ser IleSerArg Thr ArgMET SerGtylieTyr GlyLeu ProAta Vat Asn Pro AsnAsnSerAta Gtu Tyr TyrGtu It.AiaGtyArgPhe SerLeu lteSer LeuVat ProThrAsn
1452 1464 1476 1488 1500 1512 1524 1536 1548 1560
GAT GACTATCAA ACG CCA ATC GCTAATtCAGTTACCGTAAGA CM OAT TTAGAGAGACAA TtA GGCGAG TTAAGAGMGMA TTTMTTCGTTG ICA CMGAAAtAGCT GTT TCCCMCtt
Asp AspTyrGinThrProIteAlt Asn SerVatThrVat Arg GinAsp LeuGluArgGin LeuGtyGluLeuArg Gtu GtuPheAsnSerLeu SerGinGlu lieAlaVatSer GinLeu
1572 1584 1596 1608 1620 1632 1644 1656 1668 1680
AtA OATCtA GCAACATTACCGCTt CATAtG TICTCAATG TTC TCT GGA ATA AMtCAACG GTA GAGGCA GTAAAATCT ATGACTACGMTGIGATG AAAAGATtTAMACATCAAOT TTA IteAspLeuAlaThrLeu ProLeuAsp METPhe SrMET Phe SerGty lie LysSer ThrVat GtuAteVat LyeSerMET Thr ThrAsnVat MET LysArgPhe Lys thrSer SerLeu
1692 1704 1716 1728 y 1740 1752 1764 1776 1788 1800
GCAMCGCC AtAtCT GAT TtAACAAOT MTATG TCGGMGCG GCA tCA TCT GTAAGA TtOACG tCA GIG AGAtCG 0tGGGCACT Att ACA T11 CCA AGA GCT AGGGTT TCA TTGCMGIG
AlaAsnAta IteSerAspLeu ThrSerAsnMET SerGtu Ate Ala Ser SerVat ArgLeu ThrSerVei Arg Ser VatGty Thr Ile Thr LeuProArgAtaArgVaL Ser LeuGin Vat
1812 1824 1836 1848 1860 1872 1884 1896 1908 1920
GGC GAT GACTIGAGOtCCATGCAAGAC GTATCA ACACMGTGtCAMT GIGAGT AGAMT TtAAGA TtGAM GAG TtCACG ACd CMACTGATACT TTAAGC tTT OAT GACATC TCTGCA GiyAspAspLeuArgSerMETGinAspVetSer ThrGinVat SerAsn VatSer ArgAsnLeuArg LeuLyeGtuPheThr Thr Gin Thr AspThrLeu SerPheAsp Asp
Ile
SerAte1932 1944 1956 1968 1980 1992 2004 2016 2028 2040
OCTGTATtOAAGACd AMCtAGAt AMATC ACGCM ATT 1CACMCMACAATGCCAGAT ATt ATAOCT GAO tCAtCTGAGMO TlTATACCG AAG AGA TCGTAT AGAATAOTT OATGAG
Atl Vat LouLysThrLysLeuAspLysSer ThrGtn iteSerGinGin ThrMET ProAsp It liteAte Glu Ser SerGluLys PheIte ProLye ArgSer Tyr Arg IteVetAspGtu
2052 2064 2076 2088 2100 2112 2124 2136 2148 2160
GATATTCGATICGAAACT GGAATTGACGGAACGTTt TAT OCT TACAAA GTCOATACA TTt MT GMAATt CCG TTt GAT ATG GAACGATTT AATAMTTA ATAACAGAT tCACCAOTT TTA Asp Ito Arg PheGluThrGty Ite AspGlyThrPhe Tyr Ate Tyr LysVat AspThrPheAsnGtu Itc ProPhe Asp METGiuArg PheAsnLye Leu lIe ThrAsp Ser ProVat Leu
2172 2184 2196 2208 2220 2232 2244 2256 2268 2280
TCAGCAATA AtAGAC ITTtMGACG TTA MGMC TtA AAC GAC AAT TAT GGA ATAACA AAG AMCAA GCCAtG GAA CtATIA CAT TCA AATCCAMO ACATTAAM GAG TTT ATA AAT AAT
SerAteIte IteAsp PheLys ThrLeuLysAsnLeu AsnAspAsn Tyr Gly IteThrLys LysGtnAtl MET GtuLeuLeuHisSerAsnPro Lys ThrLeuLysGtuPheIteAsnAsn
2292 2304 2316 2328 2340 2352
AAT AAT CCAAtAATlAGA Mt AGA AtCGAAAAC TTA ATA TCG CAG tG0 AGO TTG TAG CTG *CTAtt TtAAGA TGTGAC C
AsnAenPro tle IieArg AsnArg lie Gtu AsnLeu lteSrGinCysArg Leu
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Nucleotide and amino acid sequence homologyof VP4 of strain K8 and of other human or simian rotavirus strains
% Homology with VP4 of strain K8 Strain (serotype)
NLIcleotide Amino acid
sequence sequence
Virulent human rotavirus
KU (1) 65.4 64.3
Wa (1) 65.6 64.3
DS-1 (2) 66.3 65.7
RV-5 (2) 65.9 65.2
P(3) 66.0 65.0
VA70 (4) 65.2 63.7
Asymptomatic humanrotavirus
M37(1) 64.4 65.8
1076(2) 64.0 65.0
McN13(3) 64.2 65.7
ST-3 (4) 64.3 65.4
Simian rotavirus
SA-ll(3) 67.6 70.5
RRV (3) 66.8 69.7
SA-11 VP5, two trypsin cleavage sites (arginine residues 241
and 247) have been identified previously (18). These two
arginine residues are also conserved in strain K8.
However,
the
six amino
acids of the connecting peptide of
K8,
i.e.,
amino acids
242 to247, were
quite different from those found
in other human rotavirus strains (Fig. 2). Furthermore, when
compared with other
HRVstrains, VP4
of strain K8 was
found
tohave
aninsertion of one amino acid after residue
135 (as described in two
simian rotavirus strains), and in
addition, it lacked one amino acid at residue 575.
The K8
nucleotide and amino acid sequences were
com-pared with those of several virulent and asymptomatic HRV
strains
aswell
astwo
simian rotavirus strains. The
overall
nucleotide sequence homology
of
VP4of strain K8 and of
other virulent HRV
strains wasonly 65
to67%
(Table 1).
This was in sharp contrast to the
high degree of homology
(90%
or
more)
found among other virulent
HRVstrains (6,
7). In
recentstudies (6, 7), asymptomatic neonatal
HRVstrains
representing each of four serotypes
werefound to
have
ahigh degree
of nucleotide homology (more
than90%)
among
themselves but
arelatively low degree of nucleotide
sequence
homology (68 to
72%)
with virulent strains of
corresponding serotype. The K8 strain also had
alow
degree
of sequence homology (64
to65%) with
theasymptomatic
strains. In
addition,
alow
degree of homology (66
to68%)
between
strain K8
and twosimian rotavirus
strains wasalso
observed. These differences
werealso reflected
atthe
amino
acid level
(Table 1).
Several neutralization
epitopes
onthe
VP8fragment of
VP4have
been identified in
rhesusrotavirus (RRV) (19),
although these sites have not yet been defined
onhuman
strains. These
epitopes
seem tobe
responsible for
strain-specific
neutralization, since aminoacid
divergence
in the VP8region was high among rotavirus strains (6) and the
N-MAbs
directed
against
VP8showed limited
neutralization
of
heterotypic
rotavirus strains (19). The amino acid
se-quence homology in VP8 of strain K8 and of the other
rotavirus strains was quite low (Fig. 2). Compared with the
low
homology of amino acid sequence in VP8 of rotavirus
strains, the N-terminal half of VP5, the larger cleaved
product of VP4, was relatively conserved among rotavirus
strains including strain
K8(Fig. 2). In our previous study
(27), we identified an amino acid critical to individual
cross-reactive
neutralization epitopes on the VP5 fragment of VP4
by
using KU (serotype
1human
strain) antigenic mutants
resistant to each of the
sixanti-VP4 cross-reactive N-MAbs.
At
the
positions where amino acid substitutions occurred in
the
antigenic mutants, there was a correlation between
conservation
of sequence among different HRVs and the
reactivity of
theseviruses with antibodies used for
selection
of
mutants. Aminoacid residue 305 (Leu)
which is found
tobe
critical to epitope
Iwas
different in strain K8 (Thr for
residue 306) from the amino acid in other strains (Fig. 3). In
addition, the amino acid
critical to epitope
IIwas
shown
tobe
position 392 (Ala) or 439 (Leu), but the amino acid
corresponding
tothese positions (393 or
440in strain
K8)
was
different in strain K8 (Asn or Ser). The amino acid
critical to epitope
IIIwas
conserved at position 434 of the
strain
K8. but
anamino
acid sequence difference
atthe
preceding position (433 for strain K8 and 432 for other
humanstrains)
wasfound. Our
previous study (27)
suggested
that
thispreceding residue is also critical
toepitope
III.These
differences
inamino
acid residues may
explain
thelack
of reactivity of
strainK8 with the
anti-VP4
cross-reactive
N-MAbs.
The
gene
encoding
VP7of
strain K8 wasalso
sequenced
by primer
extension.
Thededuced
VP7amino acid sequence
of strain
K8exhibited
anextremely
high
homology
tothose
of other serotype
1 humanstrains;
only
two orfive amino
acid residues
weredifferent between K8 and
KU orWa,
respectively (data
notshown).
Thus,
strainK8, which
shares a VP7similar
tothoseof the
serotype
1viruses, carries
aunique
VP4gene
which has
notbeen
previously
detected. These data
strongly support
theconcept that
VP4and
VP7 haveindependent
neutralization
specificities (9). With respect
toVP7among
HRV
strains,
six
different serotype
specificities
havebeen
reported
previ-ously (1,
10, 20). There have been
only
afew studies
examining the
numberof neutralization
specificities
of
VP4(6.
7,
19.
27). By
serological characterization and amino acid
sequence
analysis,
it
hasbeen shown that among virulent
strains the
VP4sof
HRVstrains of serotypes
1,3,
and
4 arehighly conserved and
thatthere
is anapparent
diversity
between
VP4of serotype
2strains and those
of serotype
1, 3,
and
4 strains(6,
7,
27, 29). Furthermore,
aconsiderably
lower
degree
of
homology
hasbeen
found between
the VP4sof virulent and
asymptomatic
HRVstrains.
Inthis
study,
the VP4of K8
wasfound
tobe
unique. Thus, there appear
toexist
atleast
four
varietiesof
VP4among
HRVstrains:
(i)
virulent strains
of serotypes 1, 3,
and4;
(ii)
virulent
strains of
serotype
2;
(iii)
asymptomatic
strains
of serotypes
1,
2,
3,
and
4; and
(iv)
the
virulent
K8 strainof serotype
1and other
strainshaving
K8-like VP4.Strain
K8 hasseveral other
characteristics
thatfollow.
(i)
In
comparison
withother
HRVstrains,
it grows
efficiently
incell culture
andproduces
relatively large
plaques (34).
(ii)
InFIG. 1. Completenucleotide sequence and deduced amino acid sequence of theVP4geneofstrainK8.Thenumbersabovethesequence refer tothe nucleotide positions. Cleavage sites are indicated by arrows. and the corresponding connecting
peptide
is underlined. The positions of nucleotide insertion and deletion foundinK8 VP4geneai-e shownboxed and indicated by the arrowhead,respectively.
on November 10, 2019 by guest
http://jvi.asm.org/
MASLIYRQLLSNSYrVTNISDEVNEIGTKKTTNVTVNPGPFAQTGYAPVDWGHGELPDSTLVQPTLDG
PYQPTSLNLPVDYWKLIAPTREGKVAEGTNTTDRWFACVLVEPNVQNTQRQYV
120 ---T--SVDLH--IEQ--SE--Q.--- --R----N .IN----T-E-I---TFKPLT--I--NSNTN-V-Y-S--NS-F-T-V-AI--H-IQVD-T
120---T--SVDLH--IEQ--SE--QS---R----N-
IN--T-E-V---TFKP-N---L--SSNTN-V-Y-S--NN-F-T-VIA--H-SQ-N---I
120---T--SVEL
I-T--SE--Q---I---N---VLESW-VN---TIE-V---FKP-S---I-LN--DQQV-L----K--I-I-LL---T-QS---T
120___.---
TVEL--I-T-SE-SQ-1-I---N----VLESW-VN--TIE-V---FKP-S---I-LN--NQQV-L----K-I-I-LL---T-QS--T
120-A---T-TVEL--IQ--ST--Q---N----N--P--TN---T-E-V---TF-P---
L---NA-V-V----N-N--L-TI-I---QVE-T-T 120-T--TVDL
IQ--ST--Q---I-L---N--P--TN---T-E-V---F-P---L---AA-V-V---N---L-TI--VA--TSET-S-T
120LDGQNVQLHVSNDSSTSWKFILFIKLTPYGTYTQYSTLSTPHKLCAWMKRDNRVYWYQGATPNASESYTINNDNSNVSSDAEYIPQSQTAMCTQYINNGLPPIQNTRNIVPVNITS
240 VF-E-K-FN-R--D+K--LFRGSSQNEFYNRR--TSDT--VGIL-YGG-IWTFH-E--R-TTDSSN-A-LNDISIIIHS---
I--R--ESK-NE---V--LSLS-
239-F-E-K-FN-E-N-D>+K---
FEKF-GSSQ-NFSNRR--TSSNR-VGML-YGG--WT
FH-E--R-TTDSSN-ADLN-ISIIIHS---I--R--ESK-NE---V--LSLS- 239-F-ETK-
IT-E-NTN+K--FEMFRKNVSAEFQHKR--TSDT--AGFL-HY-S-WTFH-E--H-TTDs-sS-as-LEVETVIHV
---I-R--ESK-VE---T---M---ALS- 239-F-ETK-IT-E-NTrN+K--FEMFRSSVSSEFQHKR--TSDT--AGFL-HY-S-WSFH-E--H-TTD-SS-S-LSEVETTIHV--I-SR--ESK-VE-
---T---K---ALS- 239-F-QVTr--Q-K---VDLS-Q-QDN-S-HGS-LSTP--YGV--HGGKI-T-N-
E---NTrG--S-T-F-TV-MTAYCD- I--LA-E-K-E---S-V- 240-F-TQE-ITIA-A-Q-Q--DVV-S-QN-S-S--GP-QSTP--YGV--HNGKI-T-N-E
---VTTK--S-T-Y-SV-KTAFCD--I--REEEST-E--- ---LALSA 240QKDV AMEDIVI SKTSLWKEMQYNRDIIIRFKFANSIIKSGGLGYKWSEISFKPMNYQYTYTRDEEEVTARTTCSVNGVNDF YNGGTLPTDFAISRFEVIKENSYVYVDYWDDSQ
S-QYK -V---T ---
C---G---V-L---Y-AA----N-L--G-Q---
N-S----S----SV-Y---K.-QYR- -V----T---G--VL-
---Y-AA----S-S--G-Q---N-S----S---S-Y--Y---.-
I---K.SVTYO+-V----I---C---N---V-L---AA----N-L--G-Q---N-S
----S---SV--Y---SVTYQ -V---I--- ---- ----A
----N-L--G-Q---N-S---L--H-SV--Y--
---.N---N-VYT
--P-Q----V--V---V----AF---G---S---V-KY---F--I---.N-ISH
--A---V---T---S--V---A---G---M---F---S---V---Y---AFRNMVYV.RSIAANWDVCSGGSYSFALPVGNHPVMSGGAVTLTSAGVTLSTQYTDYVSLNSLQFRFRLAVS
EPSFSISRTRMSGIYGLPAVNPNNSAEYYEIAGRFSLI SLVPTNDDY---S-K-T----D-SI---AW--N----S-HF---F-F---R---S-T-D---L---TVNL---A----GN---S---.-.---. S-K-T----N-R----KW-I-N----S-HF
.---FF--F----R---S-T-D---I---TINL---A---GN---MS---Q--- ---S-K----N-N-Q----AW---
S-HF---EF--F---R---S-T-E--P-L---V--L---F---SGH---F-L---S----
-L---S-K----N-N-Q----AW---S-HF---KF--F---R---S-T-E--P---L---V--L---S---SGH---S----
R--.---D--S-M-T--D---Y---T----S-H---F--F---R---S-E--P---L---V--L---AK---Q---L----.---
-S-I-T-D---QW--T----S-H---F--F---R----T-E---T---V-RL---A----GK----V
.----0
QTPIANSVTVRQDLERQLGELREEFNSLSQEIAVSQLIDLATLPLDMFSMFSGIKSTVEAVKSMTTNVMKRFKTS
SLANAISDLTSNMSEAASSV+RLTSVRSVGTITLPRARVSLQVGD ---M--- ---TD---M---L---EL---IDLT---A-S--K-RK-K--TS--EM-HSL-D---AS-SV-I-NISTISNWTNND-SN
--- ---M---L.---.IDLT--A-S---K-RK-K--TS--EM-NSL-D----.AS-SA-.INLSTISNWTNT-KS-SN --K--- ----D---MT---L---ID-A--A-K-- K--R-G--TS--E-GSL-N- - --IS-SS-I--NISSISVWTD-E-IAG---M-I-- -
--D---IT---L.---A---A----A-K--R-G--TS--E--RSL-N----S-SS-INISSISEWTD--E-IAG
-M---
---D---N---Q--M----L---
---ID-A---A---K----SV-T--DSL-D----IS-SA--SSTASAWTE--NIAS---T---A----M---L---
ID-A--A-S--K-K-G---SV-T--DSL-D----IS-GA-I---SSASAWTD-T-IT-!360:359 ;359 *359 359 360 360
480 479 -479 *479 .479 -480 -480
)599 f599 599 ;599 ;599
-600
-600
DLRSMQDVSTQVSNVSRNLRLKEFTTQTDTLSFDDISAAVLKTKLDKSTQISQQTMPDIIAESSEKF IPKRSYRIVDEDIRFETG IDGTFYAYKVDTFNEIPFDMERFNKLITDSPVLSA 719
VTN-LS-I
---T-TI-K---MI---EGM---.---I-M---GKN-L--VT-A---LKD-EVM-
INTE-KVF---I--L-V-VNK-AE-V-N---I-- 719VTD-VN---T-TI-KK---R-MI---EG---I-M---GKN-L--VT-A---VLKD-EVM-INTE-K-F---L---INK-AE-V---I--
719SSD-VSNI--M-AI--R--R-I----EGMN---I-R--H--PD-L-MT--K---A--VLKD-EVM-ADV--K-F---E-V---VDK-VD-V----I--
719 SSD-VRNI--T-AI--R--R- ---EGMN-I---I-R-H-RPD-L----T9---AA-V
VRD-EVM-ADV--K- ---E-V--VDK-VD-V----I-- 719 INVTTSSI--T-TI-R---MA ---GMN --- I---LNTN-L-E-VT-A---N-A--VIKD-EVL-AS--- KYF---E-E--VQK-AD-V.---I-- 720VSS-VSSI---T-TI-R---MR---EGMN---- ----l-R-
PN-L--VT-A---N-A--VINN-EV--A-T--R-F--R-E--D---VQK-AD-V----I--
720IIDFKTLKNLNDNYGITKKQAMELLHSNPKTLKEFINNNNPIIRNRIENLISQCRL - ---RIE-LN-IK--NV-RN--Q---
Q-L--K-
---RIE-FN-IK---NV-RN--Q---Q-L--K- ---RS--LD-IR-D-RV-RD---Q---K----Q-L--- ---RS--LD-IR-D-RV-RD---Q---K----Q--L--- --- --SRQ-LN--R-D-RV-R----QD----
S--M---.
---SRO--FN-R-D-RV-R----QD---QE--M----775 775
775
775 775
776 776
FIG. 2. Comparison ofVP4 amino acidsequence of strainK8with those of otherhuman and simian rotavirus strains. The entireVP4
amino acid sequences of KU, DS-1, M37, ST-3, SA-11,and RRV have been previously described (6, 7, 17-19, 27). Thepositions of the
predicted amino acid insertion (@) and deletion (0) in K8 VP4are indicated. Thecleavage sites are shown by arrows, and the site of
connecting peptide is boxed.
itsRNA pattern, the differencesin electrophoretic mobility
betweensegments 2and 3 and between10and 11are wider
than those for other serotype 1 and subgroup II strains
examined
thus far(29). (iii) The strain is not recognized byanti-VP2 MAb (YO-60 antibody), which is reactive
exclu-sively with
subgroup II strains (32). The relationship of theK8VP4genewithavarietyof animalrotavirus strainsneeds
further
study.
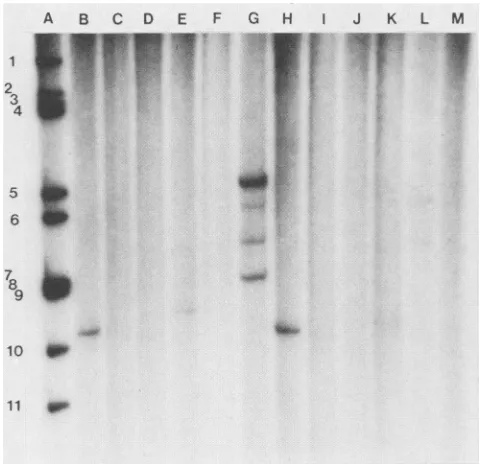
InanRNA-RNAhybridizationanalysis using32P-labeled
single-stranded K8 probeand virion RNA frombovine
(NCDV
and UK), simian (SA-11 and MMU18006),porcine
(Gottfried), equine
(H2 and FI-14), and feline (Cat97) strains,
wecould
not detect any evidence ofgeneticrelatedness
between
the VP4geneofK8and theVP4genesof theseanimal
strains (Fig.
4).Similar studies withavarietyof
naturally
occurring
human rotavirus strainsand K8 virus need tobe
carried
out. In addition, the preparation of N-MAbsspecific
to K8 VP4 would be useful in examining theprevalence of rotavirus
strains whichhaveaK8-likeVP4by using
ELISA
asthedetection method inboth humans andanimals
(31).
STRAINK8 KU DS-1 M37 ST-3
SAL1
RRV
K8 KU DS-1 M37 ST3 SAl1 RRV
K8 KU
DS-1
M37 ST3
SAL1
RRV
K8 KU DS-1 M37 ST3 SAI1 RRV
K8 KU
DS-1
M37 ST-3 SAL 1 RRV
K8 KU DS-1 M37 ST3 SA11
RRV
K8 KU DS-1 M37 ST3
SA1I
RRV
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.60.555.64.573.2]Amino acid critical to epitope II
Amino acid Amino acid critical to critical to
apitope III epitope II
KAANYQYN LDGEQVTA
-S
---PM____TT- __E_E___
__----
S----_
__---S ---__
_________ _-_______
313 313 314 313 313 313 313
380 380 381 380 380 380 380
TGGSYDFSIPVG
-___N--RL---_______ RL___
WPVMNGGA
----S--
-
--I.---400 400 401 400 400 400 400
428 SLTVI 428 429 R-A-S 428 428 428 428
PSFSI
p---RTRTVNLY
---MSGI---I
---____I___
447 447 448 447 447 447 447 FIG. 3. Comparison of amino acid residues criticaltocross-reactiveneutralization epitopesonVP4 of human rotavirusstrains. The entire
VP4amino acidsequences of thestrains other than strain K8 have been previously described(6,7, 13, 19, 27).
It has been
shown
thatantibody
to VP4 can mediateprotection
(24). The probability of antibody acquisition in
infants
against VP4 of K8-like strains
maybe less, since K8
VP4 lacks
the cross-reactive neutralization epitopes found in
serotype1,
3,
and 4 strains. The surveyof
prevalenceof
neutralizing antibodies
toHRV in different
age groupshas
shown that
antibody
distribution
patterns werequite
dif-ferent
between the
two serotype1 strains
(KU and K8); the
appearance
of antibodies
against the K8 strain after
birthwas
delayed significantly (35). This is
mostlikely reflected
by divergent VP4s, since
adifference of only
twoamino
acids
wasobserved
onVP7
between the
twostrains. Thus,
theunique
gene4 of the K8 strain
mayhave
arole in
allowing the K8 strain
to causeepidemic illness
inolder
A B C D E F
1
Ik-
;2 .
4w
G
I.!
H I J K L Mk{:
i:10 WI
[image:5.567.28.519.55.162.2]11 _
FIG. 4. Hybridization ofa labeled single-stranded RNA probe from strain K8 to genomic RNAs from human, feline, bovine, simian,equine, and porcine rotavirus strains. Before the RNA-RNA hybridization experiments, theamountofthedouble-stranded RNA
fromdifferent strainswascheckedby staining the double-stranded
RNA inthe gel withethidium bromide;we employedalmost equal
amountsof eachdouble-stranded RNA for thehybridization tests.
The RNA-RNA hybridization was performed as described
previ-ously (4). Lanes: A, K8 (human); B, P (human); C, DS-1 (human); D, Taka (feline); E, NCDV (bovine); F, SA-11 (simian); G, KU (human); H, ST-3 (human);I,RRV(simian); J, H2(equine); K, UK
(bovine); L, Gottfried (porcine); M, FI-14 (equine). Numberstothe
left ofthegel show the RNAsegmentorder.
children.
We haverecently
studiedthe
antibody
responsetoeach of several
neutralization epitopes
onVP7 and
VP4in
human
seraby competition binding
assayusing
N-MAbs(25;
K.Taniguchi
etal., manuscript in
preparation). It will also
beof
interest toexamine
theantibody prevalence
tothe
unique VP4 of strain
K8.
Wethank P. Collins andM. Hill for preparation of oligonucleotide primers. We also thank R. M. Chanock for criticism and encourage-mentand L. Jordan fortyping the manuscript.
LITERATURECITED
1. Clark, H. F., Y. Hoshino, L. M. Bell, J. Groff, G. Hess, P. Bachman, and P. A. Offit. 1987. Rotavirus isolate W161
repre-sentinga presumptivenew human rotavirus serotype. J. Clin. Microbiol. 25:1757-1762.
2. Espejo, R. T., S. Lopez, and C. Arias. 1981. Structural polypep-tides of simian rotavirus SA1l and the effect of trypsin. J. Virol. 37:156-160.
3. Estes, M. K.,D. Y.Graham, and B. B. Mason. 1981. Proteolytic enhancement of rotavirusinfectivity: molecular mechanism. J. Virol. 33:879-888.
4. Flores, J., H. B.Greenberg, J. Myslinski, A. R. Kalica, R. G. Wyatt, A. Z. Kapikian, and R. M. Chanock. 1982. Use of transcription probes for genotyping rotavirus reassortants. Vi-rology 121:288-295.
5. Flores, J., K. Midthun, Y. Hoshino, K. Green, M. Gorziglia, A. Z.Kapikian, and R. M. Chanock. 1986. Conservation of the fourth gene among rotaviruses recovered from asymptomatic newborn infants andits possible role in attenuation. J. Virol. 60:972-979.
6. Gorziglia, M.,K.Green,K.Nishikawa, K.Taniguchi,R.Jones, A. Z. Kapikian, and R. M. Chanock. 1988. Sequence of the
fourthgeneof humanrotavirusesrecovered fromasymptomatic
orsymptomatic infections. J. Virol. 62:2979-2984.
7. Gorziglia, M., Y. Hoshino,A.Buckler-White, R. Glass, J. Flores, A. Z. Kapikian, and R. M. Chanock. 1986. Conservation of amino acid sequence of VP8 and cleavage region of 84-kDa
outercapsid proteinamongrotavirusesrecoveredfrom
asymp-tomatic neonatal infection. Proc. Natl. Acad. Sci. USA 83:
7039-7043.
8. Greenberg,H.B., J.Flores,A. R.Kalica, R.G. Wyatt,and R.
Jones. 1983. Gene coding assignments for growth restriction, neutralization and subgroup specificities of the W and Ds-1 strains of humanrotavirus. J. Gen. Virol. 64:313-320.
9. Hoshino, Y., M. M. Sereno, K. Midthun, J. Flores, A. Z.
Kapikian,and R. M.Chanock.1985.Independent segregationof twoantigenic specificities (VP3 and VP7) involvedin
neutrali-zation of rotavirus infectivity. Proc. Natl. Acad. Sci. USA
82:8701-8704.
10. Hoshino, Y.,R.G.Wyatt,H. B.Greenberg,J. Flores,and A. Z.
Kapikian. 1984.Serotype similarity and diversity of rotaviruses
of mammalian and avianoriginasstudiedby plaque reduction
neutralization. J. Infect. Dis. 149:694-702.
11. Kalica,A.R., J.Flores,and H. B.Greenberg. 1983.
Identifica-tionof the rotaviral genes that code forhemagglutination and Strain
(Serotype)
Amino acid critical to epitope I
KU (1) Wa (1) K8 (1) DS-1 (2) RV-5 (2) P (3) VA-70 (4)
296 296 297 296 296 296 296
_~
!
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.567.25.265.362.594.2]protease-enhanced plaque formation. Virology 125:194-205. 12. Kalica, A. R., H. B. Greenberg, R. G. Wyatt, J. Flores, M. M.
Sereno, A. Z. Kapikian, and R. M. Chanock. 1981. Genes of human(strain Wa) and bovine (strain UK) rotaviruses that code forneutralization and subgroup antigens. Virology 112:385-390. 13. Kantharidis, P., M. L. Dyall-Smith, and I. H. Holmes. 1987.
Markedsequencevariation betweensegment4genesof human
RV-5and simian SAil rotaviruses. Arch. Virol. 93:111-121. 14. Kapikian, A. Z., and R. M. Chanock. 1985. Rotaviruses, p.
863-906. In B. N. Fields, D. N. Knipe, R. M. Chanock, J. L. Melnick, B. Roizman, and R. E. Shope (ed.), Virology. Raven Press, New York.
15. Kapikian,A. Z., J. Flores, Y. Hoshino, R.I.Glass, K.Midthun,
M. Gorziglia, and R. M. Chanock. 1986. Rotavirus: the major etiologicagentofsevereinfantile diarrheamaybe controllable by a "Jennerian" approach to vaccination. J. Infect. Dis. 153:815-822.
16. Kozak, M. 1981. Possible role of flanking nucleotide in recogni-tion of the ATG initiator codon by eukaryotic ribosomes. Nucleic Acids Res. 9:5233-5252.
17. Lopez, S., and C. F. Arias. 1987. The nucleotidesequenceof the
5' and 3' ends of rotavirus SAil gene4. Nucleic Acids Res.
15:4691.
18. Lopez, S., C.F. Arias, J. R. Bell, J. H.Strauss,and R. T.Espejo. 1985. Primary structure of the cleavage site associated with trypsin enhancement of rotavirus SAll infectivity. Virology
144:11-19.
19. Mackow, E. R., R. D. Shaw, S. M. Matsui, P. T. Vo, M. N. Dang, and H. B. Greenberg. 1988. Characterization of the rhesus rotavirus VP3gene:location of amino acids involved in
homol-ogous andheterologous rotavirus neutralization and
identifica-tion ofa putative fusion region. Proc. Natl. Acad. Sci. USA 85:645-649.
20. Matsuno, S.,A. Hasegawa, A. Mukoyama, andS. Inouye.1985. Acandidate for a new serotype of human rotavirus. J. Virol. 54:623-624.
21. Morita, Y., K. Taniguchi, T. Urasawa, and S. Urasawa. 1988. Analysis of serotype-specific neutralization epitopesonVP7of humanrotavirus by theuseof neutralizing monoclonal
antibod-iesand antigenic variants. J. Gen. Virol. 69:451-458.
22. Offit, P. A., and G. Blavat. 1986. Identification of the two
rotavirusgenesdetermining neutralization specificities.J.Virol.
57:376-378.
23. Offit, P. A., G. Blavat, H. B. Greenberg, and H. F. Clark. 1986. Molecularbasis of rotavirus virulence: role ofgenesegment4. J. Virol. 57:46-49.
24. Offit, P. A., R. D. Shaw, and H. B. Greenberg. 1986. Passive protection against rotavirus-induced diarrhea by monoclonal antibodiestosurface proteins vp3 and vp7. J. Virol. 58:700-703.
25. Shaw, R. D., K. J. Fong, G. A. Losonsky, M. M. Levine, Y.
Maldonado, R. Yolken, J. Flores, A. Z. Kapikian, P. T. Vo, and
H. B. Greenberg. 1987. Epitope-specific immune response to
rotavirusvaccination. Gastroenterology 93:941-950.
26. Taniguchi, K.,Y.Hoshino, K. Nishikawa, K. Y. Green, W. L.
Maloy, Y. Morita, S. Urasawa, A. Z.Kapikian, R. M. Chanock, and M. Gorziglia. 1988. Cross-reactive and serotype-specific neutralizationepitopes on VP7 of human rotavirus: nucleotide sequence analysis of antigenic mutants selected with monoclo-nal antibodies. J. Virol. 62:1870-1874.
27. Taniguchi, K., W. L. Maloy, K. Nishikawa, K. Y. Green, Y. Hoshino, S. Urasawa, A. Z. Kapikian, R. M. Chanock, and M. Gorziglia. 1988. Identification of cross-reactive and serotype 2-specific neutralization epitopes on VP3 of human rotavirus. J. Virol. 62:2421-2426.
28. Taniguchi, K., Y. Morita, T. Urasawa, and S. Urasawa. 1987. Cross-reactive neutralizationepitopes on VP3 of human rotavi-rus:analysis with monoclonal antibodies and antigenic variants. J. Virol. 61:1726-1730.
29. Taniguchi, K., S. Urasawa, and T. Urasawa. 1982. Electropho-reticanalysisof RNA segments of humanrotavirusescultivated incell culture. J. Gen. Virol. 60:171-175.
30. Taniguchi, K., S. Urasawa, and T. Urasawa. 1985. Preparation andcharacterization ofneutralizing monoclonalantibodies with differentreactivitypatterns to human rotaviruses. J. Gen. Virol. 66:1045-1053.
31. Taniguchi, K., T. Urasawa, Y.Morita, H. B. Greenberg, and S. Urasawa. 1987. Direct serotyping of human rotavirus in stools by anenzyme-linked immunosorbent assay using serotype 1-, 2-, 3-, and4-specific monoclonal antibodiesto VP7. J. Infect. Dis. 155:1159-1166.
32. Taniguchi, K., T. Urasawa, and S. Urasawa. 1986. Reactivity patterns to humanrotavirus strains ofamonoclonalantibody againstVP2, a component of the inner capsid of rotavirus. Arch. Virol. 87:135-141.
33. Taniguchi, K., T. Urasawa, S. Urasawa, and T. Yasuhara. 1984. Production ofsubgroup-specific monoclonal antibodies against human rotaviruses and their application to an enzyme-linked immunosorbent assay for subgroup determination. J. Med. Virol. 14:115-125.
34. Urasawa, S.,T.Urasawa,and K.Taniguchi.1982.Threehuman rotavirus serotypes demonstrated by plaque neutralization of isolated strains. Infect. Immun.38:781-784.
35. Urasawa, S., T. Urasawa, K. Taniguchi, and S. Chiba. 1984. Serotype determination of human rotavirus isolates and anti-body prevalence in pediatric population in Hokkaido, Japan. Arch.Virol. 81:1-12.
36. Vesikari, T., E. Isolauri, E. D'Hondt, A. Delem, F. E. Andre, and G. Zissis. 1984. Protection of infants against rotavirus diarrhea by RIT 4237 attenuated bovine rotavirus strain vac-cine. Lancet i:977-981.
37. Ward, R. L., D. R. Knowlton, G. M. Schliff,Y. Hoshino, and H. B. Greenberg. 1988. Relative concentrations ofserum neu-tralizing antibody toVP3 and VP7 proteins in adults infected with ahuman rotavirus.J.Virol.62:1543-1549.
38. Wyatt,R.G.,A. Z.Kapikian,andA.Mebus. 1983.Induction of cross-reactiveserumneutralizing antibody tohumanrotavirus in calves after in utero administration ofbovine rotavirus. J. Clin. Microbiol. 18:505-508.