0022-538X/87/041266-05$02.00/0
Copyright © 1987,AmericanSociety for Microbiology
Course and Extent of Variation
of Equine Infectious Anemia Virus
during Parallel Persistent
Infectionst
SUSAN L. PAYNE,' OLIVIASALINOVICH,'t SUZANNE M. NAUMAN,' CHARLES J. ISSEL,2'3 AND RONALD C.
MONTELARO1*
Departments of
Biochemistry' and Veterinary Science,2
and LouisianaAgricultural Experiment
Station,"
2andtheDepartment of
VeterinaryMicrobiology and
Parasitology,3
Schoolof
Veterinary Medicine,
Louisiana StateUniversity,
Baton
Rouge, Louisiana
70803Received 3 September 1986/Accepted 16 December 1986
Comparisons
ofpeptide
andoligonucleotide
mapsofglycoproteins
and RNA from nineisolates
ofequine
infectious
anemiavirus (EIAV) thatweregeneratedduring parallel
infections oftwoShetlandponiesrevealed
that each
isolate
wasstructurally unique. Each
EIAVisolate contained aunique
subset of variantpeptides,
oligonucleotides,
orboth, indicating
that structural variation inEIAV
isarandomand noncumulativeprocessand
thatalarge
spectrumofpossible
EIAV variantscanbegenerated
in infected animals.The
members of the lentivirus subfamily
ofretroviruses,
including equine infectious
anemia virus(EIAV),
visnavirus, and human immunodeficiency
virus(HIV),
causechronic diseases ina
variety
of animals andhumans
(2, 5, 6,
8, 16).
Thepersistent
infections
causedby
these viruses in someinstances
canbe traced
totheir
ability
tocircumvent
or overcomethe immuneresponsesof
the host(4, 6, 9,
18).
In thecaseof
EIAV, which
causesaunique episodic
diseasein
horses, it has been demonstrated that isolates recovered
from
aninfected
animalduring
different febrileepisodes
canbe
distinguished
antigenically
andstructurally by
avariety
ofbiochemical
andimmunological
assays(11, 15, 18). Results
of these studies have indicated that structural and
antigenic
variation is localized
tothe envelope glycoproteins gp9O and
gp45 (11, 18).
The occurrence ofenvelope
glycoprotein
variation has also been
demonstrated for visna virus and
HIV
(1, 4, 7, 19).
A
critical question regarding lentivirus variation is
the numberof variants that
canoccurinnature,amajor
factor
inassessing potential
vaccines. To analyze the
spectrumof
structural
variation
amongisolates
ofEIAV,
we usedpep-tide, glycopeppep-tide, and oligonucleotide mapping procedures
to compare nine EIAV
isolates
recoveredduring distinct
clinical
episodes
in twoponies infected with the
same virus.To
conduct these
experiments, identical virulent virus
inocula
wereused in
parallel infections of
twoShetland
ponies
asdescribed
previously (11, 13, 18). The clinical
histories
of these
animals
aresummarized in Fig.
1.Plasma
samples
takenduring
clinical episodes
weresubjected
toendpoint dilution in fetal equine kidney cells
torecoverthe
predominant virus
population (13). The nine
EIAVisolates
recovered were
propagated in fetal equine kidney
cells andpurified
by glycerol gradient centrifugation
(10, 11, 14). Each virusisolate
wasdetermined
tobe antigenically
distinct byusing
avariety of
immunoassays employing
serum frominfected
ponies, panels
of monoclonal antibodies, or both* Corresponding author.
t Louisiana Agricultural Experiment Station paper no. 86-12-0110.
tPresentaddress: Lovelace InhalationToxicologyResearch
In-stitute, Albuquerque, NM 87185.
(18; A. Orrego, Ph.D.
thesis, Louisiana State
University,
Baton
Rouge,
1983).
Theextent
of
structural
variationin the
viralglycoproteins
gp9O
andgp45
for each of thenine
EIAV isolates wasassayed
by two-dimensional
125I-labeled
tryptic
peptide
(11,
a)
0.
H-cL
-a a
0)
-_
a.0a. a.
r(i re5
a. a.
Doys
Post
Inoculotion
FIG. 1. Clinical histories of experimentally infected ponies showing clinical episodes from which virusisolateswererecovered. Both animalsreceivedidentical.intravenous virus inocula containing
4.8loglo50% tissuecultureinfective doses ofhost-adapted EIAV (13).Allsustainedrectaltemperaturerecordings above 39°C (dashed line) were considered abnormal and were designated febrile
epi-sodes. (A) Pony 91 exhibited classical chronic equine infectious anemia thatwascharacterized by recurring febrile episodes.Pony 91 died after fourfebrileepisodes, onday 180. (B) Pony 127 also experienced chronic equine infectious anemia. The 5 months
be-tweenthe thirdandfourth febrileepisodesrepresentaninapparent
stage of the diseasein which the asymptomatic animal remainsa
viruscamer. 1266
on November 10, 2019 by guest
http://jvi.asm.org/
[image:1.612.327.564.354.605.2]B
600
14 015
I0
eo
P
O02
Cj
32 33 03541
330 4 4
U 5
A2
0
*0*4
3
60 9.31
39410001
lo*'
160
2
U
446
onz
170
-.19
230 'ic2
310@033 350
02
32 347@ O
4
10
32
3!41012
,,
A
U~~~~~~~'O
sbt
b
_
_
a
Q- O"
[image:2.612.136.494.68.308.2]OU0
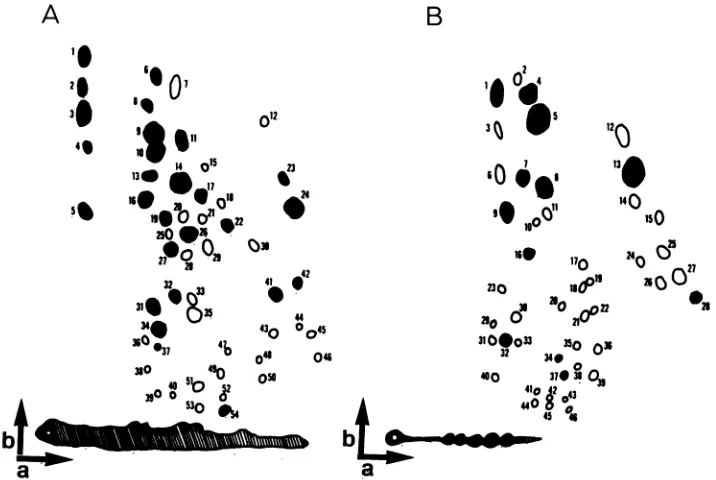
FIG. 2. Compositetwo-dimensionaltryptic peptidepatternsgenerated frommapsof 251I-labeled glycoproteinsgp9O and gp45 from each
EIAVisolate. Thepeptides generated for each virusisolateweremapped in duplicate. The compositemapsweregenerated by comparing
each virus isolate with all other isolates by directly overlaying autoradiographs. Arrowsindicate the directions ofelectrophoresis (a) and
chromatography (b). Arbitrary numberswereassignedtoeachpeptidefor identification.Closed circlesindicatepeptides thatwere common
toallvirusisolates. Open circles indicate variant peptides.Thehatchedarearepresentsradioiodinatedtryptic glycopeptides thatwerenot
resolved bystandardpeptide mapping techniques(11). (A) gp9Ocompositepeptidemap.(B)gp45composite peptidemap.
18)
and
glycopeptide (17) mapping
procedures. All peptide andglycopeptide
patterns werereproducible
onrepeated
mappings,
and virus isolates
remained stable during
contin-uedpassagein
tissue
culture. Figure 2 shows the
composite
maps
developed
todisplay
conserved and variant peptides
for
gp9O and gp45.
Atotal
of 54
peptides
wereidentified
oncomparison
of thegp9O
componentof the nine variants and
the prototype
strain of EIAV.
Twenty-seven(50%) of the
gp9O
peptides
werefound
tobe
commontoall
isolates. The
pattern
of variation
of the remaining
peptides
wasused
touniquely identify eight of
ninevariant
strains,
aswell
asthe
prototype
strain of the
virus(Table 1, gp9O peptides). Only
isolates
P3.2-2
andP3.2-3
could
notbe
distinguished by gp9O
peptide
mappinganalysis. For gp45 only
11of
45peptides
(24%)
werecommontoall
isolates. In
this
casethe
patternof
variant
peptides could be used
todistinguish all isolates
(Table 1, gp45
peptides); P3.2-2 and P3.2-3 could be
distin-guished.
Table 1 lists those
peptides
not common to all virus isolates. Some of these variantpeptides
were present in isolates from onlyone pony. Theseincluded gp9O
peptides
12, 18, 20,
and25and
gp45
peptides 6, 14, 15, 25, 27, 29,
and 30.gp9O peptides
18 and 25 wereunique
tosingle
virus isolates. Mostother
peptides
were present in and variedamong
isolates
from bothanimals,
however. Forexample,
gp9O peptide
49 was present inP3.1-2,
P3.2-1,
andP3.2-4;
gp9O peptide
52waspresent inP3.1-1, P3.1-3, P3.2-2, P3.2-3,
and P3.2-4. The data suggest that structural variations
among virus isolates do not accumulate with
time,
aspep-tides appear in one isolate, only to be lost in subsequent isolates.
Instead,
it appears that a limited set ofpeptides
varyindependently, leading
to the presence of a different subset ofpeptides
for each isolate.In contrast to the
envelope
glycoproteins, the peptide
maps
for
the viral coreproteins p9,
p15,and p26
wereidentical for all virus isolates (data
notshQwn),
asreported
previously (11, 17).
gp9O
andgp45
fromeach virus isolatewerealso
analyzed
by
glycopeptide mapping
to compare theglycosylated
tryptic
peptides
notresolved by standard
peptide mapping
procedures (11, 17, 18). Four classes of gp9O and
twoclasses
of
gp45
glycopeptide
patterns wereobserved.
Representa-tive
glycopeptide
maps areshown
inFig.
3, and
inTable
2 theglycopeptide
patternsobtained for each
virusisolate
aresummarized.
Thesedata
indicate thatchanges
inthe
pattern ofglycosylation
ofgp9O
andgp45
areindependent of
oneanother.
For
example,
all virusisolates from
pony91
(P3.2-1
through P3.2-4) share
acommongp45
glycosylation
pattern,while they
exhibit four different gp9O
glycosylation
patterns. Itisinteresting
that virus isolatesP3.2-2 andP3.2-3,
whichproduce
apparently
identicalgp9O
peptide
patterns, exhibit differentglycopeptide
maps.To
analyze
thegenomic
variation among EIAV isolates, RNAwaspurified
fromeach virus andanalyzed by
oligonu-cleotide
mapping by
previously described techniques (15,
18).
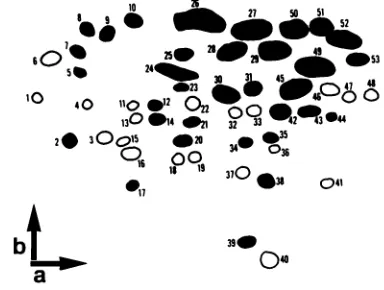
Acomposite
mapshowing
conserved and variantoligo-nucleotides
was thendeveloped (Fig. 4).
A total of 51oligonucleotides
arerepresented
inFig. 4,
and 33(65%)
of these were common toall isolates.Eight
ofthe nine virus strains and the prototype virus could bedistinguished by
analysis
of the distribution ofvariantoligonucleotides. Only
isolates
P3.2-1
andP3.2-4
couldnotbedistinguished easily.
(Table 3).
As observed with thepeptide
maps described above, thereappeared
to be no accumulation of variantoligonucleotides
amonglate virus isolates.Results of the
experiments
presented
hereprovide
aA
b4
120
lSO
24
05
0° o3
ft
0
lm-23
m
on November 10, 2019 by guest
http://jvi.asm.org/
detailed comparison of nine virus
isolates recovered from
two
ponies
infected with identical
inocula of EIAV. Several
important properties of EIAV variation
canbe concluded
from these observations.
A
distinct virus population predominates during each
febrile episode in
apersistently infected
pony.The variant
virus strains examined
in this study
wererecovered from
endpoint
dilutions of plasma and, thus,
areassumed
to representthe
predominant virus populations in the infected
animal
atthe time the plasma samples
wereobtained.
[image:3.612.319.556.69.275.2]There
appearstobe
arelatively
large number of structural
variations possible in EIAV. In
addition
tothe nine virus
isolates
described here,
werecovered five
moreEIAV
isolates from
athird
ponythat received the
sameinitial virus
inoculum (unpublished data). These virus isolates could also
be
distinguished structurally,
bringing the total number of
unique
variants generated from this virus inoculum
to14.
These 14
isolates
weredistinct
from isolates recovered from
TABLE 1. DistributionamongEIAVstrains of variantpeptides from the envelope glycoproteins gp9O and gp45
Distribution of: Virus Peptide
gp9O peptidesa
gp45peptidesb
Prototype 7, 20, 28, 36, 44, 45, 46, 47, 49, 50, 51, 52, 53
P3.1-1 7, 15, 28, 29, 30, 33, 35, 36, 38, 39, 40, 44, 45,46, 47, 51, 52, 53 P3.1-2 7, 12, 15, 21, 29, 35, 36, 38, 39, 43,
46, 47, 48,49, 50, 51
P3.1-3 7, 15, 25, 28, 29, 30, 33, 35, 39, 44, 45,46, 47, 52, 53
P3.1-4 7, 12, 15, 29, 30, 35, 38, 39, 43, 46, 47 P3.2-1 20, 28, 36,38, 39, 40, 44, 45, 46, 49,
51, 53
P3.2-2 7, 15, 20, 21, 28, 29, 30, 36, 38, 39, 40, 43, 44,45, 46, 47, 48, 50, 51, 52, 53
P3.2-3 7, 15, 20, 21, 28, 29, 30, 36, 38, 39, 40, 43, 44,45, 46, 47, 48, 50, 51, 52, 53
P3.2-4 7, 15, 18, 20, 21, 28, 29, 30, 36, 38, 39, 40, 43,44, 45, 46, 47, 49, 50, 51, 53
P3.2-5 7, 20, 21, 28, 29, 30, 33, 35, 40, 43, 44, 45, 47,48, 50, 51, 52, 53
Prototype
P3.1-1
P3.1-2
P3.1-3
P3.1-4
P3.2-1
P3.2-2
P3.2-3
P3.2-4
P3.2-5
3, 6, 12, 17, 21, 24, 26, 35, 36, 39, 41 2, 3, 21, 22, 23, 30, 31, 33, 35, 41, 42,
44,45
2, 3, 10, 18, 20, 22, 23, 29, 33, 38, 41, 42, 44, 45
2, 10, 11, 12, 17, 18, 20, 21, 22, 23, 24, 26, 33,35, 36, 38, 39, 41, 42, 43, 44, 45, 46
2, 3, 10, 17,18, 20, 22, 23, 29, 33, 36, 39, 41, 42, 44,45
3, 6, 11, 12,18, 19, 21, 23, 24, 26, 35, 36, 39, 41, 42, 43
2, 10, 11, 12, 15, 18, 19, 21, 24, 25, 27, 30, 35,39, 42, 45
2, 6, 10, 11,12, 14, 15, 17, 18, 21, 22, 23, 24, 27,30, 36, 39, 41, 43, 44, 45,46
6, 10, 11, 12,18, 21, 23, 24, 26, 30, 35, 36, 38,39, 42, 44, 45, 46 2, 6, 10, 11, 12, 14, 18, 20, 21, 22, 24,
25, 26, 27,30, 33, 35, 36, 38, 39, 42, 44, 45, 46
ICompare with Fig. 2A.
bCompare with Fig. 2B.
A B
1I
L.
[if IV 1I
FIG. 3. Examples ofglycopeptide patterns obtained for lectin-purified
'l25-labeled
gp9O and gp45 glycopeptides. Foranalysis of these patterns, the presence or absence of vertical groups of glycopeptides, as opposed to individualglycopeptides
were com-pared, such that possiblemicroheterogeneity
of the carbohydrate moiety did not interfere with thecomparisons
(17). (A) gp9O glycopeptidemaps.Class I,P3.2-1; class II,P3.2-2; class III, P3.1-3; classIV, P3.2-5(B) gp45glycopeptidemaps. Class I, P3.1-1;class II,P3.1-2. For all maps thedirection ofelectrophoresis
wasfromleft toright;thedirection ofchromatographywasfrombottomtotop.independent
serial transmissions of
EIAV betweenponies
(11, 15).
Thus,
atotal of
atleast 17 distinct structural
variants
werecataloged in our
laboratory. This large range of
EIAV variation is similar to
thatobserved
for
HIV,
inwhich
no
two virus
isolates examined have been found to be
identical
in restriction enzyme
mapping, DNA sequencing
studies,
orboth
(1).
The
evolution of EIAV variants
during
apersistent
infec-tion is
evidently random; i.e., no predictable sequence of
virus variation was observed in the two
experimentally
infected
animals, although
each animal
received
identical
virus inocula. This result differs from the data
reported for
parallel
persistent infections with visna
virus,
in which
similar
patterns
of variantevolution were observed
(3). This
difference
may reflect that a
larger
spectrum
of variation
is
possible
in EIAV
compared
with visna virus.
TABLE 2. Classification ofEIAVisolates byglycopeptide patternsof theenvelope glycoproteinsgp90andgp45
Viruses in: Glycoprotein
ClassI ClassII ClassIII ClassIV
gp9O Prototype P3.1-1 P3.1-3 P3.2-3
P3.2-1 P3.1-2 P3.2-5
P3.2-4 P3.1-4
gp45 Prototype P3.1-2
P3.1-1 P3.1-4
P3.1-3
P3.2-1 toP3.2-5
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.57.300.279.714.2] [image:3.612.315.555.611.718.2]10 a
8
D27
_ 510 214 a05
60Q4S 25 *5
24 34
0 Q023 0 73 C
40 1iO Q0
(n
2-300 340@ *5
1979 370
ba
39,o4o
FIG. 4. Composite oligonucleotidefingerprint generated by
com-parisonsofoligonucleotidemapsfromEIAVisolates. Forpurposes
ofcomparison, viralRNAfromeach virusisolatewas purified and
subjected to mapping procedures on at least two separate
occas-sions, and typically three to four maps were used for making
comparisons between any two isolates. Comparisons between all
isolates were made by directly overlaying autoradiographs. The
composite reflectsonly the high-molecular-weight (lower) region of
theoriginalmaps. Arbitrary numbers were assigned toeach
oligo-nucleotidefor identification. Closed circles indicate oligonucleotides
thatwere commontoallvirus isolates. Open circles indicate variant
oligonucleotides. The directions of first-dimension (a) (8%
poly-acrylamide; pH 3.3) and second-dimension (b) (22%
polyacryl-amide;pH 8.2) electrophoresis are indicated byarrows.
The
variations in EIAV genomic
RNA and glycoproteinsare not
necessarily cumulative. Subsequent
virus strainsmay not
arise from the virus strain which predominates
inthe
preceding febrile episode. Rather, it
appearsthat
randommutations
generated during virus replication
result in avariety of
integrated proviruses,
any oneof which may lead tothe
generation of future predominant virus
strains. In contrast toEIAV,
sequential isolates of visna display
cumu-lative
changes (3). Sequential
HIVisolates
like EIAV,however,
fail
todisplay
cumulative
changes.Noncumulative
variation of HIV has been interpreted
toindicate
parallelevolution
of variants in the infected individual
(7). Analternative
explanation for the results obtained for both
EIAV
and HIV would be
ahigh
mutation
ratewhich masks
any
direct
lineage between
sequential isolates.
The time
required for the evolution of variant populations
of EIAV
is
variable
but
canbe
remarkably rapid. The
shortest
time
observed between clinical episodes and
dis-tinct
EIAV
isolates
wasapproximately 15 days (P3.2-1
to [image:4.612.78.270.72.217.2]P3.2-2). In
contrast,the
generation of variants of visna virus
TABLE 3. Distributionof variantoligonucleotides
Virus Oligonucleotidea
Prototype 1, 4, 6, 13, 15, 16, 19, 32,41, 47 P3.1-1 1, 4, 6, 13, 16, 19, 32,36, 40, 47 P3.1-2 1, 4, 6, 16, 19,32, 36
P3.1-3 1, 4, 6, 13, 16, 19, 32,36, 46, 47 P3.1-4 1, 4,6, 13, 16, 19, 32, 33,36 P3.2-1 (1), 3,(4),6, 13, 16, 19,32, 36, 47 P3.2-2 1, 6, (11),18, 19, 32, (36)
P3.2-3 13, 18, 37,47
P3.2-4 1, 3, 4, 6,13, 16, 19, 32, 36,47 P3.2-5 1, 6, 11, 13, 18,(19), 22, 32, (36), 48
aParenthesesindicatethepresenceofaweaksignalin theposition ofthe
oligonucleotideforthat virus isolate.
usually requires at least a year (12), and the generation of
HIV variants is believed to follow a similar time course (7).
In this regard, EIAV provides a unique model to study
rapidly changing virus structure, as well as the dynamic
interaction between host immune responses and evolving
lentivirus antigens.
This work was supported by the Louisiana Agricultural Experi-ment Station, the Louisiana State University School of Veterinary Medicine, and Public Health Service grant CA-38851 from the National Cancer Institute.
LITERATURE CITED
1. Alizon, M., S. Wain-Hobson, L. Montagnier, and P. Sonigo. 1986. Genetic variability of the AIDS virus: nucleotide sequence analysis of two isolates from African patients. Cell 46:63-74. 2. Barre-Sinoussi, F., J. C. Chermann, F. Rey, M. T. Nugeyre, S.
Chamaret, J. Gruest, C. Dauguet, C. Axler-Blin, F. Brun-Vezinet, C. Rouzioux, W. Rozenbaum, and L. Montagnier. 1983. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science 220:868-871.
3. Clements, J. E., N. D'Antonio, and0. Narayan. 1982. Genomic changes associated with antigenic variation of visna virus. II. Common nucleotide sequence changes detected in variants from independent isolations. J. Mol. Biol. 158:415-434.
4. Clements, J. E., F. S. Pedersen, 0. Narayan, and W. A. Haseltine. 1980. Genomic changes associated with antigenic variation ofvisna virus during persistent infection. Proc. Natl. Acad. Sci. USA 77:4454 4458.
5. Gallo, R. C., P. S. Sarin, E. P. Gelmann, M. Robert-Guroff, E. Richardson, V. S. Kalyanaraman, D. Mann, G. D. Sidhu, R. E. Stahl, S. Zolla-Pazner, J. Leibowitch, and M. Popovic. 1983. Isolation of human T-cell leukemia virus in acquired immune deficiency syndrome (AIDS). Science220:865-867.
6. Haase,A. T.1986.Pathogenesisof lentivirus infections. Nature (London)322:130-136.
7. Hahn, B., G. Shaw, M. Taylor, R. Redfield, P. Markham, S. Salahuddin, F. Wong-Staal, R. Gallo, E. Parks, and W. Parks. 1986.Genetic variation inHTLV-III/LAVover time in patients withAIDS or risk for AIDS. Science 232:1548-1553.
8. Issel, C. J., and L. Coggins. 1979. Equine infectious anemia: currentknowledge. J. Am.Vet. Med. Assoc. 174:727-733. 9. Kono, Y., K.Kobayashi,and Y.Fukunaga. 1973. Antigenic drift
ofequine infectious anemia virus in chronically infected horses. Arch. GesamteVirusforsch. 41:1-10.
10. Montelaro, R. C., N. Lohrey, B. Parekh, E. W. Blakeney, and C. J. Issel. 1982. Isolation and comparative biochemical prop-erties ofthe major internal polypeptides of equine infectious anemia virus. J. Virol. 42:1029-1038.
11. Montelaro, R. C., B. Parekh, A. Orrego, and C. J. Issel. 1984. Antigenic variation during persistent infection by equine infec-tious anemia virus, a retrovirus. J. Biol. Chem. 259:10539-10544.
12. Narayan, O., J. E. Clements, S. Kennedy-Stoskopf, and W. Royal, III. 1986. Antigenic variation in the lentiviruses that causevisna-maedi insheep andarthritis-encephalitisin goats, p. 25-40. In T. H. Birkbeck and C. W. Penn (ed.), Antigenic variation ininfectious diseases. IRL Press, Washington, D.C. 13. Orrego, A., C. J. Issel, R. C.Montelaro, and W. V. Adams, Jr.
1982. Virulence andin vitro growth of acell-adapted strain of equine infectious anemia virus after serial passage in ponies. Am. J. Vet. Res.43:1556-1560.
14. Parekh, B., C. J. Issel, and R. C. Montelaro. 1980. Equine infectious anemia virus, a putative lentivirus, contains polypeptides analogous to prototype-C oncornaviruses. Virol-ogy107:520-525.
15. Payne, S., B. Parekh, R. C. Montelaro, and C. J. Issel. 1984. Genomic alterations associated with persistent infections by equine infectiousanemiavirus,aretrovirus. 1984. J. Gen. Virol. 65:1395-1399.
16. Popovic, M., M. G. Sarngadharan, E. Read, and R. C. Gallo.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.60.295.593.705.2]1984. Detection, isolation, and continuous production of cytopathic retroviruses (HTLV-IIL) from patients with AIDS andpre-AIDS. Science 224:497-500.
17. Salinovich, O., and R. C. Montelaro. 1986. Comparison of glycoproteins by two-dimensional mapping of glycosylated
pep-tides. Anal. Biochem. 157:19-27.
18. Salinovich, O., S. L. Payne, R. C. Montelaro, K. A. Hussain,
C. J.Issel,and K. L. Schnorr. 1986.Rapidemergenceof novel
antigenic and genetic variants ofequineinfectious anemia virus duringpersistentinfection. J. Virol. 57:71-80.
19. Wong-Staal, F., G. M. Shaw, B. H. Hahn, S. Z. Salahuddin, M. Popovic, P. Markham, R. Redfield, and R. C. Gallo. 1985. Genomic diversity of human T-lymphotropic virus type III (HTLV-III).Science 229:759-762.