0022-538X/93/084651-08$02.00/0
Copyright © 1993,AmericanSociety for Microbiology
Functional Expression and
Characterization
of
the
Epstein-Barr
Virus DNA
Polymerase Catalytic Subunit
TATSUYA
TSURUMI,l*
AKIO KOBAYASHI,2 KATSUYUKI TAMAI,2TOHRU DAIKOKU,1 RYUTARO KURACHI,1ANDYUKIHIRONISHIYAMA'
Laboratory ofVirology, Research Institute for Disease Mechanism and Control, Nagoya University Schoolof Medicine, Tsuruma-cho, Showa-Ku, Nagoya
466,1
andResearch and DevelopmentDepartment, Medicaland BiologicalLaboratories, Ohara, Terasawaoka, Ina 396,2Japan Received 25 February 1993/Accepted 13 May 1993
A recombinant baculovirus containing the complete sequence for the Epstein-Barr virus (EBV) DNA polymerase catalytic subunit, BALF5 gene product, underthe control of thebaculovirus polyhedrin promoter wasconstructed. Insect cells infected with the recombinant virus produced a protein of 110 kDa, recognized by anti-BALF5 protein-specific polyclonal antibody. The expressed EBV DNA polymerase catalytic polypeptide was purified from the cytosolic fraction of the recombinant virus-infected insect cells. The purified protein exhibited both DNA polymerase and3'-to-5' exonuclease activities, which were neutralized by the anti-BALF5 protein-specific antibody. These results indicate that the 3'-to-5' exonucleaseactivityassociated with the EBV DNApolymerase (T. Tsurumi, Virology 182:376-381, 1991) is an inherent feature of the polymerase catalytic polypeptide. The DNA polymerase and the exonuclease activities of the EBV DNA polymerase catalytic subunit
were sensitive to ammonium sulfate in contrast to those of the polymerase complex purified from EBV-producinglymphoblastoidcells, which were stimulated by salt. Furthermore, the template-primer preference for the polymerase catalytic subunit was different from that for the polymerase complex. These observations strongly suggest that the presence of EBV DNA polymerase accessory protein, BMRF1 gene product, does influence theenzymatic properties of EBV DNA polymerase catalytic subunit.
Epstein-Barrvirus(EBV)isahuman herpesvirus which is
a causative agent of infectious mononucleosis. The EBV genomeconsists ofalinear double-strandedDNA, which is 172 kb in length and encodes approximately 84 proteins (1). EBVinfects and immortalizes B lymphocytes in vitro. The EBV has both a latent state and a lytic replicative cycle. During the latent phase of the EBV life cycle, the EBV genomeis maintainedas acircularplasmidmolecule synthe-sized by host DNA polymerases (Pols) in the nuclei of the immortalized cells. ori
P,
one replication origin of EBV, mediates this type ofreplication (29).However, after induc-tion of the lytic phaseof viral replication, EBVreplication proteinsareinduced and the EBV genome is amplified100-to 1,000-fold. The replication product is a head-to-tail
con-catemer, which is synthesized by the EBV DNA Pol via a rolling-circle mechanisminitiated from the otherreplication origin, ori Lyt (8). A number of features of EBV DNA replicationmake itanattractive model system for thestudy ofeukaryoticDNAreplication.
Recently, it has been demonstrated that EBV encodes at
least sevenviral genes thatare essential for ori Lyt-depen-dent DNA replication (6). These genes and their functions which are known or predicted on the basis of sequence comparison with herpes simplex virus (HSV) type 1 are
BALF5, the DNA Pol catalytic subunit; BMRF1, theDNA Pol accessory subunit; BALF2, the single-stranded DNA-binding protein; BBLF4 and BSLF1, the helicase and pri-mase;BBLF2/3, apotentialhomolog of the third component of thehelicase-primase complex; and genes encodedby the EBVSalI-F fragment, of unknown function. Ourgoal isto
understand theenzymaticprocessesduring thelyticphase of
EBV DNAreplication.
* Correspondingauthor.
4651
Sofar,we havefocusedoureffortsonthe studyof EBV DNA Pol. The EBV DNAPol has been purifiedtovarying degrees fromculturedEBV-infectedcells andcharacterized (11, 16, 24). The 110 kDa of the EBV DNA Pol catalytic polypeptide copurifieswith the BMRF1 gene product. The neutralization of the EBV DNA Polactivity by monoclonal antibody to the BMRF1 protein (2) and thelow activity in DNAPol fraction lackingthe BMRF1 protein (12) strongly suggest that the EBV DNA Pol catalytic subunit forms a tight complexwith the BMRF1 proteinin theEBV-infected cells to function asEBV Polholoenzyme. The EBV DNA Pol activity is stimulated by ammonium sulfate when acti-vated DNA is used as template-primer and is inhibited by aphidicolin or phosphonoacetic acid (PAA) (11, 24, 25). Poly(dC)- oligo(dG) isagoodtemplate-primerfor the EBV DNA Pol (11, 25). Furthermore, the EBV DNA Pol can efficiently extend both RNA and DNA primers on the template DNAwithout ATP hydrolysis and exhibits strik-ingly high processivity, which is a desirable feature in the synthesis ofmultiple copiesofthe EBV genome in rolling-circle DNAreplication (25). More recently, 3'-to-5'
exonu-clease activityhas been demonstratedtobe associatedwith the purifiedEBV DNAPol, whichliberates 5'-deoxynucle-oside monophosphates from primer termini (24,
26).
The exonucleaseactivityis also stimulatedbyammonium sulfate and is proposed to have aproofreading
function as it preferentially excises terminally mismatched nucleotides incorporatedattheprimerterminus. Theseenzymatic
prop-ertiesarecommon toDNAPols in theherpesvirus
family.
Ininvestigationsof themolecular basis of
protein-protein
interactions between the subunits of the EBV DNA Pol holoenzyme, it is very usefultodevelopthe
overexpression
andpurificationsystemsof the individual components andto characterize them. EBV DNA Pol accessory
subunit,
BMRF1geneproduct,hasrecentlybeen
expressed,
purified,
on November 9, 2019 by guest
http://jvi.asm.org/
and characterized (27). The accessory protein exhibited higher binding affinity for double-stranded DNA but had neither DNA Polnorexonucleaseactivities. Herewereport theoverexpression of EBV DNA Polcatalyticsubunit in the recombinantbaculovirus-infected insect cells in ordertoget sufficientamountsoffunctionally activeBALF5 gene prod-uct without cross-contamination of the EBV DNA Pol accessorysubunit. Thepurificationand theenzymatic prop-erties of the catalytic EBV DNA Pol aredescribed.
MATERIALSANDMETHODS
Materials.
[3H]TTP
(50 Ci/mmol), [1',2'-3H]dGTP (50Ci/
mmol), [1',2',2,8-3H]dATP (100 Ci/mmol), and unlabeled deoxynucleoside 5'-triphosphates were from Amersham Corp. Unlabeled polynucleotides [poly(rA), poly(rU), poly(dA), and poly(dC)] and oligonucleotides
[(dT)1218,
(dG)1218,
(dA)1218,
and(rA)12_18]
were from Pharmacia LKB Biotechnology Inc.(dT)6,00
was from the Midland Certified Reagent Co. Heparin agarose andsingle-stranded DNA agarose were from Bethesda Research Laboratories. PhagemidpBS(+)wasfromStratagene. ThepVL1392trans-fer vector was a gift from Max D. Summers (Texas A&M University).
DNAsubstrate.Activated calfthymus DNAwasprepared asdescribedpreviously (25).
Template-primers used in thetemplate-primer preference assay were prepared as follows. An oligonucleotide was
hybridized at a ratio of 1:2 (wt/wt) to the complementary polynucleotide in buffer B (20 mMTris-HCl [pH8.0],5 mM MgCl2, and 0.3 MNaCl) byincubation at 70°C for 20 min, slow cooling to room temperature over 1 h, and further incubation for 1 hat 30°C.
3'-terminally labeled activated calf thymus DNA was
prepared by incubating activated DNA with the Klenow fragmentofEscherichia coliPol I in thepresenceof asingle nucleotide substrate,
[3H]dGTP.
The reactionmixturecon-tained 50 mM Tris-HCI (pH 8.0), 6 mM MgCl2, 1 mM dithiothreitol(DTT),160 ,ugofactivatedDNAperml, 6 ,uM
[3HjdGTP
(50Ci/mmol), 0.8 mM 5'-AMP, and 40 U of E. coli Pol I Klenowfragment per ml. After 30 min of incubationatroom temperature, the reaction wasstopped by the addition of100 mM EDTA and0.5% sodium dodecyl sulfate (SDS) anddeproteinizedwithphenol-chloroform.Activated
DNA.
[3H]dGMPwas separated fromunincorporated nucleotides bycentrifugation throughaspuncolumn ofChromaSpin-30 (Clontech Laboratory, Inc.). The specific activity of 3'-terminally labeled activatedDNA.
[3H]dGMP
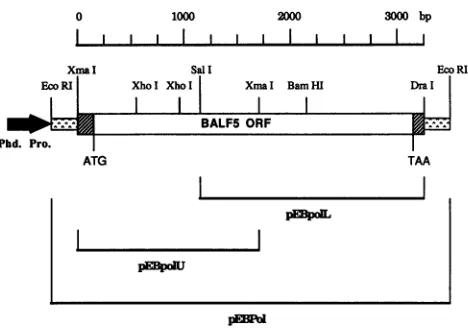
was 144,000 cpm/ilg.Construction of recombinantbaculovirus. Acosmid clone containing the EBV(strain Akata) DNA Pol open reading frame (a gift from K. Takada, YamaguchiUniversity) (23) was used to prepare a recombinant plasmid that would permit overexpression ofthe EBV DNA Pol catalytic sub-unit,BALF5protein. TwooverlappingEBV DNAPol gene clones,pEBpolU andpEBpolL, were prepared (Fig. 1). The plasmid pEBpolU contains the XmaIII (blunt)-XmaIII (blunt) fragment including the upper part of the EBV Pol openreading frame atthe HincII site of the pBS(+) vector. The plasmid pEBpolL contains the SalI-DraI (blunt)
frag-mentincluding thelower part of the EBV Pol open reading framebetween the Sall and SmaI sites of the pBS(+) vector. To generate pEBPol containing the full-length EBV Pol gene,theSalI-PvuI(blunt)fragment ofpEBPolLwasligated between the HindIII(blunt)andSallsitesofpEBPolU. The DNAfragment containing afull-length copyof the BALF5
0 1000
Eco RI
Phd. Pro.
2000 3000 bp
ma I Sal I Eco
I Xho I Xho I Xma I Bam HI Dm I
BALF5 ORF
ATG
RI
TM
pEBp)L
FIG. 1. Schematic representation of theEBV DNA Polgeneof the recombinantbaculovirusvectorpVL1392/BALF5.Thediagram showsa restriction map of theEBV DNA Polregion ofpVL1392/
BALF5.Open region, BALF5 openreading frame; hatchedregions, upstream and downstreamregions of BALF5 openreading frame;
dottedregion, multiplecloning site ofpBS(+) DNA; blackregion, polyhedrin promoter of pVL1392. Details of the construction ofthe recombinant transfervector aregiven in Materials and Methods.
genewasisolatedfrom theplasmidpEBPol by digestionwith EcoRI and inserted into the EcoRI site of the baculovirus transfer vector, pVL1392 (18), to generate a recombinant plasmid, pVL1392/BALF5.The correct orientationwas ver-ified byrestriction-mappinganalyses. The recombinant plas-mid pVL1392/BALF5 wastransformed into E. coliHB101 and isolated. The
pVL1392/BALF5
transfervector (60 ,ug) wascotransfected with theSauI-digestedlinear baculovirus DNAAcRP23LacZ (1 ,ug) (21) intoSpodopterafrugiperda
cells (Sf9 cells) as described by Kitts et al. (13). Viruses producedfrom the transfected cells were used to infectSf9 cells,andtheplaqueswerestained with 50 ,ug of neutral red perml and 250p,gof 5-bromo-4-chloro-3-indolyl-,-D-galac-topyranoside (X-Gal) per ml at 3 days postinfection. The recombinant baculovirus plaques were distinguished from theoriginaltypeofbaculovirusplaques bythe lack of blue in theinfected cells. The recombinantbaculovirus,AcBALF5,
waspurified by five rounds ofplaquepurification.
Preparation of infected cellextracts.Sf9 cells infected with the recombinantbaculovirusAcBALF5 at a multiplicity of infection of 1 were harvested at 36 h postinfection and centrifugedat 1,400 xgfor 10 min. The cellswerewashed in ice-cold phosphate-buffered saline (PBS) and recentri-fugedasdescribed above. The cellpelletwasresuspendedin ice-coldhypotonic buffer[20mMTris-HCl (pH7.6), 5 mM EDTA, 1 mM ethylene glycol-bis(,-aminoethyl ether)-N,N,N',N'-tetraacetic acid (EGTA), 1 mM DTT, 1 mM PMSF,4 ,ugofleupeptinperml,and4,ugofpepstatin A per ml] for 30 minat0°Cand thensubjectedto Dounce homog-enization. Nucleiwere removed fromthe lysate by centrif-ugationat1,500 xgfor 10 minat4°C.Thecytosolicextract wasfrozen inliquidnitrogen and storedat-80°C until itwas tobeused.
Purification of theEBV DNAPolcatalytic subunit.Unless otherwise noted, all steps were performed at 4°C. The cytosolicextractwasquicklythawed at37°Candappliedto a column ofheparin-agarose (3.0-ml bedvolume) that had been equilibrated with buffer A (25% glycerol, 20 mM Tris-HCl[pH 7.6], 1mMEDTA, 1 mMEGTA, 1mMDTT,
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.311.545.66.231.2]1 mM
phenylmethylsulfonyl fluoride,
4 ,g ofleupeptinperml,
and 4 ,gofpepstatin
A perml) containing
0.1 MNaCl. The columnwaswashed with20 mlofthe samebuffer and eluted witha36-ml lineargradientfrom 0.1to1.0 MNaClin buffer A. Fractions(0.6 ml)
were assayed for DNA Polactivity,
and somealiquots
were subjected toSDS-poly-acrylamide gel electrophoresis (PAGE) (8%
polyacrylamide) and stainedwith silveroranalyzed by
Western blot(immu-noblot)
with anti-BALF5protein-specific
antibody. Peak fractionswerepooled
anddialyzed
overnight against buffer Acontaining
0.1 MNaCl.The
dialysate
wasapplied
to a column ofsingle-stranded
DNA agarose
(1-ml
bedvolume) equilibrated
with buffer Acontaining
0.1 MNaCl. Thecolumnwaswashed with 6 ml of thesamebuffer,
andalineargradient
(22 ml)
from 0.1to0.8 M NaCl inbuffer Awasapplied.
Fractions(0.36
ml)wereassayed
for DNA Polactivity
andsubjected
to SDS-PAGE(8% polyacrylamide)
and stained with silver. Peak fractions werepooled
anddialyzed overnight against
buffer Acon-taining
0.1 MNaCl.The
dialysate
wasapplied
to acolumn ofheparin-agarose(0.4 ml) equilibrated
withbuffer Acontaining
0.1 M NaCl. The columnwaswashed with 3 ml ofthe same buffer and eluted withan8-ml lineargradient
from 0.1to0.6 M NaClin buffer A. Fractions(0.2 ml)
wereassayed
for DNA Polactivity.
Peak fractions werepooled,
concentratedby
cen-trifugation
inaCentricon 30microconcentrator,
and stored at-80°C.
Thespecific activity
of the EBV DNA Polcatalytic
subunitwas6,190,000
U/mg.
Preparationof EBV DNAPol fromEBV-producing lympho-blastoid cells. EBV DNA Pol
complex
waspurified
from B95-8cellstreated withphorbol 12-myristate
13-acetate and sodiumn-butyrate
asdescribedpreviously
(24).
Thespecific
activity
of thepurified
EBV DNAPol was1,317,000 U/mg,
whichwascorrected
by
thedefinition of thisstudy. Enzymeassays.DNAPolactivity
wasroutinely
assayedin the absenceof ammoniumsulfate.Each reaction mixture(25
,ul)
contained50 mMTris-HCI(pH 8.0);
10%glycerol;6 mMMgCl2;
100 ,gof bovineserumalbumin(BSA)
perml;80 ,ug of activated calfthymus
DNA perml;
1 mMDTT;
25 ,uM(each)
dATP,
dCTP,
andTTP;
and 10,uM
[3H]dGTP (900
cpm/pmol).
The reactionwas startedby
the addition of the enzymefraction,
andincubationwasfor20 minat35°C.One unit of enzymeactivity
wasdefinedasthe amountrequired
for
incorporation
of 1pmol
ofdeoxynucleoside
monophos-phates
into acid-insoluble material in 60 minat35°C.The3'-to-5' exonuclease
activity
wasassayed bymeasur-ing
the release of[3H]dGMP
from3'-terminally
labeled activatedDNA.
[3H]dGMP
(144,000 cpm/,ug)
as describedpreviously (26).
Each reactionmixture(25 ,ul)
contained 50 mMTris-HCl(pH 8.0),
6 mMMgCl2,
1mMDTf,
100 ,ug of BSA perml,
and3 ,ug of activatedDNA.
[3H]dGMP
perml. The reaction was startedby
the addition of the enzyme, incubated for 5 minat35°C,
andstopped by
the addition of 10 ,ul of 2 mg of sonicated salmon sperm DNA per mlcontaining
0.01 M sodiumpyrophosphate
and 100 ,ul of100%(wt/vol)
trichloroacetic acid. The reactionmixturewas lefton ice for 10 min and
centrifuged
at11,000
xgfor 10 min. One hundred microliters of thesupernatant
wasaddedto1.5 mlof ACS II(NEN),
and then theradioactivity
wascounted. Antibodies. Ananti-BALF5protein-specific antibody
wasproduced against
afusionprotein consisting
ofT7
phage4i10
protein (280
aminoacids)
and a truncatedregion
of EBV BALF5protein containing
a 3'-to-5' exonuclease domain and a nucleotidebinding
domain(361
aminoacids)
as fol-lows. The SalI-BamHIfragment
frompEBPolwasinsertedin frame between theSalI and BamHI sites of the expression vector pGEMEX2 (Promega). The recombinant plasmid pGEMEX2/BALF5 was transformed into BL21 (DE3), an
E. coli cell line which expresses T7 RNA Pol. The BALF5 fusion protein was expressed in E. coli harboring pGE MEX2/BALF5 by induction with 1 mM isopropyl-1-thio-,B-D-galactopyranoside (IPTG). The E. coli cell pellet was collectedandsuspendedin 10 volumesoflysis buffer (10 mM sodiumphosphate [pH 7.5], 30 mM NaCl, 0.25% Tween 20, 10 mM 2-mercaptoethanol, 10 mMEDTA, 10 mM EGTA) andlysed by freeze-thawing and sonication. After centrifu-gation at 12,000 x gfor 30 min, the cell pellet was resus-pended with sample buffer for SDS-PAGE. The soluble sample was then subjected to SDS-PAGE. The band of BALF5 fusion protein was excised, and the protein was
extracted with a buffer containing 25 mM Tris, 195 mM glycine, and0.1% SDS. The purity of the obtained BALF5 fusionproteinwas morethan90%asjudged bySDS-PAGE. AJapaneseWhiterabbitwasimmunized subcutaneously by
astandardprotocol(9)with thepurifiedfusion proteinseven
timesatbiweeklyintervals. ImmunoglobulinG (IgG) of the immunized rabbit serum was purified through the affinity columncoupledwith thepurified BALF5 fusion protein as described previously (9). The protein concentration of the anti-BALF5protein-specific antibodywas200p.g/mlinPBS. An anti-BBLF4 protein-specific antibody was raised against BBLF4 carboxy-terminal oligopeptides (20 amino acids) coupled tokeyhole limpet hemacyanin. Preparation for the anti-BBLF4 protein-specific IgG was performed accordingtothesameprocedureasthat for the anti-BALF5 protein-specific antibody. The protein concentration of the anti-BBLF4-specific IgGwas 1 mg/ml in PBS.
RESULTS
Expression of the EBV DNA Pol catalytic subunit in Sf9 cells.MonolayersofSf9 cellsweremockinfectedorinfected
at a multiplicity of infection of 5 PFU per cell with AcRP23LacZ or the recombinant baculovirus AcBALF5. The protein products were resolved by SDS-PAGE (8% polyacrylamide) (Fig. 2A).TheAcRP23LacZ-infected insect cells expressed large amounts of ,-galactosidase with a
molecularweightof116,000 (Fig. 2A, lane2). By
compari-son with the controls (mock-infected and AcRP23LacZ-infected cells), a 110-kDapolypeptide was detected in the AcBALF5-infectedcells. Theexpressed proteinwasshown
to be EBV BALF5 gene product by its reaction with anti-BALF5protein-specific antibody duringWestern immu-noblotanalysis (Fig. 2B). Theexpressed BALF5gene
prod-uctmadeupabout20%of the total cellularproteins. Purification of the EBVDNAPol catalyticsubunitexpressed
in
Sf9
cells. A monolayerculture of Sf9 cellswas infected withthe recombinant baculovirus AcBALF5. At 36 h postin-fection, the cells were harvested, suspended in hypotonic buffer for 30 min at 0°C, and then subjected to Dounce homogenization. Alarge part of the expressedEBV DNA Pol catalytic subunit was transported to the nuclei of the infected Sf9cells,but itwasalmostinsoluble. No DNA Pol activitywasassociatedwith the insoluble material (datanot shown). However, the cytosolicfraction contained a smallamount ofsoluble enzyme asjudged byWestern blot anal-ysis with anti-BALF5 protein-specific antibody. Thus, the cytosolicfractionwasloadedonto a
heparin-agarose
column and eluted with a linear gradientfrom 0.1 to 1 M NaCl in bufferA.The recombinantBALF5 geneproductof 110 kDa eluted at 0.36 M NaCl asjudged
by silverstaining
ofon November 9, 2019 by guest
http://jvi.asm.org/
A
E
M
1
2
3
200 we *
116 97 70
43
DF
FIG. 2. Expression of the BALF5gene prc
cellswere mock infected (lanes 1) orinfecte (lanes 2) (20) or the recombinant baculovirus
andharvested at3dayspostinfection. Protein
cellextracts wereresolvedbySDS-PAGE (89
stained with Coomassie blue (A) oranalyzed
with anti-BALF5 protein-specific antibody. '
molecularweight standards (lane M) (inthous; the left ofpanel A. The position of the BA indicatedattheright of eachpanel (closedtri
SDS-PAGE and Western blot analyses
DNA Pol activity was complicated
b3
cellular, baculovirus, and the expressed single infected cell.
The peak fractions were loaded ontc DNAagarosecolumnand elutedwithali
0.1to0.8MNaClinbuffer A.Single-stra chromatographyyieldedtwopeaksof DN firstmajorpeakofDNA Polactivity,whi
NaCl,wascoincidentwith theexpressed
asjudged bysilverstainingofSDS-PAG]
analyses (data not shown). The DNA I
second minor peak, which eluted at 0.3 coincident with the expressed BALF5
activitywasstimulated threefold in the I
ammonium sulfate, it most likely reflec DNAPolactivity(20).
The peak fractions of the expressed I
againloadedontoaheparin-agarosecolui
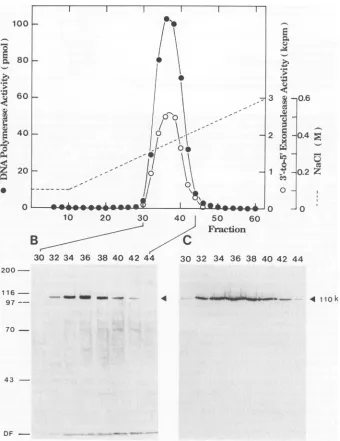
a linear gradient from 0.1 to 0.6 M Nal shown inFig. 3,thepeakofDNA Polacd
M NaCl, and the silver-stained polyacr
fractionscontainingthe Polactivityshow band that had an intensity coincident
enzymatic activity (Fig. 3). The band apparent molecularmassof110kDa,a
v-theexpectedmolecularmass oftheprot
EBV BALFS (113 kDa) gene and wa
mobilityof thecorrespondingsubunitoft fromchemicallyinducedEBV-infectedc emblotanalysis clearlyshowed that the
in factan EBVBALF5 gene product (E maryofthe purificationisgiven inTable
I Detection of a
3'-to-5'
exonucleaseactivity.
The 3'-to-5'exonucleaseactivity is known to be associated with the EBV
1
2
3
DNA Polpurified
fromchemically
induced EBV-infected cells (24). To test whether the single subunit of the EBV DNAPol has the 3'-to-5' exonuclease activity, the activity was assayed foreachfraction. The exonuclease activity was precisely coincident with the 110-kDa protein band of the expressed EBV DNA Pol catalytic subunit on the second*4
*step
ofchromatography
ofsingle-stranded
DNA agarose (data not shown) and the last step of chromatography of heparin agarose (Fig. 3). The 3'-to-5' exonuclease activity wasassayed by using 3'-terminally labeled activated DNA. [3H]dGMP as substrate. The reaction product was dGMP determined by polyethyleneimine thin-layer chromatogra-phy(data notshown). The 5'-to-3' exonuclease activitywasalso assayed under the same assay condition as the3'-to-5' exonuclease activity except with terminally labeled
5'-32P-poly(dA)255
poly(dT)6,000
as substrate. However, the 5'-to-3' exonuclease activity was not detected under the )duct inSf9 cells. Sf9 condition (data not shown). These observations indicate that dwith AcRP23LacZ the EBV DNA Pol catalytic subunit hydrolyzed3'-terminal AcBALF5 (lanes 3) nucleotide. Thus, we conclude that the3'-to-5' exonuclease 5srecovered from the activity associated with the EBV DNA Pol is an intrinsicpolyacrylamide) and feature of the EBV BALF5 gene product.
Iby Western blot (B) Inhibition of DNA Pol and 3'-to-5' exonuclease activities by
ands) are indicated at an antibody raised against BALF5 fusion protein. An
anti-LFS
gene product is BALF5protein-specific antibody
wasraised against a fusion angle). proteinconsisting of T7 4)10 protein and a truncatedregion ofthe BALF5protein including the 3'-to-5' exonucleaseand a part of nucleotide binding domains. The antibody was tested for its ability to neutralize the Pol and the 3'-to-5' exonuclease activities. The purified catalytic subunit of EBV (data not shown). DNA Pol was incubated with the antibody at 0°Cfor 30min
y the presence of and then assayed for these activities. As shown in Table 2, EBV enzymes in a the antibody inhibited both the Pol and the exonuclease activities to the same degree, whereas anti-BBLF4
protein-a single-stranded specific antibody did not. These results are consistent with inear gradient from the view that both DNA Pol and 3'-to-5' exonuclease activ-nded DNA agarose itiesarepresent onthesame polypeptide.
[APol activity. The Enzymaticproperties of the single subunit of the EBV DNA ch eluted at 0.18 M Pol. (i) Reactivity to aphidicolin and PAA. EBV DNA Pol is EBV geneproduct distinct from DNA Pol a for its hypersensitivity to the E and Westernblot pyrophosphate analog, PAA. Another compound, aphidi-Pol activity of the
colin,
a potent DNA synthesis inhibitor in vivo, inhibits all M NaCl, was nota-like
DNAPols in vitro(15).
The EBV DNA Polpurified
rotein. Since the from EBV-producingcellsis known tobe sensitive to bothpresence
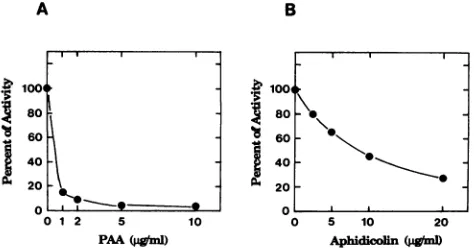
of 80 mM PAAand aphidicolin (11, 25). The single subunit of the EBV DNA Pol was assayed for DNA Pol activity in the presencected
a baculovirus ofincreasing
concentrations of PAA or aphidicolin (Fig. 4).The single subunit of the EBV DNA Pol was also sensitive to EBV enzyme were these compounds.
nnand eluted with (ii) Salt
sensitivity.
It is well known that the Pol activityCl in buffer A. As associated with DNA Pols in the herpesvirus family is tivityeluted at 0.36 stimulated by salt when activated DNAis used as
template-ylamide
gel ofthe
primer in the Pol assay (11, 14, 24). To know the salt effectred
a single distinct on the DNAPol and the3'-to-5'exonuclease activities of the with that of the EBV DNA Pol catalytic subunit, both activities wereas-migrated with an sayed in the presence of increasing concentrationsof
ammo-alue that is close to nium sulfate (Fig. 5). Both the Pol and the 3'-to-5' exonu-einencodedby the clease activities were inhibited by the salt in a dose-Is identical to the dependent manner, whereas both activities associatedwith
he enzyme isolated the enzymepurified fromEBV-producing lymphocytes were ells (11, 24). West- stimulated by the salt.
purifiedenzyme is (iii) Template-primer preference. One of the conspicuous Fig. 3C). The sum- enzymatic properties of herpesvirus DNA Pols is template-1. primer preference for poly(dC). oligo(dG) (11, 25). The
ls".Ijg--
.,:".'$-qoi4- .4w-''I-' 144P
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.57.296.71.255.2]100
-C4 80
-*5:
36040
0
20
S
0
10 20 3
B
30 32 34 36 38 40 42
200
-a _a
0 40 50 60
1
F'~Faction
C
44 30 32
4
E
C4
C.)
t
13
2
X
-1-I
3 v
-0Z
o
o"
1 0.6
-1-1
10.4 h2 Q 0.2 Z
0
34 36 38 40 42 44
- 4 110k
FIG. 3. The last step inchromatography of theheparin-agarose column of theBALF5 Pol catalytic protein. The peak fractions of the single-strandedDNAagarosecolumnchromatographywereloadedontoaheparin-agarose column and eluted withalineargradient from 0.1
to0.6 MNaClin buffer A.Fractionswereassayed for either DNA Polor3'-to-5' exonucleaseactivity. Resultsareexpressedaspicomoles
ofdGMPincorporatedin 20min (DNAPolactivity),orcountsperminute of released dGMP in 5min (3'-to-5' exonuclease activity). Aliquots
of theindicated fractions from thechromatographyweresubjectedtoSDS-PAGE (8% polyacrylamide) and stained with silver (B)oranalyzed
byWestern blot withanti-BALF5 protein-specific antibody (C). The positions of the molecular weight standards (in thousands)areindicated
atthe left ofpanelB. Thepositionsof theBALF5protein areindicatedattheright of panels B and C (closed triangles).
EBV DNA Pol purified from EBV-producing cells utilized
poly(dC). oligo(dG) 29-foldmore efficiently than activated DNA as template-primer, while the single subunit of the
EBV DNA Pol utilized it poorly (Table 3). However, poly(rA). oligo(dT), poly(rU) oligo(dA), and poly(dT)
were not utilized at all as well. Thus, the absence of the BMRF1 Polaccessory protein drastically changedthe tem-plate-primer preference. The BMRF1 protein may have a
nucleotidesequencespecificityfor its double-stranded DNA
binding and influence the Pol andprimer-terminus interac-tion.
DISCUSSION
We haveexpressedthe EBV DNA Polcatalyticsubunitin insectcellsby usingthe baculovirus system. Up to 20% of the total cellular proteins produced in the infected insect cells is the recombinant EBVDNA Polpolypeptide. Unfor-tunately, almost-expressedEBV Polproteinremainedas an
insolublefraction,whichwasenzymatically inactive,as was
observed for the expressionof humancytomegalovirusand varicella-zoster virus DNA Pols in insect cells(5). However,
the soluble andenzymaticallyactiveEBV DNA Polcatalytic
116 97
--70
-43
-OF
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.150.490.70.511.2]TABLE 1. Summary of purification of the BALF5 protein expressed in SF9 cells
Fraction Ste Total protein TotalPolactivity Spact Yield
no. P (mg) (U) (U/mg)a (%)
I Cytosolicextract 49 3,400,000 69,400 100
II Heparin-agarose 2.7 1,240,000 459,000 36
III Single-stranded DNA-agarose 0.17 503,000 2,960,000 15
IV Heparin-agarose 0.03 187,000 6,170,000 5
a1U isdefinedas1 pmol of deoxynucleosidemonophosphate incorporation for60min.
subunit was recovered from the cytosolic fraction of the recombinant virus-infected insect cells. The levels of the soluble EBV Polcatalytic protein reached maximum prior to 48 hpostinfection. After 72 hpostinfection,mostof the EBV Pol protein became insoluble. So, the optimal time for isolation of enzymaticallyactive EBV Pol was from 36 to 48 hpostinfection.
In a previous report (24), it has been reported that the 3'-to-5' exonucleaseactivity is associated with the 110-kDa subunitof the EBV DNAPol. However, the Polpreparation contained additional 48- to 50-kDa polypeptides. Anti-BMRF1 protein-specific monoclonal antibody recognized these 50- and 48-kDapolypeptides(28).ThepurifiedBMRF1 protein byitself exhibits no exonucleaseactivity (27). How-ever, there remained a possibility that the BMRF1protein can exhibit 3'-to-5' exonuclease activity only when the BMRF1proteinforms a complexwiththe BALF5 protein. The copurification of the exonuclease activity and appar-ently homogeneousEBVBALF5proteinlacking otherEBV proteins permitsus toconcludethat the 3'-to-5' exonuclease activity is intrinsictothe EBV Polcatalyticpolypeptide,as
istruewith HSV DNA Pol catalytic subunit(19).
Others havereportedtheexpressionofactiveEBV DNA Polcatalytic subunit by in vitrotranscription-translation
(12,
17). Kiehl and Dorsky (12) have shown that the EBV Pol catalytic subunitaloneexhibitsnoPolactivity;however,the expression system of both EBV Pol and BMRF1 protein generatedDNAPolactivityinthereticulocyte
lysate.
Since their Pol assay reaction mixture contained 100mMammo-nium sulfate, DNA Pol activity of the EBV Pol catalytic subunit alonemightbe inhibitedcompletely bythehighionic strength. In contrast, Lin et al. (17) havereported that the Polactivityof the EBVPolcatalyticsubunitwasresistantto
ammonium sulfate; however, salt-stimulated Pol activity was not observed. The discrepancy between them is not
known withcertainty. Thereticulocyte lysate contains cel-lularDNA Pols and many other proteins. It is possible to speculate that those factors may mask theenzymatic
prop-TABLE 2. Effect of anti-BALF5protein-specific antibodyonthe Poland the3'-to-5'exonuclease activities of the EBV DNA
Polcatalytic
subunit'
DNAPol 3'-to-5'exonuclease
Additive activity activity
pmol % cpm %
BSA 84 4 100 1,826 88 100
Anti-BALF5IgG 50 + 5 54 785 + 78 43 Anti-BBLF4IgG 91 ± 7 109 1,862 - 63 102
aPriortothe assay,8 ,ul of thepurifiedBALF5proteinwasincubated with 8,ulof anti-BALF5protein-specific antibody (200 jig/ml),anti-BBLF4
pro-tein-specificantibody (1 mg/ml),orBSA(200jig/ml)at0'Cfor30 min.The Pol and the exonucleaseactivitieswerethendeterminedasdescribedin Materials and Methods.Resultsarethe means ±standard errors of the mean(n= 3).
erties of the expressed EBVPol catalytic subunit. Sincethe EBV Polcatalyticsubunitwaspurifiedtonearhomogeneity
in this study, it was suitable to examine for its specific activity on activated DNA. The addition of ammonium
sulfate reduced the Pol activity of the EBV DNA Pol
catalyticsubunitdrastically, whiletheactivityof theenzyme
purified from EBV-producing lymphocyteswas stimulated
(Fig. 5). It might be possible to speculate that thebinding affinityof the Polcatalyticsubunit for theprimerterminus is weak and that highionic strength destabilizes the catalytic protein-primerterminus interaction.The BMRF1 Pol acces-sory protein may increase the affinity of the Pol for the
primerterminusbyits double-stranded DNAbinding
activ-ityand decrease the dissociation of the Pol from thetemplate
DNA. Further biochemical studies are needed to clarify
thesespeculations.
Most replicative DNA Pols consist of a Pol catalytic
subunit and Pol accessoryproteinsand display highly
pro-cessivepolymerization (15). BacteriophageT7 DNA Pol is
composedofgene 5 protein (gp5) and E. coli thioredoxin. TheT7 DNA Polcatalytic subunit,gp5,alone hasverylow Pol and 3'-to-5' exonuclease activities (22). An accessory
protein to gp5, E. coli thioredoxin, is near-absolutely
re-quiredfor both the Pol and the3'-to-5' exonuclease activities in vitro by extending the lifetime of a gp5 complexwith
template-primerfrom less thanasecondtoabout5minand
by increasingthe processivityofpolymerization.
Bacterio-phageT4 DNA Pol is composedofagp43Pol coreprotein
and three distinctive accessory proteins (gp44-gp62 and
gp45) to help it function as a holoenzyme, while ATP
hydrolysis is needed to form holoenzyme at the primer
terminus (30). Unlike the case of T7 DNA Pol catalytic
subunitgp5,thegp43 proteinalone hashighPol and 3'-to-5'
A
. 100
801
g 60 f 40
20
0
B
1004
80 60 40 20
0 1 2 5 10 0 5 10 20
[image:6.612.59.546.82.148.2]PAA Wml) Aphidicolin(4InI
FIG. 4. Effect of PAA(A)andaphidicolin (B)onthe Polactivity
of the EBV DNAPolcatalytic subunit. TheDNAPolactivitywas
assayedonactivated DNAtemplateas described in Materials and
Methods, supplementedwith PAAor aphidicolintothe final
con-centrations indicated. The 100% value of the EBV DNA Pol catalyticsubunitwas60pmolofdeoxynucleosidemonophosphates incorporatedper20min.
0"'I~~o
I,
----r
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.310.547.527.651.2] [image:6.612.51.291.604.673.2]A
200
t 150 1004 50
DNAPbbmeraseAcvity
B
120
100
80
60
40 20 0
0 50 100 150 0 50 100 150
(NE14)29S (mM) (NHOZSOs (mM)
FIG. 5. Effect of ammonium sulfateon the Pol and the 3'-to-5'
exonuclease activities of the EBV DNA Pol catalytic subunit (0)
and the EBV DNA Polpurified from EBV-producing lymphocytes
(0).(A) The DNA Pol activitywasassayed by using activated DNA as substrate as described in Materials and Methods except that ammonium sulfate concentrations were varied as indicated. The
activity is expressedasthepercentageof the activity obtained inthe absence of ammoniumsulfate.(B) The 3'-to-5' exonuclease activity
was assayed by using activated DNA- [3H]dGMP as substrate as
describedin Materials and Methodsexceptthat ammonium sulfate
concentrationswerevariedasindicated. The activity is expressedas
thepercentageof the activity obtained in the absence ofammonium
sulfate.
exonucleaseactivities. Theaccessoryprotein complex
stim-ulates theprocessivity ofthe gp43protein.Human DNAPol
ais madeupofacluster ofa180-kDa Pol catalyticsubunit,
a 70-kDa phosphoprotein of unknown function, and two
polypeptides of 55 and 49 kDa with primase activity (15). The DNA Pol a catalytic subunit has been functionally
expressed by arecombinant baculovirus in insect cells (3).
The catalytic polypeptide exhibits Pol activity but has no
3'-to-5' exonuclease activity. The kineticparameters of Pol catalysis, sensitivitytoinhibitors, andprocessivity of the Pol catalytic subunit are identical to that observed with the four-subunit Pol a-primase complex. HSV type 1 DNA Pol also consists of UL30 Pol catalytic subunit and UL42 Pol
accessorysubunit (4). The HSV DNA Pol catalyticsubunit
[image:7.612.68.306.73.218.2]has been alsooverexpressed in the baculovirus system (10, 19). The HSV Pol is resistanttosalt althoughsalt-stimulated
TABLE 3. Template-primerpreferences for the catalytic subunit
orthecomplex form of theEBVDNAPor
EBV DNAPol(%) Template-primer
Catalytic subunit Pol complex
Activated DNA 100 100
Poly(dC) oligo(dG) 48 2,900
Poly(dT) oligo(rA) 190 670
Poly(rU) oligo(dA) <1 <1
Poly(rA) oligo(dT) <1 <1
Poly(dT) <1 <1
a AllDNAPolassayscontained 50 mMTris-HCl (pH8.0), 6mMMgC92,1
mM DTr,100,ugof BSAperml, 10% glycerol, 40IM3H-deoxynucleoside triphosphate complementary to the template, and the indicated
template-primer. Reactions were performed for 20 min at 35'C. All templates and
primerswerepresent at80 and 40,ug/ml, respectively. Thetemplate-primer concentration in reaction with activated DNAorpoly(dT)was80pg/ml.All
assays wereperformed in triplicate,andthe datarepresent themeanvalues
relativetothat withactivated DNA.
Pol activitywasnot observed (19). Furthermore, its chro-matographicbehaviors are similar tothose for the enzyme
purified from HSV-infected cells (19). Although UL42 Pol
accessory protein does not change the HSV DNA Pol
activityon activated DNAtemplate, it stimulates the
pro-cessivityoftheHSVPolcatalytic subuniton singly primed
M13single-stranded DNA template (7, 10, 19).In thecaseof EBVDNAPol, theEBV Polcatalytic subunit alone exhib-itedboththe DNAPoland the3'-to-5' exonucleaseactivities
in the absenceofBMRF1 Polaccessoryprotein. However, the EBVDNAPolcatalytic subunitwasdistinguishedfrom theenzymepurifiedfromEBV-producingcellsinsalt
sensi-tivity, template-primer preference, and chromatographic properties (24). The functional expression of the EBV Pol
catalytic subunit provides uswith anamenable system for
pursuingstructure-function studies. For example, it isvery
interesting to know how many nucleotides the EBV Pol
catalyticsubunitalonepolymerizes and how theBMRF1 Pol
accessoryproteinchanges the enzymatic activityof the Pol
catalytic subunit. In vitrostudy of the interactions between
the BALF5Polcatalytic protein and the BMRF1Pol acces-soryproteinwithregardtothebinding affinityfor theprimer terminus andtheprocessivity isnowin progress.
ACKNOWLEDGMENTS
WearegratefultoK.Takada, YamaguchiUniversity,for provid-ingtheDNAlibrary of the EBVgenome.Wethank M. D.Summers, TexasA&M University; M. Dodson, Stanford University; Y.Ito, Mie University; S. Morikawa, NIH, Japan; and Y. Fujii, Nagoya University, for discussionsabout the baculovirusexpressionsystem andforproviding cells, virus,andvectors.We thank K.Maeno and I. R. Lehman forencouragement.
T.T.was supported by agrant-in-aid from the Ministryof
Edu-cation, Science and Culture of Japan, by the Uehara Memorial
Research Foundation, by the Ishida Foundation, andbythe Nitto
Foundation.
REFERENCES
1. Baer, R., A. T. Banlier, M. D. Biggin, P. L. Deininger, P. J.
Farrell, T.J.Gibson, G.Hatfull, G. S. Hudson,S. C.Satchwell, C. Seguin, P. S. Tuffnell, and B. G. Barrell. 1984. DNA
sequence and expression ofthe B95-8 Epstein-Barr virus
ge-nome.Nature (London) 310:207-211.
2. Chiou,J.F., J.K. KLi,and Y.-C.Cheng.1985. Demonstration
ofastimulatory protein for virus-specified DNApolymerase in
phorbol-ester treated Epstein-Barr virus carrying cells. Proc.
Natl. Acad. Sci. USA 82:5728-5731.
3. Copeland, W. C., andT. S.-F.Wang. 1991.Catalyticsubunit of
human DNA polymerase a overproduced from
baculovirus-infectedinsect cells. J. Biol. Chem. 266:22739-22748.
4. Crute, J. J., and I. R. Lehman. 1989. Herpessimplex-1 DNA
polymerase. Identification ofan intrinsic 5' to 3' exonuclease
withribonuclease H activity. J. Biol. Chem.264:19266-19270.
5. Ertl, P. E., M. S. Thomas, andK. L Powell. 1991. High level
expression of DNA polymerases from herpesviruses. J. Gen.
Virol. 72:1729-1734.
6. Fixman, E. D., G. S. Hayward, and S. D. Hayward. 1992.
trans-acting requirements for replication ofEpstein-Barrvirus
ori-Lyt. J. Virol. 66:5030-5039.
7. Gottlieb, J., A. I. Marcy, D. M. Coen, and M. D. Chaliberg. 1990. The herpes simplex virus type 1 UL42gene product: a
subunit ofDNA polymerase that functionstoincrease
proces-sivity. J. Virol. 64:5976-5987.
8. Hammerschmidt, W., and B. Sugden. 1988. Identification and
characterizationoforiLyt,a lyticoriginof DNAreplicationof
Epstein-Barrvirus. Cell 55:427-433.
9. Harlow, E., and D. Lane. 1988. Antibodies. A laboratory manual. Cold SpringHarborLaboratory, Cold SpringHarbor, N.Y.
10. Hernandez, T.R.,and I. R. Lehman. 1990. Functional
interac-0~~~~
0
WIl
--J-
0-f-
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.66.307.568.656.2]tion between the herpes simplex-1DNApolymerase and UL42 protein.J. Biol. Chem. 265:11227-11232.
11. Kallin,B., L.Sternas, A. K Saemundssen, J. Luka, H. Jornvall, B. Eriksson, P.-Z. Tao, M. T. Nilsson, and G. Klein. 1985. Purification of Epstein-Barr virus DNA polymerase from P3HR-1 cells.J.Virol.54:561-568.
12. Kiehl, A., and D. I.Dorsky. 1991. Cooperation of EBV DNA polymerase andEA-D(BMRF1) in vitro andcolocalization in nuclei of infected cells. Virology184:330-340.
13. Kitts,P.A.,M. D. Ayres, and R. D. Possee.1990.Linearization of baculovirus DNA enhances the recovery of recombinant virus expression. Nucleic Acids Res. 18:5667-5672.
14. Knopf, K.-W. 1979. Properties of herpes simplexDNA poly-merase and characterization of its associated exonuclease ac-tivity.Eur. J.Biochem. 98:231-244.
15. Kornberg, A., and T. Baker. 1992. DNA replication, 2nd ed. W. H. Freeman &Co., NewYork.
16. Li,J..S.,B.-S. Zhou, G. E. Dutschman, S. P. Grill, R.-S. Tan, andY.-C.Cheng. 1987. Association ofEpstein-Barr virus early antigen diffusecomponentandvirus-specifiedDNApolymerase activity.J.Virol. 61:2947-2949.
17. Lin, J.-C., N. D. Sista, F.
Besengon,
J. Kamine, and J. S. Pagano. 1991.Identification and functional characterization of Epstein-Barr virus DNA polymerase byin vitro transcription-translation ofaclonedgene.J. Virol. 65:2728-2731.18. Luckow, V. A., and M. D. Summers. 1988. Trends in the development of baculovirus expressionvectors. Bio/Technol-ogy6:47-54.
19. Marcy, A.I., P. D.Olivo, M. D. Challberg,and D. M.Coen. 1990. Enzymatic activities of overexpressed herpes simplex virusDNApolymerase purified from recombinant baculovirus-infected insect cells. Nucleic AcidsRes. 18:1207-1215. 20. Miller,L. K., J. E. Jewell, and D. Browne. 1981. Baculovirus
induction ofaDNApolymerase. J. Virol. 40:305-308.
21. Possee, R. D., and S. C. Howard. 1987.Analysis of the polyhe-drin gene promoterof the Autographacalifornica nuclear poly-hedrosis virus. Nucleic AcidsRes.15:10233-10248.
22. Tabor, S., H. E. Huber, and C. C. Richardson. 1987. Esche-richiacoli thioredoxin confers processivityontheDNA poly-merase activity of thegene5 protein of bacteriophage T7. J. Biol. Chem.262:16212-16223.
23. Takada, K., K. Horinouchi, Y. Ono, T. Aya, T. Osato, M. Takahashi, and S. Hayasaka. 1991. An Epstein-Barr virus-producerlineAkata:establishment of the cell line and analysis of viralDNA.Virus Genes5:147-156.
24. Tsurumi, T. 1991. Characterization of 3'-to-5' exonuclease activity associated with Epstein-Barr virusDNApolymerase. Virology 182:376-381.
25. Tsurumi, T. 1991. Primer terminus recognition and highly processive replication byEpstein-Barr virusDNApolymerase. Biochem. J. 280:708-713.
26. Tsurumi, T.1992.Selective inhibition of the3'-to-5' exonucle-ase activity associated with Epstein-Barr virus DNA poly-meraseby ribonucleoside monophosphates. Virology 189:803-807.
27. Tsurumi, T.1993.Purification and characterization of the DNA-binding activity of the Epstein-Barr virus DNA polymerase accessoryproteinBMRF1 geneproducts,asexpressed in insect cellsby using the baculovirussystem.J.Virol. 67:1681-1687. 28. Tsurumi, T.Unpublished result.
29. Yates, J. L., N. Warren, D. Reisman, and B.Sugden. 1984.A cis-acting element from the Epstein-Barr viral genome that permits stable replication of recombinant plasmids in latently infected cells.Proc. Natl.Acad. Sci. USA81:3806-3810. 30. Young, M. C., M. K. Reddy, and P. H. vonHippel. 1992.
Structure and function of the bacteriophage T4 DNA poly-meraseholoenzyme.Biochemistry 31:8675-8690.