0022-538X/93/042412-09$02.00/0
Copyright
©
1993, AmericanSociety
forMicrobiology
Analysis
of Nonnucleoside
Drug-Resistant
Variants of Human
Immunodeficiency
Virus
Type
1 Reverse
Transcriptase
PAUL L.
BOYER,"
MICHAEL J.CURRENS,2
JAMESB.
McMAHON,2 MICHAEL R. BOYD,2
ANDSTEPHENH.
HUGHES1*
ABL-Basic
ResearchProgram,
NCI-Frederick CancerResearch andDevelopment Center,
1and
Laboratory of Drug Discovery
Research andDevelopment,
DTP,
DCT,2 National CancerInstitute,
P.O. BoxB,
Frederick,
Maryland 21702-1201
Received 19 October1992/Accepted 10January1993A number of
chemically
distinct nonnucleoside inhibitorsof humanimmunodeficiency
virustype
1(HIV-1)
reverse
transcriptase (RT)
have beenreported.
Several linesofevidence, including
the isolation of RT mutants that showcrossresistance,
suggestthat, despite
theirstructuraldiversity,
manyof these inhibitors bind toacommonsiteon
HIV-1
RT. We haverecently reported that,
onthe basis ofanalysesofHIV-1/HIV-2 chimeras,
the naturalproduct
calanolide Amayinteract withadifferentsiteorsites inHIV-1
RT. We have usedBspMI
cassettemutagenesis
toprepareacollection ofHIV-1
RT mutantsthat show resistance to the knownmembers ofthe general
class of nonnucleoside inhibitors. This collection ofmutantscanbe used to determine whethera new
drug
will showcrossresistance with known inhibitors and to define amino acidpositions
critical for the action of thedrugs.
The mutantswereused toanalyze
calanolideA,
1IH,3H-thiazolo[3,4-albenzimidazole(4i),
and the
acyclic
nucleosideanalog
1-[(2-hydroxyethoxy)methyll-6-(phenylthio)thymine.
Theseanalysessuggest
that all threedrugs
interact withHIV-1
RT within thepreviously
defined commonbinding
site fornonnucleoside
inhibitors.However,
thedrugs respond
differently
tothepanel
ofdrug-resistant HIV-1 RTs,
indicating
that while thebinding
sites of thedrugs overlap they
arenotidentical.The reverse
transcriptase (RT)
of humanimmunodefi-ciency
virus type 1(HIV-1)
has been the target of both nucleosideanalogs,
suchaszidovudine anddideoxyinosine,
and nonnucleosidedrugs, including nevirapine
and the var-ioustetrahydroimidazo[4,5,1-jk][1,4]benzodiazepin-2(1H)-oneand -thione
(TIBO)
derivatives.However, drug-resistant
HIV-1
RT variants have been isolated from clinical materials and fromsamples
of virusinvitro(8, 10-12, 14, 21-23, 26,
28, 32-34, 37).
Nonnucleosidedrugs
arehighly specific
inhibitors of HIV-1 RT andarenotactiveagainst
theRT of theclosely
related lentivirus HIV-2 orother retroviral RTs(6, 7, 9, 11, 13,
18,20, 24, 27, 29, 35).
This suggests that HIV-1 RT containsspecific
structural elements which the other RTs do not possess that interact with nonnucleoside inhibitors. Therecently published
structure of HIV-1 RTcomplexed
withnevirapine
shows that thisdrug
binds to ahydrophobic pocket just
underneath thepolymerase
active site andappearstobeinteracting directly
with thetyrosine
residues
atpositions
181 and 188(19).
Since several other nonnucleoside inhibitors have beenreported
to showcross resistance withnevirapine,
these other nonnucleoside inhib-itorsmaybindtothis siteaswell.Unfortunately,
HIV-1 RT is able to accommodate substantialchanges
in the amino acids that surround this criticalbinding pocket
so thatdrug-resistant
mutants areeasily
obtained.This makes it
especially important
toidentify drugs that bind
to partsof
theenzymethat the virus is
notable
tochange
soeasily.
We haveused
BspMI
cassettemutagenesis
(3, 4)
toconstruct
various HIV-1 RTmutantsthat have been reported
toplay
arole in resistanceto
several nonnucleoside
drugs. Since the mutant enzymesdifferonly
atthe
designated sites of
muta-tion,
differences in resistance tothedrugs
canbeattributed
directly
tothespecific
mutations. This collection ofmutants*Correspondingauthor.
canbe used to test new nonnucleoside inhibitors to deter-minewhether
they
bindtothesamehydrophobic pocket
atwhich the
knowndrugs
appear to bindand
toinvestigate
which
aminoacids
areinvolved inbinding
aparticular drug.
Inthepresentstudy,
these variant HIV-1 RTs have beenassayed
forsensitivity
to two nonnucleosidedrugs,
calan-olide A(18)
and1H,3H-thiazolo[3,4-a]benzimidazole(4i)
(TBA) (6, 7), and
tothe acyclic nucleoside analog
1-[(2-hydroxyethoxy)methyl]-6-(phenylthio)thymine (HEPT) (1,
2, 10, 25). Calanolide
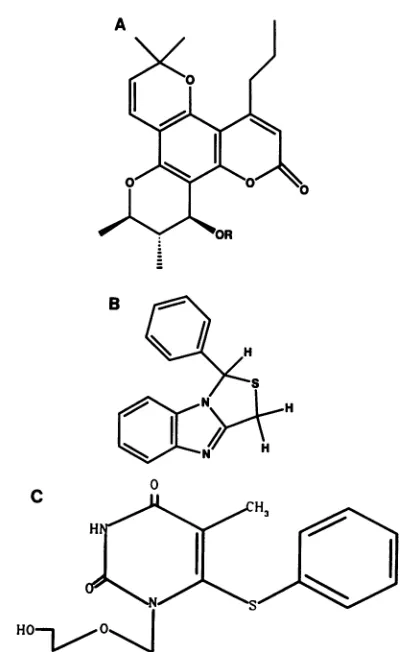
A(Fig.
1A) is
acoumarin
derivative
isolated
from extracts ofthe
fruit and twigs of the
treeCalophyllum lanigerum (18).
Thiscompound
inhibits HIV-1replication
andcytopathicity by noncompetitive inhibition of
HIV-1 RT and is inactiveagainst
HIV-2 RT(18). Calanolide
A isfully
activeagainst
HIV-1strain A17 (26), which
reportedly
is resistanttomanyknownnonnucleoside drugs,
such asTIBO and BI-RG-587(nevirapine). Analysis of the
resistanceand
sensitivity of
aseries of enzymatically active
HIV-1/HIV-2
RTchimeras suggested that calanolide
A does not interact with the same segmentsof HIV-1
RT as the TIBOcompounds (16).
This suggeststhat calanolide
Abelongs
to apharmacologic
classdifferent from that of
the otherknownnonnucleosideRT-inhibitory
drugs(18).
TBA(Fig.
1B)
is asynthetic compound which
also has beenreported
to inhibit HIV-1 RTnoncompetitively
but not toinhibit HIV-2 RT
(6, 7). The
A17strain (26),
however, is resistant to thiscompound, which
suggests that TBA mayfall into the known class of
nonnucleoside inhibitors (5).
HEPT(1, 2, 10, 25) (Fig.
1C) is
anacyclic
nucleoside analog,but,
unlike othernucleoside analogs
suchasdideoxyinosine,HEPT does not appear to
inhibit HIV-1
RT by acting asa
competitive inhibitor
or achain
terminator when poly(rC). oligo(dG)
is used as thesubstrate (1, 2, 10).
HEPTappearstobe
mechanistically similar
tothe class of
noncom-petitive
inhibitors which includesnevirapine and TIBO (10).
Likethe nonnucleosideinhibitors,
HEPTis
inactive against
2412on November 9, 2019 by guest
http://jvi.asm.org/
B
>-S
) t_
FIG. 1. Structures of the nonnucleoside drugs. (A) Calanolide A (15). (B)TBA(4, 5).(C)HEPT(2,8,23).
HIV-2 and other
retroviral RTs.Most of the other
nucleo-sideanalogs,
suchaszidovudine and2',3'-dideoxycytidine,
are
equally
effectiveagainst
HIV-1 or HIV-2 RT(2).
The results obtained in thisstudy
indicate thatalthough
thebinding
sites of the threedrugs overlap
with each other andwith those of other
knownnonnucleoside
inhibitors,
thedrugs
differ inprecisely
howthey
interact with theenzyme.Results
and discussion. We have constructedeight
single
amino acid substitutions and fourmultiple
amino acid sub-stitutions within thepolymerase
domain of HIV-1 RT(Table
1).
The amino acid substitutionscorrespond
to mutations that have beenreported
toplay
a role in resistance to nonnucleoside inhibitors(8, 10, 11, 14, 23, 26, 28, 30, 32,
35,
37). The substitutions
weregenerated
withBspMI
cassettes(3, 4), which
arederivatives of theplasmid
HIV-1 RT. Theparental plasmid
HIV-1 RT inducesexpression
of the 66-kDa form of HIV-1 RT in Escherichia coli(15, 17).
Onceaclonewas
constructed,
thepresence of themutationwasverifiedby dideoxynucleotide sequencing. Appropriate expression
of each of the variant HIV-1 RTs was determinedby
fractionation of the E. coli extracts on sodiumdodecyl
sulfate(SDS)-polyacrylamide gels.
All of the variant HIV-1expressed equivalent
Mutation % Polymerase
activity
None... 100
Leu-100-le... 40
Lys-103--Asn
... 110Lys-103-+Asn/Tyr-181-d*le
... 100Lys-103--Asn/Tyr-188--Leu
... 100Lys-103--Asn/Tyr-181- Ilefryr-188--3Leu
... 100Val-108--Ile... 100
Ile-135--Thr... 20
Val-179-+Asp
... 100Tyr-181-*Ile
... 110Tyr-181--Ile/Tyr-188--*Leu
... 120Tyr-188--*Leu
... 110His-221-
Tyr
... 130a The assays were performed as described previously (3) by with poly(rC) oligo(dG)as thetemplate-primer.Activity of the wild-type HIV-1 RTwas set at100%,and theactivitiesofthemutant RTs were normalized to thewild-typeactivity.
RT. These two mutants
have approximately 40 and
20%
the
wild-type
level of
polymerase
activity, respectively. All of
the assays were done on three separate occasions, with
independently prepared extracts. All three assays gave
sim-ilar results.
This
panel of drug-resistant mutants was constructed to
simplify
the
testing of new nonnucleoside inhibitors of
HIV-1 RT. We
have tested two nonnucleoside inhibitors,
calanolide A
(18)
and TBA
(6,
7),
and an
acyclic nucleoside
analog,
HEPT
(1,
2, 10, 25),
against
the
various
mutant
forms of HIV-1 RT. A number of
research
groups
have
shown that the
tyrosine
residues
atpositions
181 and 188
arecritically involved
in
sensitivity
tomany
nonnucleoside
drugs, including
the
pyridinones
(26,
30,
37), nevirapine (23,
28,
35),
R82150
(11, 30),
and
5-ethyl-1-ethoxymethyl-6-(phe-nylthio)uracil, which
is related
toHEPT
(11). Substitution
of
the
lysine residue
atposition
103
with
anasparagine
has been
shown
toconfer
partial resistance
topyridinone-class
drugs
and
has
alarge effect
ondrug resistance when combined with
substitutions
atposition 181 (26,
30).
The
drugs
wereinitially
tested
against variant
HIV-1 RTs
with
single amino acid
substitutions
atLys-103,
Tyr-181, and
Tyr-188 and with
avariant that has amino acid substitutions
atboth
Tyr-181
and
Tyr-188.
The
tyrosine
at181
waschanged
to anisoleucine
residue and leucine
wassubstituted for the
tyrosine
atposition 188,
since these amino
acids
arefound
atthe
equivalent
positions
in HIV-2 RT. The
lysine
atposition
103
waschanged
to anasparagine
residue,
which is the
samesubstitution
that
wasreported
by Nunberg
etal.
(26).
As
shown
in
Fig. 2A,
mutantforms of HIV-1 RT
with each of
these amino acid substitutions
wereall
resistant
orpartially
resistant
toTBA.
A mutantHIV-1 RT
with the amino acid
substitution
Tyr-181--Ile
wascompletely
resistant
tothe
drug
up
to aconcentration of 50
,uM,
indicating
that
Tyr-181
is
important
in
the
sensitivity
of HIV-1 RT
to TBA.Inter-estingly,
the HIV-1 RT
with the double amino acid
substitu-tion
Tyr-181--Ile/Tyr-188-->Leu
wasslightly
less resistant
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.83.288.69.395.2]I Tyr 181 Ile
I Tyr181Ile/Tyr188 Leu
IT eTyr188 Leu
I Lys 103 Asn
HIV- RT
uM 1H,3H-Thiazolo[3.4-a]Benzlmidazole(41)
Tyr181Ile/Tyr188Leu Tyr 188 Leu
Lys 103 Asn Tyr 181 Ile
HIV-1 ET
uMCalanolide-A
A
80%
._ ci
40%
20%
B
C.) U
on November 9, 2019 by guest
http://jvi.asm.org/
Tyr188Leu
Lys 103 Asn
Tyr181 Ile
--:=O H1V-IRT
1.0 2.0 3.0 4.0 5.0 uMHEPT
FIG. 2. Drug resistance of variant HIV-1 RTs with thesingle amino acid substitutions Lys-103--Asn,Tyr-181---Ile,andTyr-188--Leuand the double amino acid substitution Tyr-181-dle/Tyr-188-*Leu. Assays were performed as previously described (3) by using
poly(rC) oligo(dG)asthetemplate-primer. The variant HIV-1RTswereassayedwith the indicatedamountsof TBA (A),calanolideA(B), andHEPT(C). Activity of each RT in the absence ofaninhibitorwasconsideredto represent 100% activity. Activity of each RT in the
presenceofaninhibitorwasnormalizedtothis value.
resistant to
calanolide A, the level of
resistance
wasmuch
lowerthan that
seenwith
TBA (Fig. 2A). Mutants that
contained the
amino acid substitutions
Tyr-188--
Leuand
Tyr-181--*Ile/Tyr-188---Leu
showedsubstantial resistance
tocalanolide
A, suggesting
thatTyr-188
isveryimportant
for
calanolide Asensitivity in HIV-1 RT. Theobservation that
Tyr-181
andLys-103
are lessimportant for calanolide
Asensitivity
thanfor TBAsensitivity
suggests thatwhile thebinding
sites for thetwodrugs partially overlap they
arenot identical. The HIV-1 RT mutants showed a pattern of resistance to HEPT(Fig. 2C)
which was similar to that shownforcalanolide A(Fig. 2B).
Thissuggeststhat thetwodrugs,
whilestructurally
verydissimilar,
maybindatsimilar locationswithin thehydrophobic pocket.
Amutant HIV-1 RT with two amino acid
substitutions,
Tyr-181-IleFlyr-188-
Leu,
hadahigh
level of resistancetoTBA,
calanolideA,
and HEPT.Therefore,
we constructed theotherpossible
combinations of the amino acid substitu-tions andtested them for resistance to the threedrugs.
As shown inFig. 3A,
all of themutantHIV-1RTs withmultiple
mutationswerehighly
resistant to TBAcompared
with theresults of the in vitro
enzyme assaysmirror the
previously
reported in vitro
antiviral data; the
pyridinone-resistant virus
A17(26), which contains
analteration
atTyr-181 (to
acysteine)
andan alterationatLys-103 (to
anasparagine), is
resistant
to TBA(5) but is sensitive
tocalanolide
A(18).
HEPT did not show the same pattern of resistance as calanolide A in theanalysis
with themultiple-mutation RT
variants
(Fig. 3C).
Forexample,
whilethe
double
mutantLys-103-
Asn/Tyr-181--+Ile
wasonly partially
resistant to calanolideA,
itwas almostcompletely
resistanttoHEPT.Other amino acid substitutions have been
reported
to be presentinHIV-1 RTvariants which areresistantto nonnu-cleosidedrugs.
HIV-1 RT variantscontaining
these alter-ations,Leu-100-Ile (31, 37), Val-108---Ile (31),
Ile-135--Thr(31), Val-179--Asp (36),
andHis-221-eTyr
(31),
were con-structed with theBspMI
cassettemutagenesis
system.These HIV-1 RT variants were likewise tested for resistance toTBA,
calanolideA,
and HEPT. As showninFig.
4A andB,
only the
Leu-100-dIle
mutation gave anysignificant
resis-tanceto either TBAorcalanolide
A,
with theresistance tocalanolide A
being
the most marked. TheLeu-100--Ile
mutant,
however,
did besubstantially
80%.
60%7
._j 0
40%
20%
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.142.476.82.379.2]Lys 103 Asn/Tyr181 IleTyr 188Leu Lys 103 Aan/Tyr 181 le
Lys103AsnTyr188Leu
I
H1V-1RT
uM1H.3H-Thiazolo[3.4-a]BenzinUdazole
Ly 103AxnTyr181Ile/Tyr 188 Leu Lys103Asn/Tyr188Leu
Lys103AsnTyr 181Ile
) HIV-IRT
uM Calanolide-A
A
80%
ci
-C
209Y
B
-._
ci
--C
on November 9, 2019 by guest
http://jvi.asm.org/
,Lys 103Asn/Tyr 181 lIe/Tyr188 Leu
Lys103Asn/Tyr181Ile
80%
60%
40%
20%
H]V-I RT
1.0 2.0 3.0 4.0 6.0
uM HEPT
FIG. 3. Drug resistance of variant HIV-1 RTs with the multiple amino acid substitutionsLys-103-+Asn/Tyr-181-Ile,Lys-103- Asn/Tyr-188-*Leu,and
Lys-103-*Asn/Tyr-181--dle/Tyr-188--Leu.
The variant RTs were assayed with the indicated amounts ofTBA (A), calanolide A(B),andHEPT
(C).not,
by themselves, appear to be important in determining
the
sensitivity of HIV-1 RT to TBA, HEPT, or calanolide A,
it is still
possible that these substitutions could have an effect
if
they were tested against other nonnucleoside drugs or if
they
were to occur aspart
of a cluster of amino acid
substitutions.
In
the
analysis with the point mutants, TBA appears to
behave like many
previously
described nonnucleoside drugs.
Calanolide
A,
however, is
anunusual
compound and
ap-pears
tobelong
to apharmacologic
class different from that
of the other known nonnucleoside
RT-inhibitory
drugs.
Calanolide
Ais
fully
active
against
HIV-1
strain A17
(26),
and
analysis
of
the resistance and
sensitivity of
aseries of
enzymatically
active HIV-1/HIV-2
RTchimeras
suggested
that
calanolide
Adoes
notinteract with the
samesegments
of HIV-1
RT asthe TIBO
compounds
(15).
While the
pattern
of calanolide
Aresistance and
sensitivity
of the
point
mu-tantsagrees
with the A17
data,
the pattern of calanolide A
sensitivity
of the
HIV-1/HIV-2
chimeras is
moredifficult
toexplain (16).
With
the
chimeras,
sensitivity
tocalanolide
A
appeared
tobe
primarily dependent
on tworegions
from
HIV-1
RT, amino acids 93
to157 and amino acids 224
to427.
The
region
from 93
to157
contains amino
acids 100 and
103,
data derived with the
point
difficult
toimagine how alteration of this region could affect
the
drug
binding pocket either directly or indirectly. What is
more
difficult
to
reconcile with the present results are the
data with chimera
Cl,
in
which the body of the
chimera is
from HIV-1 RT
and the region between amino acids 158 and
190
derives from HIV-2 RT.
On the basis of the results with
the
drug-resistant point mutants, we would have expected
that
this
chimera, which is resistant
toTIBO, should also be
resistant
tocalanolide A. It
is,
in
fact,
only slightly less
sensitive
(50% inhibitory
concentration about threefold
higher)
tocalanolide
Athan the
wild-type
enzyme.
The
Cl
chimera is much
moresensitive
tocalanolide
Athan
is the
HIV-1 RT
with the
twomutations
Tyr-181--Ile
and
Tyr-188-+Leu.
This difference in
sensitivity
is
not aresult of the
particular
amino acids that
arepresent
atthese
positions
since the
Cl
chimera and the
HIV-1RT
with the
twomutations both contain
anIle
atposition
181
and
aLeu
atposition
188.
Therefore,
weareleft
with the
hypothesis
that
sensitivity
and
resistance
tothese
drugs
arethe
result of the
precise
geometry
of
the
drug
binding
and that this
geometry may be
quite
sensitive
tochanges
that do
notnecessarily
manifest themselves
asparticular
sites
atwhich
individual amino acid substitutions lead
directly
todrug
resistance. Amino acid
positions
within RT which have
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.123.525.82.382.2]A
10070-i
80%
Leu 100 lie
60%
Val 108 lie
>-, His221Tyr
Ile 135 Thr HIV-I RT Val 179Asp 401%
20%
10 20 30 40 50
u) IH.3H-Thiazolo[3.4-a]Benzimidazoie(4i)
B
109.
-LoU100Ile
60%
40%.
le135 Thr His 221Tyr Val108lie
Vl179Asp
HIV-1IRT
10 20 50
uM Calanolide-A
on November 9, 2019 by guest
http://jvi.asm.org/
Val179 Asp Ile 135Thr
Leu 100lle Val 108lle His221Tyr HIV- IRT
1.0 2.0 3.0 4.0 5.0
uM HEPT
FIG. 4. Drugresistance ofvariant RTs with the single aminoacidsubstitutionsLeu-100-)Ile, Val-108-dIle,Ile-135- Thr, Val-179--Asp, andHis-221-+Tyr. The variant RTswereassayedwiththe indicatedamountsof TBA (A), calanolideA(B), and HEPT (C).
idea
is further
underscored by the
recentobservation
thatanHIV-2 RT
with the amino
acid substitutions Ile-181-
Tyr andLeu-188--*Tyr
is notsensitive
tothe
nonnucleoside
inhibitors
nevirapine and
R82913 (TIBO) (14, 35). There is
hope, however,
thatadditional work
onthe
three-dimen-sional structure ofthe RT-drug
complexes and of
drug-resistant mutantswill
help
toelucidate the
exactrelation-ships
between the sequenceof HIV-1 RTand
its
structure. WearegratefultoourcolleagueAmnonHizi forhelpful discus-sions,MarilynPowers for theoligonucleotides,andHilda Marusio-disforpreparing the manuscript.This researchwassponsored by the National CancerInstitute, DHHS, undercontract no.NO1-CO-74101 with ABL.
REFERENCES
1. Baba, M.,E.DeClercq,H.Tanaka,M.Ubsawa,H.Takashima, K.Seiya,I.Nitta,K.Umezu,H.Nakashima,S.Mori,S.Shigeta,
R. T. Walker, and T. Miyasaka. 1991. Potent and selective inhibitionof humanimmunodeficiencyvirus type1 (HIV-1) by 5-ethyl-6-phenylthiouracil derivativesthrough their interaction with theHIV-1 reverse transcriptase. Proc. Natl. Acad. Sci. USA88:2356-2360.
2. Baba, M.,H.Tanaka,E.DeClercq,R.Pauwels, J. Balzarini,D. Schols, H. Nakashima, C.-F. Perno, R. T. Walker, and T. Miyasaka. 1989. Highlyspecificinhibition of human
immuno-type1reversetranscriptase.J. Virol. 66:7533-7537.
5. Buckheit, R. W., M. G. Hollingshead, J. Germany-Docker,E. L. White, J. B. McMahon, L. B. Allen, L. Westbrook, W. M. Shannon,0.Weislow, J. P. Bader, and M. R. Boyd. Submitted
forpublication.
6. Chimirri, A., S. Grasso, A.-M. Monforte, P. Monforte, and M. Zappala. 1991. Anti-HIV agents. I. Synthesis and in vitro
anti-HIVevaluation ofnovel 1H,3H-thiazolo[3,4-a]benzimida-zoles. Farmaco46:817-823.
7. Chimirri, A.,S.Grasso,A.-M.Monforte,P.Monforte, andM. Zappala. 1991. Anti-HIV agents. II. Synthesis and in vitro anti-HIV activity of novel 1H,3H-thiazolo[3,4-a]benzimida-zoles. Farmaco 46:925-933.
8. Condra, J. H., E. A. Emini, L. Gotlib, D.J. Graham, A. J. Schlabach, J.A.
Wolfpng,
R.J. Colonno,and V. V. Sardana. 1992. Identification of the human immunodeficiency virusreverse transcriptase residues that contribute to the activity of diverse nonnucleoside inhibitors. Antimicrob. Agents Chemother. 36:1441-1446.
9. Debyser, Z., R. Pauwels, K.Andries, J. Desmyter, M. Kukla, P. A.J. Janssen,and E.DeClercq. 1991. An antiviral targeton reverse transcriptase of human immunodeficiency type-1
re-vealedby
tetrahydroimidazo[4,5,1-jk][1,4]benzodiazepin-2(1H)-one and -thione derivatives. Proc. Natl. Acad. Sci. USA 88: 1451-1455.
10. DeClercq, E. 1992. HIV inhibitors targeted at the reverse
transcriptase.AIDS Res. Hum. Retroviruses 8:119-134.
807.
ci
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.158.477.68.374.2]virustype 1 resistantto 3'-azido-3'-deoxythymidine and 2',3'-dideoxyinosine. J. Virol. 66:12-19.
13. Goldman, M. E., J. H. Nunberg, J.A.O'Brien, J.C.Quintero, W. A.Schleif, K. F. Freund, S.L.Gaul,W. S.Saari, J.S.Wai, J. M. Hoffman,P. S. Anderson,D.J. Hupe,E.A. Emini,and A. M. Stem. 1991. Pyridinone derivatives: specific human immunodeficiency virustype1reversetranscriptase inhibitors with antiviral activity. Proc. Natl. Acad. Sci. USA 88:6863-6867.
14. Grob, P. M., J. C. Wu, K. A. Cohen,R. H.Ingraham, C.-K. Shih,K. D.Hargrave,T. L.McTague,and V.J.Merluzzi. 1992. Nonnucleoside inhibitors ofHIV-1reversetranscriptase: nevi-rapine as a prototype drug. AIDS Res. Hum. Retroviruses 8:145-152.
15. Hizi, A., C. McGill, and S. H. Hughes. 1988. Expression of soluble, enzymatically active, human immunodeficiency virus
reverse transcriptase in Eschenchia coli and analysis of
mu-tants.Proc.Natl. Acad. Sci. USA85:1218-1222.
16. Hizi, A., R. Tal,M. Shaharabany, M. J. Currens, M. R. Boyd, S. H.Hughes,andJ.B.McMahon. Submitted forpublication. 17. Hughes, S. H.,A. Ferris, and A. Hizi. 1990. Analysis of the
reverse transcriptase of human immunodeficiency virus
ex-pressed inEschenichia coli, p. 297-307. In W. G. Laver and G. M. Air(ed.), UseofX-raycrystallographyinthedesignof antiviralagents.AcademicPress, Inc.,NewYork.
18. Kashman, Y., K. R. Gustafson, R. W. Fuller, J. H. CardellinaII,
J. B. McMahon, M. J. Currens, R. W. Buckheit, Jr., S. H. Hughes, G. M. Cragg, and M. R. Boyd. 1992. The calanolides,a
novel HIV-inhibitory class of coumarin derivatives from the tropical rainforesttree,Calophyllum lanigerum. J. Med. Chem. 35:2735-2743.
19. Kohlstaedt, L. A., J. Wang, J. M. Friedman, P. A. Rice,and T.A. Steitz.1992. Crystalstructure at3.5 Aresolution of HIV-1
reverse transcriptase complexed with an inhibitor. Science
256:1783-1790.
20. Koup, R. A., V. J. Merluzzi, K. D. Hargrave, J. Adams, K. Grozinger, R. J. Eckner, and J. L. Sullivan. 1991. Inhibition of humanimmunodeficiency virus type 1(HIV-1) replication by thedipyridodiazepinone BI-RG-587. J. Infect. Dis. 163:966-970. 21. Larder, B. A., K. E. Coates, and S. D. Kemp. 1991. Zidovudine-resistant humanimmunodeficiency virus selected bypassagein cell culture.J. Virol.65:5232-5236.
22. Larder, B. A., and S. D. Kemp. 1989. Multiple mutations in HIV-1 reverse transcriptase confer high-level resistance to
zidovudine(AZT). Science 246:1155-1158.
23. Mellors, J. W., G. E. Dutschman, G.-J. Im, E. Tramontano, S.R.Winkler, and Y.-C. Cheng. 1992. In vitro selection and molecularcharacterization of human immunodeficiency virus-1 resistant to nonnucleoside inhibitors ofreverse transcriptase.
Mol.Pharmacol.41:446-451.
24. Merluzzi,V. J., K. D. Hargrave, M. Labadia, K. Grozinger, M. Skoog, J.C.Wu, C.-K. Shih, K. Eckner, S. Hallox,J. Adams, A. S.Rosenthal, R. Faanes,R. J. Eckner, R. A. Koup, and J. L. Sullivan. 1990. Inhibition ofHIV-1 replication bya
nonnucleo-sidereversetranscriptase inhibitor. Science 250:1411-1413. 25. Miyasaka, T., H. Tanaka, M. Baba, H. Hayakawa, R. T.
Walker, J. Balzarini, and E. DeClercq. 1989.A novel leadfor
specific anti-HIV-1 agents: 1-[(2-hydroxyethoxy)methyl]-6-(phenylthio)thymine. J. Med. Chem. 32:2507-2509.
26. Nunberg, J. H., W. A.Schleif, E. J. Boots, J. A. O'Brien, J. C. Quintero, J. M. Hoffman, E. A. Emini, and M. E. Goldman. 1991. Viral resistance to human immunodeficiency virus type 1-specific pyridinone reverse transcriptase inhibitors. J. Virol. 65:4887-4892.
27. Pauwels, R., K.Andries, J. Desmyter, D. Schols, M. J. Kukla, H. J. Bresline, A. Racymaeckers, J. van Gelder, R. Woeslen-borghs, J. Heykants, K. Schellekens, M. A. C. Janssen, E. DeClercq, and P. A. J. Janssen. 1990. Potent and selective inhibitionofHIV-1replicationinvitrobyanovel series of TIBO derivatives. Nature (London) 343:470-473.
28. Richman, D., C.-K. Shih,I.Lowy, J. Rose, P. Prodanovich, S. Goff, and J. Griffin. 1991. Human immunodeficiency virus type 1 mutants resistanttononnucleoside inhibitors ofreverse tran-scriptase arise in tissue culture. Proc. Natl. Acad. Sci. USA 88:11241-11245.
29. Romero, D. L., M.Busso,C.-K. Tan, F. Reusser,J. R. Palmer, S. M.Poppe,P. A.Aristoff,K. M.Downey,A. G.So,L.Resnick, and W. G.Tarpley. 1991.Nonnucleosidereversetranscriptase inhibitors thatspecifically block human immunodeficiencyvirus type 1replication.Proc. Natl.Acad. Sci. USA 88:8806-8810. 30. Sardana, V. V., E. A.Emini,L. Gotlib,D.J.Graham,D. W.
Lineberger, W. J.Long,A.J.Schiabach,J. A.Wolfgang, and J. H. Condra. 1992. Functional analysis of HIV-1 reverse
transcriptase amino acids involved in resistance to multiple non-nucleoside inhibitors.J. Biol. Chem. 267:17526-17530. 31. Schleif, W. A., E. A.Emini, A. Rhodes, D. L. Titus, L.Gotlib,
J. H. Condra, and V. W. Byrnes. 1992. Development and analysis of human immunodeficiency virus type 1 resistant to HIV-1 specific pyridinone reverse transcriptase inhibitors. J. Cell.Biochem.Suppl. 16E(Q552):87.
32. Shaharabany, M., and A. Hizi. 1991.The catalytic functions of chimeric reverse transcriptases of human immunodeficiency virusestype 1andtype2.J.Biol. Chem. 267:1-5.
33. Shih, C.-K.,J. M. Rose, G. L. Hansen, J.C.Wu, A.Bacolla,and J. A. Griffin. 1991. Chimeric human immunodeficiency virus type1/type2reversetranscriptases displayreversedsensitivity tononnucleoside analog inhibitors.Proc.Natl.Acad. Sci. USA 88:9878-9882.
34. St.Clair,M.H., J. L. Martin, G. Tudor-Williams, M. C. Bach, C. L.Vavro, D. M. King, P. Kellam, S. D. Kemp, and B. A. Larder. 1991. Resistance toddland sensitivity to AZT induced by a mutation in HIV-1 reverse transcriptase. Science 253:1557-1559.
35. Tan, G. T., A. D. Kinghorn, S. H. Hughes, and J. M. Pezzuto. 1991.Psychotrine andits0-methylether are selective inhibitors of human immunodeficiency virus-1 reverse transcriptase. J. Biol.Chem. 266:23529-23536.
36. Vandamme, A.-M. (Rega Institute, Leuven, Belgium). Personal communication.
37. Vasudevachari, M. B., C. Battista, H. C. Lane, M. C. Psallido-poulos, B. Zhao, J.Cook, J. R. Palmer, D. L. Romero, W. G. Tarpley, and N. P. Salzman. 1992. Prevention of the spread of HIV-1 infectionwith non-nucleoside reverse transcriptase in-hibitors.Virology 180:269-277.