0022-538X/96/$04.0010
Copyrightq1996, American Society for Microbiology
Cloning, Expression, and Characterization of the Proteinase
from Human Herpesvirus 6
NATALIE J. TIGUE,1PHILIP J. MATHARU,1NOEL A. ROBERTS,2JOHN S. MILLS,2
JOHN KAY,1
ANDRAY JUPP2*
Department of Molecular and Medical Biosciences, University of Wales College of Cardiff, Cardiff CF1 3US,1and
Department of Biology, Roche Research Centre, Welwyn Garden City AL7 3AY,2United Kingdom
Received 4 December 1995/Accepted 5 March 1996
After the U53 gene encoding the proteinase from human herpesvirus 6 (HHV-6) was sequenced, it was expressed inEscherichia coli, and the activity of the purified, recombinant HHV-6 proteinase was characterized quantitatively by using synthetic peptide substrates mimicking the release and maturation cleavage sites in the polyprotein precursors of HHV-6, human cytomegalovirus (CMV), murine CMV, and Epstein-Barr virus. Despite sharing 40% identity with other betaherpesvirus proteinases such as human CMV proteinase, the one-chain HHV-6 enzyme was distinguished from these two-chain proteinases by the absence of an internal autocatalytic cleavage site.
Primary acute infection with human herpesvirus 6 (HHV-6) frequently occurs early in childhood and appears to result in the mild disease exanthema subitum (15). At this time, anti-body titers are readily detectable but normally decline with age (15) and indicate a change to a low but persistent level of replication. HHV-6 was first isolated from peripheral blood mononuclear cells of patients with lymphoproliferative disor-ders including AIDS (24). The common CD41T-cell tropism of HHV-6 and human immunodeficiency virus (HIV) has led to the speculation that HHV-6 may influence the progression into AIDS of HIV-infected patients (17). In addition, HHV-6 has also been associated with myalgic encephalomyelitis and chronic fatigue syndrome (1, 29). Whether HHV-6 has a direct or indirect effect on the progression of these diseases remains to be proven. However, since HHV-6 has a high incidence among the normal population, this virus could have a serious impact on the outcome and clinical treatment for any disease in which it plays a role.
Assembly of the capsids of herpesviruses requires the for-mation of a scaffold to enable the capsid proteins to coalesce. One of the proteins present in this scaffold is the virally en-coded proteinase which is produced initially in the form of a precursor (Fig. 1a). Autolytic cleavages at junctions referred to as the release and maturation sites generate the N-terminal proteinase and the mature form of the viral assembly protein domain, respectively (Fig. 1a). The critical role of proteolytic processing in maturation of the viral particles (22, 28) was demonstrated when a mutant virus containing an inactivated proteinase gene was shown to be capable of forming only aberrant capsids (4, 8, 23). Consequently, the proteinases from herpesviruses have considerable potential as new antiviral tar-gets, especially since current therapies (e.g., ganciclovir and acyclovir) are not without their limitations (18). Whereas con-siderable attention has been focused on the proteinases from human cytomegalovirus (HCMV) and herpes simplex virus type 1 (HSV-1) (2, 5, 14, 20, 25, 31, 32) and on the role of these enzymes in viral particle assembly (4, 8, 22, 28), proteinases from other herpesviruses such as Epstein-Barr virus (EBV) are less well understood (7). In this report, we present a sequence
for HHV-6 proteinase derived independently from that in-cluded in the recent report of the entire HHV-6 genome (9) and describe the production and characterization of recombi-nant HHV-6 enzyme.
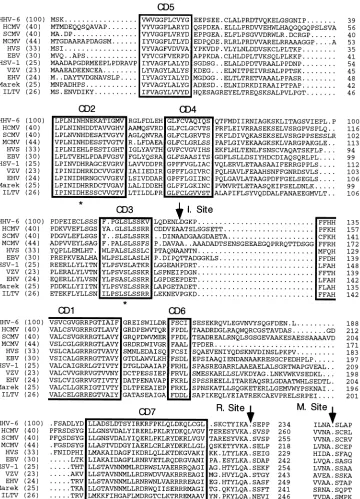
The 14W2 lambda clone that contains a fragment from the U1102 strain of HHV-6 was kindly provided by M. D. Jones (Royal Postgraduate Medical School, London, United King-dom). This clone was used as template DNA in a PCR (Fig. 1b) with forward (F1 [59-GCCACCTCCAATAACGCT-39]) and reverse (R1 [59-TCCCATAGATCTTCCCCG-39]) oligo-nucleotide primers that were designed on the sequences at the 39end of the assembly protein gene of HHV-6 (the L1 open reading frame in the SAD6 restriction fragment) (13) and the 39 end of the CB3 (12, 19)-specific restriction fragment (Eu-ropean Collection of Animal Cell Cultures, Porton Down, Wiltshire, United Kingdom). The 4.2-kb fragment thus ob-tained was digested withEcoRI and XhoI (Fig. 1c) and sub-cloned into the vector pSP72 (Promega, Madison, Wis.) and subjected to dideoxynucleotide sequencing. Subsequently, 1.7-kbp fragments were generated in two independent PCRs of the 14W2 clone utilizing new forward (59-ATCAGACGTCGACA GCATTGATGTA-39) and reverse (59-GCGCGTCGACTTA ACTATCCATTTTATTTAACGCGTCA-39) primers (Fig. 1c) and directly cloned into the pGemT vector (Promega). Two clones, one from each of the independent PCRs, were se-quenced in their entirety. The nucleotide sequence (EMBL database; accession no. X87419) was in precise agreement with that included subsequently in the report describing the com-plete organization of the HHV-6 genome (9). The sequence predicts a total of 528 amino acids for the proteinase and its associated assembly protein domain. The predicted amino acid sequence of the proteinase is aligned (Fig. 2) with those from each of the three herpesvirus subtypes (alpha, beta, and gam-ma). Since the homology among the assembly proteins is poor, these domains have not been included in Fig. 2 except for the sequences around the maturation sites. Within the proteinase itself, there are a number of conserved domains (CDs) which are boxed (Fig. 2) and labelled 1 to 7 in accordance with previous suggestions (16, 30). An active-site serine residue has previously been identified for the herpesvirus proteinases (6, 27, 30), and this invariant serine residue is present in HHV-6 proteinase in the same motif CD3 as in the other herpesviruses (Fig. 2). In addition, Fig. 2 shows the percent conservation of
* Corresponding author. Phone: 366946. Fax: 44-1707-373504.
4136
on November 9, 2019 by guest
http://jvi.asm.org/
identical amino acids between HHV-6 and each of the other herpesvirus proteinases. These data demonstrate that HHV-6 proteinase is most homologous to the CMV (betaherpesvirus) proteinases sharing about 40% identity, followed by the gam-ma- and alphaherpesvirus proteinases with which it shares 30 and 25% identity, respectively.
The HHV-6 release site that connects the proteinase to the assembly protein domain also shows a high degree of conser-vation with its counterparts from other herpesvirus protein-ases. In particular, it contains the tyrosine, alanine, and serine residues at P4, P1, and P19, respectively that are invariant in all the herpesviruses (except for an asparagine in P19 of ILTV; Fig. 2). In addition, HHV-6 proteinase like the other beta- and gamma- but not alphaherpesvirus proteinases possesses a lyzsine residue in P2. The proline residue in P49is also
con-served among the betaherpesvirus group and EBV proteinase. Similarly, the maturation site at the C terminus of the HHV-6 assembly protein shares a high degree of homology with the other betaherpesvirus proteinases with the notable exception of the P29 residue which is leucine in HHV-6 proteinase but cysteine in all of the others.
The 14W2 clone was used as a template with the oligonu-cleotide 59-GCTGCGGCTAGCAAAGTTTGGGTAGGTGG ATTC-39together with the oligonucleotides 59-GCGCGTCG ACTTAACTATCCATTTTATTTAACGCGTCA-39or 59-GC CCTCGTCGACTCAAGCTTTAATATACGTACATTTAG-39 in separate PCRs to amplify the full-length HHV-6 polyprotein and the N-terminal proteinase (from amino acid residues 2 to 230), respectively. In a parallel set of reactions, the DNA fragment encoding the HCMV proteinase domain from
nucle-FIG. 1. Schematic representation of the HHV-6 genome. (a) The HHV-6 proteinase and assembly protein are shown, together with the positions of the release and maturation sites. (b) The 162-kbp HHV-6 genome (U1102) is composed of a unique region (UL) flanked by two direct terminal repeats (TR1 and TR2). (c) An
expanded view of the region contained in the 14W2 lambda clone. The positions of the PCR primers within the CB3 and SAD6 restriction fragments are indicated. The originalEcoRI-XhoI subclone constructed for sequencing is also shown, together with the position of the U53 proteinase gene. The restriction sitesEcoRI (R1), XhoI (X1), andBglII (B2) are also indicated.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.143.471.77.510.2]otide positions 115207 to 115965 (3) was generated by PCR from the AD169 strain of HCMV by using the upstream (59 -CGCCGCTAGCGACGAGCAGCAGTCGCAGGCTGT-39) and downstream (5GGATCCTTACGCCTTGACGTATGA CTCGCGCT-39) oligonucleotide primers. These fragments were inserted (separately) into the pDS56, 6His expression vector (7) by using the restriction endonuclease sites (indicated by underlining in the oligonucleotide sequence) and verified by
dideoxynucleotide sequencing. Following transformation into
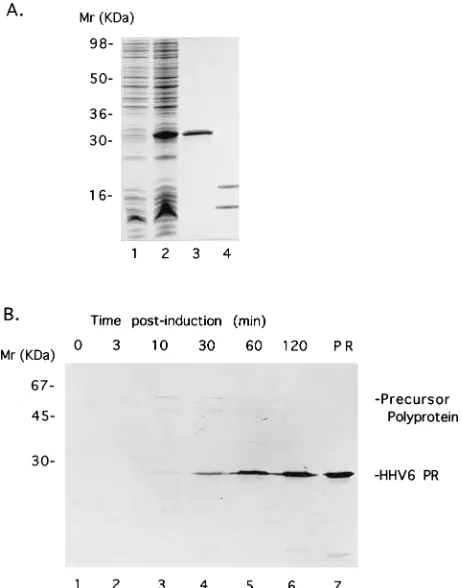
Escherichia coliM15(pDM1.1) (7), the cells were induced with 200mg of isopropyl-b-D-thiogalactopyranoside (IPTG) per ml for 120 min (Fig. 3A, lanes 1 and 2). For protein purification, cells were lysed in buffer containing 50 mM sodium phosphate (pH 7.8), 1 mM 4-(2-aminoethyl)-benzenesulfonyl fluoride hy-drochloride (ABSF), 1% Tween 20, 1 M NaCl, and 1 mg of lysozyme per ml for 20 min. Following sonication and
centrif-FIG. 2. Multiple alignment of herpesvirus proteinases and assembly protein maturation sites. The nucleotide sequence for each viral proteinase (HCMV, simian CMV [SCMV], [MCMV], herpesvirus saimari [HVS], EBV, herpes simplex virus type 1 [HSV-1], varicella-zoster virus [VZV], equine herpes virus [EHV], Marek’s disease virus [Marek], and infectious laryngotracheitis virus [ILTV]) was retrieved from GenBank and used to deduce the amino acid sequences shown. The percent identity of the HHV-6 proteinase with each of the others in this alignment is shown in parentheses at the left-hand side of each sequence. The numbering of the amino acid residues in each proteinase sequence is given on the right-hand side of each line. The conserved domains (CD) 1 to 7 are boxed, and the positions of the release (R.), maturation (M.), and internal (I.) cleavage sites are indicated.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.129.489.69.568.2]ugation at 18,000 rpm in a Sorvall SS34 rotor, the supernatant was subjected to Ni21chelate chromatography (Qiagen,
Chats-worth, Calif.) over a column equilibrated in buffer containing 50 mM sodium phosphate (pH 7.8), 500 mM NaCl, and 10% glycerol. After the column was washed with this buffer, it was washed in a similar buffer at pH 6.0. A final wash was per-formed with the latter buffer containing 75 mM imidazole prior to elution in the same buffer containing 500 mM imidazole. Fractions containing proteinase were pooled and dialyzed against a solution containing 50 mM Tris-HCl buffer (pH 8.0), 100 mM NaCl, 1 mM dithiothreitol, 1 mM EDTA, and 2% glycerol. Purified protein was analyzed by sodium dodecyl sul-fate–16% polyacrylamide gel electrophoresis (SDS-PAGE) and Coomassie blue staining, and the concentration of each purified preparation was determined with the Micro BCA Pro-tein Assay Kit (Pierce, Rockford, Ill.). The purified HHV-6 proteinase migrated as a single band of protein (Fig. 3A, lane 3) with a molecular size consistent with that predicted by the gene sequence (Fig. 2). In contrast, HCMV proteinase ex-pressed and purified under identical conditions, was found to consist of two peptide chains (Fig. 3A, lane 4). Previous reports (2, 11, 25) have indicated that HCMV proteinase consists of a
mixture of one- and two-chain polypeptides after purification and the proportion of the two-chain enzyme increased upon storage in one study (25). Proteinases from betaherpesviruses such as HCMV may be distinguished from their counterparts from the other subfamilies by having an internal, autocatalytic (-AlapAla-) cleavage site, in addition to the release and mat-uration cleavage sites. Thus, the expression system employed in the present study may have facilitated more-efficient con-version from the one- to the two-chain form of the proteinase prior to extraction and purification. At present, it is unclear what purpose, if any, this cleavage serves, since identical prop-erties were observed for a mutant form of HCMV proteinase which was unable to process itself at this site compared with the wild-type enzyme (11, 21). However, this complication of internal autoprocessing within HCMV proteinase was not ob-served with the recombinant HHV-6 proteinase (Fig. 3A, lane 3). Inspection of the amino acid sequence of HHV-6 protein-ase in the region between CD3 and CD1 (Fig. 2) reveals that the internal (-AlapAla-) cleavage junction of the other beta-herpesvirus proteinases is not present in the HHV-6 enzyme. On this basis, an explanation for the one-chain appearance of HHV-6 proteinase is readily forthcoming.
A polyclonal antiserum was raised in rabbits against the purified HHV-6 proteinase and used in Western blots (immu-noblots) (Fig. 3B) to monitor the time course of processing of the full-length HHV-6 precursor polyprotein. Only a faint band migrating (at;60 kDa) in the position anticipated for the precursor was observed at early time points (Fig. 3B, lanes 3 and 4). The predominant band in later time points (Fig. 3B, lanes 5 and 6), however, comigrated with authentic HHV-6 proteinase (lane 7) as obtained after nickel chelate chroma-tography (see above), indicating that the polyprotein precursor had undergone posttranslational processing. Since this product was stable to further processing and only a single predominant band was detected, it would appear that processing of the polyprotein precursor ultimately results in the generation of the mature HHV-6 proteinase. Furthermore, the relative ab-sence of the full-length precursor and the product following cleavage at the M site compared with mature HHV-6 protein-ase indicates that hydrolysis of both the M and R sites in
Escherichia coliis rapid.
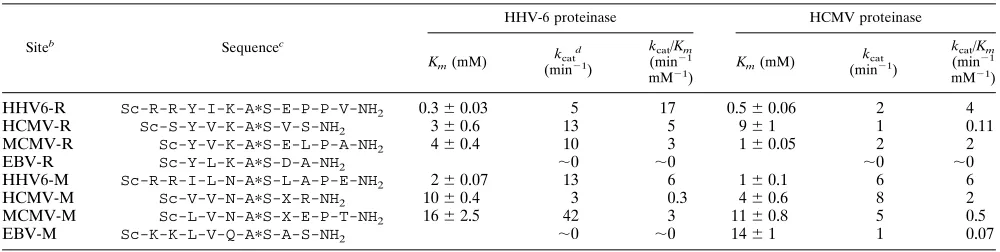
The purified HHV-6 proteinase was also utilized in experi-ments to assess its ability to hydrolyze synthetic peptides cor-responding to the release and maturation cleavage sites in its own and related protein sequences (Table 1). The peptide substrates were synthesized on a Milligen 9050 Pepsynthesiser and were purified to .95% by reverse-phase high-pressure liquid chromatography. The cleavage assay was performed in a solution containing 50 mM Tris-HCl buffer (pH 8.5), 15 mM NaCl, 1 mM EDTA, and 10% glycerol in a total volume of 50
ml at 218C. The reactions were stopped with 200ml of 0.05 M triethylammonium phosphate (TEAP) (pH 2.5)–10% acetoni-trile and analyzed by reverse-phase HPLC on a Hypersil C18 column. TheA215of the eluate was monitored. TheKmvalues
were derived by using the program Enzfitter (Biosoft, Cam-bridge, United Kingdom) and thekcatvalues were determined
by using the equation kcat 5 Vmax/[E], where E is enzyme
concentration, assuming that all of enzyme used in the assay was active. TheKmvalue for the peptide corresponding to the
release site in HHV-6 was about sevenfold lower than that measured for the HHV-6 M-site peptide (Table 1) so that a higher value for the specificity constant (kcat/Km) was derived
[image:4.612.63.294.72.366.2]for the R-site peptide. A similar trend was observed with HHV-6 proteinase acting on the R- and M-site peptides from HCMV, and the proteinase also hydrolyzed peptides based upon the cleavage sites in murine CMV (MCMV) (Table 1). In
FIG. 3. Expression and purification of recombinant HHV-6 and HCMV pro-teinases. Samples were analyzed by SDS-PAGE followed by staining with Coo-massie blue (A) or Western blotting (B). (A) Aliquots were taken for analysis before (lane 1) and after (lane 2) induction with IPTG and from the purified HHV-6 proteinase sample recovered from the nickel chelate affinity column (lane 3). An aliquot of recombinant HCMV proteinase purified in a parallel manner is shown for comparison (lane 4). (b) Western blot ofE. coliexpressing the full-length 528-amino-acid protein precursor before (lane 1) and after in-duction for 3, 10, 30, 60, and 120 min (lanes 2 to 6, respectively). Immunostaining utilized a polyclonal antiserum raised against HHV-6 proteinase (amino acids 2 to 230). A sample of affinity-purified HHV-6 proteinase is included for compar-ison (lane 7). The positions of migration of the full-length 528-amino-acid pre-cursor polypeptide, HHV-6 proteinase (PR), and standard protein markers are indicated.
on November 9, 2019 by guest
http://jvi.asm.org/
contrast, the HHV-6 proteinase was ineffective against pep-tides based upon the cleavage sites in the EBV polyprotein precursor, despite the EBV M-site peptide containing two ba-sic residues in the P6 and P5 positions. When two baba-sic resi-dues were included in these positions in the HHV-6 R- and M-site peptides (to improve water solubility of the substrates), these two peptides proved to have the highest values ofkcat/
Km. However, the specificity constants measured for HHV-6
proteinase acting on both the HCMV R- and M-site peptides and on the MCMV M-site peptide (which lacks basic residues in these positions) were comparable to that determined for the HHV-6 M-site substrate and only three to sixfold lower than that of the HHV-6 R-site peptide. This suggests that HHV-6 proteinase may not require residues in the P6 and P5 positions of a substrate, giving some indication of the length of the active-site cleft. Whereas it has been reported (5) that HSV-1 proteinase does require a substrate to contain a P5 residue for cleavage to occur, HCMV proteinase does not appear to have such a requirement (25, 27). Consistent with this, the two peptides from HHV-6 (both of which contained P6 and P5 residues) used in the present study were cleaved by recombi-nant HCMV proteinase with efficiencies comparable to those of the HCMV M-site and MCMV R-site peptides (Table 1) which did not contain P6 and P5 amino acid residues. The EBV-derived peptide sequences were not cleaved efficiently by HCMV proteinase, just as was described above for HHV-6 proteinase (Table 1). The HHV-6 proteinase showed compa-rable if not better activity than HCMV proteinase against most of the peptide substrates tested (Table 1) with the exception of the HCMV M-site peptide.
In order to characterize the recombinant HHV-6 proteinase more extensively, a peptide substrate (Dabcyl-R-R-V-V-N-ApS-X-R-L-N-Edans) containing two internally quenching flu-orescent substituents was utilized. Kinetic parameters for its hydrolysis by both recombinant HHV-6 and HCMV protein-ases were determined fluorimetrically, as described previously (10). Briefly, 0.23mM proteinase was incubated with various concentrations of substrate in buffer containing 50 mM Tris-HCl (pH 8.5), 15 mM NaCl, 1 mM EDTA, and 10% glycerol in a total volume of 500ml at 218C for 90 min prior to the addition of 500ml of dimethyl sulfoxide to terminate the reaction. The increase in fluorescence resulting from substrate cleavage was measured by using excitation and emission wavelengths of 340 and 530 nm, respectively. The values obtained for HHV-6 proteinase (Km, 3365mM;kcat, 0.5 min21;kcat/Km, 15 min21
mM21) were comparable to those derived for the proteinase
from HCMV (Km, 961mM;kcat, 0.8 min2
1,k
cat/Km, 89 min2
1
mM21); however, a lower K
m value for HHV-6 proteinase
dictated that the specificity constant for this enzyme was ap-proximately sixfold lower than that determined for HCMV proteinase. A similar relationship in the kcat/Km values was
measured (Table 1) for the unmodified substrate (the HCMV M-site peptide) upon which the fluorescent derivative was based. However, the absolute magnitude of kcat/Km was
im-proved by around 50-fold for both proteinases over that of the unmodified peptide; this effect was achieved primarily by a substantial improvement in theKmvalues as a consequence of
the increased length and presence of the bulky Dabcyl and Edans substituents. Having thus validated the fluorescent pep-tide substrate, this assay was used as a more convenient means by which to elucidate the effects of pH and temperature on the hydrolysis of this peptide by the recombinant HHV-6 protein-ase. The pH profile was determined at 218C in buffer contain-ing 20 mM 2-N-morpholinoethanesulfonic acid, 20 mM Tris-HCl, and 15 mM NaCl, which was adjusted to the indicated pH by the addition of NaOH. At pH values below about 6.5, relatively little activity was observed. The apparent pH opti-mum measured against the fluorescent substrate at 218C was pH 8.5, and at pH 9.5, approximately 50% of this maximal activity was measured. For comparison, HCMV proteinase had a pH optimum of 8.0 with this substrate. Whereas the activity of the enzyme was affected only marginally at temperatures between 4 and 218C, a fourfold decrease in activity was ob-served between 21 and 308C, followed by a further threefold decrease between 30 and 378C. Similar temperature dependen-cies have been observed with recombinant HCMV and EBV proteinases with this substrate (data not shown).
The proteinases of herpesviruses appear to be catalytically inefficient compared with archetypal serine proteinases (2, 5, 21, 25). Furthermore, since the recombinant HHV-6 protein-ase was considerably less active at 378C than at 218C (and below), the limited proteolytic capability of the proteinases from this virus family may be constrained even further under physiological conditions. These characteristics, which have also been observed for alphaviruses (26), may be important for controlling the processing of viral structural proteins and/or be required to limit proteolysis of cellular proteins to reduce possible deleterious effects on the host cell.
N.J.T. is supported by an MRC Collaborative Studentship Award. We are very grateful to Mick Jones, Royal Postgraduate Medical School London, for kindly providing the 14W2 lambda clone. In ad-TABLE 1. Cleavage of synthetic peptide substrates by recombinant HHV-6 and HCMV proteinasesa
Siteb Sequencec
HHV-6 proteinase HCMV proteinase
Km(mM)
kcatd
(min21)
kcat/Km
(min21
mM21)
Km(mM)
kcat
(min21)
kcat/Km
(min21
mM21)
HHV6-R Sc-R-R-Y-I-K-ApS-E-P-P-V-NH2 0.360.03 5 17 0.560.06 2 4
HCMV-R Sc-S-Y-V-K-ApS-V-S-NH2 360.6 13 5 961 1 0.11
MCMV-R Sc-Y-V-K-ApS-E-L-P-A-NH2 460.4 10 3 160.05 2 2
EBV-R Sc-Y-L-K-ApS-D-A-NH2 ;0 ;0 ;0 ;0
HHV6-M Sc-R-R-I-L-N-ApS-L-A-P-E-NH2 260.07 13 6 160.1 6 6
HCMV-M Sc-V-V-N-ApS-X-R-NH2 1060.4 3 0.3 460.6 8 2
MCMV-M Sc-L-V-N-ApS-X-E-P-T-NH2 1662.5 42 3 1160.8 5 0.5
EBV-M Sc-K-K-L-V-Q-ApS-A-S-NH2 ;0 ;0 1461 1 0.07
aIncubations were performed at 218C and the optimal pH values of 8.5 and 8.0 for HHV-6 and HCMV proteinase, respectively.
bThe sequences from the release and maturation sites in each viral polyprotein (Fig. 1) are indicated by the suffix R and M, respectively.
cThe scissile peptide bond is indicated by an asterisk. X, 2-aminobutyric acid; Sc, succinyl; NH
2, C-terminal amide.
dValues fork
catwere calculated fromVmax/[E], assuming that all of the proteinase present was in an active form.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.57.557.82.208.2]dition, we thank Raj Handa for synthesizing the peptide substrate and Anne Broadhurst and Alison Ritchie for valuable contributions to the work described.
REFERENCES
1.Buchwald, D., P. R. Cheney, D. L. Peterson, B. Henry, S. B. Wormsley, A.
Geiger, D. V. Ablashi, S. Z. Salahuddin, C. Saxinger, R. Biddle, R. Kikinis,
F. A. Jolesz, and T. A. Folks.1992. A chronic illness characterized by fatigue,
neurologic and immunologic disorders, and active human herpesvirus type 6 infection. Ann. Intern. Med.116:103–113.
2.Burck, P. J., D. H. Berg, T. P. Luk, L. M. Sassmannshausen, M. Wakulchik,
D. P. Smith, H. M. Hsiung, G. W. Becker, W. Gibson, and E. C. Villarreal.
1994. Human cytomegalovirus maturational proteinase: expression in Esch-erichia coli, purification, and enzymatic characterization by using peptide substrate mimics of natural cleavage sites. J. Virol.68:2937–2946.
3.Chee, M. S., A. T. Bankier, S. Beck, R. Bohni, C. M. Brown, R. Cerny, T.
Horsnell, C. A. Hutchison, T. Kouzarides, J. A. Martignetti, E. Preddie, S. C.
Satchwell, P. Tomlinson, K. M. Weston, and B. G. Barrell.1990. Analysis of
the protein-coding content of the sequence of human cytomegalovirus strain AD169. Curr. Top. Microbiol. Immunol.154:125–169.
4.Desai, P., S. C. Watkins, and S. Person.1994. The size and symmetry of B
capsids of herpes simplex virus type 1 are determined by the gene products of the UL26 open reading frame. J. Virol.68:5365–5374.
5.DiIanni, C. L., C. Mapelli, D. A. Drier, J. Tsao, S. Natarajan, D. Riexinger,
S. M. Festin, M. Bolgar, G. Yamanaka, S. P. Weinheimer, C. A. Meyers, R. J.
Colonno, and M. G. Cordingley.1993. In vitro activity of the herpes simplex
virus type 1 protease with peptide substrates. J. Biol. Chem.268:25449– 25454.
6.DiIanni, C. L., J. T. Stevens, M. Bolgar, D. R. O’Boyle, S. P. Weinheimer,
and R. J. Colonno.1994. Identification of the serine residue at the active site
of the herpes simplex virus type 1 protease. J. Biol. Chem.269:12672–12676.
7.Donaghy, G., and R. Jupp.1995. Characterization of the Epstein-Barr virus
proteinase and comparison with the human cytomegalovirus proteinase. J. Virol.69:1265–1270.
8.Gao, M., L. Matusick-Kumar, W. Hurlburt, S. F. DiTusa, W. W. Newcomb,
J. C. Brown, P. J. McCann, I. Deckman, and R. J. Colonno.1994. The
protease of herpes simplex virus 1 is essential for functional capsid formation and viral growth. J. Virol.68:3702–3712.
9.Gompels, U. A., J. Nicholas, G. Lawrence, M. Jones, B. J. Thomson, M. E. D.
Martin, S. Efstathiou, M. Craxton, and H. A. Macaulay.1995. The DNA
sequence of human herpesvirus-6: structure, coding content and genome evolution. Virology209:29–51.
10.Handa, R., E. Keech, E. A. Conway, A. Broadhurst, and A. Ritchie.1995.
Design and synthesis of a quenched fluorogenic peptide substrate for human cytomegalovirus proteinase. Antivir. Chem. Chemother.6:255–261.
11.Holwerda, B. C., A. J. Wittwer, K. L. Duffin, C. Smith, M. V. Toth, L. S. Carr,
R. C. Wiegand, and M. L. Bryant.1994. Activity of two-chain recombinant
human cytomegalovirus protease. J. Biol. Chem.269:25911–25915.
12.Josephs, S. F., D. V. Ablashi, S. Z. Salahuddin, L. L. Jagodzinski, S. F.
Wong, and R. C. Gallo.1991. Identification of the human herpesvirus 6
glycoprotein H and putative large tegument protein genes. J. Virol.65:5597– 5604.
13.Lawrence, G. L., M. Chee, M. A. Craxton, U. A. Gompels, R. W. Honess, and
B. G. Barrell.1990. Human herpesvirus 6 is closely related to human
cyto-megalovirus. J. Virol.64:287–299.
14.Liu, F., and B. Roizman.1991. The herpes simplex virus 1 gene encoding a
protease also contains within its coding domain the gene encoding the more abundant substrate. J. Virol.65:5149–5156.
15.Lopez, C., and R. W. Honess.1990. Human herpesvirus-6, p. 2055–2062.In
B. N. Fields and D. M. Knipe, Virology. Raven Press Ltd., New York.
16. Loutsch, J. M., N. J. Galvin, M. L. Bryant, and B. C. Holwerda.1994.
Cloning and sequence analysis of murine cytomegalovirus proteinase and capsid assembly protein genes. Biochem. Biophys. Res. Commun.203:472– 478.
17. Lusso, P., A. De Maria, M. Malnati, F. Lori, S. E. De Rocco, M. Baleser, and
R. C. Gallo.1991. Induction of CD4 and susceptibility to HIV infection in
human CD81T lymphocytes by human herpesvirus 6. Nature (London)
349:533–535.
18. Markham, A., and D. Faulds.1994. Ganciclovir: an update of its therapeutic
use in cytomegalovirus infection. Drugs48:455–484.
19. Martin, M. E., B. J. Thomson, R. W. Honess, M. A. Craxton, U. A. Gompels,
M. Y. Liu, E. Littler, J. R. Arrand, I. Teo, and M. D. Jones.1991. The
genome of human herpesvirus 6: maps of unit-length and concatemeric genomes for nine restriction endonucleases. J. Gen. Virol.72:157–168.
20. McCann, P. J., D. R. O’Boyle, and I. C. Deckman.1994. Investigation of the
specificity of the herpes simplex virus type 1 protease by point mutagenesis of the autoproteolysis sites. J. Virol.68:526–529.
21. O’Boyle, D. R., K. Wager-Smith, J. T. Stevens, and S. P. Weiheimer.1995.
The effect of internal autocleavage on kinetic properties of the human cytomegalovirus protease catalytic domain. J. Biol. Chem.270:4753–4758.
22. Preston, V. G., M. Al-Kobaisi, I. M. McDougall, and F. J. Rixon.1994. The
herpes simplex virus gene UL26 proteinase in the presence of the UL26.5 gene product promotes the formation of scaffold like structures. J. Gen. Virol.75:2355–2366.
23. Preston, V. G., J. A. Coates, and F. J. Rixon.1983. Identification and
characterization of a herpes simplex virus gene product required for encap-sidation of virus DNA. J. Virol.45:1056–1064.
24. Salahuddin, S. Z., D. V. Ablashi, P. D. Markham, S. F. Josephs, S.
Stur-zenegger, M. Kaplan, G. Halligan, P. Biberfeld, F. Wong-Staal, B.
Kramar-sky, and R. C. Gallo.1986. Isolation of a new virus, HBLV, in patients with
lymphoproliferative disorders. Science234:596–601.
25. Sardana, V. V., J. A. Wolfgang, C. A. Veloski, W. J. Long, K. LeGrow, B.
Wolanski, E. A. Emini, and R. L. LaFemmina.1994. Peptide substrate
cleavage specificity of the human cytomegalovirus protease. J. Biol. Chem.
269:14337–14340.
26. Sawicki, D. L., and S. G. Sawicki.1994. Alphavirus positive and negative
strand RNA synthesis and the role of polyproteins in formation of viral replication complexes. Arch. Virol. Suppl.9:393–405.
27. Stevens, J. T., C. Mapelli, J. Tsao, M. Hail, D. O’Boyle, S. P. Weinheimer,
and C. L. DiIanni.1994. In vitro proteolytic activity and active-site
identifi-cation of the human cytomegalovirus protease. Eur. J. Biochem.226:361– 367.
28. Thomsen, D. R., L. Roof, and F. L. Homa.1994. Assembly of herpes simplex
virus (HSV) intermediate capsids in insect cells infected with recombinant baculoviruses expressing HSV capsid proteins. J. Virol.68:2442–2457.
29. Wakefield, D., A. Lloyd, J. Dwyer, S. Z. Salahuddin, and D. V. Ablashi.1988.
Human herpesvirus 6 and myalgic encephalomyelitis. Lancet1:1059. (Let-ter.)
30. Welch, A. R., L. M. McNally, M. R. T. Hall, and W. Gibson.1993.
Herpes-virus proteinase: site-directed mutagenesis used to study maturational, re-lease, and inactivation cleavage sites of precursor and to identify a possible catalytic site serine and histidine. J. Virol.67:7360–7372.
31. Welch, A. R., E. C. Villarreal, and W. Gibson.1995. Cytomegalovirus protein
substrates are not cleaved by the herpes simplex virus type 1 proteinase. J. Virol.69:341–347.
32. Welch, A. R., A. S. Woods, L. M. McNally, R. J. Cotter, and W. Gibson.1991.
A herpesvirus maturational proteinase, assemblin: identification of its gene, putative active site domain, and cleavage site. Proc. Natl. Acad. Sci. USA
88:10792–10796.