0022-538X/94/$04.00+0
Copyright© 1994, AmericanSocietyforMicrobiology
Physical Interaction
between the
Herpes Simplex
Virus
Type
1
Immediate-Early Regulatory
Proteins ICPO and ICP4
FENGYAO AND PRISCILLA A. SCHAFFER*
Division of Molecular Genetics, Dana-Farber CancerInstitute, and Department ofMicrobiology
and Molecular Genetics, Harvard MedicalSchool, Boston, Massachusetts 02115
Received10 March 1994/Accepted 31 August 1994
The herpes simplex virus type 1immediate-early protein ICPO enhancesexpressionofa spectrumof viral genesaloneand
synergistically
withICP4.TotestwhetherICPOand ICP4 interactphysically,weperformed far-Westernblotting analysisofproteinsfrommock-,wild-type-,and ICP4 mutant virus-infected cells within vitro-synthesized [35S]Met-labeledICPOandICP4asprobes.TheICP4andICPOpolypeptides synthesizedin vitro exhibited molecularweightssimilar to those of theircounterpartsinherpes simplexvirustype1-infected cells, and theinvitro-synthesized ICP4wasable to bind to aprobecontainingthe ICP4 consensusbinding site. Far-Westernblotting experiments demonstratedthat ICPO interactsdirectly and specificallywith ICP4and with itself. To further define the interaction between ICPO and ICP4, we generated a set of glutathione S-transferase(GST)-ICPOfusionproteinsthat contain GST andeitherICPON-terminal amino acids 1 to 244 or 1 to 394 or C-terminal amino acids 395 to 616 or 395 to 775. Using GST-ICPO fusion protein affinitychromatography and in vitro-synthesized [35S]Met-labeled ICPO and ICP4, ICP4 was shown to interact
preferentially
with the fusion protein containing ICPO C-terminal amino acids 395 to 775, whereas ICPO interactedefficiently
withboththe N-terminalGST-ICPO fusionproteinsand theC-terminal GST-ICPOfusion proteinscontaining amino acids 395to775.Fusionproteinaffinity
chromatographyalso demonstrated thatthe C-terminal 235 amino acid residues of ICP4 are important forefficientinteraction with ICPO.Collectively, these results revealadirectand specific physical interactionbetweenICPO andICP4.Regulation ofherpes simplexvirus(HSV) geneexpression
haslong been recognized to proceed inasequential andhighly regulated manner. The HSV-1 genome contains at least 70 unique genes (35) which have been divided intothree major classes, designated immediate-early, early, and late, based on the temporal order of their expression during productive
infection (29, 52). Transcription of the five immediate-early
genes is activated by VP16, a constituent of infecting virus particles(4,10, 44). Theimmediate-earlygenesaretranscribed
by cellular machinery prior to the initiation of viral protein synthesis (7,30, 48, 63).The protein products of the
immedi-ate-earlygenes,designated infected cell polypeptide 0(ICP0),
ICP4, ICP22, ICP27, and ICP47, are the major regulatory proteins of the virus. ICPO, ICP4, ICP22, and ICP27 are nuclearphosphoproteinsthat orchestrate the expression of all classes of viral genes (60). ICP0 and ICP4 are the major activators of HSV gene expression. ICP27 plays a role in regulating the expression of both early and late genes at the level of mRNAprocessing (34, 36, 39, 54, 56). Little is known about the role of ICP22 in productive infection except that it appears to enhance the level of ICPO in a cell-type-specific manner (49). ICP47 is a cytoplasmic protein of unknown function. ICP4 and ICP27, but notICPO,ICP22, orICP47, are essential for virusreplication.
Studies of temperature-sensitive and deletion mutants in the geneencoding ICP4 have shown that this protein is the major transcriptional regulatory protein of HSV-1 and that its func-tion isrequired at very early times in infection (8, 9, 11, 14, 16,
47). Withan apparentmolecular mass of 175 kDa, ICP4 is a DNA-binding protein that interacts directly with DNA at the consensus sequence, ATCGTCNNNNYCGRC (where R is a
*Corresponding author. Phone: (617) 8240. Fax: (617)
375-8255.Electronicmail address:[email protected].
purine, Y is pyrimidine, and N is anybase). Consensus ICP4
binding sites are found in the promoters of the
immediate-earlygenesencodingICPO and ICP4(15, 24,25, 40) and in the promoter of the gene specifying the familyof joint-spanning
transcripts,the L/STs (68).ICP4-DNA interactions atATCG TCNNNNYCGRC motifs near the transcription initiation sites of the ICP4, ICPO, and L/ST promoters result in the repression oftranscriptionfrom these promoters(2,14, 19, 22,
43,68).ICP4also bindsto avarietyof related sequences inthe regulatory regions of early and late genes (15, 38). Although poorly understood, the interaction of ICP4 with these related sequences appears to be critical for ICP4's ability to activate the expression of early and late genes (14, 43, 53, 58). Imbalzano et al. (31) have suggested that ICP4 may activate the expression of early and late genes by a mechanism that does not require sequence-specific DNA binding. Recent evidence has indicated that ICP4exertsboth its repressing and itsactivating activities through a mechanism involving interac-tions with the general transcription factors, TATA-binding
protein (TBP) and TFIIB, as well aswith DNA(59).
Collec-tively,available evidenceindicatesthat ICP4 exerts its repress-ing and activatrepress-ing activities differentially as a function of the location of the ATCGTCNNNNYCGRC motif within the gene, the strengthofthe interaction of ICP4 with this motif, and the kind and extent of posttranslational modification which ICP4itself undergoes (e.g., phosphorylation).
Like ICP4, the 110-kDa ICPO is a nuclear phosphoprotein
(19, 32, 45, 64). Although ICPO is not essential for virus
replication,studies ofICPO mutant viruses have indicated that functional ICPO significantly enhances the efficiency of virus replication, especially at low multiplicities of infection (3, 20,
55,
61).
Among the five HSV immediate-early regulatoryproteins, only ICPO is capable of activating all three kinetic classes of HSV genes,aswellasanumber ofheterologous viral and cellular genes, without apparentDNAsequencespecificity 8158
on November 9, 2019 by guest
http://jvi.asm.org/
lCPO
-c0-A. I I
S DN No HH P P
B. i II
C.
ICPO ICP22 ICP47
-10 -4-
-0-ICP27 ICP4 ICP4
xI
pSXCl pSPCl
pCJ06DP
pCJ07NP
-D. -_
I I I
1 00 200 300 400 500
1 2
Zinc finger
3
3 4 5
[image:2.612.148.469.67.364.2]Nuclear localization
FIG. 1. (A)Diagram of the HSV-1genomeshowing the approximate locations anddirections of transcription of HSV-1 immediate-earlygenes.
(B) Expanded diagramof the 8.4-kb SacI-XbaI fragment frommutantvirus l1OC1,which containsone copyofanintronless ICPOgene.Relevant
restriction sites:S, Sacl;D,DrdI;N, NcoI; No,NotI;H, HpaI; P, PstI;X, XbaI.(C) Beneaththe diagram of the SacI-XbaIfragmentareshown
theHSV DNAsequencesinplasmids pSPC1, pCJ06DP,and pCJ07NP, which contain the intronlessICPOgene.Thefilledrectanglerepresents
ICPOcodingsequence.(D) Function ofdomains of the ICPO polypeptide identified byEverett(19). (17, 18, 26, 41, 42, 50). Indeed, evidence todate indicates that
ICPOhas theabilityto further enhancethe expression ofany
gene that exhibits a basal level of transcription. Because
immediate-earlygeneshaveahigh basal level oftranscription,
and sinceICP4 activatestranscription ofearlyand lategenes,
ICPOserves tofurther enhanceexpression ofall threeclasses
of viralgenes.Notably,both ICP4 andICPOarepresentwithin the tegument ofpurified HSV-1 virions (66, 67), suggesting that upon entering newly infected cells, these proteins may
function to initiate viral gene expression. Consistent with its broad transactivating activity, ICPO plays a major role in
enhancing thereactivation of HSV fromlatencyin vivo(33). Moreover, of thefive immediate-early proteins, only ICPO is abletoinducereactivation of HSV-2 fromlatencyinanin vitro system(28).
Despiteitsrecognizedroles inenhancingviralgene
expres-sion during productive infection and in reactivation from latency,the mechanismsbywhichICPOactivatesgene
expres-sion alone andsynergisticallywith ICP4are unclear. Various insertion and deletion mutationshavebeen introduced into the cloned ICPOgene, and the resulting mutant forms of ICPO have beentested for theirtransactivatingactivities in transient
assays (3, 6, 19). Five functional regions of theprotein have
beenidentifiedtodatebyusingthisgenetic approach (Fig. 1). Ofparticular significance are regions 1 and 5 as defined by Everett (19). Region 1 (aminoacids 105 to222)containstwo
zincfinger bindingmotifs. Mutations inregion 1result either in complete elimination or in a significant reduction in the ability of ICPO to activate the HSV-1 gD promoter in the
absence of ICP4. Mutationsin region 5 (amino acids 633 to
775) markedly reduce the synergistic activation of the gD promoterby ICPOin thepresenceofICP4 (19). Thefunctional significance ofregion 5 in the synergistic activation of gene expression by ICPO and ICP4 is further supported by the studies of Chen et al. (6), which showed that deletion of C-terminal aminoacid residues 628 to769impaired
coopera-tive gene activation by these two proteins. Moreover, in a
search forICPO-responsive cis-actingelements inanearlygene efficientlyactivatedbyICPO,Ralphand Schafferdemonstrated that deletion of the 5' untranslated leadersequence of ICPO-responsiveHSV-1genesimpairedtheabilityofthesegenesto respondtoICPO(51). Notably,deletionof the leadersequence
also impaired responsiveness to ICP4 and to the combined effects of ICPO and ICP4. Collectively, these observations suggest that ICPO-ICP4 synergy may result from a direct
association between thetwoproteinsor anindirectinteraction mediatedbyotherviralorcellularproteins.
As partofongoingstudies designedto elucidate the mech-anism of ICPO-ICP4synergy,wehaveexaminedthepossibility that thetwoproteinsinteractphysically. Because of difficulties associated with the solubilization of ICP4 from the nuclei of HSV-1-infected cells under nondenaturingconditions (1),
co-immunoprecipitation is not the approach ofchoice to assess potential interactionsbetween ICPO and ICP4 solubilization. In thisreport,wedescribe the results of far-Westernblotting analysis and glutathione S-transferase (GST)-ICPO affinity chromatographywhich indicate that ICPOiscapableof
inter-acting directlyandspecificallywith itself andwith ICP4.
I I
600 700
on November 9, 2019 by guest
http://jvi.asm.org/
MATERIALSANDMETHODS
Cells and viruses.Vero cellsweregrown andmaintained as described previously (54). The wild-type virus, HSV-1 strain KOS, and KOS-derived ICP4 deletion mutantviruses d2 and
d156 were propagated and assayed as previously described
(14). The mutantICP4 polypeptides specified by d2and d156
are unableto interact detectablywith theconsensus sequence
ATCGTCNNNNYCGRC in gel shift assays (14). Cells
in-fectedwiththesemutantvirusesexpressonly immediate-early proteins (14).
Plasmids. Plasmid pCJ06DP contains an intronless ICPO
gene under control of the SP6 promoter in the vector pGEM-4Z (PromegaBiotec, Madison,Wis.).Thisplasmidwas constructed as follows. 11OC1 (kindly provided by Roger Everett,MRCVirologyUnit,Glasgow, UnitedKingdom)isan HSV-1mutant virus inwhichboth intronsof thegeneencoding
ICPO have been deleted (21). 11OC1 DNA was isolated and purifiedfrom infectedVerocellsas described previously (12).
The purified viral DNA was digested first with BamHI and EcoRI and thenwithSacl andXbaI (Fig. 1B).TheSacI-XbaI fragment containing the ICPO gene was isolated from an
agarose gel byelectrophoresisontoaDEAE-cellulosecolumn.
Theisolatedfragmentwasinserted intotheSacIandXbaIsites
inthe vector pGEM-4Z toyield pSXC1. ThreePstI sites are
present in pSXC1, two in viral DNA sequences and one
immediately to theright of theXbaI sitein vector sequences. PlasmidpSXC1wasdigestedwithPstI,religatedwithT4DNA ligase,anddesignatedpSPC1 (Fig.1C).TogeneratepCJ06DP, pSPC1 was digested with DrdI, treated with T4 DNA poly-merase,anddigestedwithPstI,andtheDrdI-PstIfragmentwas
then inserted attheSmaI andPstIsites inpGEM-4Z.
TogeneratepGEX-2TK0, aplasmidinwhichtheintronless
ICPOgenewasfused in frame tothe GSTgenein thevector pGEX-2TK (kindly providedbyBillKaelin, Dana-Farber
Can-cer Institute, Boston, Mass.), aplasmid, designated pCJ07NP
(Fig. 1C), was first generated by inserting the NcoI-PstI ICPO-containing fragmentofpSPC1 intotheBamHI andPstI
sites in the vector pGEM-3Z asfollows. Briefly, pSPC1 and
pGEM-3ZweredigestedwithNcol andBamHI, respectively.
Endswerefilled inwith theKlenowfragment, and the
linear-izedplasmidsweredigestedwith PstI.TheNcoI-PstIfragment containingthe ICPO gene and thevectorDNAwere purified
andligated.Blunt-endligationof theKlenow-treatedNcol and BamHI ends regenerates BamHI and NcoIsites at the
junc-tion. The BamHI-HpaI fragment containing the ICPO gene
frompCJ07NPwasthenclonedintotheBamHI andSmaIsites
ofpGEX-2TKtoproducepGEX-2TK0 (see Fig. 5).An EcoRI
site shown in Fig. 5 is derived from pGEX-2TK, which is
locatedimmediatelytotherightofthe SmaIsite inthevector
sequences.
To generate pGSTO:1-394, a plasmid expressing a hybrid
protein containing N-terminal amino acids 1 to 394 ofICPO
fused tothe GST gene, pGEX-2TK0was digestedwith NotI
and EcoRI and then subjected to treatment with Klenow fragment. The vector-containing fragment was then isolated
andrecircularized.pGSTO:1-244wasgenerated by digestionof pGEX-2TK0withSnaBI andEcoRI, endswerefilled in with
theKlenowfragment,andthevector-containing fragmentwas
thenrecircularized.TogeneratepGSTO:395-775, pGEX-2TK0
was digestedwithNotIandBamHIand treated with Klenow fragment,andthevector-containing fragmentwasisolatedand
recircularized.pGSTO:395-616wasgeneratedasfollows.
pGEX-2TKO was linearized with
Sfil,
treated with T4 DNA poly-merase,andthendigestedwithEcoRI.After fillingin with theKlenow
fragment,
thevector-containing
fragment
waspurified
and
ligated
with T4ligase.
Plasmid pCJ4-S6 contains the ICP4 gene under control of the SP6 promoter in thevector
pGEM-3Z.
Thisplasmid
was constructedby
inserting
the SalI-EcoRIfragment
containing
the ICP4 gene from
pnll
(13)
into theSall and EcoRI sites inpGEM-3Z.
In vitro transcription and translation.
Capped
mRNAsencoding
ICPOandICP4weresynthesized by
invitrotranscrip-tion of 6
,ug
ofHpaI-linearizedpCJ06DP
andpCJ4-S6,
respec-tively, using SP6 polymerase in a final volume of 50
,ul
oftranscription buffer for 30 min at
39°C
as instructedby
the manufacturer (PromegaBiotec).
Afterphenol-chloroform
ex-traction and ethanolprecipitation, RNAswere
resuspended
in 10 ,ul of 0.1% diethyl pyrocarbonate-treated distilled water. Five microliters of RNA was then used for the in vitro translation reaction with 35RI
of nuclease-treated rabbitreti-culocyte
lysate
(Promega Biotec) containing
40,uCi
of[35S]
methionine (DuPont,NENResearchProducts, Boston,
Mass.)
in avolume of 50 ,ul for 1 h at
30°C.
A2.5-,ul
sample of the translation mixture was thenanalyzed
by
sodiumdodecyl
sulfate (SDS)-polyacrylamide gel electrophoresis
(PAGE).
Preparation ofcell extracts. Vero cells were either mock infected or infected with 20 PFU of either wild-type HSV-1 strain KOS or ICP4 deletion mutant d156 per cell. At 9 h
postinfection,cellswerewashed with cold Tris-buffered saline (TBS; 137 mM NaCl, 5 mMKCl, 25 mM Tris-HCl [pH
7.4])
containing0.4 mMphenylmethylsulfonyl fluoride(PMSF)and 50 jigofL-1-chlor-3(4-tosylamido)-4-phenyl-2-butanon(TPCK)
per ml. Cells were then suspended in 500 ,ul of lysis buffer
containing 150 mM NaCl, 50 mM Tris(pH8.0), 1% Nonidet P-40(NP-40), 0.5% deoxycholate, 0.1% SDS, and 0.5%
1-mer-captoethanol in the presence of 0.4 mM PMSF, 100
,ug
of TPCK perml, and 10jig
ofleupeptin per,Il
for30minonice. Cell debriswasremovedby centrifugationat 10,000xgfor 20 min. The supernatant fluid was stored at-70°C.
DNAbinding assays.The30-bp oligonucleotide used in this
study for ICP4 DNA binding assays, 5'-GATCCCGCCCCG ATCGTCCACACGGAGCGG-3',was
synthesized by
the Mo-lecular Biology Core Facility, Dana-Farber Cancer Institute. Complementary oligonucleotides in 100 ,ulof 10 mM Tris(pH 8.0)-i mM EDTA-100 mMKClwere annealed by heating at 100°C for 5 s, cooling to 68°C, incubation for 1 h, and slow cooling to room temperature. The annealed oligonucleotides werelabeled with T4 polynucleotide kinase in the presence of [,y-32P]ATP. For ICP4 DNA binding assays, 2.5RIl
of in vitro-translated ICP4 or unprogrammed reticulocyte lysate was incubated at 25°C for 30 min with the 1 ng of32P-labeled
double-stranded oligonucleotides described above in the pres-ence of 2 ,ig of poly(dI-dC)-poly(dI-dC) in DNA binding
buffer(10% glycerol, 20 mM N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid [HEPES; pH 7.4], 50 mM NaCl, 1 mM EDTA) in a final volume of 25 ,ul.For experiments involving antibody supershift assays, after the initial30-min incubation, reactions were carried out for an additional 10 min in the presence of anti-ICP4 monoclonal antibody 58S as described by DeLuca and Schaffer (14). The protein-DNA complex was analyzed on a native 4%polyacrylamidegel containing 45 mM Tris-borate-1 mM EDTA (pH 8.0). In the DNA binding assays which utilized cell extracts, 2.5 ,ul (10 jig) of extract from mock-infected cells or cells infected with either KOS or the ICP4 mutant virusd156 was used.
Expression and purification of GST fusion proteins. Esche-richia coli BL21 cells transformed with plasmids expressing either GST orGST-ICPO fusion proteins were grown overnight in LBmedium containing 50
jig
of ampicillin per ml, dilutedon November 9, 2019 by guest
http://jvi.asm.org/
1:10 in fresh LB medium in the presence of 50 ,ug of ampicillin perml, and grownforan additional 2 h at 37°C with shaking. Protein expression was then induced by addition of 0.1 mM
isopropylthio-3-D-galactoside
(IPTG) for 2 h at 37°C. Forpurification of GST or the GSTO:1-394 fusion protein, bacte-rial cultures were centrifuged at 5,000 xgfor 10 min at 4°C and suspended in lx TBS containing0.5% NP-40, 100 ,ug of TPCK per ml, and 0.4 mM PMSF. Bacteria were lysed by
sonication for 1.5 min in three 30-s bursts at 4°C, and the
suspensionwascentrifugedat25,000xgfor 30 min at 4°C. For
purification of the GSTO:395-775 fusion protein, after centrif-ugationat5,000 xgfor 10 min at 4°C, bacteria were lysed by
three cycles of freezing and thawing in lx TBS containing
0.5% NP-40, 1 mM EDTA, 2 mM dithiothreitol, 100 ,ug of TPCK per ml, 0.4 mM PMSF, and 4 mg of lysozyme per ml. Cellular debris was removedby centrifugation at 25,000xgfor 30 min at 4°C. The supernatant fluids were aliquoted and stored in liquid nitrogen.
For purification and functional analysis of GST and
GST-ICPO fusion proteins, 1-ml aliquots of bacterial supernatant
were incubated for 1 h at 4°C with 50 ,ul of
glutathione-Sepharose beads that had been washed three times and
resuspended in TBS containing 0.5% NP-40 and 3% bovine serum albumin (BSA). After incubation, the glutathione-Sepharosebeadswerewashed three times with TBScontaining 0.5% NP-40. Beadswere thensuspendedin 1x samplebuffer
(1% SDS, 1%
P-mercaptoethanol,
0.5 M urea, and 10% sucrose in 100 mM Tris-HCl[pH 6.7]),
and eluted proteins were resolvedby SDS-PAGE.SDS-PAGE and Western blotanalysis.The
procedures
used forSDS-PAGE have been describedpreviously
(46).
Proteins were electrophoresed on 7 or 9%polyacrylamide gels
cross-linked with N,N-methylenebisacrylamide. Proteins ongels
were eithervisualized directly byCoomassie blue
staining
or transferredto hydrophobic polyvinylidene difluoride(PVDF)
membranes and immunoblotted with anti-ICPO monoclonal
antibody H1083 (kindly provided by Lenore
Pereira,
Univer-sity of California, San
Francisco)
or anti-ICPOpolyclonal
antibody J17 as describedpreviously
(67)
except that theantigen-antibody complexeswereeither reacted with
chemilu-minescence reagent as described
by
the manufacturer(Du-Pont,
NEN)
andexposed
on Kodak X-AR5 film(Fig.
4)
orvisualized by an alkaline
phosphatase
assay(Bio-Rad,
Her-cules,Calif.) (Fig. 6).
Far-Western blot
analysis
andprotein binding
assaysusing
GST fusion proteins. For far-Western blot
analysis,
proteins
separated
on 7%gels by SDS-PAGE
were electrotransferred to PVDF membranes in lxTris-glycine
buffer(25
mM Trisbase, 192 mM
glycine)
and renatured as follows. Blots werewashedoncefor 10 min with 1x
protein
binding
buffer(20
mMHEPES
[pH 7.4],
50 mMNaCl,
5 mMMgCl2,
1 mMEDTA,
0.1 mM
ZnCl2,
1 mMdithiothreitol,
10%glycerol)
and incu-batedat4°C
with fresh 1xprotein
binding
buffersequentially
for 1
h,
overnight,
and for 3 h. The blotswerethenreacted withlx
protein
binding
buffercontaining
5% BSA for 3 h at4°C
andprobed
with in vitro-translated[35S]methionine-labeled
ICPOorICP4or
equivalent
amountsofunprogrammed
reticu-locyte lysate in 10 ml of lx
protein
binding
buffer in the presence of 3% BSAovernight
at4°C.
Excessprobe
wasremoved by
washing
the blot with 10 20-ml volumes of 1x proteinbinding
buffercontaining
0.1% NP-40 for 20 minat room temperature. The blotwas then dried andexposed
to KodakX-AR5 film.Forprotein
binding
assaysusing
GST fusionproteins,
equal
amountsofGST andGST-ICPOfusion
proteins
werepurified
on
glutathione-Sepharose
beads as described above. Thepu-c 0
_ ICP4
ICPO -
e-1 2 3 4
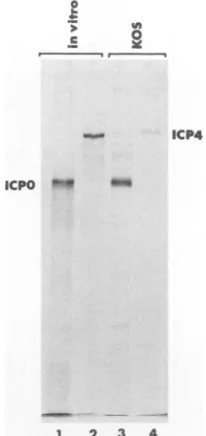
FIG. 2. Molecular
weights
of in vitro-translatedICPOandICP4.An intronlessICPOgene(derived
frommutantvirus110C1)
and the geneencoding
ICP4 werecloned into an invitrotranscription
vectorandexpressed
inrabbitreticulocyte
lysate
in thepresenceof[35S]Met,
and theresulting
proteins
were resolvedby
SDS-PAGE(lanes
1 and2).
Lanes3 and 4showICPOand ICP4
synthesized
inKOS-infected cells andimmunoprecipitated
with anti-ICPO(H1083)
oranti-ICP4(58S)
monoclonal
antibody.
rified
GSTorGST-ICPO
fusionproteins
bound tobeadswereallowed to react
overnight
at4°C
with 25,ul
of in vitro-translated[35S]Met-labeled
ICPO or ICP4 in 225,ul
of lxprotein
binding
buffercontaining
3% BSA. Beads were thenwashed three times with 1x
protein
binding
buffercontaining
0.1%
NP-40,
suspended
insample
buffer(2%
SDS,
1 M urea,2%
mercaptoethanol,
10% sucrose, 125 mM Tris[pH
6.7]),
andresolvedby
SDS-PAGE. Thegels
weredried andexposed
to Kodak
X-AR5
film.RESULTS
Properties
ofICP4 andICPOsynthesized by
invitrotran-scription-translation.
The ICP4 gene andan intronlessICPOgene were cloned
individually
into an in vitrotranscription
vector and
expressed
in a rabbitreticulocyte
lysate
as[35S]
Met-labeled
proteins.
Although
the genesencoding
ICP4
and
ICPO
specify
polypeptides
of1,298
and 775 aminoacids,
respectively,
withpredicted
molecular masses of132,835
Da(for
ICP4)
and78,452
Da(for
ICPO),
theICP4
and ICPOproteins
synthesized
in HSV-1-infected cellsmigrate
aspro-teins with
apparent
molecular masses of 175 and 110kDa,
respectively,
by
SDS-PAGE. To determine if the molecularweights
of the ICP4 andICPOspecies
translatedin vitroweresimilar to those of their
counterparts
expressed
in HSV-1-infectedcells,
proteins
from thetwosources werecompared by
SDS-PAGE.As shownin
Fig.
2,
theICP4
andICPOmoleculessynthesized
in vitro were stable and exhibited molecularweights
similar to those theircounterparts
synthesized
inKOS-infected cells. These
findings
suggest
that theapparent
differences observed in the
predicted
molecularweights
and themolecularweights
seen onSDS-PAGEaredueprimarily
toinherent
properties
of thetwoproteins,
such ashigh
proline
content.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.384.487.75.293.2]Anti-ICP4 * 0
Cz z
E °0
°J
u a_.Y .
-m
...s
g.AiN
>e
-j
*,
5F
vF ICP41 2 3 4 5 6 7 5 9
FIG. 3. Gel shiftassaywithin vitro-translated ICP4. Extracts from mock-infected cells(lane2),cellsinfectedwithwild-typeHSV-1strain KOS(lanes4and5)ortheICP4deletionmutantvirusd156(lane 3), in vitro-translated ICP4(lanes7and8),andunprogrammed reticulo-cytelysate(lanes6and 9)wereincubatedwitha32P-labeled double-strandedoligonucleotide containingtheICP4binding site,ATCGTC. Lane 1containsprobealone.Insupershift experiments(designated by dots above lanes5, 8, and 9),after the initial 30-min incubation of
extractwithprobe,reactionswerecarriedoutforanadditional10 min
inthepresenceofanti-ICP4 monoclonalantibody58S.
Interaction ofinvitro-translated ICP4witha
probe
contain-ing
theICP4consensusbinding site,
ATCGTCNNNNYCGRC. To determine whether theICP4synthesized
invitrowasable to interact with its consensus DNAbinding site, gel
shiftexperiments
wereperformed.
As shown inFig. 3,
the ICP4 translated in vitro boundto asynthetic probe containing
theconsensus sequence ATCGTCNNNNYCGRC
(lane 7),
andthe
ICP4/DNA complex
wassupershifted by
addition of anti-ICP4 monoclonalantibody
(lane 8)
butnotanti-ICPO
mono-clonalantibody (data
notshown).
Moreover, the ICP4-DNAcomplex
formed whenKOS-infectedcellextract wasusedas a source of ICP4(lane
4)
exhibited amobility
on nativepoly-acrylamide gels
similar to that of the ICP4-DNA complexformed
by
invitro-translated ICP4 with thesameprobe(lane
7).
Thesupershifted
bands were also of similar mobilities(compare
lanes 5 and8).
As anticipated,themutantform of ICP4specified by
d156 bound poorly to the probe (lane 3). These observations demonstrate that in vitro-translated ICP4 isabletobindto aprobe containingaconsensusICP4binding site.Moreover,
because of the similarities in the migrationpatterns of the
complexes
shown in lanes 4 and 7 and lanes 5 and8,
otherviralproteins
arelikely
not associated with the ICP4-DNAcomplex
formed with infected cell extracts.Physical
interaction ofICPOwithICP4and withitself.In an efforttoinvestigate
thepossibility
thatICPO
andICP4interactphysically,
far-Westernblotting analysis
was performed. The ICP4deletionmutantd2wasused in thesetests as anegativecontrol because the d2
polypeptide
has been shown to beincapable of down-regulating the expression of
immediate-early genes or activating the transcription of early and late genes.Thus, in d2-infectedcells, thesynthesis of immediate-early proteins is significantly enhanced and the synthesis of
earlyand lateproteinsis undetectable (14).
Figure4Ashows the results of Western blot
analysis
of theprotein blot shown in
Fig.
4C, using
anti-ICPO monoclonalantibody. Consistentwith the
previous
observation of DeLuca andSchaffer(14),
the level ofICPOexpression
at 9 hpostin-fection was
significantly higher
in d2-infected cells than inKOS-infected cells. The far-Western blots shown in
Fig.
4B and C demonstrate that aprotein expressed
in d2-infectedcells,withamolecular
weight
similartothat ofICPO,
interacts with invitro-translatedICPO and ICP4(designated by
dots in theright-hand
margins)
but notwithproteins
in anunpro-grammed
reticulocyte
lysate
(Fig. 4D). Although
Western blotanalysis
inconjunction
with Coomassie bluestaining
indicated that themutantICP4peptide
specified by
d2(with
anapparent molecularmassof -160kDa)
(data
notshown and reference14), ICP22, andICP27
(14)
werepresentathigh
levels ind2-infected cells, these
peptides
did not interact with in vitro-translatedICPOorICP4(Fig.
4B andC).
When in vitro-trans-lated[35S]Met-labeled
HSV-1UL8.5protein
wasusedas aprobe,
far-Westernblot
analysis
of d2-infected cell extractsrevealed littleto nointeraction betweenUL8.5protein
andICPO(data
not
shown).
Itshouldbe noted thattheheavy
bandmigrating
below the ICPO bandis
nonspecific,
asitis present inFig.
4B toDaswellas inmock-infected and infected cellextracts.Collectively,
theseobservations indicate that ICPO interactsspecifically
with ICP4 and with itself.Notably,
in addition to interactingwithICPO,
the in vitro-translated ICP4 also inter-actedspecifically
with a cellularprotein
with an apparent molecularmassof 130 kDa(Fig. 4B).
Wehavedesignated
this cellularprotein
p130.
Little or no interaction was detected betweenp130
andICPO(Fig.
4C).Interaction ofICPO and ICP4 withGST-ICPO fusion pro-teins.
Having
shown that ICPO interacts with ICP4 andwithitselfby far-Western blotting analysis, we next attempted to confirm and further define these interactions by using
GST-ICPO fusion proteins. Initially, an intronless ICPO gene was fused in frametotheGSTgene
(pGEX-2TKO; Fig. 5),
andthefusion
protein
wasexpressed
in several different strains of E.coli. The
full-length
GST-ICPO fusionprotein proved
to be highlyunstable whenexpressedinbacteria. Toovercomethisproblem, we generated two GST-ICPO fusion proteins that contain GST and either N-terminal amino acids 1 to 394 or C-terminal amino acids 395 to 775 of ICPO
(Fig. 5).
These fusionproteins
weredesignated
GSTO:1-394
andGSTO:395-775, respectively.
GST and thetwoGST-ICPOfusionproteins wereexpressed
in E. coli BL21 and purified onglutathione-Sepharose
4B beads. Thepurified proteins
were resolvedbySDS-PAGE and visualizeddirectly byCoomassiebluestaining
(Fig. 6A)
ortransferredtoPVDFmembranes and subjected to Westernblot analysiswithanti-ICPO
polyclonal antibodyJ17(Fig. 6B)
or monoclonal antibody H1083 (Fig. 6C). On the basis of theirmigrationpatternsingelsandtheirreactivity withanti-ICPO antibodies
(Fig.
6B and C), the bands marked by dots in lanes 1 and 2 ofFig. 6A representfull-length fusionproteinsGSTO:1-394 and
GSTO:395-775,
respectively.Faster-migratingbandsbelow the dotsmostlikelyrepresent degrada-tion products of GSTO:1-394 and GSTO:395-775. The poly-clonal
antibody
J17reacted with bothGSTO:1-394andGSTO:395-775 and their
degradation products (Fig. 6B);
however,monoclonalantibodyH1083 reactedonlywith the
GSTO:395-775fusion
protein
(Fig. 6C).
This observation isconsistent withon November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.87.269.72.343.2]A.
Anti -ICPO
8 C
0 E l
B. 35S IC D.
P4 Cs 35S lCpo
Y Y
17S 175 Omm
m
-_.~_F_ pN
I .. 6
1753j
--
m~
. * ..~0
_ _-q
__ _
...
_i
__wwww. l
1 2 3 4 1 2 3 4 1 2 3 4
FIG. 4. ICPOinteracts directly with ICP4 and with itself. Vero cells were either mock infected (lanes 2), infected with wild-type HSV-1 strain KOS (lanes 3),orinfected with ICP4 mutant d2 (lanes 4) at a multiplicity of 20 PFU per cell. At 9 h postinfection, cells were harvested and proteins were resolved by SDS-PAGE and transferred to PVDF membranes. Lanes 1 (dots) contain in vitro-translated ICP4 as a marker. After renaturation, blots were probed with in vitro-translated [35S]Met-labeled ICP4 (B), in vitro-translated [35S]Met-labeled ICPO (C), or [35S]Met-labeledunprogrammed reticulocyte lysate (D). The dots on the right of panel B and C indicate the locations of proteins that interact with 'S-labeledICP4 andICPO,respectively. Panel A shows aWesternblotanalysis of the blot shown in panel C, using theanti-ICPOmonoclonal antibody H1083todetectICPO-specificpeptides. Sizes are indicated in kilodaltons.
previous studies demonstrating that H1083 recognizes a
C-terminal epitopeofICPO (65).
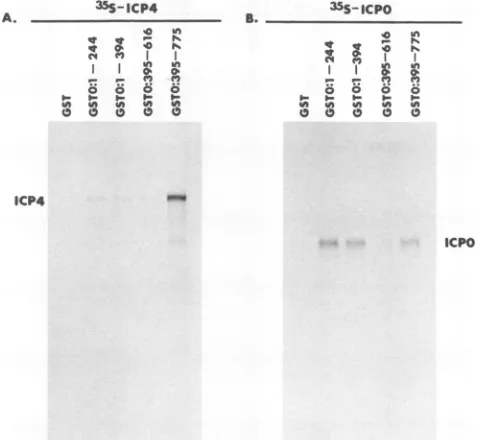
Totesttheir abilities to interact with ICP4 andICPO, GST,
GSTO:1-394,andGSTO:395-775werepurifiedon
glutathione-Sepharosebeads and allowed toreactwith in vitro-translated
full-length[35S]Met-labeledICP4(Fig.7A) orICPO (Fig.7B). The boundproteinswereanalyzed by SDS-PAGE and
visual-ized by autoradiography. The intensities of the
[35S]Met-labeled bands in each lane werequantified byPhosphorlmager scanning(model400 EPhosphorImager; MolecularDynamics, Sunnyvale,Calif.).These data indicated that the interaction of
BamHI SnaBi NotI Sfi1 EDoR
pGEX-2TKO
394 775
pGSTO:1-244 Ei _
1 244
pGSTO:1-394
1 394
pGSTO :395-616
pGSTO:395-775
395 616
395 775
FIG. 5. CompositionsofGST-ICPO fusionproteins. Plasmid pGEX-2TKOcontainsanintronlessICPOgenewasfusedin frametothe GST gene in vector
pGEX2TK.
The GSTportion ofthe hybrid protein productisshownas anopenbox,and theICPOaminoacidsequence is shownas ablackbox. TheGST-ICPOfusionproteinsGSTO:1-244,GSTO:1-394, GSTO:395-616, and GSTO:395-775 were generated as
describedinMaterials andMethods.
ICP4 withGSTO:395-775was85-fold greater than that of ICP4 with GST alone and 13-fold greater than that of ICP4 with
GSTO:1-394. As shown in Fig. 7A, in vitro-translated ICP4 interacted strongly with the fusion protein containing
ICPO
C-terminal amino acids 395to775. In these tests, about 15to 30%ofinputICP4wasretainedontheGSTO:395-775column. Notably, ICP4 interacted significantly less strongly with the fusion protein containing ICPO N-terminal amino acids 1 to 394. In Fig. 7B, ICPO interacted equally well with both the N-terminal and C-terminal
GST-ICPO
fusionproteins.In an effort to map the domains of
ICPO
required for its interaction with ICP4 and with itself more precisely, two additional GST-ICPO fusion proteins were generated. Theseproteins contain GST andeitherN-terminalICPOamino acids 1 to 244(GSTO:1-244) orC-terminal amino acids 395 to 616
(GSTO:395-616) (Fig. 5).Todetermine the effect of deletion of amino acids 245 to 394 from fusion protein GSTO:1-394 or amino acids 617 to 775 from GSTO:395-775 on the abilityof theseproteinstointeract with ICP4 and
ICPO, equal
amounts of GST and GST-ICPO fusion proteins GSTO:1-244, GSTO:1-394, GSTO:395-616, and GSTO:395-775 werepurified
on glu-tathione-Sepharose beads and reacted with either invitro-translated 35S-labeled ICP4
(Fig.
8A)
or ICPO(Fig. 8B).
As demonstrated inFig. 8A, deletion of amino acids 617 to775 from fusion protein GSTO:395-775significantly impaired
itsability to interact with ICP4, whereas, aswith
GSTO:1-394,
aweak interactionwasdetected between GSTO:1-244 and ICP4. As shown inFig. 8B,deletion of amino acids 245to 394 from
GSTO:1-394 had no effect on the ability of the
protein
toassociate with invitro-translated
ICPO,
whereas little interac-tion was detected between the in vitro-translated ICPO and fusionprotein GSTO:395-616,demonstrating
arole for amino355-NoRNA
9
u In
° O "C
* X Y 1
1 2 3 4
1
ICPO
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.115.495.73.302.2] [image:6.612.55.299.530.655.2]Anti-ICPO A. Coomassieblue
stain_
in
N o,
C'I
0 0
.- P- I-.. on V
0 0 0
K polyclonal
In
0' Ln
I.. I.- I In
°I
WI0 00
monoclonal
b
I
in VI
0 0 0
C.
205-
116-80-~~
I...
49-3=~~~~~~~~~~~~S
[image:7.612.148.483.74.381.2]1 2 3 1 2 3 1 2 3
FIG. 6. Expressionand purification of GST-ICPOfusionproteins GSTO:1-394 andGSTO:395-775. GST-ICPO fusionproteins GSTO:395-775 (lanes 1)andGSTO:1-394(lanes 2),aswellasGST(lanes 3),wereexpressedin E. coli andpurified by using glutathione-Sepharosebeads. The purified proteinswereresolvedbySDS-PAGEand either visualizeddirectly byCoomassie bluestaining (A)ortransferredtoPVDFmembranes andsubjected toWestern blotanalysiswith anti-ICPO polyclonal antibodyJ17(B) ormonoclonal antibodyH1083 (C). Sizesare indicated in kilodaltons.
acids 616to775, butnot245to394,inthe interaction of ICPO with itself.
Interactions of ICP4 and truncated ICP4 peptides with GSTO:1-394 and GSTO:395-775. ICP4 contains multiple func-tional domains thatarerequiredforautoregulation, activation
ofearly and late geneexpression, nuclearlocalization, DNA
binding, intranuclear distribution, andvirion association
(14,
65). To determine whether a particular domain or primary aminoacid sequence within the ICP4proteinis critical for the interaction ofICP4withICPO,three truncated ICP4 polypep-tidesspecifyingamino acids 1 to522, 1 to681, and 1 to 1063 weresynthesized invitro and labeled with[35S]Met.
Approxi-mately equalamountsof[35S]Met-labeled full-length ICP4 and
truncated ICP4 peptides were then allowed to interact with GSTor theGST-ICP0 fusion proteins bound to
glutathione-Sepharose beads, andbound proteinswere analyzed by SDS-PAGE (Fig. 9).
In this experiment, the wild-type form of ICP4 interacted moreefficientlywithGSTO:395-775 than did the mutant forms of ICP4(Fig. 9; compare lane 6 with lanes 7 to 9). Moreover, just as in Fig. 7A and 8A, the wild-type form of ICP4 again
interacted more efficiently with the C-terminal GST-ICPO
fusion protein than with the N-terminal fusion protein (Fig.
8A; compare lanes 2 and6). The 1,298-amino-acidwild-type ICP4 protein interacted strongly with the
ICP0
C-terminal fusion proteinGSTO:395-775,
yet scanning analysis revealed that the truncated ICP4protein (amino acids 1 1063) lackingthe 235 C-terminal amino acid residues interacted -10-fold less strongly, suggesting that the ICP4 domain that interacts with ICPO may include C-terminal amino acid residues 1064to 1296. Alternatively, the intact ICP4 protein may be required for optimal interaction withICPO.
DISCUSSION
Several lines of evidence suggest that functional interactions mayoccuramongHSV-1immediate-early proteins. First,ICPO and ICP4 activate gene expression synergistically, and ICP27 functionsas eitheratrans activator of late geneexpressionor a trans repressor of early gene expression induced by ICP4,
ICPO, or a combination of ICP4 and ICPO (57, 62). Second, ICP27 has been shown to playarole inthe posttranslational modification of ICP4 during productive infection (37, 62). Third,theintranuclear localizationofICPOhas been reported to be affected by ICP4 (27, 32). Lastly, recent evidence has
demonstratedthat ICP4facilitates,whereas ICP27 inhibits, the nuclearlocalization ofICPO (69). To elucidate the molecular mechanism underlying the cooperative regulation of HSV-1 gene expression mediated by immediate-early proteins, we have investigated the possibility that direct physical interac-tionsoccuramongthe HSV-1immediate-earlyproteins. Using far-Westernblotting analysis,GST-ICPO fusionprotein affinity chromatography, and in vitro-translated ICPO and ICP4,we haveshown that ICPO isabletointeractspecificallywith ICP4 iffommus
9-*NRUIW
.i.: 0
on November 9, 2019 by guest
http://jvi.asm.org/
A. 35S-ICP4
..
aI
0vII 0
3.-
-00 00
B. 35S-lCPO
In
r%
N
I al.
%M C.)
ON
H 0 0
ICP4
_ _
A 35S-ICP4
-.0
4 ell m
I
_: _ n o is a
C4 I-
I-ICP4
In
Oh 0
B. 3S-lCPO
B._
.0
HI
N.
VI VI VI
0mCM
ICPO
1 2 3 4 1 2 3 4
FIG. 7. Interactions of ICPO and ICP4 with GST-ICPO fusion
proteins. GST (lanes 3)and the GST-ICP0 fusionproteins GSTO:1-394
(lanes 2) and GSTO:395-775 (lanes 1)were expressed in E. coliand
purified on glutathione-Sepharose beads. The purified GST and
GST-ICPO fusionproteins boundtobeadswereallowedto reactwith
invitro-translated [35S]Met-labeledICP4 (A)orICPO (B)overnightat
4°C.The boundproteinswereresolvedby SDS-PAGE and visualized by autoradiography.Invitro-translated [3 S]Met-labeled ICP4orICPO
(lanes4)wasused asamarker andwasnot reacted with the affinity column.
and with itself. Furthermore, the C-terminal portions of the twoproteinsappeartoplay important roles in mediating these interactions.
ICP4 isasequence-specific DNA-binding protein. Relative
to its high-affinity consensus DNA binding site, ATCGTCN NNNYGRC, othersequences towhich ICP4 binds arehighly
degenerate such that thebinding affinityof ICP4tothese sites
can vary dramatically depending on the sequence of the site (15, 38). High-affinity binding sites are present within the promotersof thegenes specifying ICP4, ICPO, and theL/STs and are absent from thepromoter regions ofmostearlyand
late genes. Because the expression ofearly and lategenes is
inducedby ICP4, andmutantforms ofICP4impaired in the abilitytointeract with DNA exhibit littleor notransactivating
activity (14, 43, 58),it islikelythat under certaincircumstances, the interaction of ICP4 with itslow-affinity bindingsitesinthe promoters and/or 5' untranslated regions of early and late
genesplaysan importantrole inmediatingthetransactivating
effect of ICP4 on thesegenes (15, 53, 58). Given itssynergy
with ICPO inactivatinggeneexpression and therecent obser-vation that ICP4canformtripartite complexeswith the basic transcription factors,TBP and TFIIB,on atemplate
contain-ing both a TATA sequence and the ICP4 binding site, it is
possible that theinteraction of ICPO with ICP4 facilitates the interaction of ICP4 with itslow-affinityDNAbindingsite and with factors in the basictranscription complexsuchasTBP and TFIIB (59).
ICPO isa potent transactivator of all three classes of HSV
genes and is associated with chromatin in infected cells in a
[image:8.612.78.278.74.310.2]manner that is not sequence specific (23, 45). Moreover,
FIG. 8. Effects of C-terminal truncationsof GST0:1-394 and GSTO:
395-775ontheir abilitiestointeract with ICP4 and ICPO. GST and the GST-ICP0 fusion proteinsGST0:1-244, GST0:1-394, GST0:395-616, andGST0:395-775werepurifiedonglutathione-Sepharosebeads and
reacted with either in vitro-translated [35S]Met-labeledICP4 (A) or
ICPO (B). After several washes with 1x protein binding buffer containing 0.1%NP-40, the bound proteinswere analyzed by
SDS-PAGE andvisualized byautoradiography.
purified, baculovirus-expressed ICPO is ableto associate with DNA-cellulose columns, although it failsto form stable
pro-tein-DNA complexes in solution (23). These observations suggestthatstableICPO-DNA complex formationmayrequire both anonspecific, low-affinity interaction of ICPOwith DNA
aswell as aninteraction withDNA-bound protein(s).
Prelim-inarystudiesfrom this laboratorysuggestthat like ICP4, ICPO
canactivategeneexpression throughamechanisminvolvingan
interaction with TBP and association with DNA(unpublished observations). From the observation reported herein, that ICPO and ICP4 can interact directly with each other, it is reasonabletospeculatethatthe interaction of ICP4 with ICPO
mayservetostabilize the association of ICPO withDNA,which in turn leads to synergistic gene activation. We are currently
investigatingthispossibility.
Duringthepast5years,considerable evidence has
accumu-latedto indicate that the C-terminal half of ICPO is criticalin mediatingthe several functions of thisprotein. Thus,this half of the ICPO molecule contains multiple functional domains, including (i) asignal requiredfor nuclear localizationmapped to amino acids 474 to 509 (19); (ii) a region required for efficient transactivation mappedtoamino acid residues 526 to 720(3);and(iii)aregionwithin amino acid residues 720 to 770 required for the intranuclear localization of ICPO(19, 69),its association withtheHSV-1virion(65), andresponsivenessto the inhibitory effect ofICP27 on the nuclear localization of ICPO (69) (Fig. 1). Using GST-ICPO fusion protein affinity chromatography,wehave shown that ICP4 interacts preferen-tiallywith C-terminal aminoacid residues 395 to 775 of ICPO.
The observation that deletionof amino acids 617to 775 from
fusion protein GSTO:395-775 significantly impaired its ability to interact with ICP4 suggeststhat amino acids 617 to 775 of
ICPO mayrepresent themajordomain for the physical
inter-ICPO
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.317.556.76.296.2]GST GST0 1-394 GSTO:395-775
L IL, 'O IL >in UUI U l l
ICP4
1 2 3 4 5 6 7 8 9
FIG. 9. Effects of C-terminal deletions on the ability of ICP4 to
interact withICPO. GST (lane 1) andthe GST-ICPO fusion proteins GSTO:1-394 (lanes 2 to 5) and GSTO:395-775 (lanes 6 to 9) were
purifiedonglutathione-Sepharose beads and reactedat4°Covernight
with in vitro-translated[35S]Met-labeledwild-type ICP4(lanes2and 6)
orwith truncated ICP4polypeptidesspecifyingamino acids 1to 522
(lanes3 and 7),1to1063(lanes4and8), and1to681(lanes5 and 9).
The beadswerewashed, and boundproteinswere resolved by
SDS-PAGEand visualizedbyautoradiography.
action of ICPOwith ICP4. Notably, this region of ICPO also contains the domain involved in the synergistic activation of
geneexpressionwithICP4(19). Together, these observations strongly support the hypothesis that the synergisticeffects of ICPOand ICP4result fromadirect interactionbetween these
twoproteins.
Byusing glycerol gradient centrifugation(23) and Sephacryl chromatographyelutiontechniques (5), ICPOhas been shown tooligomerizein solution.Theobservation thatamutantICPO polypeptide specifying only N-terminal amino acid residues 1 to525 and lackingthe virion associationdomain (residues 720
to 770) is able to inhibit association of wild-type ICP0 with
HSV-1 virions alsosuggests that ICPOcan dimerize and that
thedimerizationdomain lies within the N-terminal portionof
the molecule (65). Using far-Western blotting analysis and
GST-ICPO affinitychromatography, in this reportwe provide
directevidencethatICPOcaninteract with itself. Furthermore,
we have shown that in contrast to ICP4, ICPO is capable of interacting both with N-terminal GST-ICPO fusion proteins
GSTO:1-244and GSTO:1-394 and with C-terminal GST-ICPO fusion protein GSTO:395-775, whereas little interaction was detected between ICPOand GST-ICP0 fusion protein GSTO: 395-616.Theseresultsindicate thatICPOis abletooligomerize
andthat bothN-terminalamino acids1 to244and C-terminal
amino acids617 to775 ofICPO areinvolved in this process. Studies of temperature-sensitive, deletion, and nonsense mutantshaveclearlydemonstrated that theC-terminal portion of ICP4(aminoacid residues775 to1298)plays acritical role
in the multiple functions ofthis protein. This region of the proteinspecifies(i)autoregulation and activation of early and
late gene expression; (ii) intranuclear localization (14); and
(iii) virion association, which has been mapped to C-terminal
amino acid residues 1029 to1298(65).Deletion of these amino acid residues and the presence of
temperature-sensitive
muta-tions in this regionof the proteindrasticallyimpair
theability
of ICP4 to interactwith DNA, suggestingthatresidues within the C-terminal portion of ICP4 alsoplay an important role in controlling theoverallconformation of the ICP4 molecule
(11,
12, 14). The studies described in this report demonstrate that C-terminal amino acid residues 1064 to 1298 of ICP4 are required for the efficient interaction of ICP4 with ICPO.
Although the findings presented herein indicate that the physical interaction between
ICP0
and ICP4 isrequiredforthesynergistic activity of these two proteins, definitive functional
evidence to support thishypothesis is not yet available. Such
evidence will require the isolation of mutantformsof ICP4 or
ICPO that are unable to interact physically but that maintain full transactivating activity.
ACKNOWLEDGMENTS
We thank C. Dabrowski, M. Hardwicke, and R. Jordan for helpful discussion, R. Courtney and L. Pereira for providing useful antibodies, and M. Shea for manuscript preparation.
These studies were supported by research grant R37-CA20260 from the National Cancer Institute. F.Y. is the recipient of a postdoctoral fellowship from the National Multiple Sclerosis Society.
REFERENCES
1. Ackermann, M., D. K. Braun, L. Pereira, and B. Roizman. 1984. Characterization of herpes simplex virus 1 a proteins 0, 4, and 27 with monoclonal antibodies. J. Virol. 52:108-118.
2. Bohenzky, R. A., A. G. Papavassiliou,
I.
H. Gelman, and S. Silverstein. 1993. Identification of a promoter mapping within the reiterated sequences that flank the herpes simplex virus type 1 UL region. J. Virol. 67:632-642.3. Cai, W., and P. A.Schaffer.1989. Herpes simplex virus type 1ICPO plays a critical role in the de novo synthesis of infectious virus following transfection of viral DNA. J. Virol. 63:4579-4589. 4. Campbell, M. E. M., J. W.Palfreyman,and C. M. Preston. 1984.
Identification of herpes simplex virus DNA sequences which encode a trans-acting polypeptide responsible for stimulation of immediate early transcription. J. Mol. Biol. 180:1-19.
5. Chen, J., C. Panagiotidis, and S. Silverstein. 1992. Multimeriza-tion of ICPO, a herpes simplex virus immediate-early protein. J. Virol. 66:5598-5602.
6. Chen, J., X. Zhu, and S. Silverstein. 1991. Mutational analysis of the sequence encoding ICPOfrom herpes simplex virus type 1. Virology 180:207-220.
7. Clements, J. B., R. J. Watson, and N. M. Wilkie. 1977. Temporal regulation of herpes simplex virus type 1 transcription: location of transcripts in the viral genome. Cell 12:275-285.
8. Courtney, R. J., and M.
Benyesh-Melnick.
1974. Isolation and characterization of a large molecular weight polypeptide of herpes simplex virus type-1.Virology 62:539-551.9. Courtney, R. J., P. A.Schaffer,and K. L. Powell. 1976. Synthesis of virus-specific polypeptides by temperature-sensitive mutants of herpes simplex virus type 1. Virology75:306-318.
10. Dalrymple, M. A., D. J. McGeoch, A. J. Davison, and C. M. Preston. 1985. DNA sequence of the herpes simplex virus type 1 gene whose product is responsible for transcriptional activation of immediate early promoters. Nucleic Acids Res. 13:7865-7879. 11. DeLuca, N., A. M. McCarthy, and P. A. Schaffer. 1985. Isolation
and characterization of deletion mutants of herpes simplex virus type 1 in the gene encoding immediate-early regulatory protein ICP4. J. Virol. 56:558-570.
12. DeLuca, N. A., M. A. Courtney, and P. A.Schaffer.1984. Temper-ature-sensitive mutants in herpes simplex virus type 1 ICP4 permissive for early gene expression. J. Virol. 52:767-776. 13. DeLuca, N. A., and P. A. Schaffer. 1987. Activities of herpes
simplex virus type 1 (HSV-1) ICP4 genes specifying nonsense polypeptides. Nucleic Acids Res. 15:4491-4511.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:9.612.102.262.77.309.2]14. DeLuca, N. A., and P. A. Schaffer. 1988. Physical andfunctional domains of the herpes simplex virus transcriptional regulatory protein ICP4. J.Virol.62:732-743.
15. DiDonato, J. A., J. R. Spitzner, and M. T. Muller. 1991. A predictive model forDNArecognition by the herpes simplex virus protein ICP4. J. Mol. Biol. 219:451-470.
16. Dixon, R. A. F., and P. A. Schaffer. 1980.Finestructuremapping and functional analysis of temperature-sensitive mutants in the gene encoding the herpes simplexvirus type 1 immediate-early proteinVP175. J. Virol. 36:189-203.
17. Everett, R D. 1984. Trans-activation of transcription by herpes virus products: requirement for two HSV-1 immediate-early polypeptidesfor maximumactivity. EMBO J. 3:3135-3141. 18. Everett, R. D. 1986. Theproducts of herpes simplexvirus type 1
(HSV-1) immediate-early genes 1, 2, and 3 can activate HSV-1 geneexpressionintrans. J.Gen. Virol. 67:2507-2513.
19. Everett, R D. 1988. Analysis of the functional domains of herpes simplexvirus type 1immediate-earlypolypeptideVmwllO. J.Mol. Biol. 202:87-96.
20. Everett, R D. 1989. Construction and characterizationofherpes simplex virus type 1 mutantswith defined lesions in immediate-earlygene 1. J. Gen. Virol. 70:1185-1202.
21. Everett, R. D. 1991. Constructionand characterizationofherpes simplextype 1viruseswithout intronsin immediateearlygene 1. J.Gen. Virol.72:651-659.
22. Everett,RD.,and A.Orr.1991. TheVmwl75bindingsite in the IE-1 promoterhasnoapparentrolein theexpression ofVmwllO during herpes simplex virus type 1 infection. Virology 180:509-517.
23. Everett,R D.,A.Orr,and M. Elliot. 1991.Highlevelexpression and purification of herpes simplexvirus type 1 immediate early polypeptideVmwllO. Nucleic AcidsRes. 19:6155-6161. 24. Faber, S. W.,and K. W.Wilcox. 1986.Association ofthe herpes
simplex virus regulatory protein ICP4 with specific nucleotide sequences inDNA. Nucleic AcidsRes. 14:6067-6083.
25. Faber, S. W.,and K.W. Wilcox.1988.Association ofherpesvirus regulatory proteinICP4withsequences spanningthe ICP4gene transcriptioninitiationsite. NucleicAcids Res.16:555-570. 26. Gelman,L.H.,andS. Silverstein. 1985.Identification of
immedi-ate-early genes from herpes simplexvirus that transactivate the virus thymidinekinase gene. Proc.Natl. Acad. Sci. USA 82:5265-5269.
27. Gelman,L.H.,and S.Silverstein.1986. Co-ordinateregulationof herpessimplexvirus geneexpressionismediatedbythe functional interaction oftwo immediate-early gene products. J. Mol. Biol. 191:395-409.
28. Harris, R A., R D. Everett, X. Zhu, S. Silverstein, and C. M. Preston. 1989. Herpessimplexvirus type 1
immediate-early
pro-teinVmwl 10reactivates latentherpessimplexvirustype2inanin vitrolatencysystem. J. Virol. 63:3513-3515.29. Honess,RW.,and B. Roizman. 1974. Regulationof
herpesvirus
macromolecularsynthesis.I.Cascaderegulationof the
synthesis
of three groupsof viralproteins.J. Virol. 14:8-19.30. Honess,RW.,and B. Roizman. 1975.
Regulation
ofherpes
virus macromolecular synthesis: sequential transition ofpolypeptide
synthesisrequiresfunctional viralpolypeptides.Proc.Natl.Acad. Sci. USA72:1276-1280.31. Imbalzano,A. N.,A. Shepard, and N. DeLuca. 1990. Functional relevance of specific interactions between
herpes
simplex
virus type 1ICP4 andsequencesfrom thepromoter-regulatory
domain ofthe viral thymidinekinasegene.J.Virol.64:2620-2631. 32. Knipe,D.M.,andJ.LSmith. 1986.Amutantherpesvirus
protein
leadsto ablockinnuclearlocalization of otherviral
proteins.
Mol. Cell.Biol.6:2371-2381.33. Leib,D.A.,D. M.Coen,C.L.Bogard,K.A.Hicks,D.R
Yager,
D. M. Knipe,K. L.Tyler, andP. A. Schaffer. 1989. Immediate-early regulatory gene mutants define different stages in the establishment and reactivationofherpes
simplex
viruslatency.
J. Virol. 63:759-768.34. McCarthy,A.M.,L.McMahan,and P. A.Schaffer. 1989.
Herpes
simplex virus type 1 ICP27 deletion mutants exhibit altered patternsoftranscriptionandareDNAdeficient.J.Virol. 63:18-27.35. McGeoch,D.J.,M. A.Dalrymple,A.J.Davison,A.Dolan,M. C. Frame,D.McNab,L.J.Perry, J.E.Scott,and P.Taylor.1988.The
complete
DNA sequenceof thelong unique
regionin thegenome ofherpes simplexvirustype1.J. Gen. Virol. 69:1531-1574. 36. McLauchlan, J.,A. Phelan,C.Loney, R M. Sandri-Goldin,andJ. B. Clements. 1992. Herpes
simplex
virus IE63 acts as the posttranscriptionalleveltostimulate viral mRNA 3'processing.
J. Virol. 66:6939-6945.37. McMahan, L., and P. A. Schaffer. 1990. The
repressing
and enhancingfunctionsof theherpessimplex
virusregulatory
protein
ICP27 maptoC-terminalregions and are
required
to modulate viral gene expression veryearly in infection. J. Virol. 64:3471-3485.38. Michael, N.,D.Spector,P.Mavromara-Nazos,T.M.Kristie,and B. Roizman. 1988. The DNA
binding properties
of themajor
regulatoryprotein
a4ofherpes
simplex
viruses. Science 239:1531-1534.39. Mosca, J. D., P. M. Pitha, and G. S. Hayward. 1992.
Herpes
simplex
virus infectionselectively
stimulatesaccumulation of beta interferon reportergene mRNAby
aposttranscriptional
mecha-nism. J.Virol. 66:3811-3822.40. Muller,M. T. 1987.
Binding
of theherpes simplex
virusimmedi-ate-early
geneproduct
ICP4toitsowntranscription
startsite.J. Virol. 61:858-865.41. Nabel, G. J., S. A. Rice, D. M.Knipe, and D. Baltimore. 1988. Alternative mechanisms for activation of human immunodefi-ciencyvirusenhancerin T cells. Science239:1299-1302. 42. O'Hare, P.,and G. S.Hayward.1985.Evidenceforadirectrole for
both the 175,000-and
110,000-molecular-weight
immediate-early
proteins
ofherpes
simplex
virusinthe transactivationofdelayed-early
promoters.J.Virol. 53:751-760.43. Paterson,T.,and R D. Everett.1988.Mutational dissection of the HSV-1
immediate-early
protein
Vmwl75 involved in transcrip-tionaltransactivation andrepression. Virology
166:186-196. 44. Pellet,P.E.,J.L C.McKnight,
F.J.Jenkins, andB. Roizman.1985.Nucleotidesequenceand
predicted
amino acidsequenceofa
protein
encoded inasmallherpes
simplex
virus DNAfragment
capable
oftrans-inducing
agenes. Proc.Natl.Acad.Sci. USA82:5870-5874.
45. Pereira, L., M. H. Wolff, M. Fenwick, and B. Roizman. 1977.
Regulation
ofherpesvirus
synthesis. V.Properties
ofapolypep-tides
specified by
HSV-1 and HSV-2.Virology
77:733-749. 46. Powell,K.L.,and R J.Courtney.
1975.Polypeptides synthesized
in
herpes
simplex
virustype 2-infectedHEp-2
cells.Virology
66: 217-228.47. Preston,C. M. 1979.Control of
herpes
simplex
virustype1mRNAsynthesis
incells infected withwild-type
virusorthetemperature-sensitivemutanttsK. J.Virol. 29:275-284.
48. Preston,V.G.,A.J.
Davison,
H.S.Marsden,M.C.Timbury,J.H.Subak-Sharpe,
and N. M. Wilkie. 1978. Recombinants betweenherpes
simplex
virus types1 and2:analysis
ofgenomestructureand
expression
ofimmediate-early
polypeptides.
J.Virol. 28:499-517.49. Purves,F.C.,W.0.
Ogle,
and B. Roizman.1993.Processing
of theherpes
simplex
virusregulatory
protein
a22mediatedby
theUL13
protein
kinasedeterminestheaccumulation ofasubset ofaandgmRNAsand
proteins
ininfected cells.Proc.Natl.Acad.Sci.USA 90:6701-6705.50. Quinlan, M.,and D.
Knipe.
1985. Stimulationofexpression
ofaherpes
simplex
virusDNAbinding
protein
by
twoviralfunctions. Mol. Cell. Biol. 5:957-963.51. Ralph, W.,and P.A.Schafer.Personalcommunication. 52. Roizman, B.,and A. E. Sears. 1993.
Herpes
simplex
virusesandtheir
replication,
p. 11-69. In B. Roizman, R.J.Whitley,
and C.Lopez
(ed.),
The humanherpesviruses.
Raven Press, New York.53. Romanelli, M. G., P. Mavromara-Nazos, D.
Spector,
and B.Roizman. 1992. Mutational
analysis
of the ICP4binding
sites in the5'transcribednoncoding
domainsof theherpes
simplex
virus 1UL49.5Y2
gene. J.Virol.66:4855-4863.54. Sacks,W.R,C. C. Greene, D. P.Aschman,and P. A.
Schaffer.
1985.Herpes
simplex
virustype1ICP27isanessentialregulatory
protein.
J.Virol.55:796-805.on November 9, 2019 by guest
http://jvi.asm.org/
55. Sacks, W. R., and P. A. Schaffer. 1987. Deletion mutantsinthe gene encoding the herpes simplexvirus type 1 immediate-early protein ICPO exhibit impaired growth in cell culture. J. Virol. 61:829-839.
56. Sandri-Goldin, R., and G. E. Mendoza. 1992. A herpesvirus regulatory proteinappears to actpost-transcriptionally by affecting mRNAprocessing. Genes Dev. 6:848-863.
57. Sekulovich,R.E., K.Leary,and R M.Sandri-Goldin. 1988.The herpes simplex virus type 1 protein ICP27 can act as a trans-repressor ortrans-activator in combination with ICP4 and ICPO. J. Virol. 62:4510-4522.
58. Shepard, A. A., A. N. Imbalzano, and N. A. DeLuca. 1989. Separation of primarystructural componentsconferring autoreg-ulation, transactivation, andDNA-bindingpropertiestothe her-pessimplex virus transcriptionalregulatory protein ICP4.J.Virol. 63:3714-3728.
59. Smith, C. A., P. Bates, R Rivera-Gonzalez, B. Gu, and N. A. DeLuca.1993. ICP4,the majortranscriptional regulatory protein ofherpes simplex virus type 1, forms a tripartite complex with TATA-bindingproteinandTFIIB. J.Virol. 67:4676-4687. 60. Spector, D., F. C. Purves, R. King, and B. Roizman. 1993.
Regulationof geneexpression inanimal viruses, p. 25-42. Plenum, NewYork.
61. Stow, N. D., and E. C. Stow. 1986. Isolation and characterization of a herpes simplex virus type 1 mutant containing a deletion within the gene encoding the immediate-early polypeptide VmwllO. J. Gen.Virol. 67:2571-2585.
62. Su,L., and D. M. Knipe. 1989. Herpes simplexvirus a protein ICP27 can inhibit or augment viral gene transactivation.Virology 170:496-504.
63. Weinheimer,S.P.,and S. L.McKnight. 1987.Transcriptionaland post-transcriptional controlsestablishthe cascade ofherpes sim-plexvirusprotein synthesis.J.Mol.Biol. 195:819-833.
64. Wilcox, K. W., A.Kohn, E. Sklyanskaya,andB. Roizman. 1980. Herpes simplexvirusphosphoproteins.I.Phosphate cyclesonand off someviralpolypeptidesand can alter theiraffinityfor DNA.J. Virol. 33:167-182.
65. Yao,F. 1991.Virion- andmembrane-associated immediate-early proteins of herpessimplex virustype 1.Ph.D. thesis.The Louisi-anaStateUniversity MedicalCenter, Shreveport.
66. Yao, F., and R J.Courtney. 1989.Amajortranscriptional regu-latory protein (ICP4) ofherpes simplex virustype 1isassociated withpurifiedvirions. J.Virol. 63:3338-3344.
67. Yao, F., and R J. Courtney. 1992.Association of ICPO but not ICP27withpurified virionsofherpes simplex virustype 1.J.Virol. 66:2709-2716.
68. Yeh, L., and P. A. Schaffer. 1993. A novel class of transcripts expressed with late kinetics in the absence of ICP4 spans the junction between the long and short segments of the herpes simplex virustype1 genome. J.Virol. 67:7373-7382.
69. Zhu, Z., W. Cai, and P. A. Schaffer. 1994. Cooperativityamong herpessimplexvirus type 1 immediate-early regulatory proteins: ICP4 and ICP27 affect theintracellular localization of ICPO. J. Virol.68:3027-3040.