JOURNALOFVIROLOGY,Feb.1973,p.232-237
Copyright ©1973 AmericanSocietyforMicrobiology
Vol.11, No.2 Printed inU.S.A.
Effect of Viral
Double-Stranded
RNA
on
Protein
Synthesis
in
Intact Cells
B. CORDELL-STEWART AND MILTON W. TAYLOR
Departmentof Microbiology, Indiana University, Bloomington, Indiana47401
Received for publication 17August 1972
The addition ofpurified, noninfectious, double-stranded RNA ofbovine en-terovirus, a picornavirus, to intact cells in culture results in a rapid cessation ofcellularpolypeptide synthesis. Thisinhibition isspecificforhostcell protein
synthesis since the translation of picomavirus-specific proteins isnot affected
bythe double-stranded viralRNA.
It has been recently reported that
heterolo-gous double-strandedRNAshave aninhibitory
effect on in vitroproteinsynthesis(4, 8, 9). We
report a similar inhibition ofcellular
polypep-tide synthesis when purified, noninfectious,
double-stranded RNA isolated from cells
in-fected with a picornavirus, bovine enterovirus
-1 (BEV), is added to intact cells in
suspen-sionculture. This inhibition is specific for
cell-directed protein synthesis asdemonstrated by
a lack of inhibition when the viral
double-stranded RNA is added to either BEV or
Mengo virus-infected cells at the time of
maxi-mum viral-specific protein synthesis.
MATERIALS AND METHODS
Cell, virus, and viral infections. Propagation of mouseL cells andEhrlich ascitestumor(EAT) cells, virus isolation, and infections have previously been described (5, 14, 15).
Isolation and purification of viral double-stranded RNA. After 7 to 7.5hr ofinfection, cells
were harvested by centrifugation, resuspended in
phosphate-buffered saline, and homogenized gently
in a glass Douncehomogenizer. Cell debris was
re-movedby centrifugation at 700 x g for20 min,and RNAwas extracted from the supernatant fluid at 37 C by the method of Penman(11). Theviral double-stranded RNAwas isolated by differential salt pre-cipitation (2), treatedwithribonuclease AandT,at 15
gg/ml
and 2Ag/ml,
respectively, deoxyribonu-clease at 20 ug/ml, and Pronase at 1.0 mg/ml.Theenzymatic treatment procedure is as described by Stern and Friedman (13). The double-stranded RNA was purified by centrifugation through sucrose gra-dients as described by Bishop et al. (3) and then dialyzed against0.15MNaCl with0.001 M ethylene-diaminetetraacetic acid(EDTA)for12hr at 4 C. Radi-oactively labeled double-stranded RNA was obtained by adding3.0gCiof[5-_3H uridine perml tothe viral incubation medium1hrpostinfection in the presence
of5
Ag
ofactinomycin D per ml.Infectivity assay of viral double-stranded RNA.Titers of infectious RNA were determined by
aplaque assay in which confluentL-cellmonolayers onFalcon plastic dishes (60 mm)wereused. Double-stranded RNA, appropriately diluted in 1 M NaCl containing 100 gg of diethylaminoethyl
(DEAE)-dextran per ml,wasaddedin avolumeof 0.05mlto
the monolayer which had previously been washed with saline and then0.6MNaCl. Aftera30-min incu-bationperiodat 37 C, themonolayerswereoverlayed
with a 1:1mixture of1.2%Noble agar and 2x Earle minimal essential media. The disheswereincubated
for 48hr, after which the agaroverlaywasremoved andthe cell sheetwasstained with0.8%crystal violet in20%ethanolforthe enumeration ofplaques.
Determination of amino acidincorporation into
polypeptides. Cellsinsuspensionwere incubatedin a 37 C shaker bath at various concentrations in
Hanks minimal essential medium and with various concentrations of radioactive protein precursors as describedfor eachexperimentinthelegends (Fig. 1, 2, 4, 5, 6). Samples werewithdrawn, and incorpora-tionwashaltedby freezingin adryCO,-ethanolbath (-70 C). Samples werethawed, bovine serum albu-minwasaddedascarrier, andsamplesweremadeto 10% trichloroacetic acid with 20% trichloroacetic acid. Next, the samples were boiledat 100C for 5 min and the resulting precipitate was washed three times with5% trichloroaceticacid, driedat37 C for 15 hr, and solubilized in 0.5 ml ofNuclear-Chicago
solvent(New England Nuclear, Boston, Mass.)for 2
hrat37 C.Samples werecounted in aBeckman liq-uid scintillation counter in a toluene-2,5-diphenyl-oxazole (NewEngland Nuclear, Boston, Mass.)
scin-tillation cocktail. Using the wide-window, fixed ISO-SET, 14C was counted at90% efficiency, while
3H was counted at 35% efficiency.
Radioactive materials. Radioactive materials included:
[5-'Hluridine,
20 to 30 Ci/mmole;["4C]-leucine, 58.2 Ci/mmole;
["4C]phenylalanine,
383Ci/mmole(all threefromNewEngland Nuclear); and
[I4C]protein
hydrolysate, 54 Ci/mA (Amersham/Searle, Arlington Heights, Ill.).
RESULTS
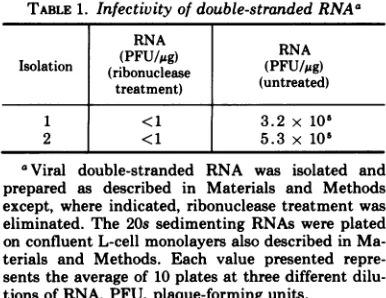
Infectivity of viral double-stranded
RNA. The viral double-stranded RNA used in
232
on November 10, 2019 by guest
http://jvi.asm.org/
the experimentsto be describedwastested for infectivity. From the same isolation, one half
ofthe total RNA initiallyisolated was prepared
as described in Materials and Methods, the
other halfwas treated similarly excepting the
ribonuclease treatment. Both 20s sedimenting
RNAs were then tested for infectivity by the
plaque assay method outlined in Materials
and Methods. It can be seen in Table 1 that
treatment of the double-stranded viral RNA
with ribonuclease drastically reduces (5 log
units) the plaque-forming ability ofthe RNA.
Therefore, the followingresults are not a
con-sequence of the replicative capacity of this
viral RNA, since ribonuclease-treated RNA
was used in subsequent experiments.
Effect of viral double-stranded RNA
on
protein
synthesis.
The effect of viraldouble-stranded RNA on cellular protein
syn-thesisin intactEATcellswasexamined. A
sus-pension ofcells in culture, actively
incorpora-ting aminoacids, wassupplemented with
puri-fied BEV double-stranded RNA. As
demon-strated inFig 1, the addition ofthe viral RNA
resulted in a cessation in the incorporation of
amino acids. This probably reflects an overall
inhibition in cellular translation rather than a
reduction in the rate ofsynthesis. The
inhibi-tion can be seen to occur within 10 min after
the addition ofthe double-stranded RNA.
By
40 min aftertreatment, therate of aminoacid
incorporation has decreased by 70% as
com-pared to the control culture. Also, the
level
ofinhibition
induced by the double-strandedRNA is identical to that
eventually
obtainedwhen cells are infected with complete BEV
virus (Fig. 2). The
following
experiment wascarriedoutto eliminate the
possibility
that theinhibition by the viral double-stranded RNA
was aconsequence of analterationincell
mem-brane permeability, which would have resulted
in suppressed incorporation of protein
precur-TABLE 1. Infectivity ofdouble-stranded RNAa
RNARN
Isoltion
(PFU/,ug)
RNA/ug
Isolation (ribonuclease (uReAte) treatment) (untreated)
<1 3.2 x 105
2 <1 5.3 x 10'
aViral double-stranded RNA was isolated and
prepared as described in Materials and Methods except, whereindicated, ribonucleasetreatment was eliminated. The 20ssedimenting RNAswere plated
onconfluent L-cellmonolayersalso describedin Ma-terials and Methods. Each value presented
repre-sentstheaverageof 10platesatthreedifferent
dilu-tions ofRNA. PFU,plaque-formingunits.
!2~0
10
z<
~~~~~~~0/
/OO /o i O o
10 20 30 40 S0 *0
FIG.1. Effect of viraldouble-stranded RNA on cel-lularprotein synthesis. "4C-protein hydrolysate was added to a culture of EAT cells (2.5 x 0l cells per ml) to a final concentration of 2.0
;Ci/ml.
7he cul-ture wasimmediately divided. After 10 min of incu-bation at 37 C with slight agitation, BEV double-strandedRNA, in a smallvolume of2x SSC(SSC=0.15M NaCl, 0.015 M sodium citrate, pH 7.0), was added to one culture to a concentration of 20ug/ml,
and anequal volumeof2 x SSCwasadded to the control culture. Samples were prepared for analysis ofamino acidincorporationasdescribed in Materials and Methods. Symbols: 0, untreated control cells; 0, double-stranded RNA-treated cells. Arrow indi-catestimeofRNAaddition.
sors. Viral double-stranded RNA-treated cells
and untreated cells were monitored for their
respective ability to incorporate amino acids.
The rate and extent of uptake ofradioactive
amino acids in Fig. 3 indicatesnodifference in
membrane
permeability
between cells culturedwithorwithout the viral RNA.
To test the specificityofthis
inhibition,
theeffect ofBEV double-stranded RNA on
viral-specific protein synthesis was investigated. At
145 min postinfection with BEV virus, a time of linear viral-specific protein synthesis (Fig.
4), BEV double-stranded RNA was added to
the culture. Noinhibition in the accumulation
ofviral-specificproteinsbythe viral RNA can
be detected, even 60 min aftertreatment(Fig.
5). To show that this was viral-specific protein
synthesis, the culture wasassayedat6 hr after
virus infection for virus production. A normal
burst size for BEV-infected EAT cells (25
plaque-formingunits per cell) was obtained by
plaqueassay determination.
To further test the specificity of the
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.493.253.444.66.270.2] [image:2.493.48.241.510.659.2]CORDELL-STEWARTAND TAYLOR
wasobtained for
virally
infected cells (datanotD
shown).
. Nature of the cell-associated RNA. Cells
were incubated with labeled double-stranded
RNA for 30minatwhich timetheywereplaced
in RNA-free medium and incubated further for various intervals. The cell-associated RNA
D * was then analyzed as to its double- or
single-stranded nature on the basis of ribonuclease resistance. After a total of 30 min of incuba-tion, 98.8% ofthe RNA was foundto have
re-tained its double-strandedness. However, at
60 min incubation, only 65% was resistant to A
.,,
ribonuclease,
andby
120 minonly
25.5% wasdouble-stranded RNA (Table 2). Although
the double-stranded RNA appears to become
QC5 i.'o
2.
. single-stranded (ribonuclease sensitive) witht
0.5
1.0
1.S
2.0
time, it would seem from Fig. 2 that the RNAHOURS
is
not capable of translation, precluding theFIG. 2. Inhibition of cellular protein synthesis by viral double-stranded RNA. BEV double-stranded RNAataconcentration of25gg/mlin 2x SSC (see Fig. 1 legend) was added toasuspension culture of EATcells(107cellsperml). After the addition ofthe RNA, samples of 101 total cells were removed and pulse labeled for10minutes with5,uCiof l 4C-phenyl-alanine. The pulsewashalted byfreezing the sample at-70Cinadry ice-ethanol bath. Sampleswerelater
assayed forhot trichloroacetic acid-precipitable ma-terial. The data are expressed aspercentages of an
identically labeled control towhichanequal volume of2 x SSCwasadded.Also shown isthe inhibitory effect by BEVvirus in a culturehandled similarly. (Infection by the virus is described in reference 5.) Each point was determined from theaverage of duplicate samples. The arrow indicates the initial
time of treatment with the double-stranded RNA
and virus. Symbols: A, double-stranded RNA
treated; A, BEVinfected.
tor,the effect of BEVdouble-stranded RNAon the protein synthesis of another picornavirus, Mengo virus, wasexamined. Adding BEV RNA at a time of increasing accumulation of viral proteins in thereplication cycle of Mengo virus (15),noalterationintranslationwasnoted(Fig. 6).
Extent of incorporation of viral double-stranded RNA into whole cells. Using radi-oactively labeled BEV double-stranded RNA, the rate and amount of RNA associated with whole cells was determined. Cells were
incu-bated in the presence of the double-stranded RNAat20,ug/ml. Thoroughlywashed cell
sam-pleswere then assayed for uptake of the RNA. A linear increase in cell-associated label could be observed over a2-hr incubationperiod (Fig. 7). At this time, this represented approxi-mately 1.8 pg per cell. Similar incorporation
possibility that a newly
synthesized
viral
pro-tein is the mediatorofthe inhibition observed.
DISCUSSION
The data presented here further suggest
that the inhibition of cellular protein synthesis
\ 3
70
, 2
0
o 1. v
10 20
MINUTES
FIG. 3. Effect of viral double-stranded RNA on
cellpermeability. EATcellswereincubatedat37 C in phosphate-buffered saline at a concentration of
4 x 106cellspermlfor1hr. Thecellswere then pel-leted andresuspended toa concentration of2 x 106
cellsper mlinphosphate-buffered saline containing
2
gCi
of "4C-protein hydrolysateper ml plus25 Ag ofviraldouble-strandedRNAperml in2x SSC(seeFig. 1 legend) oranequivalent volumeof2 x SSC.
Samples were extractedanddiluted1:100 insaline,
washedthreetimes insaline,andthefinal cell pellet
wassolubilized inNCSat37Covernight. Symbols:
A, untreated control; A, double-stranded RNA
treated. lOC
wso
c
6C
0
z 0
I
4C
20
234 J. VIROL.
4,a
a
A
4,A
AA
I
aA
a A
II
A
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.61.249.69.266.2] [image:3.493.260.454.343.530.2]--1EAT -0-L-CELLS
hoursafter infection
FIG.4. BEV-induced inhibitionofhostprotein syn-thesis. Cells were infected as previously described (5).Atvarious time intervals after infection, samples of2x 101cellswerepulse labeled for 10 min with 10
ACi
oft4C-phenylalanine. Sampleswereanalyzed for hot trichloroacetic acid-precipitable material. Values are reportedaspercentages ofanuninfected control culturepulse labeled in thesamefashion.by picornavirus double-stranded RNA seen
in vitro reflects the inhibition occurring in
in-fected cells. This is demonstrated by an
inhi-bition oftranslation in intact cellswhen
nonin-fectious viral double-stranded RNA is added
tothe culture medium. The inhibitionisrapid,
observable as early as 10 min after treatment
with the viral RNA. Allowing 5 min for the in-corporation of the double-stranded RNA (2),
this inhibition may actually be commencing
within 5 min ofaddition ofthe inhibitor. The
inhibition inthe rate of amino acid
incorpora-tion was morethan 70%, 40 min after thewhole
cellscome in contact withthedouble-stranded
RNA. Thisinhibitionintranslationby the viral
double-stranded RNA is not a consequence of
decreased membrane permeability since the cells show no alteration in theuptake of amino acids after treatment.
Reisolation ofthe viral double-stranded RNA fromtreated cells at a time of maximum inhibi-tion indicated that the RNA had retained nearly 99% of itsdouble-stranded nature deter-mined by its resistance to ribonuclease. This suggests that single-stranded viral RNA by
it-self is not the inhibitor. That viral
double-stranded RNA is responsible for inhibition of
host cell translation in virally infected cells is
also evidenced, though only indirectly, by the
necessity of a functional, single-stranded ge-nome; ultraviolet-inactivated virus (12) or hy-droxylamine-treated virus (10) has no effect on
cell-specific protein synthesis. Also, Hunt and
Ehrenfeld (9) demonstrated that
single-stranded viral RNA did not inhibit in vitro syn-thesis of hemoglobin at concentrations at which the double-stranded RNA is inhibitory. This inhibition of translationby picornavirus
double-stranded RNA appears specific. There
is nodetectable inhibition of protein synthesis inpicornavirus-infected cellsbythe viral inter-mediate. This is seen for both homologous
(BEV) and heterologous (Mengo virus)
poly-peptide synthesis employing BEV double-stranded RNA. This is consistent since BEV
and Mengovirus can replicate productively in
simultaneously infected cells. Therate of
incor-poration ofthe double-stranded RNA is
identi-cal for both picornavirus-infected and
unin-fected cells. Approximately 0.8 pg per cell can
be foundafter a 1-hrincubation period
reflect-180 190 200 210
.1.UT 5,
"I",r[ Po"t S C.6 V0",I,,, -" T
'-FIG. 5. Effect of BEV double-stranded RNA on protein synthesis in BEV-infected cells. EAT cells
wereinfected with BEVat amultiplicity of infection of50 aspreviously described (5).At 135min
postin-fection, 2.0MCiof "4C-leucineper ml was addedto the culture. The culture was then divided. After10 min of incubation at 37 C, BEV double-stranded
RNA wasaddedto oneculturetoaconcentrationof 20 ug/ml. The control was treated with an equal volumeof RNA suspension buffer. Symbols: 0, un-treated BEV-infected cells; 0, double-stranded RNA-treated BEV-infected cells. Arrow indicates timeofRNAaddition.
.3
i.
la
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.493.48.238.65.311.2] [image:4.493.251.444.339.537.2]CORDELL-STEWART AND TAYLOR
1S
v s
10~~~~~~
6/0
10 20 30 40 SO 60 70
'145 1SS 165t '175 185 195 20S
II-INUI1S POST St"V INICtTION
FIG. 6. Effect of BEV double-stranded RNA on protein synthesisinMengo virus-infected cells. EAT
cellswereinfected with Mengovirusatamultiplicity of infection of80aspreviouslydescribed(13).At 180 min postinfection, 1.0 pCi of "4C-protein hydrolysate permlwasaddedtotheculture and then the culture
was equally divided. The procedure follows thatas described in the legend of Fig. 4. Symbols: 0,
un-treated Mengo virus-infected cells; 0,
double-stranded RNA-treated Mengo virus-treated cells.
TimeofRNA addition markedbyarrow.
ing 20% adsorption. Weigers and Koch report
that, in the absence of polycations,
approxi-mately20% ofpoliovirus double-stranded RNA
is adsorbed to HeLa cells after a 15-min incu-bation period (16). These authors also state
that the degree of double-stranded RNA ad-sorbed is not dependent on RNA
concentra-tion and has only a slight dependence on cell concentration. In oursystem, a 15-min incuba-tion results in about 7 to 9% RNA adsorbed,
and this amount increases as the incubation
period is lengthened. It should be noted that
the poliovirus double-stranded RNA used by
Weigers and Koch was not treated with ribo-nuclease which may increase its adsorption to cells. Whether the BEV double-stranded RNA
taken up by cells is intracellular oradheres to the cell surface iscurrently being investigated.
Preliminary evidence suggests it is
intracellu-lar.
The mechanism of inhibition of host cell translation by the viral intermediate is un-known. Theinhibition observed mayrepresent
an interaction between the double-stranded RNA and ribosomes, transfer RNA (tRNA), essential factors, or messenger RNA (mRNA).
Ribosomes and tRNA canbe eliminated since
such interactions should produce nonspecific
inhibition. Thus the possible siteofspecificity
is the mRNA, the essential factor(s), or both.
From in vitro studies (1, 9), there is indirect evidence that the initiation processof thehost
is inhibited. It is also suggested by Ehrenfeld
and Hunt (9) thatan initiation factorrequired for polypeptide synthesis is depleted during
incubationwiththe viral double-stranded RNA
and that its formation or regeneration is
pre-2.0 10
0 20 40 O 100
FIG. 7. Rate and extent of incorporation ofBEV double-stranded RNA into whole cells. EATcells, at a concentration of5 x 106 cellsperml, were incu-batedin thepresenceof20pgof 3kH-double-stranded
RNAperml(510countsperminper,g).Cellsamples were removed and washed three times with saline. Samples were thenanalyzed for cold trichloroacetic acid-insoluble material by precipitation onto
mem-branefilters (Millipore Corp.).
TABLE2. Secondary structure of cell-associatedRNAa Total Ribonuclease- % Ribo-time of Untreated treated nuclease-resist-incubation (counts/min)
(min) (counts/mm) ant RNA
30 987 975 98.9
60 1026 667 65.0
120 1002 256 25.5
aEhrlichascites, at aconcentration of 4 x 106 cells per mlin Hanks minimal essential medium were treated with 3H-double-stranded viral RNA at a concentration of 20
ug/ml. The suspension culture was kept at 37 C under slight agitationfor 30min,afterwhich thecells were washed with 37 C minimal essential medium, resuspended to the samecell concentrationinRNA-free medium, and incubated further. Samples of8 x 101total cells were removed, pel-leted, resuspendedin 2 x SSC, and then phenol extracted at 37C. Theresulting aqueous phase was divided equally; one-halfwas treatedwith 10pgofpancreatic ribonuclease per ml and2 pgofribonucleaseT, perml at 37 C for 20 min, the other half was incubated similarly without ribo-nuclease. Thesampleswerethen analyzedforcold trichloro-aceticacid-insoluble material.
236
J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.54.243.57.275.2] [image:5.493.259.446.197.343.2] [image:5.493.261.447.455.531.2]ventedby this RNA. Further evidence from in vitrostudieswithahemoglobinprotein
synthe-sizing system suggest that, in the absence of
mRNA,complexes areformed between the
ini-atorformylmethionyl tRNA (met-tRNAF) and
40sribosomal subunits,areversalofthe normal
function ofcodon recognition. In the presence
of poliovirus double-stranded RNA this
com-plex formation is prevented (6). This
met-t-RNAF subunit complex seems to require an
initiation factor(s) (7). Ascione and Vande
Woude (1) report the existence of a
virus-spe-cific initiation factor isolated only from cells infected with foot and mouth disease virus,
another picornavirus. This factor stimulates
the binding of N-acetyl-aminoacyl-tRNA to
viralmRNA in a foot-and-mouth disease virus
RNA-dependent system in vitro. Host
initia-tion factors isolated from uninfected cells do
not permit translation of the viral message.
This factor may be insensitive to the
transla-tional inhibitory action of the viral
double-stranded RNA, whereas a corresponding
cellu-larfactor may not.
However, the workof
Speyer
etal. (4) wouldsuggest that the inhibition seen by
double-stranded RNA is specific for the message. In
an E. coliin vitroprotein synthesizing system
directed by f2 and
T4
mRNAs, thesynthetic
double-stranded RNA,
polyinosinic-polycyti-dylic acid inhibitsonly theformersystem.
Also,
it was shown that the conformation of the f2
RNA
plays
aroleinthe inhibition.A well defined in vitro
study
employing
cellular,
viral,
andsynthetic
messages andcell and viral initiation factors may
help
toelucidate the mechanism ofinhibition
by
picor-navirus double-stranded RNAs.
ACKNOWLEDGMENTS
The authors wish to acknowledge the finetechnical as-sistance of Elodee A. Hopper andMengovirus donatedby
Suzanne Prather.Thiswork wassupported bygrantsfrom thePublic HealthService,no. CA10417fromthe National Cancer Institute, and from the DamonRunyon Memorial fund(DRG1018). B. C.S.is apredoctoraltraineesupported byPublic Health ServicegeneticstraininggrantGM12from theNationalInstituteforGeneral Medical Sciences.
LITERATURE CITED
1. Ascione, R., and G. F. Vande Woude. 1971. Ribosomal factors effecting the stimulation of cell-free protein synthesis in the presence of foot-and-mouth disease virus ribonucleic acid. Biochem. Biophys. Res. Com-mun.45:14-21.
2. Bishop, J.M., and G. Koch. 1967. Purification and char-acterization of poliovirus-induced infectious double-strandedRNA. J. Biol. Chem. 242:1736-1743. 3. Bishop, J. M., D. F. Summers, and L. Levintow. 1965.
Characterization of ribonuclease-resistant RNA from poliovirus-infected HeLa cells. Biochemistry 54:1273-1281.
4. Chao, J., L.Chao,and J.Speyer. 1971.The inhibitionof f2 RNA directed E. coli protein synthesis by Poly rI rC in vitro. Biochem. Biophys. Res. Commun. 45: 1096-1102.
5. Cordell-Stewart, B., and M. W. Taylor. 1971. Effect of double-stranded viral RNA on mammalian cells in cul-ture. Proc. Nat. Acad. Sci. U.S.A. 68:1326-1330. 6. Darnbrough, C., T. Hunt, and R. J. Jackson. 1972. A
complex betweenmet-t-RNAFand native 40s subunits in reticulocyte lysates and its disappearance during incubation with double-stranded RNA. Biochem. Bio-phys. Res. Commun. 48:1556-1564.
7. Editorial. 1972. Protein synthesis at Cold Spring Harbor. Nature N. Biol. 239:223-224.
8. Ehrenfeld, E., and T. Hunt. 1971. Double-stranded poli-ovirusRNA inhibits initiation of protein synthesis by reticulocyte lysates. Proc. Nat. Acad. Sci. U.S.A. 68: 1075-1078.
9. Hunt, T., and E. Ehrenfeld. 1971. Cytoplasm from polio-virus-infected HeLa cells inhibits cell-free haemoglo-binsynthesis. Nature (London) 230:91-94.
10. Koschel, K., H. Tauber, and E. Wecker. 1971. The inhi-bitionofcellular protein synthesis in HeLa cells after infection with active and inactivepoliovirus. Z. Natpr-forsch 26:798-803.
11. Penman, S. 1966. RNA metabolism in the HeLa cell nu-cleus. J. Mol. Biol. 17:117-130.
12. Penman, S., and D.Summers. 1965. Effects on host cell metabolism following synchronous infection with polio-virus.Virology 27:614-620.
13. Stern,R., and R. M. Friedman. 1971. Ribonucleic acid synthesis in animal cells in the presence of actinomycin D.Biochemistry. 10:3635-2645.
14. Taylor, M. W., B. Cordell, M. Souhrada, and S. Prather. 1971.Viruses as an aid to cancertherapy: regression of solid and ascites tumors in rodents after treatment with bovineenterovirus. Proc. Nat.Acad. Sci. U.S.A.
68:836-840.
15. Wall, R., and M.W.Taylor. 1970. Mengovirus RNA syn-thesis inproductive and restrictive cell lines. Virology 42:78-85.
16. Weigers, K. J., and G. Koch. 1972. Interaction of polio-virus-induced double-stranded RNA with HeLa cells. Arch. Biochem.Biophys. 148:89-96.