Copyrighti 1977 AmericanSociety forMicrobiology Printedin U.S.A.
Isolation
of the Viral DNA
Replication
Complex
from
Adeno-Associated Virus
Type
1-Infected Cells
HIROSHIHANDAtANDHIROTOSHIMOJO*
InstituteofMedicalScience, University of Tokyo,4-6-1,Shirokanedai, Minato-ku, Tokyo 108,Japan
Received for publication 6 June 1977
The
replication
complex
active in
adeno-associated virus type
1(AAV-1)
DNA
synthesis
in vitro wassolubilized,
with anonionic
detergent,
fromthenuclei ofhuman
embryonickidney
cells coinfectedwith AAV-1andanearly
temperative-sensitive mutant
(ts125)
of human adenovirustype
5 at thenonpermissive
temperature
(40.50C).
Thecomplex
sedimented with a mean size of 23S andcontainedparental AAV-1 DNA. Most ofthe DNA
synthesized
withtheAAV-1DNA
replication complex in vitrowasAAV-1DNA,
asrevealedby DNA-DNAhybridization
andsedimentation
in aneutral
sucrosegradient. However, it
sedi-mentedinan alkaline sucrose
gradient
asmolecules smaller thanAAV-1 DNA(14.4S).
The AAV-1 DNAreplication
complex
was notformed in cells infectedwith AAV-1alone.
Adeno-associated
virus(AAV)
is a defectiveparvovirus
that iscapable
ofmultiplying
only
in cells coinfected with
adenovirus
(1, 8, 15, 23).
The
analysis
of the AAVDNAhasbeen difficultdue to the simultaneous
replication
ofhelper
adenovirus
(17).
Itwasreported
that atemper-ature-sensitive
(ts)
mutantofhuman adenovirustype
31(H31) (10,
13),
aswellasmutantsof
H5and H12
(7, 24)
and avianadenovirus
(9),
defec-tive in viralDNA
synthesis
atthenonpermissivetemperature,
assist thegrowth
ofAAV.
Only
AAV DNA
replicates without the
replication
of
adenovirus DNA under these
conditions
(6, 13).
We
reported
that nucleiisolated from
cells
coin-fected with AAV
type
1(AAV-1)
and H31tsA13
atthe
nonpermissive
temperature
(400C)
wereactive in AAV-1 DNA
synthesis
invitro(5).
Theavailability
of asubcellular
systemcapable
ofsynthesizing
AAVDNA invitrowould facilitate theanalysis
ofAAV DNA
replication.
The
pres-entcommunication
describes the
isolation of the
AAV-1 DNA
replication
complex
from nuclei of
cells coinfected with AAV-1 and H5ts125 at
40°C,
aswell
astheanalysis
of
DNAsynthesized
invitro.
It also shows that the
replication
com--plex
was notformed in cells infected with
AAV-1alone.
MATERIALS AND
METHODS
Cells,
viruses,
andinfection. Secondary culturesof humanembryonic kidney (HEK) cells and mono-layer cultures of KBcellswereused. The growth of AAV-1wasdescribedpreviously (6). A
temperature-tPresent address:Laboratoiy of ExperimentalPathology,
National Institute of Arthritis, Metabolism, and Digestive Diseases,Bethesda, MD 20014.
sensitive (ts) mutant ofH5,H5ts125,waskindly pro-vided by H. S. Ginsberg (4). H5ts125 was grown in KB cells and plaque-assayed in HEK cells at 330C
(permissive temperature). Confluent monolayers of HEK cells in 32-ounce (ca.960-ml) bottles (2 x 107 cells per bottle) were coinfected with AAV-1 and H5ts125 at 15fluorescent infectious units (2) and 30 PFU percell,respectively.Afteradsorptionfor2h at 370C, the cultures were incubated in maintenance medium (Eagle minimum essential medium [MEM]
with 1% fetal calfserum) at40.5°C (nonpermissive
temperature).
Preparation ofthe AAV-1 DNA replication
complex. HEK cells were harvested at 18 h after
coinfection with AAV-1 and H5ts125, washed with phosphate-buffered saline (PBS), and suspended in
reticulocyte standard buffer (10 mM NaCl, 10 mM
Tris-hydrochloride, pH7.4, 1.5 mMMgCl2)at2x107
cells per ml inanice bath for30min. Theswollen cells were disrupted with five strokes in a tightly
fitting Douncehomogenizer,andthe nucleiwere col-lected by centrifugation at 2,500 rpm for 5min at
40C.Thenuclearpelletwaswashed withreticulocyte
standard buffer andsuspendedat 2 x 107nuclei per ml in TK buffer (50 mMTris-hydrochloride, pH 7.9,
25 mM KCl) containing 0.1 mM EDTA and 2 mM dithiothreitol (DTT). The nuclear suspension was lysed by addition of 0.5% dodecyl poly (P = 11.2)
oxyethylene ether(KAO-AtlasCo. Ltd., Tokyo) (27)
at0°Cfor30min. Then,thelysate wascentrifuged
at 4°Cin anSW27rotor at 25,000 rpm for 30 min. Thesupernatantwasdialyzed against TK buffer
con-taining0.1mMEDTA,20%glycerol,and2mM DTT
at 4°C for 3 h and used immediately or stored at -80°C. A protease inhibitor, phenylmethylsulfonyl
fluoride(Sigma),wasadded tosolutions at a concen-tration of1 mMforpreparationofthecomplex.
Assayforendogenous DNAsynthesisinvitro.
The reaction mixture (100
pi)
contained 50 mMTris-hydrochloride, pH 7.9, 2.5 mM MgC12, 2 mM DTT,
444
on November 10, 2019 by guest
http://jvi.asm.org/
100FM each of dATP, dCTP, and dGTP, 20
.uCi
of[3H]TTP
(46Ci/mmol), andapreparation of the DNAreplicationcomplex containing about 35
gAg
ofprotein.After incubation at370Cfor various times, the incor-poration of[3H]TTP into anacid-insoluble fraction
wasmeasuredasdescribed previously (5). The protein
content was determined by the method of Lowry et aL (12).
Partial purification of the replication
complex
by sucrose gradient centrifugation. The
dialyzed
soluble complex was layered onto a linear 15 to 30% (wt/vol) sucrose gradient in TK buffer containing 0.1 mMEDTA, 10% glycerol, and 2 mM DTT. The gra-dient wascentrifugedin anSW41 rotor at 30,000 rpm
for20hat
40C.
Aportion of each fraction was assayed for endogenous DNA polymerase activity. The re-mainder of each fraction was frozen rapidly and storedat-800C.
Analysisof DNA synthesized in vitro. After the
reaction, the mixture was treated with Pronase (1 mg/ml, self-digested at 370C for 1 h) and 25 mM EDTA at 370Cfor 30min,followed by further
incu-bation for 60 min after addition of 0.1% Sarkosyl.
Then,DNAwasextractedwithphenol.The aqueous
phase containing newly synthesized DNA was
ana-lyzedbyaneutralsucrosegradient(5to20%[wt/vol]
in10mMTris-hydrochloride, pH7.5, 0.1 MNaCI,10
mMEDTA)at30,000 rpm for14hat40CinanSW41 rotor or by an alkaline sucrose gradient (5 to 20% [wt/vol] in0.5MNaCl,10mMEDTA,0.3NNaOH) at40,000 rpm for16hat40CinanSW41rotor.After
centrifugation,thegradientwasfractionated,and the
trichloroacetic acid-insoluble radioactivity in each
fractiontrappedon amembranefilter(Millipore)was
counted.DNA-DNAhybridizationwith AAV-1DNA,
H5DNA, and cellular DNAwas carried out as
de-scribed
previously
(22).RESULTS
Conditions
for
solubilization
of the
rep-lication
complex.
The
rateof DNA synthesis
in HEK cells
coinfected with AAV-1and
H5ts125
began
to increaseat 6 hpostinfection
(p.i.) and
reached its maximumat16 hp.i.
(Fig.
1). Nuclei
were isolated from cells at 18 hp.i.
and
solubilized
with various kindsof
detergents.
Only
alowlevel ofendogenous
DNApolymerase
activity
wasfound in thesupernatant,
whenthe
nuclei
weresolubilizedby Nonidet P-40, Triton
X-100,
Brij
58,
orSarkosyl.
The
highest
levelof
activity
wasfound when the nucleiweresolubi-lized
by dodecyl
polyoxyethylene
ether.Theac-tivity
of the
solubilizedcomplex
was alsode-pendent
on the concentration ofNaCl
during
solubilization.
Thecomplex
solubilized atvar-ious
concentrations
of NaCl wasdialyzed and
assayed
for the
endogenous
DNA-synthesizing
activity
in vitro(Fig. 2).
Theresult showedthat 0.5MNaCl
wasoptimal
for solubilization ofthe
complex
with thedetergent.
The
extract similarlyprepared from the nuclei
of
HEKcells infected
with H5ts125alone
x
4 8 12 14 16 18 20 22
Hoursafter coinfection with AAVI and H5ts 125
FIG. 1. RateofDNAsynthesisinHEK cells after
coinfectionwith AAV-Iand
HMts125.
Confluentmon-olayersofHEKcells in 2-ounce(ca.
60-mI)
bottles(2x
log
cells per bottle) were coinfected withAAV-1andH5ts125 at 15
fluorescent
infectious
units and30 PFU per cell,
respectively,
and incubated at40.50C.
At intervals of2 h, the cells were labeled withf3H]thymidine
(2auCi/ml)
for30min, and theradioactivity
incorporated
into the acid-insolublefractionwascounted.
0
x E
0
0)
0
I-0.
c
(I
0 0.1 0.2 0.3 0.5 0.7 1.0 Concentration ofNaCI(mole)
FIG. 2. Effectof
NaCl
concentrationonthesolu-bilization ofthe
complex
with thedetergent.
Thenuclei isolated
from
the HEKcells
coinfected
withAAV-1 and
H5ts125
werelysed
with 0.5%dodecyl
polyoxyethylene
ethercontaining
the indicatedcon-centrationsof NaCl.The
lysate
wascentrifuged.
Thesupernatantwasdialyzedand
assayed
for
DNApo-lymerase
invitro. Theamountof
PHJTTP
incorpo-rated into anacid-insolublefraction
wasmeasured(0).
The extractpreparedfrom
the nucleiof
HEK cells infectedwithH5ts125 alonewassimilarly
as-sayed
(0).
showed
noorverylittle
DNA-synthesizing
ac-tivity
(Fig.
2).
Conditions for DNA
synthesis
in
vitro
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.500.255.449.57.239.2] [image:2.500.275.429.332.516.2]446
with the
replication complex. The
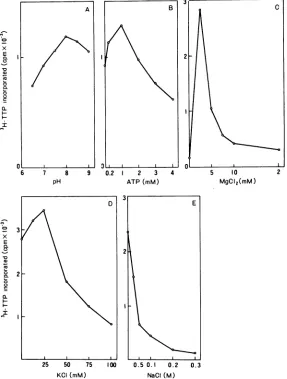
pH
opti-mum for endogenous DNA
polymerase activity
was
broad, ranging from pH
7.5 to8.5(Fig. 3A).
Although ATP and
KCI
were
notessential for
the
activity, the addition of
1mM ATP
and
25mM
KCI
gave maximal stimulation
(Fig.
3B and
D).
Mg2"
was
essential for the
activity (Fig. 3C),
and the
optimal
Mg2e
concentration
was2.5
mM. The
high NaCl concentration (0.5 M)
showed the
inhibitory
effect on the activity (Fig.
3E).
All four
deoxyribonucleoside
triphosphates
were
required for maximal
activity (Table
1).
An
appreciable activity detected in the absence
of one to three
deoxyribonucleoside
triphos-phates may be due
tothe
remains of these
x
E
C)
0
0~
Q.
I
A
01 l
6 7 8 9
pH
substrates
inthe
extract(3, 21, 26). RNase
in-hibited the reaction
slightly. Actinomycin
D
and
sodium
pyrophosphate inhibited the polymerase
activity
markedly.
This
observation suggested
the involvement of RNA
synthesis in DNA
syn-thesis. The lack
of inhibition
by
a-amanitin
in-dicated that RNA
polymerase
II
(11) was
notinvolved in DNA
synthesis
in this system.
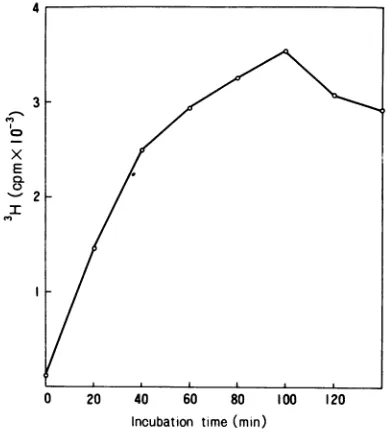
Under
optimal conditions,
DNA
synthesis
increased
linearly for 40 min. Maximum synthesis was
obtained by incubation
at370C
for 100 min
(Fig.
4).
Partial purification of
the
replication
complex by
sucrosegradient
centrifuga-tion.
The
soluble
replication complex
was
par-tially purified
by sedimentation in a neutral
2 3 4 5 10
ATP(mM) MgCI2(mM)
3
E
o 2
0.
25 50 75 100 0.5 0.1 0.2 0.3
KCI(mM) NaCI(M)
FIG. 3. Conditionsfor DNApolymerasein vitro. The crude DNA replicationcomplex was solubilized and assayed for DNApolymeraseunder thefollowingconditions: (A) at various pHvalues; (B,C, D and E) at various concentrationsofATP,
MgC42,
KCI, andNaCi,respectively.on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.500.114.401.247.626.2]TABLE 1. Requirements for DNAsynthesiswith the viral DNA replicationcomplex
Reaction mixture Percentage of max-imalactivity
Complete
... 100Deletions. -DTT ... 102
-dATP ... 73
-dCTP ... 31
-dGTP ... 38
-dATP, dCTP,dGTP ... .... 28
Additions. RNase (100ug/ml) ... ... 25
RNase (4
jig/ml)
... ... 41Actinomycin D (5
jg/ml)
... 19a-Amanitin (10
pg/ml)
... .... 99a-Amanitin (0.5
pg/ml)
... 103a-Amanitin (0.01
jig/ml)
... 98Sodium pyrophosphate (10 mM) . 6
aDNA-synthesizing
activity of the complex was measured under theoptimal conditionsdescribed in the text with thedeletionsoradditionslisted above.Theactivitywasexpressed by incorporationof
[3H]-TTP into the acid-insoluble fraction relative to that
inthecompletesystem.
4
3
0
E
0
'-' I
0 20 40 60 80 100 120
[image:4.500.46.241.79.258.2]Incubation time(min)
FIG. 4. TimecourseofDNA synthesisin vitro. The
crude DNAreplication complexwassolubilizedand
assayed forDNApolymerase.Atthe timeindicated, the acid-insolubleradioactivitywasmeasured.
sucrose gradient. The complex, detected by
DNA-synthesizing activity, sedimentedas a
sin-gle broad peak withameansize of 23S(Fig. 5).
Theapical three fractions (fractions 12, 13, and
14) were pooledas the partially purified
repli-cation complex. The replication complex
ex-tracted from cells coinfected and labeled with
3
co,
X 2
E
0
I-5 10 15 20
Fraction number
FIG. 5. Sedimentation of the soluble complex by sucrosegradients. The crude DNAreplication
com-plex wasextracted, dialyzed, andcentrifuged in a
neutralsucrosegradient.Eachfractionwasassayed
forDNA-synthesizing activityasdescribed
(0).
Thecoinfectedcells werelabeledat18h p.i. with
R3H]-thymidine (2 aCi/ml) for30minand treated in the
same manner. The nuclear extractwas centrifuged
similarly, and the acid-insoluble radioactivity in
eachfractionwascounted
(0).
Simian virus40DNAcomponents I(20S)and II(16S)wereusedasmarkers.
[3H]thymidine
sedimented similarly
(Fig. 5).
The
results indicate that the
complex
of
the
same
size is active both
invivo
and in vitro.
Nature
of
DNA
sequences
synthesized
in
vitro. The results of DNA-DNA
hybridization
revealed that the DNA
synthesized
in vitro with
the
replication
complex
consisted
mainly
of
AAV-1 DNA
(Table
2).
Very
little,
if
any,H5
DNA
orcellular DNA
wassynthesized.
The
results show that the
complex
is the AAV-1
DNA
replication complex.
Sucrose
density gradient
analysis
of
DNA
synthesized
in
vitro.
Most of the in
vitro-synthesized
DNA
sedimented
atthe
posi-tion
coinciding
with the
marker
AAV-1DNA
(14.48)
when
analyzed by
sedimentation in
aneutral
sucrosegradient (Fig.
6).
DNAsynthe-sized
ina 5-min reaction sedimentedassmallercomponents
inanalkaline
sucrosegradient (Fig.
7A).
However,
prolonged
incubation didnotre-sult
inthe
elongation
of the
smaller molecules
(Fig.
7Band
C). No decrease in the size of3H-labeled
AAV-1 DNAwasdetected
when AAV-1DNAwasincubated
with thereplication
com-plex under the
sameconditions, suggesting
thaton November 10, 2019 by guest
http://jvi.asm.org/
[image:4.500.49.244.332.548.2]TABLE 2. DNA-DNA hybridization of DNA synthesized in vitro
DNAimmobilized(%)
InputDNA(cpm)~ ~ ~ ~ ~ ~ ~ ~ ~ ~ - Blankfilter
Input DNA (cpm) AAV-1 DNA H5 DNA Cellular DNA
(cpm)
(cpm) (cpm) (cpm)
DNAsynthesizedinvitroa (8,400 2,856(34)b 391(4.7) 428(5.1) 60
cpm)
AAV-1 DNAc(5,800cpm) 2,088(36) 93(1.6) 87
(1.5)
58H31 DNAC (5,200cpm) 104(2.0) 2,236 (43) 88(1.7) 53
HEK DNAc(8,800 cpm) 185(2.1) 167(1.9) 1,848 (21) 64
aDNAsynthesizedin vitrowasextractedasdescribed in Materials andMethods,
dialyzed against
O.JX
SSC(0.15MNaCl plus0.015 M sodiumcitrate)and 10 mMEDTA,and annealed with AAV-1DNA,H5DNA,and humanembryolivercell DNA immobilizedonmembranefilters. DNA-DNAhybridizationwascarried outas
describedpreviously (22).
bFigures inparenthesesshow percentages of DNAhybridizedtoinputDNA.
cAscontrols,labeledDNAfrompurified
visions
ofAAV-1, H5,and fromgrowingHEKcellswereincludedin thetest.
15
x
E
5
5 10 15 20 25
Fraction number
FIG. 6. Analysis of DNA synthesized in vitro in neutralsucrosegradients.DNAsynthesizedinvitro
with thecomplexat37°C for60 minwassedimented
in aneutralsucrosegradient. Theposition of 3P-labeled AA V-I DNA(amarker) isshown byan arrow.
the formation ofsmaller-sized DNAduring the
in vitro reaction wasnot due to the action of
DNasein thecomplex (Fig. 7D).
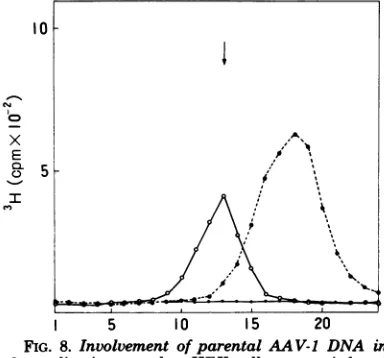
InvolvementofparentalAAV-1 DNAin
thereplication complex.Confluent
monolay-ersof HEK cellswerecoinfectedwithH5tsl25
and[3H]thymidine-labeled AAV-1. At18 hafter
coinfection,thecomplexwasextracted and
sed-imentedina sucrose gradient. The radioactive
peak coinciding with the position of the AAV-1
DNAreplication complexwasdetected(Fig. 8).
About 1% of theinput AAV-1 DNA was found
in the
replication complex.
When HEKcells
were infected with3H-labeled AAV-1
alone,
noradioactivepeakwasdetected.
DISCUSSION
The
AAV-1 DNAreplication complex
wassolubilized and
partially
purified from the nuclei
of HEK cells coinfected with AAV-1 and
H5ts125
at40.5°C,
toestablish
anin
vitro
systemfor AAV DNA
replication. The complex, with
a meansize of 23S, synthesized exclusively
AAV-1DNA
asidentified
by DNA-DNA
hybridiza-tion and sedimentahybridiza-tion in
aneutral
sucrosegra-dient.
However,
sedimentation in
analkaline
sucrose
gradient showed that the products
con-sisted
mainly of shorter chains than
AAV-1DNA. The
results suggestthat the
replication
complex
lacks
somefactors,
such
asDNA
ligase,
which
are present in the intact isolatednuclei
and
necessaryfor the formation of
complete
AAV-1 DNA molecules, since AAV-1 DNA of
the
maturesize
wassynthesized
in vitro with
isolated nuclei
(5).
The
analysis
of the
parental
AAV-1 DNA in
analkaline
sucrosegradient
would also
provide
anexplanation
for this
ob-servation. No AAV DNA
replication complex
wasformed
incells infectedwith
AAValone. It hasbeen
reported
that AAVvirions contain
single-stranded DNA,
which is either aplus
orminus
linear
strand(14, 18). After single
infec-tion
withAAV,
theeffective
adsorption and
penetration
ofthe
AAV genomeinto the nucleus
was
observed
(20;
H.Handa, unpublished data).
The
conversion of
single-stranded
DNA todou-ble-stranded
DNA(replicative
form)
must benecessary
for the
AAVreplication cycle,
as inthe
replication
of otherparvoviruses (16, 25).
The above
results
suggest that thereplicative
form is not
formed
incells after
infectionwith
AAV alone. Further studies
are necessary to448
HANDA AND SHIMOJOon November 10, 2019 by guest
http://jvi.asm.org/
x E
o-
0.5-I
.q
5min
41
6
x E
I 31
21
[image:6.500.96.392.60.402.2]15min
FIG. 7. AnalysisofDNA synthesized in vitro in alkaline sucrosegradients. DNA synthesized in vitro
with thecomplexat37Cfor(A) 5, (B) 15,and(C)60 minwassedimented inalkalinesucrosegradients. (D)
3H-labeledAAV-1 DNA was sedimentedsimilarly afterpreincubation with the AAV-1 DNA replication complexat370Cfor60min. Theposition of32P-labeledAAV-1 DNA(a marker)is shownbyanarrow.
elucidate the defective step of the AAV DNA
10 replication.
We have suggested that a helper factor(s)
induced by infection with adenovirus is closely
related to the formation ofadenovirus-specific
T antigen (H. Handa, K. Shiroki, and H. Shi-mojo, submitted
for
publication).
Furtherpuri-x E.' 8 fication of the AAV DNA replicationcomplex
5 and the establishment of a complete system
I, t .would provide amethodto investigatethe
na-ture of factors involved in the AAV DNA
repli-f\: \ cation.
ACKNOWLEDGMENT
We aregratefultoK.Shirokiforcooperation and toK. Oda forcritical reviewof themanuscript.
1 5 10 15 20
FIG. 8. Involvement ofparentalAAV-1 DNA in thereplication complex. HEKcells werecoinfected
with
13H]thymidine-labeled
AA V-1(0.1cpm/ fluores-centinfectious unit)andH5ts125at40.50C.Symbols: AAV-1 DNA replication complex extractedsedi-mented in aneutral sucrosegradient
(0);
extractpreparedfromthe cellsinfectedwith
13Hlthymidine-labeledAAV-I alone
(-4*);
and 3H-labeledAAV-1DNAextractedfrom purifiedvirions(---0).
Theposition ofthe AAV-1 DNAreplication complex is
shownbyan arrow.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.500.44.236.454.633.2]This work wassupported bygrants from theMinistryof Education, Science and Culture, Japan, and the Princess TakamatsuFund for Cancer Research.
LITERATURE CiTED
1.Atchison,R. W.,B. C.Casto, andW. M. Hammon. 1965. Adenovirus-associated defective virusparticles. Science 149:754-756.
2. Blacklow, N. R., M. D. Hoggan, and W. P.Rowe. 1967.Immunofluorescent studies of the potentiation of anadenovirus-associatedvirusby adenovirus7. J. Exp. Med.125:755-765.
3. Chang,L. M. S.1973. Lowmolecularweight deoxyribo-nucleic acidpolymerase from calf thymus chromatin. I.Preparationof homogeneousenzyme. J.Biol. Chem. 248:3789-3795.
4. Ensinger,N.R., andH.S.Ginsberg. 1972.Selection and preliminarycharacterization of temperature-sensi-tivemutantsof type 5adenovirus.J. Virol.10:328-339. 5. Handa, H., andILShimojo.1977.ViralDNAsynthesis in vitro with nuclei isolated from adeno-associated virus type 1-infected cells.Virology77:424-428.
6. Handa, H., H. Shimojo, and K. Yamaguchi. 1976. Multiplicationof adeno-associated virus type 1 in cells coinfectedwith atemperature-sensitive mutant of hu-manadenovirustype 31.Virology 74:1-15.
7. Handa, H.,K.Shiroki,and H.Shimojo.1975. Comple-mentation ofadeno-associatedvirus growth with tem-perature-sensitivemutants ofhumanadenovirus types 12and5.J.Gen.Virol. 29:239-242.
8. Hoggan,M.D., N. R. Blacklow, andW. P. Rowe. 1966.Studiesofsmall DNA viruses found invarious adenoviruspreparations: physical, biological,and im-munological characteristics. Proc. Natl. Acad. Sci. U.S.A.55:1467-1471.
9. Ishlbashi, M., andM. Ito. 1971. The potentiation of type1adeno-associatedvirusbytemperature-sensitive conditional-lethalmutantsof CELO virus at the restric-tive temperature.Virology45:317-320.
10.Ito, M., and E.Suzuki.1970.Adeno-associatedsatellite virusgrowth supported by a temperature-sensitive mu-tantof human adenovirus. J.Gen. Virol. 9:243-245. 11.L~ndeli,T.J., F. Weinberg,P.W.Marris,R. G.
Roe-der, andW. J. Rutter. 1970. Specific inhibition of nuclear RNA polymerase II by a-amanitin. Science 170:447-449.
12.Lowry,0. H.,N. J.Rosebrough,A. L Farr,and R. J.Randall.1951.Protein measurementwiththe Folin phenolreagent.J. Biol. Chem.193:265-275.
13. Mayor, H.D., and J. Ratne. 1973.Analysis of
adeno-associatedsatellitevirusDNA. Biochim.Biophys.Acta 299:189-195.
14. Mayor, H.D., K.Torikai,J. LMelnick, and M. Man-del. 1969.Plus andminussingle-strandedDNA sepa-ratelyencapsidated inadeno-associated satellite viri-ons.Science166:1280-1282.
15.Parks,W.P.,J.LMelnick,R.Rongey, and H.D. Mayor. 1967.Physicalassay andgrowthcyclestudies of adefectiveadeno-satellitevirus. J.Virol.1:171-180. 16.Rhode, S. L1974.Replicationprocessof theparvovirus H-I.L.Isolation and characterization of H-Ireplicative formDNA.J. Virol. 13:400-410.
17.Rose,J. A. 1974.Parvovirusreproduction, p. 1-62. In H. Fraenkel-Conrat and R.R. Wagner(ed.), Comprehen-sivevirology,vol. 3.PlenumPress,New York. 18.Rose,J. A.,K I. Berns,M. D. Hoggan, andF. J.
Kocmot.1969.Evidencefor asingle-stranded adenovi-no-associatedvirusgenome: formation of a DNA den-sity-hybridonreleaseofviral DNA.Proc. Natl. Acad. Sci.U.S.A.64:863-869.
19. Rose,J.A.,M. D.Hoggan,and A. J.Shatkin.1966. Nuclei acidfrom anadeno-associated virus: chemical and physical studies. Proc. Nati. Acad. Sci. U.S.A.
56:88-92.
20. Rose,J.A.,andF. J. Koczot. 1972. Adenovirus-associ-atedvirusmultiplication.VII.Helper requirementfor viral deoxyribonucleic acid synthesis.J. Virol.10:1-8. 21. Sedwick, W.D.,R.S.-F. Wang, andD.Korn. 1972. Purificationandpropertiesofnuclear andcytoplasmic deoxyribonucleic acid polymerases from human KB cells.J.Biol. Chem. 247:5026-5033.
22. Shimojo, HI, and T. Yamashita. 1968. Induction of DNA synthesis by adenoviruses incontact-inhibited hamstercells.Virology61:474-485.
23. Smith, K. O., W. D. Gehle, andJ. F. Thiel. 1966. Propertiesof asmallvirusassociatedwithadenovirus type 4. J.Immunol.97:754-766.
24. Straus, S.E.,H.S.Ginsberg,and J. A. Rose. 1976. DNA-minustemperature-sensitive mutantsof adeno-virus type 5help adenovirus-associatedvirus replica-tion. J.Virol.17:140-148.
25. Tattersall, P., L V.Crawford, and A. J. Shatkin. 1973.Replicationof theparvovirusMVM.II.Isolation and characterizationof intermediates in thereplication oftheviraldeoxyribonucleicacid. J.Virol.12:1446-1456. 26. Weissbach, A., S.-C. L Hong, J. Aucker, and R. Muller.1973.Characterizationofherpessimplex virus-induced deoxyribonucleic acid polymerase. J. Biol. Chem.248:6270-6277.
27. Yonetani,T. 1959.StudiesoncytochromeA.III. Effect ofsynthetic detergentsupon theactivityofcytochrome A. J.Biochem. 46:917-924.