0022-538X/80/01-0272/14$02.00/0
Identification of the Herpes Simplex Virus DNA Sequences
Present in
Six Herpes Simplex Virus Thymidine
Kinase-Transformed
Mouse Cell Lines
JEFFREY M.
LEIDEN,'
NIZAFRENKEL,'* ANDFREDRAPP2Department of Biology, The University of Chicago, Chicago, Illinois60637,1 and Department of Microbiology and Specialized Cancer Research Center, The Milton S. Hershey Medical Center, The
Pennsylvania State University College of Medicine, Hershey, Pennsylvania170332
We
have
used
anovel filter
hybridization approach to detect and map theherpes
simplex virus
(HSV) DNA
sequences which are presentin four HSV
thymidine kinase (HSVtk+)-transformed cell lines which
werederived by
exposureof
thymidine kinase negative (tk-)
mouse cells to UV light-irradiated HSV type 2(HSV-2).
Inaddition, we have mapped theHSV-1
DNAsequences which are presentin
twoHSV-ltk+-transformed
cell lines
produced bytransfection of
tk-mousecells with sheared HSV-1 DNA. The results of these studies
canbe
summarized
asfollows.
(i)
The
only
HSV DNA
sequenceswhich
werecommon toall HSVtk+-transformed
cells
werethose
located between
mapcoordinates
0.28and
0.32.Thus, this region contains
allof the viral
DNAsequenceswhich
are necessaryfor the expression
of HSV-mediated tk transformation. (ii)
Manyof the
cell lines also contained variable
amountsof non-tk
geneviral DNA
sequenceslocated between
mapcoordinates
0.11 to 0.57and
0.82 to 1.00,suggesting that
incorporation of the viral DNA
sequenceslocated between these
mapcoordinates
is
arelatively random
event.(iii) The viral DNA
sequenceslocated between
mapcoordinates
0 to 0.11and
0.57 to 0.82 wereuniformly absent from all of the
HSVtk+ cell lines
tested,
suggesting that there is
a strongnegative
selective
pressureagainst incorporation of these viral DNA
sequences.During their
lytic cycle, herpes simplex
vi-ruseshave been
shown
toprogramthe
synthesis
of
avirus-specific thymidine
kinase.
(tk)
enzyme(6, 15). This tk has been shown
tobe distinct
from
both the nuclear and mitochondrial
mouseand hamster cell
enzymeswith
respect to anumber
of biochemical and
immunological
prop-erties, including thennal
stability,
electropho-retic
mobility,
and
antigenic specificity
(16, 26).
The
enzymehas
been shown
tobelong
tothe
,Bclass
(11) of viral
proteins,
in that its
synthesis
requires
the
presenceof
functional
early
(a)
viral
polypeptides and is turned off late in infection
by late
(y)
viral
polypeptides (7,
18; R. Honess
and B.Roizman,
unpublished observations).
Munyon
etal.
(27)
werethe
first
toshow that
asmall
proportion
of
tk-negative (tk-)
mousecells could
bestably
transformed
tothe tk-pos-itive(tk+)
phenotype
after exposureof
thetk-cells
toUV
light-irradiated herpes
simplex
virus type 1(HSV-1).
Thetk
enzymemade in
suchbiochemically
transformed
cells has beenshown
topossess thebiochemical
andimmunological
characteristics ofthe
tkproduced during lytic
HSV infections
(4, 17, 26,
34).
More
recently,
transformation of tk- mousecells
to thetk+
phenotype has beenaccom-plished by transfecting the tk-
cells
with either
sheared HSV
DNA,
orisolated restriction
en-zymefragments of HSV DNA (1,
21, 23,37). Byusing these
approaches, Wigler
etal. (37) have
shown
that
a3.4-kilobase
fragment
of HSV-1 (F)
DNA is sufficient
tostably
transform tk- cells
tothe
HSV
tk-positive (HSVtk+)
phenotype.Furthernore, Maitland and McDougall
(21)have
reported that the purified restriction
en-zymefragments
of
HSV-2 (333) DNA which
contain the viral DNA
sequenceslocated
be-tween mapcoordinates
0.53and
0.65 aresuffi-cient
totransform tk-
mousecells
tothe
HSVtk+
phenotype.
This
maplocation
for the HSV-2
tk-transforming
genewassurprising
inlight
of re-centstudies of HSV-1
xHSV-2 recombinants
which
haverevealed
that most,if
notall,
of
the genesof these
twoviruses
arecolinear
(24,
25,
29),
andwhich have
mapped
theHSV-1 tk
genebetween coordinates
0.27and 0.35(25).
The studies
describedin
this paper werede-signed
tobothunambiguously
mapthe HSV-1
andHSV-2
tk genes andtostudy
thepatterns
of
incorporation
of non-tk geneviral
DNAse-quences in
HSVtk+
transformantsproduced by
infection of tk-mouse cells with UV light-irra-diatedHSVorby
transfection of thesecells with272
on November 10, 2019 by guest
http://jvi.asm.org/
HSV DNA
SEQUENCES
IN HSVtk+-TRANSFORMED 273sheared viral DNA. Toward these endswehave
developed
a novelhybridization mapping
ap-proach
which has allowed the detection and identification of the HSV DNA sequences which are present in a number ofHSVtk+
cell lines. Our results have revealed that(i)
theonly region
of the HSV genomewhich
is present inall
HSVtk+-transformed cells is located between
map
coordinates 0.28 and0.32, and (ii)
awide
variety
of non-tk gene viral DNA sequences can bestably incorporated
into theHSVtk+
cells.However, specific regions
of theHSV
genome appear to beuniformly
absent from suchHSVtk+
transformants.MATERIALS AND METHODS
Virus. HSV-2 strains 333 and 324 (both isolated frompenile lesions) and Silow (isolated from a vaginal lesion)wereobtained from W. Rawls (McMaster Uni-versity,Hamilton, Ontario,Canada). HSV-1 (1023) is arecombinant betweenHSV-1 (HFEM) and HSV-1 (MP) (32).HSV-2 strain G, HSV-1 strains F and MP, and HSV-1 (1023) were obtained from B. Roizman
(UniversityofChicago,Chicago,Ill.). HSV-1 (Justin) wasobtained from A.Sabin.
HSVtk -transformed mouse cells. HSVtk+-transformed cell lines 33A+, 39A+,
59W,
and Silow wereobtainedby the biochemical (tk)transformation method ofMunyon etal. (27), as modified by Rapp and Turner (30). NclAcllO (tk-) cells, inbred Swiss mouse cells lacking tk activity, were originally ob-tained from R. Goldberg (National Institutes ofHealth, Bethesda, Md.). The stocks of HSV-2 to be used fortransfornationwereUV-irradiated for5min
at 46 ergs/s per
mm2.
This UV-irradiated viruswasthen usedtoinfect the NclAcllOcells insuspension
(6x 105cellsper 100-mmplate) at a multiplicity of 2 to 4PFU/cell (as calculated before UV irradiation). After 72 h,selectivemediumcontaining MTAGG (0.28 ygofmethotrexate sodium perml,4,ugof thymidine perml, 13
jug
of adenosine perml, 14,ugofguanosine perml, and7.5,ugofglycineperml) replaced nonse-lectivemedium(27).Three weeks afterinfection,foci of tk+-transformed cells appeared on the infected plates and werepicked witha Pasteurpipette. Celllines were grown and continuously propagated in
MTAGG-containingmedium.HSVtk+ cell lines33A+ and39A+weretransformedbyHSV-2(333), cellline
59W
wastransformedbyHSV-2 (324), andcellline SilowwastransformedbyHSV-2(Silow).Cellswerepassagedatleast50 to 100times (every4to 5days) before being used in the studies described in this paper. Cell lines 8N and 5Awere obtained from D.
Polacek and B. Roizman. These celllines were
pro-duced after transfection of L tk- CllD cells with sheared HSV-1 (1023) DNA, by using the calcium-phosphateprecipitationmethodof Grahametal. (8). After transfection, the cellswereswitchedto Little-field's HAT medium (0.01 M hypoxanthine,4.4,IM methotrexate, and 16,uMthymidine) (20). Surviving colonieswerepicked by using small glass cylinders and werefurtherpropagated inHAT-containing medium. Passage15of each of thesecell lineswasused inthe experiments describedbelow.
Assay of transformed-cell tk activity. HSV-2tk+ cells between passages50 to 100 wereassayedfor tk activityby using the method of Lin andMunyon
(19) asmodifiedby Rapp and Turner(30). Both non-infected and infected (18 h with HSV-2 (333), at 2 PFU/cell) controlcultures,composedofnormalNclA cllO(tk-)and NIH Swissmouse(tk+) cells,werealso assayed.
Toselectively inactivate the viral tk activity,cell
extracts wereincubatedat40°C for 30 min (28).
Purification of viral DNA. Purification of viral DNAwas performed asfollows. Vero cells were in-fected with 5 PFU/cell of HSV in 199-V medium containing 1.5
juCi
of [3H]thymidine (Amersham Corp.) per ml. After 20 hat 37°C,the infected cells werescraped into the medium andpelleted. The cell pelletwasrinsed twice with coldphosphate-buffered saline andresuspended in lysisbuffer (0.1 M NaCl,0.01 MTris-hydrochloride(pH8.0),0.01MEDTA;13 ml/109cells). Sodiumdodecylsulfate(SDS) and pro-nase (Sigma Chemical Co.), pretreated according to Hotta andBassel(12),wereaddedtofinal concentra-tions of 0.6% and1mg/ml,respectively,and the
mix-ture was incubated at 37°C for 4to 6h. The lysis
mixturewasthen dilutedto afinal volume of150ml with 0.01 M Tris-hydrochloride (pH 8.0)-0.001 M EDTA(TE)and addedto195gof solid CsCl(Eastern Chemical).The refractive index of theresulting solu-tion was adjusted to 1.4005, and the mixture was
centrifugedat40,000rpmfor 20 hat20°Cina Beck-manVTi5OrotorinanL-5centrifuge.Fractions con-taining viral DNAwerecombined. Therefractive in-dex of this pooled peak material wasreadjusted to 1.4005withaCsClsolution,and theresulting mixture wasrecentrifugedtoequilibrium asdescribed above. The viralpeak material from these secondgradients
waspooled anddialyzed against TE buffer.
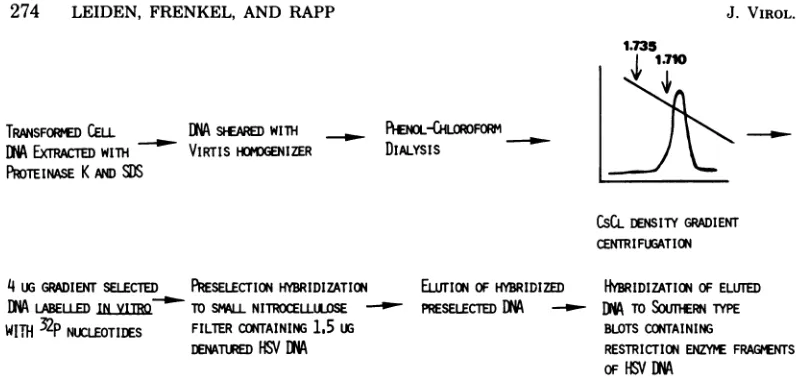
Mapping approach.Themapping approachwhich has been usedto identify the viral DNA sequences presentin theHSVtk+celllines isschematically rep-resented in Fig. 1. Basically, this approach involves threesequential steps: (i) aninitial density gradient
centrifugation step of the transformed-cell DNA,to enrichfor transformed cell-associated viral DNA se-quences, (ii)invitrolabelingof thegradient-selected
DNA followedbyapreselectionhybridizationtoHSV DNA immobilizedon nitrocellulosefilterstofurther enrich for the cell-associated viral DNA sequences, and(iii) hybridization of thepreselected transformed-cell DNA to blots containing restriction enzyme
frag-mentsof HSV DNA.
(i) Extraction and CsCl density gradient
cen-trifugationoftransformed-cell DNA. DNA from either1.25x 108or2.5x108cellswaslabeled in vivo by growing the cells for2daysin the presenceof 0.5 ,uCiof[3H]thymidine (Amersham Corp.) perml.The cellswereharvestedby rinsing the monolayers three times with ice-cold phosphate-buffered saline, fol-lowedby lysis in25mlofamixturecontaining 0.01 M
Tris-hydrochloride (pH 8.0),0.01 M EDTA, 0.01 M NaCl, 0.5% SDS, and1mgof Proteinase K (Merck &
Co., Inc.) per ml. The lysis mixture was incubated overnightat37°C and sheared on icefivetimes (30s/
time) at20,000 rpm inaVirTis45homogenizer. The
sheared DNA wasextractedwith 1volumeofphenol
and1volume ofchloroformandwasdialyzedat room VOL. 33,1980
on November 10, 2019 by guest
http://jvi.asm.org/
TRANSFORMED CELL DN SEAREDWITH _ ENOLCHLOROFORM
DNM
EXrRATDwim VIRTIS ZVGENIZER DIALYSISPROTEINASE KAm
SDS
CSCLDENSITYGRADIENT
CENTRIFFUGATION
4
UG
GRADIENT
SELECiTE
PRESELECTION
HYBRIDIZATION DN LABELLED INVITRO TOSMA-LLNITROCELLULOSE-WITH
32P
NUCLEOTIDES FILTERCONTAINING 1.5 uGDENATUED HSVDNA
EhLTION OFHYBRIDIZED PRESELECTED DNA
-_-HYBRIDIZATIONOF ELUTED
DMA
TOSOUMERN TYPE BLOTS CONTAININGRESTRICTION ENZYMEFRAGMENTS
[image:3.504.60.458.52.241.2]OF HSV
DNA
FIG. 1. Diagrammatic representation of the hybridization approach. The individual steps of this approach aredescribed in detail in the text.
temperatureforapproximately12h against TEbuffer. The DNA mixture was then resheared five times as above, adjustedto atotal volume of 25 ml, and added to 32.5 gof solid CsCl (Eastern Chemical) to yield a final refractive index of 1.4005. Equilibrium density centrifugationwasdone inaVTi5O Beckman rotor at 40,000 rpmat20°Cfor 20 h.Sixty
600-pl
fractions were collected per gradient. The fractions between densities 1.710and 1.735g/cm3 (asdetermined from refractive indices) were pooled and dialyzed against several changes of TE bufferat4°C for3days. The DNA was then precipitated with ethanol. Figure 2 shows the results of a reconstruction experiment in which a mixture ofpurified 32P-labeled HSV-2 DNA and 3H-labeled tk- cell DNA wassheared andsubjected toequilibrium densitycentrifugation as described above. As shown in Fig. 2, the sheared viral and cell DNAs sedimented in discrete bands. The fractions between densities 1.710and 1.735g/cm3contained 75% of the
32P-viral DNA label and only 2% of the 3H-labeled
cellularDNA, representinga40-fold enrichment.
(ii) Invitro labeling of DNA. Viral DNA and CsCl gradient-selected transformed-cell DNAswere labeled in vitro with [a-32P]dGTP and[a-32P]dCTP
(350 Ci/mmol; Amersham Corp.) with Escherichia coli DNA polymerase I, essentially as described by
Maniatis et al. (22). The specific activities of DNA labeledbythisprocedure rangedbetween 7x 107and 1.6x 108
cpm/4g.
(iii) Preparation of nitrocellulose filters
con-taining HSVDNA. The small nitrocellulose filters used in thepreselectionhybridizationstepwere pre-paredasfollows. A10-to15-,ugamountofHSV-1 or HSV-2 DNAwassuspendedin0.9mloflxSSC(lx
SSC=0.15MNaClplus0.015Msodiumcitrate)and
sheared fivetimesthrougha26-gauge needle. A
0.1-ml amountof3NNaOHwasadded,and the mixture
wasincubated for10minat roomtemperature.This
mixture was then added to 9 ml of 28x SSC and
filteredtwotimesthroughaHAWP02500 nitrocellu-lose membrane filter (Millipore Corp.), which had
1250-2
0
0.
NL
1000- 750-500
250
N
1-I\'
~~~~
.1~~~~~~~
N-'
-1.74
-1.70
11.66 r400
300
200
100
z 0
s
2
8 16 24 32 40 48 56
FractionNumber
FIG. 2. CsClequilibrium densitycentrifugationof amixtureofshearedtk-cell DNA andHSV-1DNA. A mixtureof
32P-labeled
HSV-1DNA and3H-labeled
CIIDtk- cellularDNA wasshearedto an average molecularweight of5x 106and sedimentedtoequi-librium inaCsCldensity gradientasdescribed in the text.
been prewashed with 28x SSC. After filtration, the filterwaswashed with10ml of28xSSC, dried over-nightat roomtemperature, and bakedat80°C for5h. Eight replicate 2-mm-diameter filterswere punched out of each 25-mm-diameter filter, with a leather punch. It wasfoundthat filtration in alkalias
origi-nallydescribedby Baker (2) along with theuseof very high salt(28x SSC) resultedinthe best retention of HSV DNA.
Nitrocellulose "blots" containing various restriction enzyme fragments of HSV DNA were prepared as follows. HSV DNA wasdigested with restriction en-donucleases(NewEngland Biolabs),andsubjectedto
r
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.266.457.283.443.2]electrophoresisat50 V for 20 h inahorizontalagarose gel (10 by30cm;0.4%),asdescribed byHayward (10).
A 10-to15-,ugamountof HSV DNAwasloadedonto 4-cm-wide wells. DNA wastransferred from the gel
ontoasheet ofBA 85 nitrocellulose filter(Schleicher &Schuell Co.), essentially asdescribedbySouthern
(33). Afterblotting, replicate blotswereprepared by cutting the nitrocellulose filter into 3-mm vertical stripswhichwere allowedto dry overnight atroom
temperatureandthenbakedat80°Cfor 5 h.
(iv)Preselectionhybridizations. Nitrocellulose
ifiters(2-mm diameter), preparedasdescribed above
andcontaining1.5
tLg
of immobilizeddenatured HSV DNA,werepreincubated for6 h at65°Cinasolutioncontaining6x SSC and 0.02% bovineserum albumin
(BSA), 0.02% polyvinylpyrrolidone, and0.02% Ficoll (5).
Invitro-labeled transformed-cell DNA (4ug) was
precipitated with ethanol ina1.5-ml microfuge tube
andresuspended by heatingat65°C for30minin 20
pl
ofhybridization solution containing6xSSC,0.01 M EDTA, 30% formamide, and 1 mg of E. coli DNA (Miles Laboratories, Inc.) per ml. The resuspendedDNAwas thensealed in acapillary pipette and
de-natured for 7minat117°C.Theresultingdenatured
DNAwasplacedintoaBeemcapsule (Beem Corp.),
and 2
pl
of 1Ox Denhardt solution (lx Denhardtsolution is 0.02%BSA,0.02%Ficoll, 0.02%
polyvinyl-pyrrolidone)inwaterwasadded. Apreincubated
ni-trocellulosefilter (2-mm diameter) containing
immo-bilizedHSVDNAwasblotteddryandsubmergedin
thehybridizationsolutionatthe bottomof the Beem capsule. The filter and hybridization solution were
overlaid with mineraloil,andhybridizationwas
car-ried outat65°Cfor 16 h. Afterhybridizationthe small
nitrocellulose filterwaswashed threetimesatroom
temperature with 2x SSC and twice for 1 h at65°Cin
10 ml of 6x SSC-30% formamide-lx Denhardt
solu-tion-0.5% SDS. After these washes, the hybridized
DNAwaselutedfrom the filtersbyincubation of each
filterin 100
pl
of elution buffer (0.1MNaOH,0.01 M EDTA, 0.1%SDS) at45°Cfor 2h,asoriginallyde-scribedby Gronebergetal.(9).Thesolution of eluted
DNAwasneutralized with 1 NHCI.Itshould be noted
that the efficiency of the preselection hybridization
stepwasgreatly improved by minimizingthe volume
of the hybridization solution. For convenience, we
choseafinalvolume of 20 tdperfilter,yieldinga50%
hybridization efficiencyforpurifiedviral DNA anda
background of0.02 to 0.04% for cellular DNA and, therefore, representing a 1,200- to 2,500-fold
enrich-mentfor viral DNAsequences.
(v)Blothybridizations.Nitrocellulose blotswere
preincubatedfor6 hat65°CinalxDenhardt solution
in 6x SSC.Thesepreincubatedfilterstripswerethen
hybridized inaglassscintillation vialcontaining the
elutedDNA fromthepreselection hybridizationstep
in1ml ofasolutioncontaining6xSSC,lxDenhardt
solution, 30%formamide, 10% dextransulfate
(Phar-macia FineChemicals, Inc.),0.5%SDS, and200ytg of
denatured E. coli DNA per ml. Hybridization was
carried out at 65°C under mineraloilwith vigorous
shakingfor 20h.It should benotedthatthe inclusion
of 10% dextran sulfate(36)intotheblothybridization
mixtures was found to result in a two- to fourfold
increase in thehybridization efficiency.Inaddition, it wasfound that theincorporationofexcessunlabeled E. coli DNA into the hybridization solutions resulted insignificantlyreduced levels ofbackground,as com-paredtohybridizationsperformedinthe presence of
excessunlabeled salmon sperm DNA. After
hybridi-zation, the nitrocelulose strips were washed three timesat roomtemperaturewith2xSSC and twice for 2 h with 6x SSC-lx Denhardt solution-30% form-amide-0.5% SDS at 650C. Finally, thenitroceUulose strips were dried andautoradiogrammed onCronex 2c medical X-ray film (Du PontCo.) either with(at -70°C),orwithout(atroomtemperature)high speed
intensifyingscreens(Du PontCo.). Whenapplicable,
the relativeintensities ofautoradiographicbandswere determined by scanning the autoradiograms with a Transidyne2955scanningdensitometerequipped with anautomaticintegrator.
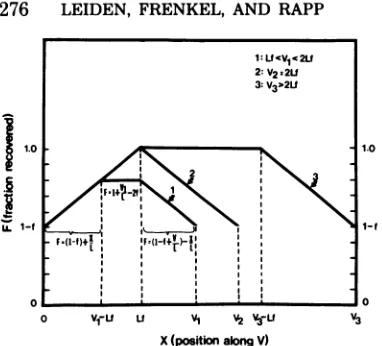
Theoreticaltreatmentof the fractional recov-eryofintegratedviral DNA from the CsCl
equi-librium density centrifugation step. Consider a piece of HSV DNA of average viralbuoyant density,
Pv (=
1.729g/cm3)
(14),
and sizeV,
which isintegrated
intoceUular DNA sequences of average buoyant den-sity
pc
(= 1.690g/cm3).
After randomshearing of the transformed-ceU DNA to double-stranded fragmentsof sizeL, anyintegrated sequence of viral DNA
wil
be present in anassortmentofpieces of sheared DNA, containing various amounts of viral and ceUular se-quences. In order for a piece of this sheared trans-formedcellDNAtoberecoveredfrom the CsClequi-librium density centrifugation step (i.e., to display a
buoyantdensitygreaterthanorequaltop=1.710g/
cm3),
itmustcontainaminimumfraction of viral DNA sequences. This minimum fraction(t)
canbe easilycalculated from thefollowing equation:p=
(t) pv
+(1 - f)pc (equation 1). Itfollows that for apiece of
transformed-cellDNAoflength L,Lfrepresentsthe minimum amountof viral DNA whichmust be con-tained in thatpieceifit istobe recoveredfrom the
densitygradient preselectionstep.
Assumingthat
pv
= 1.729g/cm3,
Pc = 1.690g/cm3,
p=1.710
g/cm3,
and L=6 x106(i.e., the average sizeof the sheared transformed-cell
DN4),
then fromequation1f=0.5andLf=3x106.
Thds,
anypiece
of integrated viral DNA of molecularweightVless than 3x10'
willnotberecovered from the CsCl gradient purification step. On the other hand, when the size of anintegrated piece of viral DNA is greater than 3x106, allofthe viral DNA sequences contained in this piece of integratedDNA wiU be recovered tosome
extentduring the CsCl purification step. Theexact
fractional recovery ofagivenviral DNA sequence will
dependuponitspositionXalongthe stretch of
inte-gratedviral DNA.Therelationshipbetween the frac-tional recovery F and the position X, for the viral DNA sequences contained in variouspieces of inte-grated viralDNAof sizes V>Lf is shown in Fig. 3. As seeninthatfigure, those viral DNA sequences located at both edges of the piece of integrated viral DNA (i.e.,at X=0andX= V)willberecovered from the equilibrium density step with anefficiency of F= 1
-fwhich is independent ofboth L, the size of the
shearedtransformed-cellDNA, and V, the size of the integrated viral DNA sequence. As described above
on November 10, 2019 by guest
http://jvi.asm.org/
276
LEIDEN, FRENKEL, AND RAPP1.0
1.00 V1-fu U VI V2 V3U V3
X(positionalongV)
FIG. 3. Diagrammatic representation of the frac-tionalrecoveryofintegrated viral DNAsequences as afunction of the position of the integrated viral DNA sequences. L = average length of sheared
trans-formedcellDNA,X=distanceofagiven viral DNA sequencefromoneend of the integrated piece of viral
DNA, F=fractionalrecoveryfrom CsClequilibrium
density gradient ofagiven viral DNAsequence
lo-catedatpositionX,f=minimumfraction ofapiece
of shearedtransformed-cell DNA whichmustbe
com-posed of viral DNAsequencesif thatpiece of
trans-formed cell DNA is to be recoveredfrom the CsCl equilibrium density centrifugationstep,and V=the
sizeofan integrated stretch of viral DNA. As
de-scribed in thetext, agivenviralDNAsequencewill
be included inan assortment of fragments of
ran-domly sheared transformed-cell DNA. Fwas
calcu-latedasthefraction ofthese fragments which contain
atleast Lfamountof viral DNA sequences. Fwas
calculatedfor threecasesdiffering withrespect tothe size Vof theintegrated viral DNAasdepicted in the upperright handcornerofthefigure.
usingourexperimentalconditionsf=0.5.Therefore,
those viral DNAsequenceslocated atboth edgesof
thepiece ofintegratedviral DNAwillbe recovered
with an efficiency F = 0.5. Viral DNA sequences
locatedatinternalpositionsof theintegratedstretch
of viral DNA (V> X> 0) will be recovered with
efficiencieswhichvarylinearlybetween1-f(= 0.5)
and 1.0. Insummary,by usingourexperimental
con-ditions this theoretical treatmentpredictsthat all of
the viral DNAsequencespresentinintegrated pieces
ofviral DNA withmolecular weightsgreaterthan 3
x
10'
willbe recoveredwitha minimum efficiencyof50%.
RESULTS
Biochemical characterization ofHSVtk+ transformed celllines.
HSV-2tk+
transform-antswereobtainedasdescribedabove after ex-posureofNclA cllO(tk-)mousecells toseveralstrains of UV
light-irradiated
HSV-2(30).
Theresultanttk+ colonieswerecontinuously
propa-gatedinMTAGG selective medium.The various
celllineswereassayedfortkactivityafter 50to
100passages.
These
tests(Table
1)
haveshown
that
all of the cell lines
express atk
activity
which
displays
the
characteristic thermal
labil-ity of the
HSV-2
enzyme(28).
HSV-ltk+ transformants
were produced bytransfecting
L tk-C11D
mouse cells with HSV-1(1023)
DNAwhich
wasrandomly sheared
tofragments of molecular weight
20 x106
to 30 x106
(D.
Polacek and B.
Roizman,
unpublished
data).
After
24h, the cells
wereplaced in
Little-field's
HATmedium
(20)
toselect for
tk+
cells.
Surviving colonies
werepicked
after 2 to 3 weeksand
growncontinuously for
15 to 20 passages inHAT-containing medium.
Characterization of the
mapping
ap-proach.
The
mapping approach which
wede-veloped
(Fig. 1) involves three sequential
steps.In the
first
step,transformed-cell
DNA
is
ran-domly sheared
tofragments
of molecular weight 5x 10 to 7x106
and subjected toCsCl
equilib-rium density
centrifugation.
Because HSV andcellular
DNAsdisplay widely different buoyant
densities (1.726 and
1.729g/cm3
for HSV-1
andHSV-2 DNAs,
respectively, [14] and
1.69 to 1.70g/cm3
for
cellular DNA), this
stepresults in the
partial purification of
anyviral
DNA sequenceswhich
are presentin
the transformed cells.
Inthe second
step,the DNA
banding
atdensities
greaterthan
orequal
to 1.710g/cm3
is labeled,
in
vitro, with
a-32P-nucleotides
and further
en-riched for viral DNA
sequencesby hybridization
to asmall nitrocellulose
filter
containing
dena-tured unlabeled HSV DNA.
Finally,
the
hybrid-ized, labeled transformed
cell
DNA is
eluted
from the
filters and
rehybridized
toSouthem
typeblots
containing
unlabeled restriction
en-zymefragments
of
HSV DNA
(33). The resulting
bands
arevisualized
by
autoradiography and
areTABLE 1. Viral tkactivity in biochemically
transformedmouse cells nmolof[3H]TdR
in-corporatedper mgof
protein in 45 mina
Cellextract Residual
Preincu- activity
Noprein.
bation
atcubation 4oocb
NclA cllO(tk-) 0.029 0.043 148.3 NclA cllO(tk-)+ 0.30 0.081 27.0
HSV-2(333)
NIH Swissmouse(tk-) 0.20 0.18 90.0 NIHSwissmouse(tk-) 0.81 0.15 18.5
+HSV-2(333)
33A+cells 0.27 0.039 14.4
39A+cells 0.23 0.066 28.7
59D+cells 0.48 0.11 22.9
Silow cells 0.35 0.057 16.3
The assay forthymidine kinase wasperformed as
de-scribedin thetext.TdR, Thymidine.
bPreincubationwas
performed
for30min.J.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.70.261.51.224.2] [image:5.504.274.465.479.629.2]identified
by
comparison
toautoradiograms
of
replicate
control blots which
werehybridized
topurified, in vitro-labeled HSV DNA.
Specificity and
sensitivity
ofthe
mapping
approach.
To
assessthe
specificity
of the
map-ping
approach which is
described
above, DNAs
from
uninfected
Veroand
Ltk-
CllD cells,
aswell
ascalf
thymus DNA
wereprocessed
through the
preselection
stepsdescribed above
and
hybridized
toblots
containing restriction
enzymefragments
of viral DNA.
Autoradi-ogramsof these blots
weredevoid of
identifiable
bands
(data
notshown).
In
anattempt todetermine the
sensitivity
of
this
approach
asregards
unintegrated
viral DNA
sequences,either
3 x10-2
or 3 x 10-ljig
ofpurified viral DNA
(representing
1.0and
10cop-ies
percell,
respectively)
wereadded
tolysates
of
2 x 108CllD
tk-
cells, and the resultant
mixtures
weresequentially
processed
asde-scribed above.
The band
patterns of these
recon-struction
hybridizations
werecompared
tothose
of
control
hybridizations in which
replicate
ni-trocellulose blots
werehybridized
topurified, in
vitro-labeled HSV DNA. The results of
thesehybridizations
areshown
inFig.
4and
5and
revealed the
following.
(i)
As seeninFig.
4,the
mapping
approach is
sufficiently
sensitive
todetect
anunintegrated
piece of HSV DNA of
size
106
(e.g.,
KpnI
fragment
S,
1.1 x106
in
molecular
weight),
present in anaverageabun-dance
of
one copy percell.
(ii) In Fig.
5, wecompared
the relative molarities of the
corre-sponding
bands from the
reconstruction and
control
hybridizations.
As seen inthat
figure,
there
was nosignificant difference between the
molarities of the control and reconstruction
bands,
indicating
that the
preselection
stepsdid
notresult
inthe loss of
specific subsets of viral
DNA
sequences.(iii) To determine whether the
mapping
approach
described above would
yield
information
concerning
the
number of
copies
percell of the viral DNA
sequences presentin
agiven
transformed cell
line,
wecompared the
absolute intensities of corresponding bands from
autoradiograms
of the
1and
10 copy percell
reconstruction
experiments. These ratios ranged
from
7.9to21.9(Table
2).
Thus, the technique
isquantitative within limits of
approximately
twofolderror.Because
thebuoyant density
ofcell-associated
HSV
DNA sequencesdepends
uponwhether
these sequences areintegrated into
thehost
genome, it
wasalso necessary
toconsider the
sensitivity
ofthe
mapping approach
forinte-grated
transformed-cell associated HSV
DNA sequences. Whetheraspecific
sequence ofinte-grated viral
DNA will be recoveredfrom
theKpn I
C
AB
p
C ,
D 0
E
s
F t..
G o
H "
I
J
K LM
N
0
Id 5d 5d
ld
10 10 1
C
10II
9DE3-I~I
I I LmS
#N
p
0
RS
PQ
~~~~T
RFIG. 4. Hybridization reconstruction experiment.
Either 3x10-2or3x101pgofpurifiedHSV-2DNA
(representing 1 and 10copiespercell, respectively),
weremixedwithalysateof2x 108CIIDtk- cells.
The resultant mixtureswerepreselectedtoenrich for
viral DNAsequencesandhybridizedtonitrocellulose
blotscontainingtheKpnIorEcoRI/HpaIrestriction
enzymefragments of HSV-2(G)DNAasdescribed in
the text. C = control: replicate blots which were
hybridizedtopurifiedinvitro-labeled HSV-2DNA,
10=invitro-labeled DNAfromthe 10copy percell
reconstruction mixture, 1 = in vitro-labeled DNA
fromtheIcopy per cellreconstructionmixture,Id= 1day of autoradiographicexposure,and 5d=5days
of autoradiographicexposure.
CsCl
density gradient purificationstep (i.e., will be includedinpiecesofshearedtransformed-cell
DNA with buoyant densities greater than orequalto1.710g/cm3) will dependupontheexact
size and density of the integrated viral DNA
sequence, as well as upon the density of the surrounding cell DNA sequences. All of these parameters will, mostprobably, vary from cell
linetocellline,andit istherefore
impossible
toEcoRI/Hpa I
5d
5d
10
1
I
i..
O.
.Y.
0.
.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.250.445.75.409.2]278 LEIDEN, FRENKEL, AND RAPP
FIG. 5. Relative abundances of hybridized bands from autoradiograms of control and reconstruction
hybridizations. Top:densitometric scansoftheautoradiogramsshown in Fig. 4.Bottom: relativemolarities
ofthe various bands. These molarities were calculated relative to the molarities of the corresponding
fragments in virion DNA. See reference 31 for a review of the relevantstructural features of HSV DNA. The relativemolaritieswere obtainedbydividing the fractional area under each peak of thedensitometricscan by theexpectedfractionalarea, asdeterminedfrom the sizesand knownmolaritiesofthe fragments contained in eachpeak,-i.e., for1Mfragments ofsize M (daltons), molarity= (fractional area)
(M/108)-';
for0.5 Mfragments, molarity=2(fractionalarea)
(M/108)
-1;for 0.25 Mfragments,molarity=4(fractionalarea)(MI108)
-1.Eachtripletofbarsrepresents,from lefttoright,therelativemolarities of the bandsfrom
the 10 copy per cellreconstruction,thecontrol,andthe1copy per cell reconstructionexperiments.*, Agroup offragmentsincluding either a 0.5 M or a 0.25 Mfragment, i.e.,containing sequences
from
theinvertedrepeats of HSV DNA(ab and ac). It is noteworthy that these groups of fragments exhibit relative molarities greater than 1.0. Thisfindingmostlikely reflectsthefactthat the ab and ac repeatedsequencesarepresentingreater than unitmolarity in the labeledprobe DNA and, therefore, confirms thehypothesis
that thehybridization issensitive to the amountsoflabeledprobeDNApresent in thehybridization solution.
experimentally
ortheoretically
determine the
absolute
sensitivity
of the
mapping approach
for
all
casesof
integrated
viral
DNA sequences.However, in the theoretical
treatmentpresented
above,
wehave calculated the fractional
recov-ery(from
the
CsCl
density gradient)
of the viral
DNA
sequencescontained in
pieces
of
viral
DNAof average
buoyant density,
which
areintegrated into cellular DNA of
averagebuoyant
density.
Thesecalculationspredict
that,
by
using
ourexperimental
conditions,
any HSV
DNA sequence which is present inapiece
ofintegrated
viral DNA ofmolecular
weight
greater than 3 x 10will be recovered from
theCsCl
gradient
preselection
stepwith
aminimum
efficiency
of 50%.Mapping
of the HSV-2
DNA sequences present inHSV-2tk+
33A+
cells. Cell
line33A+
wasproduced
by infection
oftk-
mousecells
withUV
light-irradiated
HSV-2
(333).
DNA extractedfrom
passage 50 ofthis cell line
waspreselected and
hybridized,
as describedabove,
toblots
containing
either theKpnI
orEcoRI/HpaI
restrictionenzyme
fragments
of
HSV-2
(G)
DNA. Theresults ofthese
hybridi-zations
(Fig. 6)
canbe
summarized
asfollows.
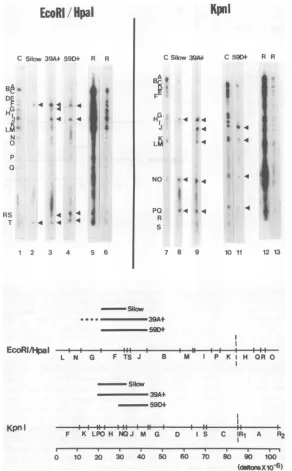
(i) The
mapsgenerated from the
twodifferent
restriction
enzymecleavages used in these
ex-periments
werein
agreementand
indicated that
this
cell line contains
acontiguous
setof
HSV-2
(G)
DNAsequences
located between map
coor-dinates
0.14and
0.57.(ii)
Acomparison
of the
intensities of the
bands
produced by 33A+
DNA with thoseproduced by
the 1 copy per cell reconstructionmixture,
which washybridized
toreplicate
nitrocelluloseblots, revealed
that33A+
cells
contain between 1and
5copies
per cell
ofthese
HSV-2 DNA sequences(Fig.
6). (iii)
Toassess
the
reproducibility
of the
mapping
ap-proach,
twoadditional
preparations of 33A+
DNA(passages
60-70)
wereseparately
prese-lected and
hybridized
todifferent
setsoffilters
andblots
containing
HSV-2
DNA.The
threepreparations of transformed-cell DNA
yielded
identical
hybridization patterns
(Fig.
6).
Mapping of the viral DNA
sequences present inthree
additional
HSV-2tk+
trans-formants.
In an attempt tofurther delineate
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.99.410.65.281.2]TABLE 2.
Comparison of the
intensities of the
ditional
HSV-2tk+-transformed
cell lines:
39A+
bandsfrom the1and 10copy per-cell reconstruction cells, producedby
exposureof tk-
cells
toUV
experinents
light-irradiated HSV-2 (333) and
59D+
and
Silow
Restriction endonu-
Fra
tIntensity 10/inten-
cells, produced by
infection of
tk- cells with UV
clease
agmen
sity 1"
light-irradiated HSV-2 (324) and HSV-2 (Silow),
KpnI
A 8.3respectively.
The results of these
mapping
ex-B
periments
(Fig. 7) revealed the
following:
39A+
c
cells
contained
acontiguous
setof
viral
DNAD
16.44
sequences located between map coordinates 0.14
and
0.42.Silow cells contained
acontiguous
E
10.08
stretch of viral DNA sequences mapping
be-tween
coordinates
0.21 and 0.32.
Finally, 59D+
F
8.43
cells
contained
acontiguous
setof
viral
se-G
12.3
quencesspanning
coordinates 0.28 to 0.42.
H
In addition to the
viral DNA sequences
de-I827
scribed
above,
cell lines
39A+,
59D+,
and
Silow
I*8.27
all
contained
two setsof viral DNA sequences
J
8.86
which
werepresent
in
significantly
lower
abun-dances (less than 0.1 copy per cell). These
low-L
8.03
abundance
viral DNA sequences have been
ten-M
tatively mapped in
twoseparate
regions
of the
viral genome located
between map coordinates
N
7.89
0.06 to 0.20 and 0.82
to 0.93.
o
Acomparison of the map
positions of the viral
p
12.2
DNA sequences present in the four
HSV-2tk+
transformants
(Fig.
6and
7) reveals that the
only region of the HSV-2 genome which is
com-EcoRI/HpaI
A 7.99mon to all four cell
lines
is located between map
B
coordinates
0.28
and 0.32.
Thus, this region of
C
the
HSV-2 genome
mustcontain the HSV-2 tk
D 9.03 gene.
E
Mapping
of the HSV-1 DNA sequences
F
9.94
presentin
twoHSV-ltk'-transformed
cell
lines. Cell lines 5A and 8N
wereproduced
by
G13.91
transfection
of L tk-
CllD
mouse
cells
with
HSV-1
(1023) DNA which was
randomly
H
8.54
sheared to fragments of molecular weight 20 x
I 10.55
106 to 30
x106. DNAs from
passage 15 of these
J
cell lines
werepreselected
and
hybridized,
asdescribed
above,
tonitrocellulose
blots
contain-L
9.27
ing various restriction enzyme
fragments
of
M
HSV-1 DNA.
Autoradiograms
of the
hybridized
blots from these
experiments (Fig. 8 and 9)
re-N8.84
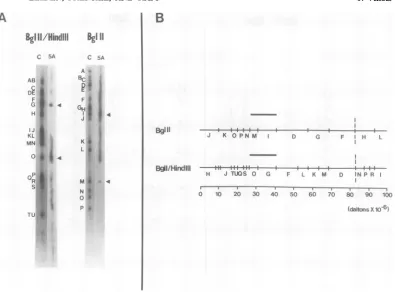
vealed that (i)
cell
line 5A contains a contiguous
O
9.45
setof HSV-1
DNAsequences
mapping between
coordinates
0.27and
0.41, and
(ii) cell
line
8N
P
20.74
contained four sets of
noncontiguous L region
viral
DNAsequences
mapping between
coordi-Q
21.96
nates0.11
to0.17,
0.29 to0.32,
0.34to 0.40, and
R10.0
0.52 to 0.56. In
addition,
as seenin
Fig. 9,
8N
s DNA
hybridized
to theBglII/HindIII
fragment
'Ratios of theintensities of the
corresponding bands (as
Q of HSV-1(Justin)
DNA,
and to theBglII/
determiined bydensitometric scanning as described in the HindIIIfragment R,
but not to theBglll/
text)of the 10 and 1copy per cell reconstruction experiments HindIH fragment P, of HSV-1 (F) DNA.
More-shown inFig. 1. The5-day autoradiographic exposures were
over,
it hybridized tothe0.5 M fragmentsfromused for the densitometric
scanning,
the
right
sideof the S
region of HSV-1
DNA moreefficiently than to those from the left side
the
map
position
ofthe tk
gene
ofvarious strains
of the S region.
Therefore, we have concluded
ofHSV-2,
wemapped
the viralDNA
sequences
that this
cell
contains a contiguous set of S
which arepresent
inlate
passages
ofthree
ad-
region DNA
sequences, composed of the
U.
se-33,
on November 10, 2019 by guest
http://jvi.asm.org/
LEIDEN, J. VIROL.
Kpn I
LcoKI
/IHpaI
A B c A P
;r:'A + :5, C R33A4C
fE*<w <
.oR9b0.:*k
DPt~~~~~~~~~~~~~~~~~
F* w .. Z~~DDE g t s v *w
**@4tW~ ~ ~ ~ ~ J
N
K *
N X 4
F:.~ ~ ~ ~ ~ .
6 7 .
33A
HSV-2(G)EcoRIfipal LIN 4FT+ -- t M+--
-G FTSJ B M.
HSV-2(G)KpnI - 4+ Ht it ---
-F K LPO H NOJ M 0 D0
MW , -I-.,---r--- --.
0 10 20 30 40 5 50 *'A%
ft4hC,
FIG. 6. Mapping ofthe HSV-2 DNA sequences present in the
HSV-2tkt
cellline 33A . Top: control DNA(purifiedin vitro-labeled HSV-2[G] DNA) (C)orin vitro-labeled33A+ DNA(33A+) whichwasprocessed
through thepreselection steps washybridized tonitrocellulose strips containing either the KpnI, orthe
EcoRI/HpaI restriction enzyme fragments of HSV-2(G) DNA. A, B, and C, Autoradiograms from three separatehybridizationexperiments,eachdone withaseparatepreparationof 33A + DNA (passages50-70of thecellline)andaseparatesetofnitrocellulose blots. InexperimentCa1copy percellreconstruction mixture (seelegendtoFig. 4)washybridizedto areplicate nitrocellulose strip (R).Inexperiment A,autoradiography
wasperformed for either 5 days
(panels
2 and 10) or 3 weeks (panels 3 and 11). Bottom: schematic representation ofthe regions of homology between 33A+ DNA andthe KpnI, or EcoRI/HpaIrestriction enzymefragments ofHSV-2(G)DNA. The restrictionenzyme mapsweretakenfrom G. S.Hayward, T. G. Buchman, andB.Roizman(unpublisheddata).The dotted line represents aregionof uncertain homology, duetothecomigration oftheKpnIfragmentsLand M.quences
mapping
betweencoordinates
0.91and present in sixHSVtk+-transformed cell lines.
0.96and
aportion of
theacsequences.Whereas
thismethod
requires
relatively
small
DISCUSSION
amounts oftransformed-cell
DNA,
itpossessessufficient
sensitivity
todetect
a small piece of In thestudies
reported
inthis
paper, wehave
HSV
DNA which ispresent in an averageabun-described
anovel
mapping
approach
which hasdance
of1 copy percell.
Thus, this
procedureallowed
thesimultaneous detection
andidenti-
could
beapplied
instudies
ofcells
transformed
ficationof
theHSV
DNA sequences which areby
other virusescontaining
genomes whichon November 10, 2019 by guest
http://jvi.asm.org/
[image:9.504.104.408.70.465.2]EcoRI/
Hpal
Kpnil
C Silow 39A+ 59D+ R R C Siow39A4 C59D R R BA
F F
F 4 H
H1 2 4301G 21
K
LM'a'
N K
O LM *
P~ ~ ~ ~ ~ ~ ~~P
T R~~~~~~~~~~~~
1 2 3 4 5 6 7 8 9 10 11 1213
-ASIow
39A+
590+
EcoRI/lpal
i- i Hi -i--- i i i H"s ^ C rCe I a u I D e I u D^O
KpnlI
L N G r- IS J C MV IMr In11UR Q
-Sibw
39A4
- D59D+
F K LPO H NQJ M G D I S C 1R1 A R2
,.. -- ---_,-
--~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
0 10 20 30 40 50 60 70 80 90 100
(daetonX10o6)
FIG. 7. Mapping ofthe HSV-2 DNAsequences present in theHSV-2tk+ celllines Silow,
39At,
and59Dt.
Top: control[purifiedin vitro-labeledHSV-2(G)DNA](panels 1, 5, and 8)orin vitro-labeledSilow (panels 2and6), 39A+(panels3and7), and 59D+(panels4and9)transformed-cellDNAs, whichwerepreselected to enrichforviral DNA sequences,werehybridizedtonitrocellulose blotscontaining either the KpnIor
EcoRII
HpaIrestriction enzymefragments of HSV-2(G)DNA.Autoradiographywascarriedoutfor 4 days. Bottom: schematicrepresentationoftheregionsof homologybetweenSilow, 39A+,and59D+ DNAs and the KpnIorEcoRI/HpaIrestriction enzymefragmentsofHSV-2(G) DNA. The restriction enzyme mapsweretakenfrom G. S. Hayward, T. G. Buchman, and B. Roizman (unpublished data). Dotted lines representaregion of uncertainhomology,duetocomigration of the KpnIfragmentsLand M. It should be noted that as described in thetextallthreeHSVtk+cellDNAsshowed lowintensityhybridizationtofragmentslocated between map coordinates 0.06 to0.20and0.82 to 0.93.Becauseof their extremely low abundances, these regions are not displayed in the bottom partofthefigure.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.504.104.396.70.541.2]282 LEIDEN, FRENKEL, AND RAPP
A
BgIII/HindIII
Bgl
11
B
Bglll
Bgll/Hlirlclll
--+I-- t4$t-s--- i---- t - t
J K 0PN M D G F H
H J TUOS 0 G F L K M D IN P R
0 10 20 30 40 50 60 70 830 90 100C
(dialtonsX
FIG. 8. Mapping ofthe HSV-1 DNA sequencespresent in theHSV-ltk+ cell line 5A. A, Control DNA
(purifiedin vitro-labeled HSV-1(F)DNA(C)orin vitro-labeled 5ADNA, whichwaspreselectedasdescribed
in thetext, washybridized toreplicate nitrocellulose blotscontainingeither theBglIIrestriction enzyme
fragments of HSV-1(MP) DNA, orthe BglII/HindIII restriction enzymefragments of HSV-I(F) DNA. B,
Schematic representation ofthe regions ofhomology between 5A DNA andtheBglII and BglII/HindIII
restrictionenzymefragments ofHSV-I DNA. The restrictionenzyme maps weretaken from G. S.Hayward,
T. G.Buchman,and B. Roizman(unpublished data).
play buoyant densities different from those of
host cellular DNA. On the other hand, this
approach suffers from two distinct disadvan-tages. First, ityieldsnoinformationconcerning theintegrationorarrangementof the viral DNA sequenceswithin the transformedcells.Second, theincorporationof the
CsCl
gradient preselec-tion step could result in the failure to detect small stretches of integrated viral DNA se-quences. However, theoretical considerationshave indicated that theshearing and
centrifu-gation conditionsused in the studiesreportedin
thispaperwould haveallowed the detectionand
mappingofpiecesofintegratedviral DNA with
molecularweights greaterthan 3 x 106 (i.e.,3%
of the HSVgenome).
Mappingof theHSV-2 tkgene. Themaps
shown in
Fig.
10 indicate that the sequenceslocated betweenmapcoordinates 0.28and 0.32 are the only viral DNA sequences which are common to all of the HSV-2tk+-transformed
cells.Similarmappingstudies of the HSVDNA sequences which are present in six
HSV-ltk+
transformants (including the cell lines 5A and
8N discussedin thispaper,andfour additional HSV-ltk+-transformed cell lines which will be discussed elsewhere) have shown that the se-quenceslocated between mapcoordinates0.28
and 0.32arealso theonly viral DNAsequences common to all HSV-ltk+-transformed cells.
Therefore, both the HSV-1 and HSV-2 tkgenes arelocated between map coordinates 0.28 and
0.32, and this location does not vary between
viral strains. These results are in accord with
thefindings of Morseetal. (25). However, they
areindisagreement with the results of Maitland andMcDougall (21).
Non-tk gene viral DNA sequences
pres-ent in HSVtk+ cells. Inaddition to the viral DNA sequences located between map coordi-nates 0.28 and 0.32, all of the
HSVtk+-trans-formed cell lineswhichwehave tested contain a substantial, but variable, amount of non-tk
geneviral DNA. Severalpointsarenoteworthy
concerning the possible expression and
func-tion(s) of these viral DNA sequences. First, if
C 5A
AB
ID
F
KL MN
p RS
TU
C 5A A
F t
GH l
J 4
.: 4
K6.
L f
*::
NJd oh P -i
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.504.67.462.62.354.2]F Jus BglII/Hindill BglII/Hirdill
CBN
| A . I
I
Gf .s.A<
1-
N1
[image:12.504.50.444.70.328.2] [image:12.504.47.239.463.574.2]-4 0o* <
|<ST st*
U
1 2 3 4 5 6
Jus
KmI
C SN
7 8
B
(MP)Bgl11
(F) BglII/Hindill
(Jus)BglIl/Hindill
(Jus)Kpn
J K OPNM D G F H L
-8tt -144t4}tt1---l-i---F-1$ H JTUOSO G F L K M D IN PR
_.j-^P4 --*-.
-+-H U J STPRVO G F M L N E 0 K
RUG Z E aWH MSNPv.X B LC' A YT J OUR: F K AA
0 10 20 30 40 50 60 70 so 90 10(0o
FIG. 9. Mapping ofthe HSV-1 DNA sequencespresent in the
HSV-ltk'
cell line8N. A, ControlDNA(purifiedinvitro-labeled HSV-1DNA)orinvitro-labeled 8NDNA,whichwaspreselectedto enrichforviral DNA sequences, was hybridized to replicate nitrocellulose blots containing various restriction enzyme
fragmentsofDNAfromHSV-1 strainsMP,F,and Justin(Jus). KpnIfragmentsR1 andR2areboth derived
fromthe endsofthe Lregionandreflecttheheterogeneity ofthe viral DNAsequencesin thisregion ofthe
HSV-1genome(35;H.Locker andN.Frenkel, manuscriptinpreparation). B,Schematicrepresentation ofthe
regions ofhomologybetween 8N DNAandtheBglII, BglII/HindIII,andKpnIrestrictionenzymefragments ofHSV-1 DNA. TheBglIIandBgIII/HindIIIrestrictionenzymemapsweretakenfromG.S.Hayward,T.G. Buchman,andB. Roizman(unpublished data). TheKpnIrestrictionenzyme mapwastakenfromH.Locker
and N. Frenkel(manuscriptinpreparation).Dotted linesrepresentregions ofuncertainhomology,whichare
presumablyduetothepresenceofthe invertedrepeatacsequencesin8N DNA(seetext).
Sequencearrangwnwit
1-a L_
s-J-ab Lk dAikhc
0 0.1 CQ2 Q3 0.4 Q5 06 0.7 08 Q.9 1.0
r . , . . .
0 Q W 0.4 0I 0I 0. OI Q ID
I~~~~~~~~~~~~~~~~~~~
0.1
liQ
0 S 6 0. S 9 1FIG. 10. Summary ofthemapping data oftheHSV DNA sequences present in the six HSVtk+-trans-formedcel lines.Thedottedlinesrepresentaregion
ofuncertainhomology (see legendtoFig. 6).
thesesequences encodeanyoftheknownlytic
functions ofHSV, the expression of these lytic functionsmustbesuppressed inthe
transfonned
cells. Second, it has been previously suggested
that some of these non-tk gene viral DNA se-quences may exert a positiveregulatory effect on viral tk gene expression in these cells and may,
therefore,
bepositively selectedforduring propagation of the cells inHAT medium (17). Thispossibility currentlyseemsunlikely in lightof the fact that none ofthe
HSVtk+
cell lines tested in this study contain the viral DNA se-quences which are known to encode the early (a)viralgenes.The results shown in Fig. 10, when taken together with the mapping data of four addi-tional
HSV-ltk+
transformantstobepresentedelsewhere,
havealso revealedthat therearetworegionsoftheviralgenomewhichappeartobe uniformly absent frommost,ifnotall, HSVtk+ transformants. These regions are located be-tween map coordinates 0 to 0.06, and 0.57 to
0.82. One can envision at leasttwo alternative explanations for the uniform absence of these
1980
A
MP Bgl 11
C BN
K
I
,:
A
I C BN
0
q.
F GHi
A L MN
1o
p
IQ
s
T U
A
R
RII
4
TV v wx yi
(daltonsX10-6)
on November 10, 2019 by guest
http://jvi.asm.org/
284
LEIDEN, FRENKEL, AND RAPPviral
DNA sequences. First, these sequences maycontain
nonsuppressible
viral lyticfunc-tions. Thus,
anycell
originally containing
themwould be unable
tosurvive. In
this
light it
isinteresting
to notethat
Morse etal.
(25) havemapped
ahost
shutoff function
betweencoor-dinates
0.52and
0.59 on the HSV-2 genome.Alternatively, the
sequences which areuni-formly deleted from the transformed cells
couldcontain
early (a) viral functions
which areneeded
to turn onthe expression of lytic
viralfunctions
located elsewhere
onthe
genome (11).Thus the absence of these regions
could assurethe lack ofexpression of late viral lytic
functions.
This
possibility is especially attractive in
view of recentstudies which have indicated that
someof
the
early viral
genes arelocated within the
inverted
repeatregions of the L
componentof
HSV DNA
aswell
asbetween
mapcoordinates
0.56and
0.79(3,13,29;
L.Morse and B.
Roizman,
unpublished data).
Patterns
of incorporation of HSV DNA
during
HSV-mediated tk
transformation.
Our
mapping studies have revealed that
someofthe
HSV-2tk+ cell lines
whichwehave examined
(cell lines 39A+, 59D+, and
Silow)
contain viral
DNA
sequenceswhich
werepresentin
verylow
abundance. One
canenvision
atleast
twounder-lying mechanisms which could
generatethese
observed abundance
pattems.First,
it is
possible
that
each of these cell lines
representsthe
prod-uctof
several
independent
transformation
events(of
several different
cells),
allof which
resulted in the
incorporation of the tk
gene,but
each of which involved the
incorporation
of
adifferent
setof
contiguous
non-tk
geneviral
DNA
sequences.This is
possible given
that the
HSV-2tk+ cell lines used
inthese studies
werenot
extensively
cloned.
Alternatively,
it is
pos-sible that these
celllines
mayhave been the
product of
asingle
transformation
event,but
that
following
the
initial
cloning
in
HATme-dium there
was aperiod
of loss of non-tk
geneviral DNA
sequencesfrom selected cells within
the
originally
uniform
population.
These
possi-bilities
arecurrently
being
differentiated
by
mapping
studies of the DNA from subclones oflate
passages of thesecell
lines.Finally,
itshould also
benoted that
most, ifnot
all,
of
theUV
light-irradiated
HSV-produced
cell lines
appeartocontain
asingle
contiguous
sequence
of HSV
DNA,
whereas 8Ncells which
wereproduced
by transfection of tk- cells
withsheared
HSV DNAclearly
containalarge
num-ber of
noncontiguous
HSV
DNAsequences. This differencemight reflect
the fact that DNAtrans-fections result
in theincorporation
of
multiple
DNA
fragments
into thesamecell.
Moreover,
thefinding
of themultiple
noncontiguous
setsofHSV
DNA sequences in8N
cells
indicates thatthe
incorporation of viral
DNAfragments during
HSV-mediated tk transformation is
a relativelyrandom
process,i.e., that
any pieceof viral
DNAwhich does
notencode
lytic HSV functions
canbe
incorporated into the cells during
HSV-me-diated tk transformation.
ACKNOWLEDGMENTS
We thank B. Burckart forskillfullygrowing thecells and viruses used inthese studies, H. Locker and B. Roizman for useful discussions, N.Turnerforperformingthe tk assays, and D.Polacek and B. Roizman for providing the 8N and5A cell lines.
The work performed at the University of Chicago was
supported byPublic HealthServiceresearch grants AI-15488
and CA-19264 from the National Institutes of Health, and by National Science Foundation grant PCM78-16298. J.M.L. is a
predoctoral fellow supportedby Public Health Service grant
5-T32 HD-07009-03.
The workperformedattheHershey Medical Center was supported by contractNO1CP 53516 within the Virus Cancer Program of the National Cancer Institute and grant CA 18450 awarded by the National Cancer Institute. A preliminary report of thesedata waspresented in the Herpesvirus Work-shop,Cambridge,England, August 1978.
LITERATURE CITED
1. Bacchetti, S., and F. Graham. 1977. Transfer of the gene forthymidinekinase tothymidinekinase-deficient human celis by purified herpes simplex viral DNA. Proc.Natl. Acad. Sci.U.S.A.74:1590-1594.
2. Baker,R. F. 1977.Bindingof DNA tocellulose nitrate
filtersunder denaturingconditions. Analyt. Biochem. 78:569-571.
3. Clements,J.B., R. J. Watson, and N. M.Wilkie.1977.
Temporal regulation of herpes simplexvirus type 1
transcription: location of transcripts on the viral ge-nome.Cell 12:275-285.
4. Davidson, R., S. Adelstein, and M. Oxman. 1973. Herpessimplexvirusasa sourceofthymidine kinase
forthymidinekinasedeficient mouse cells: suppression
and reactivation of the viral enzyme. Proc. Natl. Acad. Sci. U.S.A. 70:1912-1916.
5. Denhardt,D.T. 1966.Amembrane filter technique for
the detection of complementary DNA. Biochem.
Bio-phys.Res.Comm.23:641-646.
6. Dubbs,D.R.,andS. Kit.1964.Mutant strains ofherpes
simplexdeficient inthymidine kinase-inducing activity.
Virology22:493-502.
7. Ga.rfinkle, B., and B. McAuslan. 1974.Regulation of
herpessimplex virus induced thymidine kinase.
Bio-chem.Biophys. Res.Commun.58:822-829.
8. Graham,F.L.,A. J. Van derEb,and H.L.Heijneker. 1974. Size and location of the transforming region in human adenovirustype5 DNA.Nature251:687-691.
9. Groneberg,J., Y.Chardonnet,and W.Doerfler. 1977.
Integrated viral sequences in adenovirus type 12 trans-formed hamster cells. Cell 10:101-111.
10. Hayward,G.1974.Uniquedouble-strandedfragmentsof
bacteriophage T5 DNA resulting from preferential
shearinducedbreakage ofnicks.Proc.Natl.Acad. Sci. U.S.A.39:679-701.
11.Honess, R. W., and B. Roizman. 1975. Regulation of herpesvirus macromolecular synthesis. Sequential tran-sition ofpolypeptidesynthesis requires frunctional viral polypeptides. Proc. Natl. Acad. Sci. U.S.A. 72:1276-1295.
12.Hotta, Y., and A. Bassel. 1965. Molecular size and
circularity of DNA in cells ofmammals and higher
plants. Proc. Natl. Acad. Sci. U.S.A. 53:356-362.
on November 10, 2019 by guest
http://jvi.asm.org/
13. Jones,P.C.,G.S. Hayward,and B.Roizman. 1977. Anatomyof herpes simplex virus DNA. VII. aRNA is homologous to non-contiguous sites in both L and S componentsof viral DNA. J. Virol. 21:268-276.
14. Kieff,E.D., S.Bachenheimer, andB.Roizman. 1971.
Size, composition and structure of the DNA of subtypes 1and 2herpes simplex virus. J. Virol. 8:125-132. 15. Kit,S.,and D. R. Dubbs.1963.Acquisitionofthymidine
kinaseactivity by herpes simplex infected mouse fibro-blastcells. Biochem. Biophys. Res. Commun. 11:55-59.
16. Klemperer,H.G., G. R. Haynes, W. I. H. Shedden,
and D. H. Watson. 1967. A virus specific thymidine kinase in BHK21 cells infected with herpes simplex virus.Virology 31:120-128.
17.Leiden, J.,R. Buttyan, and P. G. Spear. 1976. Herpes
simplex virus gene expression in transformedcells. I.
Regulation of viral thymidine kinase gene in trans-formed Lcells by products of superinfecting virus. J. Virol. 20:413-424.
18. Leung, W. C. 1978. Evidence for a herpes simplex virus-specific factorcontrolling the transcription of deoxy-pyrimidinekinase. J. Virol. 27:269-274.
19. Lin, S.-S. and W. Munyon. 1974. Expression of the viral thymidine kinase gene in herpes simplex virus trans-formed L cells. J. Virol. 14:1199-1208.
20.Littlefield, J. 1964. Selection of hybrids frommatingsof fibroblasts in vitro and their presumed recombinants. Science 145:709-710.
21. Maitland, N. J., and J. K. McDougall. 1977. Biochem-ical transformation of mouse cells by fragments of herpes simplex virus DNA.Cell11:233-241.
22. Maniatis, T., A. Jeffrey, and D. G. Kleid. 1975. Nu-cleotide sequences of the rightward operator of phage
A.Proc. Natl. Acad.Sci. U.S.A. 72:1184-1188. 23. Minson, A.C.,P.Wildy, A. Buchan, and G. Darby.
1978.Introduction of the herpessimplexvirus thymi-dine kinase gene into mouse cells using virus DNA or transformedcellDNA.Cell13:581-587.
24. Morse,L.S.,T.G.Buchman, B. Roizman, and P. A.
Schaffer. 1977. Anatomy of herpes simplex virus DNA. IX. Apparentexclusion of some parental DNA arrange-mentsin the generation ofintertypic (HSV-1 x HSV-2) recombinants. J. Virol. 24:231-248.
25. Morse,L.S.,L.Pereira, B. Roizman, and P. A.
Schaf-fer.1978.Anatomy of HSV DNA. X. Mapping of viral genesby analysis of polypeptides and functions
speci-fiedby HSV-1xHSV-2 recombinants. J. Virol. 26:389-410.
26. Munyon, W.,R.Buchsbaum,E.Paoletti,J.Mann,E.
Kraiselburd, and D. Davis. 1972.Electrophoresis of thymidine kinase activity synthesized by cells trans-formedbyherpes simplex virus.Virology 49:683-689.
27. Munyon, W.,E.Kraiselburd,D.Davis,and J. Mann.
1971. Transferofthymidinekinasetothymidine kinase-less L cells by infection with ultraviolet-irradiated herpessimplex virus. J. Virol. 7:813-820.
28. Ogino, T., R. Shiman, and F. Rapp. 1973. Deoxythy-midine kinase from rabbitkidney cellsinfected with herpessimplex virus types1and2.Intervirology 1:80-95.
29.Preston, V. G., A. J.Davison, H. S.Marsden,M. C. Timbury, J. H.Subak-Sharpe, and N. M. Wilkie. 1978.Recombinants betweenherpessimplexvirustypes 1and 2:analyses of genomestructuresandexpression of immediateearly polypeptides.J.Virol. 28:499-517.
30. Rapp, F., and N. Turner. 1978. Biochemical
transfor-mation ofmousecellsby herpessimplexvirus types1
and2:comparisonof different methods for inactivation
of viruses. Arch. Virol. 56:77-87.
31. Roizman, B. 1979. The structure and isomerization of
herpessimplex virus genomes.Cell 16:481-494.
32. Ruyechan,W.T., L.S.Morse,D. M.Knipe,and B.
Roizman.1979.Moleculargenetics ofherpes simplex
virus.II. Mapping of the major viralglycoproteinsand ofthe genetic loci specifying the social behavior of infected cells. J. Virol. 29:677-697.
33. Southern, E. M. 1975. Detection ofspecificsequences among DNAfragmentsseparatedby gel
electrophore-sis. J.Mol. Biol.98:503-517.
34. Thouless, M. and P.Wildy.1975.Deoxypyrimidine ki-nasesof herpessimplexvirusestype1and2:comparison ofserological and structural properties. J. Gen. Virol. 26:159-170.
35. Wagner, M. J., and W. C. Summers.1978.Structure of thejoint S region and the termini oftheDNA ofherpes
simplexvirustype1.J. Virol. 27:374-387.
36. Wetmur, J. G. 1975.Acceleration of DNArenaturation rates.Biopolymers14:2517-2524.
37. Wigler, M., S. Silverstein, L. Lee, A. Pellicer, Y.
Cheng,and R. Axel.1977.Transfer ofpurified herpes
virusthymidine kinase gene tocultured mousecells. Cell11:223-232.