0022-538X/85/110558-13$02.00/0
Copyright© 1985,AmericanSociety for Microbiology
Isolation and Characterization of Deletion Mutants of Herpes
Simplex Virus Type 1 in the Gene Encoding Immediate-Early
Regulatory Protein ICP4
NEAL A. DELUCA, ALICE M. McCARTHY, AND PRISCILLA A. SCHAFFER*
Laboratory of Tumor Virus Genetics, Dana-FarberCancer Institute, andDepartmentof Microbiology and Molecular Genetics, Harvard MedicalSchool, Boston, Massachusetts 02115
Received 3 June 1985/Accepted 7 August 1985
Using Vero cellstranformedwiththe wild-typegenefor ICP4asthe permissive host cell,weisolatedherpes
simplex virustype 1(HSV-1)mutantscontaining deletions in both copies of the ICP4gene.Themutants,d120
and d202, contained deletions of 4.1 and 0.5 kilobases, respectively, in each copy ofICP4. ICP4 mRNA
synthesized in d202-infected Vero cellswas0.5kilobasessmaller than that synthesized in cellsinfected withthe
wild-typevirus. No ICP4 mRNAwasdetectedind120-infectedVero cells.d120andd202 specifiedpolypeptides
that reacted with ICP4 antiserum and were smaller than the wild-type ICP4 polypeptide by 130 and 30
kilodaltons, respectively. The only other HSV-1geneproducts detectableoninfection ofVero cells with d120 and d202 were ICP6 (of the early kinetic class of HSV-1 polypeptides), ICPO (immediate early), ICP22 (immediate early), and ICP27 (immediate early). Immediate-early polypeptides ICPO and ICP27 were
expressedtoahigherlevel inVerocellsinfectedwithanICP4 temperature-sensitive(ts)mutant(tsB32)at39°C, indicating immediate-early stimulatory activity associated with the ts ICP4 polypeptide. In addition, the patternsof complementation of d120, d202,andtsB32inICP4-transformed cells alsodemonstratedinhibitory activityassociatedwith the tsformof theICP4 polypeptide.
The viralgenes expressed initially afterinfection of sus-ceptible cells withherpessimplex virustype 1(HSV-1)have been
designated
immediate-early (or a) genes (3, 19). These genes are operationally defined as those transcribed in the absence of prior viral protein synthesis (3, 19). Fiveimme-diate-early
mRNA species are translated into the polypeptides ICP0, ICP4, ICP22, ICP27,andICP47(19, 50). Functional immediate-early polypeptides are required for efficient expression ofearly(3)
orlate (y) viralgenes(20).The functional roles of individual immediate-early gene products in the regulated expression of HSV-1 genes have been examined by several methods. First,
temnperature-sensitive (ts)mutantsof HSV-1 containing mutations in the genesfor ICP4 and1CP27
havebeenisolatedand character-ized (11, 35, 37, 40). Phenotypic analysis ofthese mutants hasshown thatICP4andICP27perform essentialreplicative functionsand are involvedspecificallyinmodulatingHSV-1 gene expression (11, 36, 40, 49). Second, reconstruction experiments involvingcotransfection of plasmids containing the immediate-early genes for ICP4 and ICP0 and suitabletestgenesindicate that both stimulateexpression ofHSV-1
early
genesin trans(12, 30,39). Finally, deletionmutantsin the gene for ICP22 have been shown to be capable of growing in Vero cells, a cell line which is routinely usedas alaboratoryhost for
HSV.
Theinvolvement ofthe immediate-early polypeptides ICP22and ICP47 in HSV-1 gene expres-sion, however, remains obscure.The most extensively studied immediate-early gene of HSV-1 is thatencodingthe
175-kilodalton
polypeptideICP4. Alarge numberof
temperature-sensitivemutantsin the gene forICP4 exist.Collectively, theydefine HSV-1 complemen-tationgroup1-2
(43). Acommon characteristic ofts mutantsinthisgroup isthattheyinducedramatic overproduction of immediate-early polypeptides atthe nonpermissive
temper-*Correspondingauthor.
ature. Based onthis observation and the resultsof temper-ature-shift experiments, it has beenpostulated thatICP4 is
required for negative regulationorattenuated expression of immediate-early genes (11, 36). Other features of cells in-fected with most ICP4 ts mutants at the nonpermissive temperatureareunderrepresentationorabsenceof early and late polypeptides and absence ofviral DNA (11, 36). The basis for thesephenotypic propertiesis thatwild-type ICP4 is
required
for the accumulation ofstable transcripts from early and late genes, and the products of early genes arerequired for viralDNA synthesis. Asubset oftsmutantsin group1-2inducethesynthesis of appreciable levels of early polypeptides and viral DNAatthe nonpermissive
tempera-ture(8). The absence of detectable infectious virus in such mutantsis attributable to theabsence of late polypeptides. Basedonthecumulativeevidence, itisthoughtthatICP4is (i) required for the expression ofearly and late genes, (ii) autoregulatory,and(iii)involvedinthe inhibition of
expres-sionofotherimmediate-earlygenes.Furthermore,it appears thatthemechanismof actionof ICP4 involves modulationof gene expression at the level of accumulation of stable transcripts.
Asjust stated, ourcurrentunderstanding oftheactivities of andrequirementsfor
wild-type
ICP4 is basedprimarilyonthe phenotypic properties of ICP4 ts mutants at the nonpermissive temperature. The conclusions drawn from studies ofthesemutants rest in partonthe
assumption
that the temperature-sensitive forms of the ICP4 polypeptide havenoinfluenceonHSV-1 geneexpression.This may not, infact, be thecase. Recently, it has been shown that ICP0can actintrans tostimulateexpressionfrom
early
promoters inacotransfectionassay(30, 39).This
presehts an interest-ing paradox in that most ICP4 ts mutants overproduce immediate-earlypolypeptides (includingICPO)
and yet failto expressearlypolypeptides.
Therefore,eithercotransfection experiments donotreflectan eventoccurring in viral infec-558on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 DELETION MUTANTS IN THE ICP4 GENE 559 TABLE 1. Growth of HSV-1 ts mutants on 2-6 and Vero cells
Complement- Defective Virus grownona
Vlrusationgroup gene 2-6 Cells Vero cells
KOS 8.0 x 106 3.7 x 106
tsB32 1-2 ICP4 7.8 x 105 2.1 x 102
tsB21 1-2 ICP4 6.0 x 104 5.0 x 100
tsL14 1-2 ICP4 1.1 x 105 1.7 x 102
tsY46 1-5 ICP27 2.5 x 101 1.0 x 101
tsJ12 1-9 gB 4.0 x 102 3.6 x 102
tsA24 1-1 ICP8 3.0 x 101 3.0 x 101
a2-6 and Vero cellswereinfected with the indicated virusmultiplicity of infection of 1.0 PFU per cell and incubated at39'C for 18 h. The cultures were then harvested and assayed on Vero cells at34'C.
tion or, more interestingly, the tsform oftheICP4 polypep-tide has aninhibitoryeffect onearly gene expressionor the transactivity of ICPOorboth. Inaddition,theisolation ofts mutants in the structural gene for ICP4 which exhibit dis-tinctly different regulatory phenotypes (8) suggests that the ts form(s) of ICP4 may exhibit activities capable of modu-lating gene expression to a greater or lesser degree.
To betterdefinetheroleofICP4in the lifecycle ofHSV-1 and to answer questions regarding the possible regulatory activities oftsforms ofthisprotein on HSV-1 gene expres-sion, we isolated deletion mutants ofHSV-1 which induce the synthesis of truncated forms ofthe ICP4 polypeptide. BecauseICP4performsanessentialreplicative function,we developed a system for screening and propagating ICP4 deletion mutants by constructing cell lines expressing the wild-type ICP4 gene. Such a system has proven invaluable forthestudy of deletionmutantsin themajortranscriptional regulatory protein of adenovirus, EIA (22). Although cells transformedwith HSV-1 genes have been shown to comple-ment ts mutants(7, 41), suchcellshave notbeenusedtodate to isolate deletion mutantsofHSV-1. In the present study, twoICP4 deletionmutants wereisolated andused to address thefollowing questions. (i)What is thepolypeptide pheno-typeof cells infected witha mutantlackingafull-length ICP4 polypeptide, and (ii) doesthisphenotype differ fromthat of ts mutant-infected cellsinwhichafull-sizeyet temperature-sensitiveform oftheICP4 polypeptideis produced?
(kb) Saclfragment (Fig. 1) to yield pKX2-ASacI. pKX2-P4 was constructedby insertion of the 220-base-pair EcoRI-to-SmaI fragment(-110 to -330[Fig. 1]) into the EcoRI site of pKX2 in the proper orientation. TheSmaI site at -330 was modified with EcoRI linkers. DNA-modifying enzymes and EcoRI linkers were obtained from New England BioLabs, Inc. (Beverly, Mass.) and used as prescribed by the manu-facturer.
Nucleic acid isolation. Purified HSV-1 (14) and bacterial plasmid DNAs were isolated as described previously (8). Cytoplasmic RNA wasisolated from Nonidet P-40 extracts of Vero cells free ofnuclei,as previously described (24).
Transformation of Vero cells. pSV2neo confers resistance to the antibiotic G418 (geneticin; GIBCO Laboratories, Grand Island,N.Y.)whenintegrated intocultured mamma-lian cells (46).Vero cells were cotransformed with pSV2neo and HSV-1 ICP4 as follows. pSV2neo (0.5 p.g) was coprecipitatedwith 5.0 ,ug ofpKXCin thepresence of 15 ,ug ofsalmon testes DNA(SigmaChemicalCo.,St. Louis,Mo.) in a total volumeof1.0 mlbytheprocedure ofGraham and van der Eb (16). Freshly trypsinized Vero cells (4 x
106)
were added to the precipitate and incubated at 370C with continuous shakingfor 30 min. The cell-DNA mixture was suspended in20mlof culturemedium andtransferred to two 85-mm petri dishes. After a 4-h incubation at 37°C, theadsorbed cells were subjected to glycerol (15%) shock and incubated for an additional 2 days at 37°C. The cells were then trypsinized and seeded into 85-mm petri dishes at a densityof 5 x
103
cellsper cm2. After the cells had adsorbed to the dish, themedium was removed and medium contain-ing1.0mgof G418per ml wasadded.After 5 to 10days,the G418concentrationwaslowered to 400,ug/ml.After 12 to 18 days, individual G418-resistant colonies were isolated, am-plified, and subsequently screened for ability to support replication of tsB32 at 39°C in a plaque assay. Approxi-mately20% oftheG418-resistantclonesexhibited detectable complementing activity. One cellline, 2-6,was retained for further study.Cell lines 1-2, 1-5, and 4-2 were generated in a similar manner, except that pKX2-P4 rather than pKXC and d120 and d202 were used to generate and screen G418-resistant clones, respectively. Among
pKX2-P4-transformed
cells,MATERIALS ANDMETHODS
Cells and viruses. Procedures for the growth and maintenance of African green monkey kidney cells (Vero, ATCC P130; CV-1, ATCC P27) were previously described (51). Nero cells are an unclonedpopulation of G418-resistant Vero cells generated bytransformation withpSV2neo.
Thewild-type KOS strain ofHSV-1 and theKOS-derived temperature-sensitive mutants (Table 1) were propagated and assayed as previously described (8). tsB32, tsB21, and tsL14 are mutants in the structural gene for ICP4 (8, 11).
Plasmids. pSV2neo contains the bacterial gene for neomycin resistance under control of the simian virus 40 early promoter (46). pKXC (Fig. 1) contains the XhoI C fragment of KOS and was constructed as
described
previ-ously(10). pKBZandpKX2(pKEB-X2) are diagrammedin Fig. 1and were constructed asdescribedpreviously (8).Plasmids pKX2-ASacI and pKX2-P4 were constructed from pKX2 in the following manner. pKX2 was partially digested with Sacl, religated with T4 DNA ligase, and transformed into Escherichia coli. Ampicillin-resistant clones were screened only for deletion of the 0.5-kilobase
0 81 0 82 083 084 085 086 087
1 1
11 LI1 1 T
0 2 3 4 5 6 7 8 9 10
X X
m u
-kIb
E pKXZP4
-X pKX2 -pKX2-&SocO
-pK8Z
E
C C E
8 83
ICP4 mRNA EMORIs
FIG. 1. HSV-1 DNA inserts inplasmidsused in this study.The regionof the HSV-1 genome in theIsorientation(17) between map units(m.u.)0.806 and 0.877containingoris(47),the ICP4upstream regulatory sequences(closed box) (23),theICP4message(50),and thea sequence (27) is shown with respectto the HSV-1 inserts in pKXC, pKX2-P4, pKX2, pKX2-ASacI,andpKBZ.Thederivation of these plasmids is described in Materials and Methods. The relevantrestriction sites shownareEcoRI (E), SmaI (M), BamHI (B), SacI (C), andXhoI (X). The dashed line represents the L-S junctionof the viral genome.
R7s -8 1- "
a
VOL.56, 1985
DKXC
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.319.560.528.626.2]approximately 40% of the G418-resistant clones demon-strated complementing activity.
CAT assays. The transfection of HSV-1-chloramphenicol acetyltransferase (CAT) chimeras and in vitro assays for CATwereperformed byamodification (10) of the procedure of Gorman et al. (15). The construction of pIE3CAT, ptkCAT, and p5CATwas described previously (10). These plasmids contain the CAT gene under control of HSV-1 immediate-early, early, and late promoters, respectively.
Electrophoresis. Lysates of [35S]methionine-labeled in-fected cell cultures were prepared and electrophoresed on sodium dodecyl sulfate (SDS)-polyacrylamide gels as de-scribedpreviously (8). DNA fragments generated by restric-tion endonuclease digestion were separated on horizontal agarosegels. Cytoplasmic RNA was separated by electro-phoresis on agarose gels containing 2.2 Mformaldehyde by the procedure of Goldberg (13).
Blot hybridization. The treatment of DNA restriction fragments inagarosegels, subsequenttransferto nitrocellu-lose, and hybridization were performed by the method of Southern (45). Formaldehyde gels containing separated RNA werewashedwithwater, soakedfor30 minin 50 mM NaOH-10 mM NaCl, soaked for 30 min in 0.1 M Tris hydrochloride (pH 7.5), and then soakedfor 30 min in 20x SSC (1x SSC is 0.15 M NaCl plus 0.015 M sodium citrate). RNA wasthentransferred tonitrocellulose by the procedure of Southern (45). RNA blots were hybridized to denatured probe in50%formamide-5x SSC-10 mM Tris hydrochloride (pH
7.5)-i
x Denhardtsolution-25 ,ug of salmon testes DNA per ml-10%dextran sulfate at42°C for 18 h. ForNorthern blot hybridization, 32P-labeled probe was NaOH (0.5 M) denatured in the presence of 1 mgofsalmon testes DNA. The solution was neutralized by addition of 0.2 M Tris hydrochloride (pH 7.5)-0.5 M HCI. Sequential washes of 3 and 0.1x SSC were performed at 50°C. Probes were pre-pared by nick translation (25) in the presence of[32P]dGTP and[32P]dCTP
(AmershamCorp., Arlington Heights, Ill.).Western blot analysis. Western blot analysis of gel-separated SDS peptides was conducted as previously de-scribed (2, 48), with the following modification. Before reaction with antibody, the nitrocellulose was incubated overnightat4°C with BLOTTO(21). Theantibody reaction
was performed in 10 ml ofBLOTTO-0.1 ml of ICP4 rabbit antiserum. The filter was then stained with I251-labeled protein A(Amersham). Rabbit ICP4 antiserum (6)was the generous gift of
Richard
Courtney (University ofTennessee, Knoxville).RESULTS
Derivationof a celllineexpressing ICP4. ICP4performsan
essential replicative function. To propagate viral mutants
deleted in the ICP4 gene, we constructed cell lines which express thewild-typegenefor ICP4. Viable cellsexpressing this gene have been isolated andcharacterizedpreviously(7, 32),ruling outthepossibility of inherentcytotoxicity of the resident ICP4 gene or itsproduct.
Verocellswerecotransfected withplasmids pSV2neo and pKXC (Fig. 1). After2weeks, G418-resistantcolonieswere
isolated, amplified, and screenedforabilityto support rep-lication of tsB32 at 39°C. tsB32 is a temperture-sensitive
mutant in the structural gene for ICP4 (11). Approximately 20% of the G418-resistant lines isolated were capable of complementing tsB32. The cell line designated 2-6 demon-strated the greatest complementing activity and therefore
waschosenfor further study. As wellasall other cell linesto
BamHIZ _ _
I
*
--4~~~~~..
A
_to_A_
P5 P20 C I
2-6 0
3 10 30
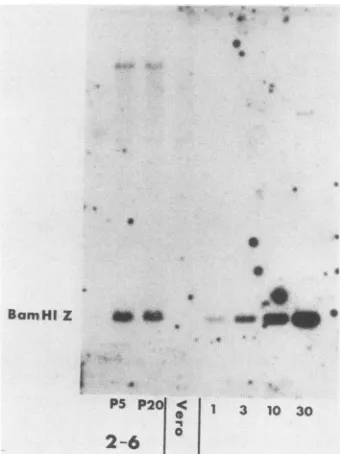
FIG. 2. Southern blot analysis of HSV-1 DNA in 2-6 cells. Electrophoretically separated BamHI digests of2-6 cellDNA (10 ,ug)(frompassages5 and20)andVerocellDNAweretransferredto nitrocellulose paper and probed with the 32P-labeled BamHI Z fragment(1.84 kb) frompKBZ(Fig. 1). We alsoincluded BamHI-digestedpKBZ (7.8kb)tovisualize1(25 pg), 3 (75 pg), 10 (250 pg), and 30(750pg)copies of viralDNAper 3 x 109basepairsofcellular DNA.
be described in this study, 2-6 cells are indistinguishable fromVerocellswith respecttomorphologyandgrowthrate.
Todemonstrate the presenceof ICP4codingsequencesin 2-6cells andtodetermine theapproximatenumberof copies of the ICP4genepresent, wecompared DNA from 2-6 cells with Vero cell DNA bySouthern blot analysis (Fig. 2). The probe used in this experiment was the purified BamHI Z fragment obtained from plasmid pKBZ (Fig. 1). This
frag-mentcontainsapproximately50% of thecodingsequencefor ICP4. The standards present on the right half of the gel indicate that 2-6cells contain approximately five copies of the BamHI Zfragmentper3 x 109basepairs.Moreover, 2-6 cells appear stable with respect to the integrity of these sequences over 15 passages(Fig. 2).
2-6 cellsexpressbiologically active ICP4. 2-6 cells contain approximately5copies ofthe BamHI Zfragmentperhaploid equivalent or 10 copies per cell. This does not necessarily imply thatall five copies serve astemplatesfor the produc-tion of ICP4. The measure ofICP4 expressionused in this studywasthecomplementationof ICP4 ts mutants at39°C. Table 1 shows thegrowth characteristics ofvarious HSV-1 ts mutants on2-6 and Vero cellsat 390C. tsB21,tsB32, and
tsL14 each specifyathermolabile ICP4polypeptide (8, 11). tsA24, tsJ12, andtsY46contain mutations in ICP8(51), gB (9), andICP27 (40),respectively. Inthisexperiment,2-6and Verocellswereinfected with1PFUof theindicatedmutant
per cell at 39°C. The cultures were harvested at 18 h
postinfectionandassayedon Verocell
monolayers
at34°C. Onlymutantsdefective inICP4grewto substantially higher levels (3to4ordersofmagnitude)
in 2-6 than in Verocells. However, eithercomplementation was notcomplete
or themutantforms of ICP4 wereabletointerfere with the action of thecomplementing wild-typeICP4polypeptide,sincethe
yieldof mutant virus was 1 to 2orders of
magnitude
below9 9
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.351.521.71.298.2]HSV-1 DELETION MUTANTS IN THE ICP4 GENE 561
Vero
0 1 3 10 K K
2-6
_
__"_F
~..md
fI
e. >* v;X -i $.
4
5
6 0
8
11
-.S
am aw.- 25
_, &a-* 27
[image:4.612.91.255.71.410.2]*6 meMO
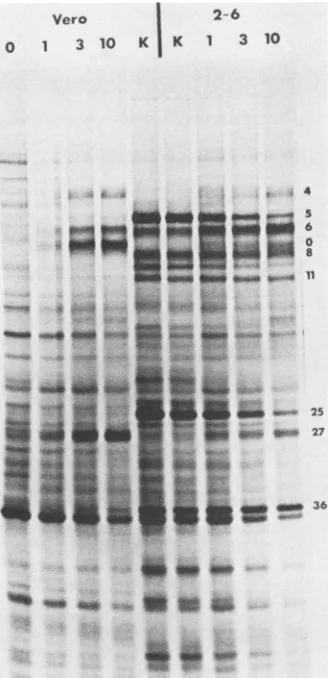
FIG. 3. Synthesis ofviral polypeptides in tsB32-infected Vero and2-6 cells.Monolayers of 2-6and Vero cellswereinfected atthe indicatedmultiplicities ofinfection with tsB32 at39°C. Cells were
also infected with KOS ata multiplicity of infection of 3 PFUper cell(K).[35S]methioninewaspresentduringincubation from 5to16 h postinfection. At 16 h postinfection, total cell lysates were
preparedfor SDS-PAGE. The infected-cellpolypeptides were elec-trophoretically separatedona 9% polyacrylamide gelcross-linked with N,N'-dicyclohexylcarbodiimide (28). Infected-cell polypeptides representative of the x (ICP4, ICPO, and ICP27), 1B
(ICP6, ICP8, ICP11,andICP36),and-y(ICP5, ICP19-20,andICP25)
kinetic classes arelabeledfor reference (28).
that of KOS in 2-6 cells. The diminished yield of ICP4 ts
mutants in 2-6 cells at 39°C relative to wild-type virus was not significantly reflected in the 39-34°C plating efficiencies of the mutants on2-6 cells. The plating efficiency (39-34°C)
ofwild-type viruson2-6 cells wasnear1.0, whereas that of themutants ranged from0.3to 0.8. The plaque sizes of the mutants on 2-6 cells at 39°C, however, were substantially
smaller thanthatof KOS (datanot shown).
An alternative means of assessing the extent of comple-mentation of ICP4 ts mutantsin 2-6 cellsisto compare the
synthesis ofviralpolypeptidesin2-6 and Vero cellsat39°C.
InVero and other cell types, tsB32 characteristically
over-produces immediate-early polypeptides and fails to
synthe-sizeearlyand latepolypeptidesat39°C (11).This is
presum-ablydue tothe lack offunctional ICP4 at thistemperature. The complementation of tsB32 by wild-type ICP4
synthe-sized in 2-6 cells, asreflected bythe synthesis of early and late viralgeneproducts, wasboth substantial and
multiplic-ity dependent (Fig. 3).Atamultiplicityof 1 PFUpercell,the
polypeptide profileof tsB32 in2-6 cells resembledthat ofthe wild-type virus.Atmultiplicitiesgreaterthan 1 PFU per cell, however, characteristics of the mutant phenotype were clearly evident and becamemore pronounced with increas-ing multiplicity. A similar multiplicity effect has also been observed forts mutants inthe HSV-1DNA-binding protein in transformed cells (41). 2-6 cellscontainapproximately10 copies of the BamHI Z fragment per cell, yet this number
was notsufficient to restore completely the wild-type phe-notype atthe multiplicities of infection tested. Three possi-ble explanations existfor this observation. (i) Only a small fractionof theintegratedICP4 sequencesserve astemplates for theexpressionof ICP4.(ii)ICP4 mRNA isnot
expressed
asefficientlywhen its templateisintegratedinto the cellular genome as in the viral genome. (iii) The mutant ICP4 polypeptide interferes with the action of wild-type ICP4 present in transformed cells. Data
presented
below support the thirdpossibility.
Derivation ofa cell lineexpressing ICP4but lacking oris. The fragment of HSV-1 DNA used to generate 2-6 cells
contains,
in thefollowing order,
thecis-acting
packaging
sequences(a)(27)derived from thejoint region
orterminiof the viral genome, the gene for ICP4 (26,28, 38),
the regulatory sequencesforexpression
of ICP4 mRNA(5, 23),
andanoriginof viral DNAsynthesis (oris) (47) (Fig. 1).
As shown previously (7), passage of ICP4 ts mutants in suchcells resulted in the
generation
of ts+ recombinants. In the present study, the number of recombinantsgenerated
incomplementation
tests afteramplification
ofasingle
tsB32 plaque isolate in 2-6 cells wasapproximately
1%(data
notshown). Because this level was well below the
yield
ofts mutantvirusgenerated by
complementation
in thesecells,
itwas felt that wild-type recombinants would not
seriously
interfere with the isolation ofdeletion mutants.Therefore,
2-6cells wereusedtogenerateandscreenputative
deletionmutants in
ICP4,
as described below. To obtain recombi-nant-free stocks of the ICP4 deletion mutants,however,
itwas necessary toisolate ICP4-transformed cell lines which would not give rise to
wild-type
recombinants. Conse-quently, the deletionmutantsgenerated
as described belowwere used to screen G418-resistant cell lines isolated after cotransfection of
pSV2neo
andpKX2-P4
(Fig. 1). pKX2-P4
containstheasequences,ICP4,
and theregulatory
elements for ICP4expression
but lacksoris.
It wasanticipated
that deletingoris
wouldreduce,
ifnoteliminate,
thegeneration
of wild-type recombinants.Recently,it has beendemonstrated thatthis isindeedthecase(7). Among
the celllines isolated after cotransfection withplasmids pKX2-P4
andpSV2neo,
approximately
40% were able to supportgrowth
of the deletion mutants. Three were retained for furtheranalysis.
The threelines,
1-2, 1-5,and4-2,containedthree, three,
and five copies, respectively, of the ICP4coding
sequence perhaploid equivalent
of DNA(data
notshown).
Isolation of deletion mutants in 2-6 cells. A
plasmid
con-taining
a 0.5-kb deletion in the ICP4coding
sequences was constructed frompKX2
by partial
Saclcleavage
and religa-tion(Fig.
1). The deletedICP4-containing
plasmid
pKX2-ASacI was first assayed for abilityto express ICP4activity before attempts to introduce the deletion into the viral genome. This was doneby
cotranfecting
CV-1 cells with either pKX2 orpKX2-ASacI
and HSV-1-CAT chimeric genes andmeasuring
the levels ofexpression
of the HSV-1-CATchimeras inducedby wild-type
ICP4 and the deleted form of theprotein.
In this way the effect of the 0.5-kb deletionon ICP4activity
could be assessed. We and others have shown that ICP4 alone can suppressimmediate-early
VOL.56, 1985
'A ii ii&Ai lmw ".0.0.w
v
'.c. 1.
-M,41
414 I
on November 10, 2019 by guest
http://jvi.asm.org/
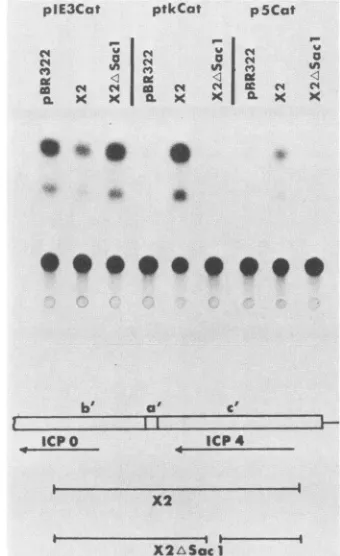
CAT chimeric gene expression (10) and simultaneously
enhance early andsomelategeneexpression intrans(10, 12, 30). Here, immediate-early-CAT (pIE3Cat) expression was
suppressed whereas early-CAT (ptkCat) and late-CAT (p5Cat) expressions wereenhanced bywild-type ICP4 (Fig.
4). The CATsignal generated fromextractsof cells cotrans-fected with the deleted ICP4 plasmid resembled that derived fromextractsof cells cotransfected with pBR322. Therefore, the0.5-kb internal deletion abolished the trans-acting activ-ity ofICP4.
pKX2-ASacI wasthenlinearized andcoprecipitated with
infectious KOS DNA, and 2-6 cellswerecotransfected with
the mixture. Individual progenyplaques from this
transfec-tionwereisolated and screened for ability toreplicate in 2-6 cells and not in Vero cells. Of 224 plaque isolates, 2, designated d120 and d202, exhibited impaired ability to replicate inVero cells.
Stocks of d120 and d202 generated in 2-6 cells always contained 1 to 5% ([titer of Vero cells/titer of 2-6 cells] x
100) wild-type recombinants which produced plaques in Vero cells. Such recombinants, when isolated and plated in Vero and 2-6cells, always yielded platingefficienciesof 1.0. When individual plaque isolates of d120 and d202 were
propagated in 1-2, 1-5, or4-2 cells, wild-type recombinants wererarely detected. Occasionally, however, when d120or
d202was amplified through several rounds of growth in
ori-ICP4+ cell lines, wild-type recombinantsweregeneratedata
frequency approaching
10-5.
The growth characteristics of d120 and d202 on control Nero (G418-resistant Vero cells[seeMaterials and Methods]), 1-2, 4-2, 1-5, and2-6 cellsare
shown in Table 2. In thisexperiment, wild-type virus was not present in lysates obtained from ori- cells. Recombi-nant-free stocks of d120 and d202 grown in 4-2 cells were
usedfor further analysis.
Characterization of deletions in d120 and d202. d120 and
d202 aretheprogeny ofcotransfection of pKX2-ASacI and
KOS DNA. To determine whetherthese isolates contained the deletion in pKX2-ASacI, we analyzed viral DNA from
d120 and d202by restriction enzymedigestion.
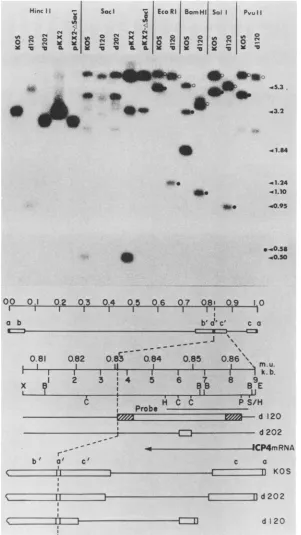
Electropho-retically separated restriction fragments of KOS, d120, and d202 were transferred to nitrocellulose paper and probed with theindicated 32P-labeled fragment (Fig. 5).
The isolate d202 contained the expected 0.5-kb Sacl deletion in bothcopies of ICP4. The 3.2-kb HincIIfragment ofpKX2 comigrated withthat ofKOS, whereas theHincIl
plE3Cat ptkCat p5Cat
04 X'R 41 X X0 4
('I~~~~~~~~~~~~~~~~~~
(a4 C'o4 C r
C') X XII IL X4
j-('4 x
* *
b' a' c'
ICPO +ICP 4
X2
[image:5.612.354.524.72.350.2]X2Asac1
FIG. 4. trans-Inducing activities of wild-type and deleted forms of ICP4. CV-1 cells were cotransfected with 2 ,ug each of the
indicated CAT plasmid and the indicated test plasmid. At 40 h posttransfection, cellextractswerepreparedand analyzed forCAT activity in vitroas previously described (15). Shownare the auto-radiographic images of the chromatographically separated input,
unacetylatedchloramphenicol (lowerspot),andthe acetylated prod-ucts (upper spots). Alsodiagrammedarethetestplasmidsusedin the cotransfection with respectto the genes forthe infected cell
polypeptides ICP4 and ICPO. b', a', and c'representtheelementsof the HSV-1joint region(17).
fragment of pKX2-ASacI comigrated with that of d202. Furthermore, theSaclfragments of d202homologoustothe 3.2-kb HincIl probe were identicaltothose of KOSexcept
for the absence of the 0.5-kb deletion inpKX2-ASacI. The HincIl and Sacl digests of d120 DNA indicated a
TABLE 2. Growth ofICP4mutants ontransformed cells Virusgrown ona
Virus
Nerocells 4-2Cells 2-6 Cells 1-5Cells 1-2Cells
KOS 1.9 x 108b 2.2 x 108 1.4 x 108 2.0 x 108 9.0 x 107
1.8 x 108c 1.5 x 108 1.1 x 108 1.7 x 108 1.0 x 108
d120 2.5 x 104b 4.0x 107 4.9 x 107 5.2 x 107 4.2 x 107
<5c <102 1.7 x 105
<102
<102d202 7.2 x 104b 6.5 x 107 3.7 x 107 6.5 x 107 3.3 x 107
<5C <102 2.2 x 105 <102 <102
tsB32 6.2 x 102d 1.0x 104 3.0 x 104 2.0 x 103 2.0x 103
<5e <5 2.0 x 102 <5 <5
aNero,4-2, 2-6, 1-5, and1-2cellswereinfectedwith theappropriatevirusunderconditions identicaltothose stated inFig.9.
bInfectedcultureswereharvestedat18 hpostinfectionandassayedat34°Con1-2cells.
' Same cultureasthatdescribedinfootnote bassayedsimultaneouslyat34°ConNero cells.
dInfected cultureswereharvested at 18 hpostinfectionandassayedat34°ConNerocells.
eSame cultureasthatdescribedinfootnotedassayedsimultaneouslyat39°ConNerocells.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.68.564.546.679.2]HSV-1 DELETION MUTANTS IN THE ICP4 GENE
Hunc II; Sacl EcoRI BamHI Sal I PvuII
io o X VI cm
a t4~x£~ 0 0
x C a9 xgx
-lb
40-.
4
4.4
00 a.
0
00 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8r 0.3 1.0
I I I I I 1 I I I, 1I
o b
>~~
b'a"c' c a-
..
... ... ... ...
0.81 082
os
--0.84
0.85 0.86 \\ f ..
I I I 1 1 I
I}
|x
-?1
2 3 14 5 6SI
8 at-_
I,__C
I--1CC I -_,. HCCc P S/
Probe
k. b. E (Hl
r
--b a'o c'I
rCPAM
c a
KOS
crzz=
d202
ar-
d120FIG. 5. Genomestructuresofd120 andd202. Viral (KOS, d120, and d202) and plasmid (pKX2 and pKX2-ASacI) DNAsweredigestedwith
theindicated restriction enzymes and electrophoretically separated on a1.2% agarose gel. Separated DNAfragments wereanalyzed as
described in thelegendtoFig. 2 with the indicated 3.2-kb HincIl fragmentasprobe. Forcomparison of KOS and d120 fragments (right half
of thegel), terminal (0) and joint (0) fragmentsareindicated. Shown below the autoradiogram in descending orderaretheKOSgenome,an
expansion of thejoint region of thegenomebetween 0.806 and 0.867mapunits(m.u.),thelocations ofrelevantrestriction sites,thelocation
ofthefragment usedasprobe, and the deducedstructuresof d120 and d202. Theopenand shaded boxesin thestructuresofd120andd202 indicate deletedsequences andregions of uncertainty, respectively. The boundaries between deleted sequencesandregions of uncertainty ind120DNAwerededuced fromthefollowing observations. (i)d120containsa4.1-kbdeletion. (ii) The PvuII site ispresentatm.u.0.863.
(iii) The functionalasequences arepresent atthejoint,asdemonstrated by evidence of inversionof L and S. (iv) The restriction sites spanned by theopenboxind202areabsent in d120. The right portions of the KOS, d120, andd202genomes arediagrammedin the lower portion of thefiguretoillustratetheeffects of deletions in both oftheinvertedrepeats(C and C')onthesizesof the shortcomponentsof thegenomes.
Adashedline is drawntoindicatetheL-Sjunction. The relevant restrictionsitesarethesameasin Fig. 1, with the addition ofHinclI (H),
PvuII (P), andSall (S).
-5.3.
.3.2
.41.84
-.cl24 -a1.O .c0.95
.40s8
{0.30
-d 120
d202
MD
I
11
VOL.56, 1985 563
I I I
I I
i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.155.456.70.605.2]more substantial deletion. The structure of d120 was ana-lyzed further by Southern blot hybridization. The right half ofFig. 5 is an example of one such experiment. The 5.3-kb terminal EcoRI K fragment of the KOS short component was absent in d120 DNA, and a new 1.24-kb band was present. The deletion in d120 spanned the twoBamHI sites located near 0.853 map units (Fig. 5) in KOS, and a new 1.1-kb band was present in d120. The 5.1-kb terminal Sall fragment of KOS was replaced by a 0.95-kb fragment in d120. Likewise, the4.7-kbterminal PvuII fragment of KOS wasreplaced by a 0.58-kb fragment in d120. The size of the novel terminal fragments in d120 was consistent with the separation between, and order of, the EcoRI, BamHI, SalI, and PvuII sites near the 5' terminus of the ICP4 gene in wild-type HSV-1 (Fig. 5). The sizes of the reductions in the abovementioned terminal fragments (Fig. 5) and in the joint fragments (Fig. 5; data not shown) are consistent with the existenceof a 4.1-kb deletion in both copies of ICP4.
Based on other Southern blots (data not shown), restric-tion sites in the long component of the genome did not appear to be affected in either d120 or d202, and no se-quenceshomologous to pBR322 were detectable in the DNA of eithervirus. In addition, the expected patterns generated by genomeisomerizationwerereflected inEcoRIandPvuII
44.3
- 3.8
< 2.8
b' ca0' c'
r1
1:=
ICPO ICP 4
X2
[image:7.612.98.267.338.627.2]X2 Sac1
FIG. 6. ICP4 mRNA synthesized in d120 and d202. Vero cell monolayers were preincubated in thepresence of 75 ,ugof cyclo-heximideperml for 1 hat37°C. Monolayerswerethen infected with 10 PFU ofKOS,d120,ord202percell in the presenceof 75 ,ugof cycloheximideperml. After 6 h of incubationat37°C, the cultures
wereharvested andcytoplasmic RNAwasanalyzedasdescribed in
the text. The locations of the ICPO and ICP4 mRNAs and of the
deletioninpKX2ASacIusedtogenerate d202areshown beneath the autoradiogram. The numbersontherightindicate size inkilobases.
b', a', and c'aredescribed in thelegend toFig.4.
!,
4 "w4v49
4
K64
infected asdescribed in the legendto Fig. 6. At 6 hpostinfection, cycloheximide was washed outwith medium containing 10 ~±g of actinomycinDperml.Incubationwascontinuedforanadditional 3 h in the presence of 10 ~tg of actinomycin D and 50 p.Ci of [35S]methionine per ml. Cell lysates were then prepared and
ana-lyzed bySDS-PAGEasdescribed in thelegendtoFig.3. Numbers
onthe leftaredescribed in thelegend toFig. 3.
digests (Fig. 5; data notshown). Thus, because the presence of a sequences at both thejoint and molecular termini are required for isomerization (33), their presence in d120 in a functionalconfigurationis implied.
ICP4 mRNAsynthesis by d202 and d120. d202 contains the 0.5-kb deletion introducedbypKX2-ASacI.Therefore,d202
was expectedto synthesize an ICP4 mRNA 0.5 kb smaller than that of the wild-type virus. d120 contains a large deletion (4.1kb)andthereforewasexpectedtosynthesizea
muchsmaller messageor none atall. Northern blotanalysis ofcytoplasmic RNA from KOS-, d120-, and d202-infected
Vero cells verified that thiswas the case (Fig. 6). Infected Verocellswereincubated in the presence ofcycloheximide for 6 h. Cytoplasmic RNAwasthenisolated, electrophoret-ically separated, transferred to
nitrocellulose,
and probed with nick-translatedpKX2 DNA.pKX2 contains sequences homologous to the immediate-early mRNA for ICPO and ICP4. KOS induced the synthesis ofa4.3-kb mRNA speci-fyingICP4 anda2.8-kb mRNAspecifyingICPO(50) (Fig.6). d120 synthesized only the 2.8-kb message, whereas d202 synthesizeda3.8- anda2.8-kb message.Therefore,
although ICP0 mRNA was synthesized in cells infected with the deletion mutants, as it was in KOS-infected cells, ICP4t,
0
0
40
"40on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 DELETION MUTANTS IN THE ICP4 GENE 565
Mock
d120 d202
KOS
.Avmm
170140
&~~~~~~~~~~~~~~~~~~~~~~~
_
1~~40
FIG. 8. Western blot analysis of d120- and d202-infected Vero cells. Non-ICP4-containing cells were infected withthe indicated
virusatamultiplicity of10 PFUpercell.At5hpostinfection, SDS
lysateswerepreparedandrunon a9%SDS-polyacrylamide gel.The SDSpeptideswereelectrophoretically transferredtonitrocellulose andanalyzedasdescribedin Materials and Methods. Numberson
theright indicateapparentmolecularweight.
mRNA was 0.5 kb smaller in d202-infected cells and not detectableby thisprocedure in d120-infected cells.
Immediate-early polypeptide synthesis in d120- and
d202-infected Vero cells. To determine what immediate-early polypeptides where synthesizedin d120- and d202-infected Vero cells, we performed an experiment similar to that shown inFig. 6, except thatat6 h postinfection,
cyclohex-imidewas washedoutand incubationwascontinued for 3 h in the presence of actinomycin D and [35S]methionine. Imnmediate-early polypeptides ICPO, ICP22,andICP27were
clearlyvisible inextracts ofdl20-, d202-,andKOS-infected cells on SDS-polyacrylamide gels (Fig. 7). As in other
cycloheximide reversal experiments, small quantities of ICP6were also observed. The ICP4polypeptide, however, was detectable only in extracts of KOS-infected cells.
Whereas one might not expect an ICP4 polypeptide to be
synthesized in d120-infected cells, it is possible that a deletedform of ICP4wassynthesizedind202-infectedcells.
Although this is not evident inFig. 7, the possibility ofan ICP4-like peptide comigrating with a peptide ofa different
electrophoretic mobility than ICP4 exists. This possibility
was investigated furtherby Western blot analysis. Electro-phoretically separatedSDS peptides preparedfrom infected
anduninfected Nero cells were transferred to
nitrocellulose,
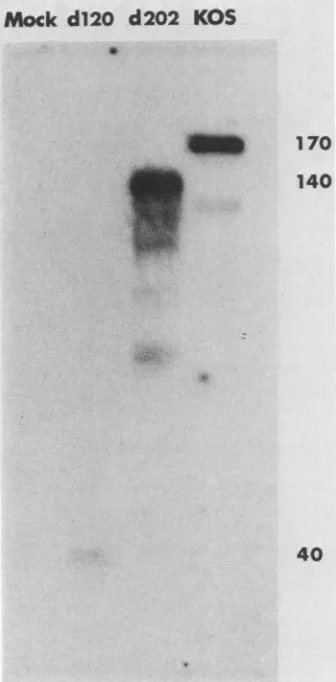
probed with anti-ICP4 rabbit serum, and visualized by staining with'25I-labeled
protein A. The anti-ICP4 serum used was generated from rabbits inoculated with a gel-eluted SDS ICP4polypeptide (6). Such polyclonal antibody would be expected to react efficiently with truncated SDS ICP4 peptides. Cells infected with d202 contained a polypeptide shorter than KOS by about30kb (Fig. 8). This polypeptide probably comigrated withICP6 orICPO (Fig. 7). The d202 formof ICP4appeared relatively unstable with respect to the wild-typeform of ICP4, asreflectedby the descending smear and several specific lower-molecular-weight bands. The or-igin of the lower-molecular-weight band in KOS is unclear and may represent adegradatiqn
product of the wild-type ICP4polypeptide. Itisnot,inall likelihood,acellular bandor another HSV-1 immediate-early peptide, since it was absent from mock-infected and d120-infected cell profiles. d120 appears to specify a peptide of approximately 40 kilodaltons, which is reactive with ICP4 antiserum. This peptide represents the amino terminusofICP4, since only the 5' end of the ICP4 genewasretained in d120 (Fig. 5).
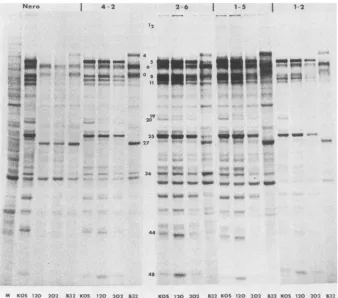
Otherphenotypicproperties of d120andd202. Thegrowth propertiesandpolypeptide phenotypesofd120, d202,tsB32, and KOS were compared in Nero and ICP4-transformed cells. Nero cells and cells of four ICP4-transformed lines
wereinfectedat amultiplicity of10 PFU percell and labeled with [35S]methionine from 5 to 16 h postinfection. All in-fected cultureswereincubatedat34WC,exceptthoseinfected with tsB32, which we incubated at 39°C to visualize the
mutant phenotype.
Duplicate
cultures were harvested for simultaneous infectivity determinations.Thepolypeptideprofiles ofd120 and d202differed drasti-callyfrom that of KOS in Nero cells (Fig. 9). Qualitatively, the profiles of d120- and d202-infected cells were quite similartoeach other andtothatoftsB32-infected cells,with the exception of ICP4, which was evident only in tsB32-infected cells. Thepredominant viral polypeptides detected in Nerocells infectedwith thedeletionmutants were ICP6, ICPO, and ICP27. The truncated forms of ICP4 are not
evident inFig.9.In ourhands, ICP22wasdiscernibleonly in cycloheximide reversalexperiments. Both ICPO and ICP27
are immediate-early polypeptides (28), whereas ICP6 is
generally notregarded as amember ofthis class of HSV-1 genes. It isclear, however,that theexpressionof ICP6 does
notrequire functional ICP4 (Fig. 9).
The polypeptide profiles of d120 and d202 in ICP4-transformed cells resembled that of the wild-type virus, indicating efficient complementation by resident, wild-type ICP4. Enhanced expression ofearly polypeptides in tsB32-infected ICP4-transformed cells relative to tsB32-infected Nero cells (i.e., ICP8, ICP11, and
ICP36)
wasalso evident. Aspects of the mutant phenotype characteristic of tsB32, however, were evident in all ICP4-transformed cell linesatthis multiplicity. This observation is consistent with the results shown in Fig. 3, in which the ts mutant was
ineffi-ciently complemented
in ICP4-transformed cells at high multiplicity of infection. The fact that early and late polypeptideswereexpressedrelatively inefficientlyin ICP4-transformed cells infected with thetsmutant, whereasthose of the deletionmutants wereefficiently expressed, indicates that the ts mutantform of the ICP4 polypeptideexhibitsaninterferingeffectongeneexpression,thusinhibiting comple-mentation.
Thegrowthcharacteristics ofKOS,d120, d202,and tsB32
are summarized in Table 2. The following statements
con-cerningthese viruses can be made. (i) The growthofd120,
VOL.56, 1985
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.93.261.74.415.2]Nero 2-6
I
1-5I
1-2 12*-U4 U-_
-*~ _ U_O8-
--M KOS 120 202 832 K05 120 202 19 20
_m 25 IMM
_ 27 -N_ *-..O ---ku ow -'1
, t ~. ;w,4 i*.
36 wom w_> 0 ,
U c:
44 _
48
-832 KOS 120 202 832 KOS 120 202 B32 KOS 120 202 832
FIG. 9. Viralpolypeptides synthesized inKOS-,d120-,d202-,and tsB32-infected Nero andICP4-transformedcells. Confluentmonolayers of each celltype wereinfectedasdescribedinthe text andanalyzed by SDS-PAGEasdescribed inthelegendto Fig. 3. All infectionswere performedat34°C,except thosewithtsB32,whichwereperformedat39°C.AnuninfectedNero cell culture was maintained for visualization of cellularbands (M). Theuninfected cell polypeptide profiles ofNero,4-2, 2-6, 1-5, and 1-2 cells were identical (data not shown). Numbers in the center aredescribed in the legend to Fig. 3.
d202, and tsB32 (at 39°C) in Nero cells was severely re-stricted. (ii) All ICP4-transformed cell lines complemented
d120 and d202 efficiently. (iii) ICP4-transformed cell lines complementedthegrowthof tsB32poorlyatamultiplicityof infection of 10 PFU percell. Only in2-6 and 4-2 cellswere
significant levels of complementation demonstrated. (iv) Wild-type recombinants were observed only when ICP4 deletionortsmutantswerepropagatedin2-6 cells. Thus, the yields ofmutant and wild-type viruses incell lines
express-ing ICP4 supportthe results ofpolypeptide analysis.
The experimental results shown in Fig. 9 illustrate a
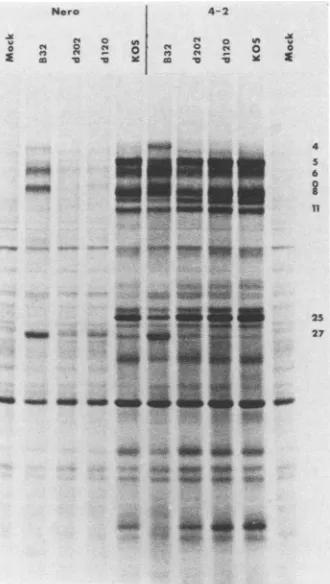
qualitative comparison of polypeptide synthesis by d120, d202, and tsB32 (at 39°C) in Nero and ICP4-transformed cells. When all of the incubations werecarriedoutat 39°C,
the same pattern of polypeptide synthesis was observed in ICP4-transformed cell lines. The polypeptide profiles of d120- and d202-infected 4-2 cells were similar to that of
KOS, whereas the profile of tsB32-infected 4-2 cells was morerestricted,asshownbythe relativeunderproductionof ICP5 andICP25 (Fig. 10).Also clear is the factthat,inNero
cells, d120 and d202 synthesized reduced levels of
immedi-ate-early polypeptides ICPO and ICP27 relative to tsB32. ICP6 was also reduced in d120- and d202-infected cells relative to tsB32-infected cells. To determine more
specifi-cally whether the rates of synthesis of ICPO, ICP27, and ICP6 in d120- and d202-infected cells differ from that in tsB32-infected cells and to examine the possible effects of
proteinturnoveronpolypeptide phenotype,welabeled Nero cells infected with these mutants for 0.5-h intervals at
selected times from 0to8 hpostinfectionat39°C. Therates
ofsynthesisofICP0, ICP27, and
ICP6
incells infected with the deletion mhutants were slower than that seen in tsB32-infectedcells(Fig. 11). This differencemay be attributedto adirect orindirect effect ofthetsB32form of ICP4. Inthis case, it would be postulated that the presence of the ts mutantform of the ICP4polypeptide
resultedinexaggerated expression of immediate-early polypeptides.DISCUSSION
The genome of HVS-1 is capable ofcoding for approxi-mately 100 polypeptides. Despite the large size of its genome,HSV-1ishighly conducivetogenetic analysis,thus pertnittingtheidentification ofspecificHSV-1-encoded func-tions. ts mutants, in particular, have
proven
invaluable in thisregard (1, 42) in that, todate, complementation groups representing approximately 30 essential replicative genes have beenlocalized ontheHSV-1 genome. Inadditionto tsmutants, mutants resistantto avariety ofanti-herpesdrugs (4),mutantswhich render cells resistanttoimmune
cytolysis
(31), and mutantsresistanttoneutralization with monoclonal antibodies (18) have proven useful inidentifying
and char-acterizingthegenomiclocations andfunctions ofindividual HSV-1 geneproducts.Using cloned HSV-1 genes and selections for both the thymidine
kinase-negative
(tk-) and-positive (tk+)
pheno-types, Post andRoizman (34) have developeda method to deletenonessential genesof HSV-1. Thisapproachledtothe isolation of viable deletion mutants in theimmediate-early
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.612.141.479.75.373.2]HSV-1 DELETION MUTANTS IN THE ICP4 GENE 567
Nero 4-2
o X X o o
-3-c co
OA__
I0
£
4 5 6
11
25 27
_ _ _ _
FIG. 10. Viral polypeptides synthesized in KOS-, d120-, d202-, and tsB32-infected Nero and 4-2 cells at 39°C. Nero and 4-2 cells were infected, labeled, and analyzed asdescribed in the legend to Fig. 8,exceptthat allincubationswerecarriedout at39°C.Numbers ontherightaredescribedin thelegendtoFig.3.
ICP4 deletion mutants d120 and d202 were isolated from the progenyof a cotransfection of pKX2-ASacI (Fig. 1) and intact KOS (wild-type) DNA. pKX2-ASacI contains a 0.5-kb deletion in the coding sequence for ICP4. In the absence of sequenceinformation in this region of the genome, it was of interest to determine if the ICP4 gene in pKX2-ASacI retained any ICP4 activity. The transient expression assay shown in Fig. 4 indicates that pKX2-ASacI does not specify ICP4-like activity. This experiment enabled us to ascertain what the consequences of such a deletion would be before introducing the deleted gene intowild-type virus. As noted above, adeletionmutant in ICP4 hasbeen isolatedrecently which does not inactivate the gene product (44).
d202contains theexpected0.5-kb deletion in both copies of ICP4. Consistent with the presence ofa 0.5-kb deletion internal to the ICP4-codingsequences wasthe synthesis of anmRNAwhich was smaller than itswild-typecounterpart by 0.5 kb and the synthesisofa correspondingly truncated form of ICP4. The structureofd120, ontheotherhand, was
quiteunexpected. Thismutant containsa4.1-kbdeletion in both copies of ICP4 and failed to express detectable cyto-plasmic ICP4 mRNA. A severely foreshortened form of ICP4 was, however, detectedby Western blot analysisand
KOS d 120 d 202 B32
A AAA A A Ai AA A AAAA A
In An if in; InAM kmem I e,in il VW VW
(a)geneforICP22, indicatingthenonessentiality of thisgene for virus replication in the cell lines tested. Moreover, a
viable deletion mutant has recently been isolated in a non-essentialregion of ICP4(44). In thisstudy,wegeneratedcell lines transformed with the gene for the HSV-1 immediate-earlygene encodingICP4.
ICP4-containing
cells were usedtoisolateand propagate twoHSV-1 deletionmutantsin the geneforICP4.Thephenotypesofthe twodeletionmutants, d120 and d202, were then analyzed on cells which do not
contain the ICP4 gene.
Isolation ofICP4 deletionmutants. ICP4-transformedcells
were generated by cotransformation of Vero cells with pKXCandpSV2neo. ICP4-containing2-6cellswereused to
screen for ICP4 deletion mutants. Two ICP4 deletion mu-tants were isolated in this manner. Consistent with the findings of Davidson and Stow (7), passage of ICP4 ts or deletion mutants on 2-6 cells resulted in the generation of wild-type (ts+ for ts mutants; host cell independent for deletion mutants) virus in the progeny. This presumably
occurs in the presence ofhelper functions provided by the superinfecting virus by replication, mobilization, and prop-agation ofwild-type sequences containing ICP4 in 2-6 cells. Amplifiedsequences arepackagedasdefective genomes (7) which are then able to recombine with the superinfecting
mutanthelpervirustoproducewild-type virus. Cells gener-atedby transformation with pKX2-P4 lack
oris
and did not result in thegenerationofwild-typevirus afterinfectionwithts ordeletion mutants (Table 2).
g m".I
A"
me b
"4000 4
11
25
5 w Sn lo _27
*-***
~~~~~~36
FIG. 11. Rates of synthesis of viral polypeptides at 39°C in KOS-,d120-, d202-,andtsB32-infectedNero cells. Nerocellswere infected with the indicated viruses and incubated at 39°C as de-scribedinthelegendtoFig.8. ["5S]methionine(100p.Ci)wasadded duringtheindicated timeintervals. At the endofthelabeling period, monolayers werewashed and celllysates were preparedand ana-lyzedbySDS-PAGE asdescribed in thelegendtoFig.3. Numbers ontherightaredescribed in thelegendtoFig. 3.
VOL.56, 1985
.f,.
"I Z.
44, 2Lk-.
4"
.:
4
p
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.612.98.263.75.367.2] [image:10.612.338.520.331.653.2]may represent expression of the amino terminus of ICP4. One wouldpredictthatanotmRNAwouldinitiate fromthe
ICP4 mRNA start site ind120, sincethisregion ofthe ICP4 gene is intact. Repeated efforts to detect an ICP4-specific mRNA in d120-infected cells have failed, however. As anticipated, the quantity and mobility of the ICP0 mRNA wereidenticaltothe analogous mRNAsynthesized in KOS-andd202-infected, cycloheximide-treated cells.
Despite differences in the size and structure of the ICP4 gene and the size of the ICP4 mRNA synthesized in d120-and d202-infected cells,
the
immediate-early polypeptides expressed in cells infected with the two mutants were identicalin a cycloheximide reversal experiment (Fig. 7). In fact, withthe exceptionof ICP4, which was notdetected by standardpolyacrylamide gelelectrophoresis (PAGE) analy-sis in extracts of deletion mutant-infected cells, the quanti-ties and mobilities of ICP6, ICP0, ICP22, and ICP27 were identicaltothose observed in cells infectedwith KOS. The experiments of Fig. 7 and 8 established two points. (i) Full-lengthICP4polypeptidesare notexpressedind120-and d202-infected Nero cells. Independentofwhether the dele-tion ind202is inframe,apeptideantigenicallyreactive with ICP4 antiserumwith an apparent molecularweightreduction of30,000 with respect to the wild-type protein was synthe-sized. Anantigenicallyreactivepeptideofapproximately40 kilodaltonswassynthesized ind120-infectedcells. Thesizes of theshortened peptides were consistent with the sizes of thedeletions ind120andd202(Fig.5).(ii) Expressionof the other immediate-early polypeptides in cells infected with d120 and d202 was identical to that seen in wild-type virus-infectedcells inacycloheximide reversalexperiment. Viral geneexpression inthe absence ofICP4. Theabsence of functional ICP4 in d120- and d202-infected cells had profound effects on the expression of other viral genes relative to that seen in wild-typeinfection.
Qualitatively, with theexceptionofICP4,thepeptidessynthesized
in d120-andd202-infectedNerocellswerethesamespeciesasthose expressed in tsB32-infected cells at39°C.
ICPO and ICP27were produced in moderate quantities in d120- and d202-infectedNero
cells
(Fig. 9, 10, and 11). Interestingly, ICP6was the most abundantly expressed polypeptide in cells infectedwith thedeletionmutants. Honess
and
Roizman(19) havepreviouslyshown thatICP6 isinefficiently
expressedin theabsence ofpriorviralproteinsynthesis, placing
ICP6in the early(1)
class of HSV-1 genes. They have also shown thatICP6 is synthesized to somedegree
in the presence of the amino acid analog canavanine (20). In ourhands,
re-ducedquantities ofICP6 relative toICPO were
synthesized
inacycloheximide reversal experiment(Fig. 7).This ICP6-ICP0 ratio was reversed in d120- and d202-infected Nero cells(Fig.
11). The synthesis of ICP6in thisgenetic
back-ground indicates that ICP4 is notrequired
for efficient expression of the ICP6 gene. This observation and theminimal expression of ICP6 in a cyclohexiinide reversal experiment may well reflect the involvement ofan a gene other thanICP4
(i.e., ICP0,
ICP22, ICP27,
orICP47)
in ICP6 gene expression.Recently it
has
been shown bya number ofinvestigators
that theimmediate-early
geneproduct ICP0can actintrans in transient expression systems to enhanceexpression
of early8ISV-1
genes (30, 39). With theexception
ofICP6,
early genes were underexpressed to the same or a greaterextentindeletion mutant-infectedcells than in cellsinfected with an ICP4 ts mutant at 39°C.
Therefore,
the lack of expression ofearlygenesinICP4tsmutant-infected cells at390Ccannotbe due
solely
toinhibitory
effects manifestedby
the
ts form of the ICP4 polypeptide. Studies of the ICP4 deletionmutantsreflect theinsufficiencyofICPOto actalone tostimulateearly
geneexpression.
The tsform of ICP4affectsgeneexpression. Thetsformof ICP4 intsB32-infected cellswasapparentlyinhibitorytothe activity of the wild-type form of ICP4 in all of the
trans-formed cellstested.Inhibitionwas moreprominentat
higher
multiplicities (Fig. 3).Atamultiplicityof10 PFU percell, all of the cell linescomplementedd120 and d202efficiently,yet tsB32wascomplemented relatively inefficientlyat39°C(Fig.
9 and10;Table2).Infact, only2-6 and 4-2cellssignificantly
enhancedtheyield oftsB32at 39°Cabove theyieldin Nero cells. Possible explanations for this inhibition include the following. (i)Thetsform ofICP4bindstightlytositesbound by wild-type ICP4, inhibiting the activity of the latter by competition. (ii)Thetsform ofICP4 formsastablecomplex
with the wild-type form of the protein in the cytoplasm, thereby inhibitingeither transportof thelatter to the nucleusor itsactivity oncein thenucleus. (iii) Thetsform of ICP4 alters cellular metabolism in such a way as to render the wild-type formof ICP4 less effective.
Another property of the ts form of ICP4, as indicated by comparisonwiththedeletion mutants, is itsabilitytoinduce overproduction of immediate-early polypeptides. This was
evident from continuous labeling experiments and by pulse-labelingat 39°C (Fig. 9, 10, and 11). The rates ofsynthesis and accumulation of ICPO and ICP27 were decreased in d120- and d202-infected Nero cells relativetotsB32-infected cells. Consistent with these findings isthe observation that
an ICP4-containing clone derived from tsB32 can induce CATactivityat39°C when CAT geneexpression is under the control of an immediate-early promoter (10). A possible explanation for this finding is thatthe ts form of ICP4 retains
some of the activity associated with that of the wild-type polypeptide.
Analysis of ICP4 deletion mutants has bothconfirmed and extended the findings previously obtained with ICP4 ts
mutants(11, 36). ICP4 is required for the expression ofearly and late genes. In the absence of functional ICP4, the remaining immediate-early genes are either insufficient or
insufficiently expressed to induce significant levels ofearly genes, with thepossible exception of ICP6. The phenotypes ofd120 andd202 also indicate that the exaggerated levels of immediate-early polypeptides induced by ICP4 ts mutants
are due, in part, to activities associated with the ts ICP4 polypeptides themselves.
Many of the contentions put forth in this studyrestin part ontheassumption that the deleted forms of ICP4 have little or no effect on gene expression or on the activity of the wild-type ICP4 in any of the transformed cell lines tested. The 40-kilodalton peptide synthesized ind120-infected cells has noeffect ontherestoration of thewild-typepolypeptide phenotype in d120-infected ICP4-containing cells. On the other hand, the slightly reduced intensities of some late polypeptides on gel profiles of d202-infected ICP4-containing cells (Fig. 9 and 10) suggest a small inhibitory effect. Althoughwe cannotformally ruleout anactivityassociated with the d202 ICP4-related polypeptide, it should be noted that the plasmid clone used to generated202 had noactivity in a transient assay (Fig. 4). It remains of interest to
determine the molecular basis for the apparent inhibitory effect of the ts ICP4 polypeptide and possibly that of the d202 peptide. An attractive hypothesis is that the mutant ICP4 polypeptides (tsB32 and possibly d202) are, for the mostpart, nonfunctional and yet may still bindtothe normal site ofaction, thereby competing with any wild-type ICP4
on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 DELETION MUTANTS IN THE ICP4 GENE 569 present. Furtherstudiesareunderway to test this
hypothe-sis.
Severalotherobservationsconcerningthegrowthof d120 andd202 in ICP4-negative cells suggest
interesting
avenuesforfurther study. Unlike the ts mutants in ICP4 (29), d120 and d202 do not induce certain cellular stress proteins on infection of HELcells (datanotshown), norarecytopathic effectsevident after 24 hat a
multiplicity
of 10PFUper cell, suggesting additional differences between the deletion mu-tants and the ts mutants. Also of interest is the genesis of d120. Efforts are in progress to determine whether the deletion ind120 was inducedas aresultof markertransferorwhether deletions such as that found in d120 arise
sponta-neously.
ACKNOWLEDGMENTS
We thank Maryellen Polvino-Bodnar forexpert assistance with Western blot experiments. We also thank Don Coen for useful comments on the manuscript and Madelon Datz for manuscript
preparation.
Thisinvestigationwas supported byPublic Health Servicegrant CA20260 from the National Cancer Institute.N.A.D.is therecipient
ofPublic HealthServicepostdoctoralfellowshipAI06790 from the National Institute ofAllergyandInfectious Diseases.
LITERATURECITED
1. Brown, S. M., D. A. Ritchie, and J. H. Subak-Sharpe. 1973. Genetic studieswithherpessimplex virustype 1. Theisolation oftemperature-sensitive mutants, theirarrangement into com-plementation groups and recombination analysis leading to a linkagemap. J.Gen. Virol. 13:329-346.
2. Burnette,W. N.1981. Westernblotting: electrophoretictransfer ofproteinsfrom sodiumdodecylsulfate-polyacrylamidegelsto unmodified nitrocellulose and radiographic detection with an antibody and radioiodinated protein A. Anal. Biochem. 112:195-203.
3. Clements,J. B.,R.J.Watson,and N. M. Wilkie.1977.Temporal regulationofherpessimplexvirustype 1transcription: localiza-tion oftranscriptsontheviralgenome. Cell 12:275-285. 4. Coen,D.M.,and P. A.Schaffer. 1980.Twodistinctlociconfer
resistance to acycloguanosine in herpes simplex virus type 1. Proc. Natl. Acad. Sci. USA 77:2265-2269.
5. Cordingly,M.G.,M. E. M.Campbell,andC. M. Preston. 1983. Functionalanalysis ofaherpes simplexvirustype 1 promoter: identification of far-upstream regulatory sequences. Nucleic Acids Res. 11:2347-2365.
6. Courtney, R. J., and M. Benyesh-Melnick. 1974. Isolation and characterization of a large molecular-weight polypeptide of
herpes simplexvirustype 1. Virology62:539-551.
7. Davidson, I.,and N. Stow. 1985. Expression ofan immediate
early polypeptide and activation of a viral origin of DNA replicationin cellscontainingafragmentofherpessimplexvirus DNA. Virology141:77-88.
8. DeLuca,N. A.,M. A.Courtney,and P.A. Schaffer. 1984.Tem-perature-sensitivemutantsinherpes simplexvirustype 1ICP4
permissiveforearlygeneexpression. J.Virol. 52:767-776. 9. DeLuca, N., S. Person, D. J. Bzik, and W. Snipes. 1984.
Location oftemperaturesensitivemutantsinglycoprotein gB of
herpes simplexvirustype 1. Virology137:382-389.
10. DeLuca,N.A.,and P. A.Schaffer.1985. Activationof
immedi-ate-early, early, and late promoters by temperature-sensitive and wild-type forms of herpes simplex virus type 1 protein ICP4. Mol. Cell. Biol. 5:1997-2008.
11. Dixon, R. A. F., and P. A. Schaffer. 1980. Fine-structure
mapping and functionalanalysis oftemperature-sensitive mu-tants in the gene encoding the herpes simplex virus type 1 immediateearlyprotein VP175.J. Virol.36:189-203.
12. Everett, R. D.1984. Transactivation oftranscription by herpes virus product: requirement for two HSV-1 immediate-early
polypeptidesfor maximumactivity. EMBO J. 3:3135-3141. 13. Goldberg, D. A. 1980. Isolation and partial characterization of
theDrosophilaalcoholdehydrogenasegene. Proc. Nati.Acad. Sci. USA77:5794-5798.
14. Goldin, A. L., R. M. Sandri-Goldin, M. Levine, and J.C. Glorioso. 1981. Cloning of herpes simplex virus type 1 se-quencesrepresenting the wholegenome.J. Virol. 38:50-58. 15. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982.
Recombinant genomeswhich express chloramphenicol acetyl-transferase in mammalian cells. Mol. Cell. Biol.2:1044-1051. 16. Graham,F.L.,and A.J.vanderEb.1973. A newtechniquefor
theassayofinfectivityofhuman adenovirus 5 DNA. Virology 52:456-467.
17. Hayward, G.,R.J.Jacob,S. C. Wadsworth,andB.Roizman. 1975.Anatomyofherpes simplexvirus DNA: evidence for four populationsofmolecules that differ in the relativeorientations of their longand short segments. Proc. Natl. Acad. Sci. USA 72:4243-4247.
18. Holland,T.C.,S. D.Marlin,M. Levine,andJ.Glorioso. 1983. Antigenicvariants ofherpes simplex virusselected with
glyco-protein-specific monoclonalantibodies. J. Virol.45:672-682. 19. Honess,R.W.,and B.Roizman.1974.Regulationofherpesvirus
macromolecular synthesis.I.Cascaderegulationofthe synthe-sis ofthreegroups of viralproteins.J. Virol. 14:8-19. 20. Honess,R.W.,and B.Roizman.1975.Regulationofherpesvirus
macromolecularsynthesis: sequential transition ofpolypeptide
synthesis requires functional viral polypeptides. Proc. Natl. Acad. Sci. USA72:1276-1280.
21. Johnson, D. A., J. W. Gautsch, J. R. Sportsman, and J. H. Elder. 1984. Improved technique utilizing nonfat dry milk for analysisofproteins and nucleic acids transferred to nitrocellu-lose. Gene Anal.Tech. 1:3-8.
22. Jones, N.,and T.Shenk.1979. Anadenovirustype 5early gene function regulates expressionofother viral genes. Proc. Natl. Acad. Sci. USA76:3665-3669.
23. Mackem, S., and B. Roizman. 1982. Structural features ofthe herpes simplex virus a. gene 4, 0, and 27 promoter-regulatory sequences which confer a regulation of chimeric thymidine kinase genes. J. Virol.44:939-949.
24. Maniatis, T.,E. F. Fritsch,andJ. Sambrook. 1982. Molecular cloning: alaboratory manual, p. 191-193. Cold SpringHarbor Laboratory,ColdSpring Harbor,N.Y.
25. Maniatis, T., A. Jeffrey, and D. G. Kleid. 1975. Nucleotide sequence of the rightward operator of phage X. Proc. Natl. Acad. Sci. USA72:1184-1188.
26. Marsden,H.S.,N. D.Stow,V.G. Preston, M. C.Timbury,and N. M.Wilkie. 1978. Physical mappingofherpes simplex virus-inducedpolypeptides. J. Virol.28:624-642.
27. Mocarski,E.S.,and B. Roizman.1982.Structureand roleofthe herpes simplexvirus DNA termini ininversion,circularization, andgeneration ofvirion DNA. Cell 31:89-97.
28. Morse, L.S.,L.Pereira,B.Roizman,andP. A.Schaffer. 1978. Anatomy ofherpes simplex virus(HSV) DNA X. Mappingof viralgenes byanalysis of polypeptides andfunctions specified by HSV-1 x HSV-2recombinants. J. Virol. 26:389-410. 29. Notarianni, E. L., and C. M. Preston. 1982. Activation of
cellular stressprotein genes by herpes simplex virus tempera-ture sensitive mutants which overproduce immediate-early polypeptides. Virology 123:113-122.
30. O'Hare, P.,andG. S.Hayward.1985.Evidencefor adirectrole forboth the175,000- and 110,000-molecular-weight immediate-early proteins ofherpessimplex virusin the transactivationof delayed-earlypromoters. J.Virol. 53:751-760.
31. Pancake, B. A., D. P. Aschman, and P. A. Schaffer. 1983. Genetic andphenotypic analysis of herpessimplexvirustype 1 mutants conditionally resistant to immune cytolysis. J. Virol. 47:568-585.
32. Persson, R. H., S. Bacchetti, and J. R. Smiley. 1985. Cellsthat constitutivelyexpresstheherpes simplex virus immediate-early protein ICP4 allow efficient activation of viral delayed-early genesintrans.J. Virol. 54:414-421.
33. Poffenberger, K. L., E. Tabares, and B. Roizman. 1983. Char-acterization ofa viable, noninverting herpes simplex virus 1 genomederived by insertion and deletion of sequencesat the junction ofcomponents L and S. Proc. Natl. Acad. Sci. USA VOL.56, 1985
on November 10, 2019 by guest
http://jvi.asm.org/
80:2690-2694.
34. Post,L. E., and B.Roizman. 1981. A generalized technique for deletion of specific genes in large genomes: a gene 22 of herpes simplex virus type 1 is not essential for growth. Cell 25:227-232. 35. Preston, C. M. 1979.Abnormal properties ofanimmediateearly polypeptide in cells infected with herpes simplex virus type 1 mutant tsK. J. Virol.32:357-369.
36. Preston, C. M. 1979. Control of herpes simplex virus type 1 mRNA synthesis in cells infected with wild-type virus or the temperature-sensitive mutant tsK. J. Virol. 29:275-284. 37. Preston, V. G. 1981. Fine-structure mapping of herpes simplex
virus type 1 temperature-sensitive mutations within the short repeat region of thegenome. J. Virol. 39:150-161.
38. Preston, V. G., A. J. Davison, H. S. Marsden, M. C. Timbury, J. H. Subak-Sharpe, and N. M. Wilkie. 1978. Recombinants between herpes simplex virus types1 and 2:analyses of genome structures and expression of immediate early polypeptides. J. Virol.28:499-517.
39. Quinlan,M.P., and D.M.Knipe. 1985.Stimulation ofaherpes simplex virus DNA-binding protein by two viral functions. Mol. Cell. Biol. 5:957-963.
40. Sacks, W. R., C. C. Greene,D. P.Aschman, and P. A. Schaffer. 1985.Herpessimplex virus type 1ICP27 isanessential regula-toryprotein. J.Virol. 55:796-805.
41. Sandri-Goldin, R. M., A. L. Goldin, L. E. Holland, J. C. Glorioso, and M. Levine. 1983. Expression ofherpes simplex virus p and y genes integrated in mammalian cells and their induction by anageneproduct. Mol. Cell. Biol. 3:2028-2044. 42. Schaffer, P. A., G. M. Aron, N. Biswal, and M.
Benyesh-Melnick. 1973. Temperature-sensitive mutants of herpes simplex virus type 1: isolation, complementation and partial characterization. Virology 52:57-71.
43. Schaffer, P. A., V. C. Carter, and M. C. Timbury. 1978. Collaborative complementation study oftemperature-sensitive mutants of herpes simplex virus types 1 and 2. J. Virol. 27:490-504.
44. Schroder,C. H.,J.DeZazzo, K.W.Knopf,H. C.Kaerner,M. Levine, andJ. Glorioso. 1985. A herpes simplex virus type 1 mutant withadeletion in thepolypeptide-codingsequences of theICP4 gene. J.Gen. Virol. 66:1589-1593.
45. Southern, E. M. 1975. Detection ofspecificsequences among DNAfragments separated by gel electrophoresis. J. Mol. Biol. 98:503-517.
46. Southern,P.J.,and P.Berg. 1982. Transformation of mamma-lian cells to antibiotic resistance with abacterial gene under control ofthe SV40early region promoter.J.Mol.Appl. Genet. 1:327-341.
47. Stow,N. D.,andC. McMonagle. 1983. Characterization ofthe TRS/IRsorigin ofDNAreplication of herpes simplex virus type 1.Virology130:427-438.
48. Towbin,H.,T.Staehelen,andJ.Gordon.1979.Electrophoretic
transfer of proteins from polyacrylamide gels to nitrocellulose sheets:procedure andsomeapplications. Proc. Natl. Acad. Sci. USA 76:4350-4354.
49. Watson, R. J., and J. B. Clements. 1978. Characterization of transcription-deficient temperature-sensitive mutantsofherpes simplexvirustype 1.Virology91:364-379.
50. Watson, R. J., C. M. Preston, and J. B. Clements. 1979. Separation and characterization of herpes simplex virustype1 immediate-early mRNA's.J.Virol. 31:42-52.
51. Weller, S. K., K. J. Lee, D.J. Sabourin, and P. A. Schaffer. 1983.Genetic analysis oftemperature-sensitive mutants which define the gene for the major herpes simplex virus type 1 DNA-binding protein.J. Virol. 45:354-366.