Copyright © 1998, American Society for Microbiology. All Rights Reserved.
Inhibition of Simian Immunodeficiency Virus (SIV) Replication by
CD8
1
T Lymphocytes from Macaques Immunized
with Live Attenuated SIV
MARIE-CLAIRE GAUDUIN,1RHONA L. GLICKMAN,1ROBERT MEANS,2 ANDR. PAUL JOHNSON1,3*
Divisions of Immunology1and Microbiology,2New England Regional Primate Research Center,
Harvard Medical School, Southborough, Massachusetts 01772-9102, and Infectious Disease Unit and Partners AIDS Research Center, Massachusetts
General Hospital, Boston, Massachusetts 021153
Received 8 January 1998/Accepted 21 April 1998
Characterization of immune responses induced by live attenuated simian immunodeficiency virus (SIV) strains may yield clues to the nature of protective immunity induced by this vaccine approach. We investigated the ability of CD81T lymphocytes from rhesus macaques immunized with the live, attenuated SIV strain
SIVmac239Dnef or SIVmac239D3 to inhibit SIV replication. CD81T lymphocytes from immunized animals
were able to potently suppress SIV replication in autologous SIV-infected CD41T cells. Suppression of SIV
replication by unstimulated CD81T cells required direct contact and was major histocompatibility complex
(MHC) restricted. However, CD3-stimulated CD81T cells produced soluble factors that inhibited SIV
repli-cation in an MHC-unrestricted fashion as much as 30-fold. Supernatants from stimulated CD81T cells were
also able to inhibit replication of both CCR5- and CXCR4-dependent human immunodeficiency virus type 1 (HIV-1) strains. Stimulation of CD81cells with cognate cytotoxic T-lymphocyte epitopes also induced secretion
of soluble factors able to inhibit SIV replication. Production of RANTES, macrophage inhibitory protein 1a
(MIP-1a), or MIP-1bfrom stimulated CD81T cells of vaccinated animals was almost 10-fold higher than that
from stimulated CD81 T cells of control animals. However, addition of antibodies that neutralize these
b-chemokines, either alone or in combination, only partly blocked inhibition of SIV and HIV replication by soluble factors produced by stimulated CD81T cells. Our results indicate that inhibition of SIV replication by
CD81T cells from animals immunized with live attenuated SIV strains involves both MHC-restricted and
-unrestricted mechanisms and that MHC-unrestricted inhibition of SIV replication is due principally to soluble factors other than RANTES, MIP-1a, and MIP-1b.
Efforts to develop an effective AIDS vaccine have been thwarted by a number of factors, including our incomplete understanding of the specific immune responses involved in protective immunity. Analysis of immune responses induced by vaccination of macaques with live attenuated simian immuno-deficiency virus (SIV) strains, an approach that has yielded the most consistent protection to date in the SIV-macaque model (11), offers an opportunity to begin to identify humoral and cellular immune responses that may play a role in mediating protection against infection. Studies of animals vaccinated with live attenuated SIV strains have demonstrated the presence of SIV-specific cytotoxic T-lymphocyte (CTL) responses (22, 52), proliferative responses (13), and neutralizing antibodies (11, 51). However, the relative contribution of each responses in mediating protective immunity remains to be determined.
In human immunodeficiency virus type 1 (HIV-1)-infected subjects, CD81T cells are believed to play a major role in
in-hibiting viral replication (40, 54). Two types of CD81
T-cell-mediated HIV-specific responses have been described: a cyto-toxic mechanism mediated by CTL that lyse HIV-1-expressing target cells in a major histocompatibility complex (MHC) class I-restricted manner, and a noncytotoxic mechanism mediated
by soluble suppressive factors secreted by T lymphocytes from HIV-infected subjects (5, 48–50). As initially demonstrated by Walker et al. (49) and subsequently confirmed by others (5, 23, 47), CD81 T cells are capable of suppressing in vitro HIV
replication in CD41T cells in a noncytolytic, MHC class
I-un-restricted manner (for reviews, see references 32 and 50). The role of CD81 T-cell-derived soluble factors in suppressing
HIV infection in vivo is not known. However, detection of this activity in HIV-infected long-term nonprogressors (35, 37, 50) and in acutely infected subjects prior to the detection of neu-tralizing antibodies (38), and its decline in subjects with ad-vanced disease (30), all suggest that soluble factors produced by CD81T cells may contribute to suppression of HIV in vivo.
Initial studies indicated that soluble factors able to inhibit HIV replication were not related to known cytokines, including alpha, beta, and gamma interferons, tumor necrosis factor alpha, interleukin-1 (IL-1), IL-2, IL-4, IL-6, or IL-12 (36). Recently, Cocchi et al. (10) reported that antiviral effects of supernatants from CD81T cells were primarily due to the
chemoattractant cytokines (b-chemokines) RANTES (regulat-ed on activation, normal T-cell express(regulat-ed and secret(regulat-ed), mac-rophage inhibitory protein 1a(MIP-1a), and MIP-1b. Inhibi-tion of HIV-1 replicaInhibi-tion by theseb-chemokines appears to occur in part by their ability to interfere with viral entry via the CCR5 coreceptor (1, 7, 12, 14). However, the presence of RANTES, MIP-1a, and MIP-1bin CD81T-cell
superna-tants from HIV-specific CTL clones does not appear to corre-late with the ability of these supernatants to inhibit HIV, and * Corresponding author. Mailing address: Division of Immunology,
New England Regional Primate Research Center, Harvard Medical School, One Pine Hill Dr., P.O. Box 9102, Southborough, MA 01772. Phone: (508) 624-8148. Fax: (508) 624-8172. E-mail: pjohnson@war ren.med.harvard.edu.
6315
on November 9, 2019 by guest
http://jvi.asm.org/
cells from animals vaccinated with live attenuated SIV strains to inhibit SIV replication. In this study, we characterized the ability of CD81 T lymphocytes from macaques immunized
with SIVmac239Dnef or SIVmac239D3 to inhibit SIV replica-tion. The potential role ofb-chemokines RANTES, MIP-1a, and MIP-1bin mediating CD81T-lymphocyte antiviral activity
in this model was also evaluated. CD81T cells from vaccinated
animals were able to inhibit SIV replication when in direct contact with infected cells and to produce soluble factors able to inhibit viral replication. Although stimulated CD81T cells
from animals immunized with live, attenuated SIV produced increased amounts of RANTES, MIP-1a, and MIP-1b, these b-chemokines did not appear to mediate the dominant effect of CD81T-cell-derived factors able to suppress SIV and HIV
replication.
MATERIALS AND METHODS
Animals.Rhesus macaques used in this study were housed at the New England Regional Primate Research Center. Three groups of SIV-infected macaques were studied: (i) five animals infected with the live attenuated SIV strain SIVmac239Dnef (26) 8 to 9 years prior to our studies; (ii) five animals infected with SIVmac239D3 (deficient in nef, vpr, and the negative regulatory elements of the long terminal repeat [LTR]) 5 to 6 years prior to our studies; and (iii) two animals infected with pathogenic strain SIVmac239 (25) or SIVmac251. At the time of the study, the two animals infected with a pathogenic SIV strain had relatively advanced disease, with CD4 counts of#300 mm3and virus loads of
123103and 10.83105copies/ml, respectively. In addition, four other macaques
from the conventional colony that were seronegative for SIV were used as control animals. All animals were maintained in accordance with the guidelines of the local institutional animal use committees and the federal government (47a). Three of the five rhesus macaques infected with SIVmac239Dnef were challenged 2 years later either with cloned pathogenic SIVmac239/nef-open (intact nef) (animals 353.91 and 397.88) or with pathogenic SIVmac251 (animal 71.88). Of macaques immunized with SIVmac239D3, two animals (358.91 and 437.91) were challenged 2 years later with uncloned pathogenic SIVmac251. All challenged macaques remained healthy without evidence for wild-type SIV in-fection at time of our study.
Cell lines.C8166-45, CEMx174, and PM1 (34) cells were maintained in RPMI 1640 medium (Sigma, St. Louis, Mo.) supplemented with 10% heat-inactivated fetal calf serum (Sigma), 10 mM HEPES 2 mML-glutamine, 50 U of penicillin per ml, and 50mg of streptomycin per ml (R-10 medium). C8166-45-SEAP and CEMx174-SEAP cell lines expressing secreted alkaline phosphatase (SEAP) under the control of the SIV LTR have been previously described (41). Stimu-lator cells consisted of autologous or allogeneic herpesvirus papio-transformed B-cell lines (B-LCL).
Virus stocks.The SIV isolates used in this study were SIVmac239, derived from the pathogenic molecular clone SIVmac239 (25), and SIVmac251, a patho-genic virus stock that has been passaged only in rhesus peripheral blood mono-nuclear cells (PBMC). In addition, three HIV-1 isolates were used: HIV-1Ba-L, a
monocytotropic primary HIV-1 isolate; HIV-1JR-CSF, a molecularly cloned
pri-mary HIV-1 isolate (29) that demonstrates many of the characteristics of a primary HIV isolate and will not grow in most transformed T-cell lines (29); and HIV-1NL4-3, which is derived from a molecular clone that grows well in
trans-formed T-cell lines.
Each virus stock was prepared from the supernatant of either macaque PBMC
by using a magnetic separation device (Dynal). Similarly, CD41lymphocytes were obtained by depleting CD81cells by using an anti-CD8 (51.1; American Type Culture Collection catalog no. HB230) antibody at 20mg/106cells and then
incubating them with anti-IgG2a-coated magnetic beads (Dynal) at a 10:1 bead-to-cell ratio for 60 min at 4°C. The CD41cell-enriched supernatant was then collected by using a magnetic separation device (Dynal). Finally, CD41cells were stimulated in R-10 medium supplemented with ConA (5mg/ml) overnight (37°C, 5% CO2) before infection with SIV. After cell separation, CD81cell
populations were greater than 85% CD81and contained,5% CD41cells as assessed by flow cytometry, and CD41T lymphocytes were greater than 95% CD41, with less than 1% residual CD81cells.
Transmembrane assay for viral inhibition.CD41T cells were stimulated with ConA (5mg/ml) for 24 h and then infected with SIVmac239 at a multiplicity of infection (MOI) of 0.01 TCID50/cell. A total of 13106to 1.53106CD41T cells
per well were placed in a 24-well plate (lower well) and overlaid with a 0.4m m-pore-size semipermeable membrane insert (Millipore, Bedford, Mass.). Within the insert (upper well) were placed 23106CD81T cells (MHC matched or mismatched) stimulated with 63106goat anti-mouse IgG-coated beads
(Per-Septive Diagnostics) previously saturated with the mouse rhesus CD3 anti-body (6G12; hybridoma provided by J. Wong, Massachusetts General Hospital, Boston) (24). Lower and upper wells contained a total of 1.4 and 0.7 ml, respec-tively, of 10% IL-2-supplemented RPMI 1640. Controls included CD81T cells mixed directly with the infected CD41T cells in the lower well, CD81T cells not stimulated by CD3-specific beads in the upper well, and CD3-specific beads alone in the upper well. Plates were incubated at 37°C for 10 days. At day 4, 6, 8, and 10, half of the supernatant fluid was removed from the lower well, analyzed for p27 antigen (Ag) quantitation by standard quantitative SIV p27 Ag enzyme-linked immunosorbent assay (ELISA; Coulter), and cryopreserved at
280°C for later use. Cryopreserved supernatant fluids from stimulated or un-stimulated CD81T cells were tested for concentrations of RANTES, MIP-1a, and MIP-1b by using human-specific ELISA kits known to cross-react with rhesus chemokines (R&D Systems, Minneapolis, Minn.).
Generation of soluble factors from CD81T lymphocytes activated by specific peptides.Soluble factors were generated as described above, using the trans-membrane assay. CD81T cells from the upper well were stimulated by specific peptides. Briefly, CD81T cells were cocultured at 23106cells/ml in the upper
well with 106autologous B-LCL labeled with specific epitope peptide. These B
cells had been labeled with 100mg of specific peptide per ml, washed twice, and gamma irradiated (10,000 rads). At day 4, 6, 8, and 10, supernatant fluid from the lower well was harvested and cryopreserved at280°C for later use. Controls included cell-free supernatants from autologous B cells alone, autologous B cells labeled with unrelated peptide, and allogeneic B cells labeled with specific peptide. Peptides used for these experiments (10mg/well) included the Mamu-A*01-restricted SIV Gag peptide 11C (TPYDINQML) (42), the SIV Gag pep-tide 7G (HQAAMQIIR) (19a), and the unrelated peppep-tide 12K (NYSETDRWG) (16).
SEAP assay.The SEAP assay has been described previously (41) and was performed as follows. Briefly, 100ml of CEMx174-SEAP or C8166-45-SEAP cells was first plated in triplicate in 96-well plates at 0.43106cells/ml of R-10
medium. Each well received 50ml of either control antibody, chemokine, or soluble factor fluid. SEAP cells were then infected by addition of 50ml of virus (SIVmac239 or SIVmac251) at the indicated concentrations. In addition, the CEMx174-SEAP cell line was cultured under the same conditions to analyze the ability of either control antibody, chemokine, or soluble factor fluid to inhibit HIV-1NL4-3replication. The plates were then incubated at 37°C for 72 to 96 h in
a 5% CO2incubator. SEAP activity was monitored at several time points
postin-fection by harvesting supernatant from each well and by using a Phospha-Light assay kit as described previously (41). Generally three types of controls were included on each plate: CEMx174 or C8166-45 cells as a background control; SIV-infected CEMx174 (or C8166-45)-SEAP cells for measurement of basal
on November 9, 2019 by guest
levels of SEAP production; and virus-infected CEMx174 (or C8166-45)-SEAP cells alone to measure SEAP expression in the absence of supernatant.
Assay for inhibition of HIV-1 replication by chemokines or supernatants from stimulated CD81T cells.PM1 cells were acutely infected with various dilutions of HIV-1Ba-Lor HIV-1JR-CSF, resuspended in RPMI 1640 supplemented with
20% fetal calf serum, and plated at 2.53106cells per well in a 24-well plate.
CD81T-cell supernatants were then tested at a final dilution of 1:2. The che-mokines RANTES, MIP-1a, and MIP-1b(PeproTech, Inc., Rocky Hill, N.J.) were also tested for antiviral activity alone or in combination at different con-centrations. In some experiments, CD81T cells, supernatants, or chemokines were first incubated at room temperature for 30 min with polyclonal neutralizing antibodies to RANTES, MIP-1a, and MIP-1b(R&D Systems) (separately or in combination) and then added to acutely infected PM1 cells. Supernatant fluid (0.5 ml) was removed on day 5 and 7 for p24 measurement and replaced with fresh medium.
RESULTS
CD81 T lymphocytes from macaques immunized with
SIVmac239Dnef or SIVmac239D3 inhibit SIV replication. In initial experiments, we investigated the ability of CD81T
lym-phocytes isolated from macaques infected with either an at-tenuated SIV strain (SIVmac239Dnef or SIVmac239D3) or a pathogenic strain (SIVmac251 or SIVmac239) to inhibit SIV replication. Inhibition of SIV replication was assessed by a transmembrane assay where CD41 T lymphocytes were
in-fected in vitro with SIVmac239 (MOI of 0.01 TCID50per cell) and then cultured either in direct contact with autologous CD81 T lymphocytes or separated from them by a 0.4-m
m-pore-size membrane. CD81T cells not in direct contact with
CD41T cells were either unstimulated or activated by
CD3-specific antibodies bound to paramagnetic beads. SIV rep-lication was then assessed by SIV p27 Ag ELISA at multiple time points during a 10-day period. The results presented in Fig. 1 are representative of experiments using cells from five SIVmac239Dnef-infected animals, five SIVmac239D3-infected animals, two wild-type SIV-infected animals, and four unin-fected animals. CD41 T lymphocytes from animals infected
with attenuated SIV strains were readily infected with SIV in vitro, resulting in peak levels of viral replication that did not differ significantly from those obtained for CD41T cells from
normal controls (Fig. 1A and data not shown). When in direct contact with infected CD41 T cells, CD81T cells from
ma-caques immunized with SIVmac239Dnef or SIVmac239D3 were able to potently inhibit SIV replication (Fig. 1A). In ad-dition, CD3-stimulated CD81T lymphocytes from immunized
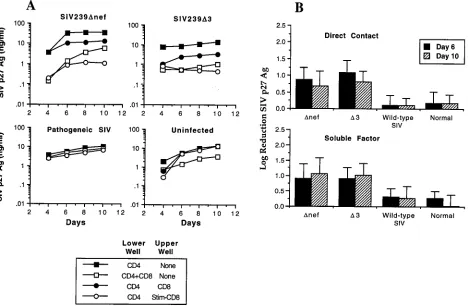
[image:3.612.70.538.71.377.2]animals were able to secrete soluble factors that inhibited viral replication by 1 log or more in autologous SIV-infected CD41 FIG. 1. Inhibition of SIV replication by CD81T lymphocytes from macaques immunized with SIVmac239Dnef or SIVmac239D3. (A) Transmembrane experiments
using PBMC from macaques infected with either wild-type SIV or live attenuated SIV and from uninfected controls. PBMC were separated into CD41and CD81
populations by using immunomagnetic beads. CD41T cells were ConA stimulated overnight and acutely infected with SIVmac239 at an MOI of 0.01 TCID
50/cell. Infected CD41T cells were then cultured in the lower well either alone, in direct contact with CD81T cells, or with stimulated or unstimulated CD81T cells placed
in the upper well. Inhibition of SIV replication by CD81T cells was assessed by measuring the production of SIV p27 Ag over a period of 10 days. Representative assays
are shown for individual animals infected with the indicated SIV strain and for an uninfected control. (B) Quantitative analysis of suppression of SIV replication by CD81T lymphocytes from uninfected animals and animals infected with pathogenic or live attenuated strains of SIV. Experiments were carried out as described above,
and log reduction of SIV replication (90% inhibition51 log, 99%52 logs, etc.) was analyzed on days 6 and 10. The data represent the means6standard deviations for a total of 16 animals: 5 SIVmac239Dnef-, 5 SIVmac239D3-, and 2 wild-type (SIVmac239 and SIVmac251)-infected animals and 4 uninfected animals. Suppression of SIV replication by CD81T cells from animals infected with either SIVmac239Dnef or SIVmac239D3 was statistically significant compared with inhibition by either
wild-type SIV-infected animals or normal controls (P,0.01), for both cells in direct contact and for soluble factors. The difference between wild-type SIV-infected animals and normal controls was not significant.
on November 9, 2019 by guest
http://jvi.asm.org/
T cells (Fig. 1A). The addition of CD3-specific immunomag-netic beads alone to the upper well had no effect on SIV replication (data not shown). Although we occasionally ob-served a low level inhibition of replication in PBMC from uninfected animals, comparison of results from multiple ani-mals showed that the level of inhibition of SIV replication by CD81T cells from macaques infected with live attenuated SIV
strains was 30-fold greater than that for normal controls (Fig. 1B) for cells in direct contact with infected CD41T cells and
20-fold greater for soluble factors produced by activated T cells (Fig. 1B) (P,0.01). Thus, immunization of macaques with live attenuated SIV strains induces a specific CD81T-cell response
that is able to inhibit SIV replication and was significantly greater than that found in animals infected with wild-type SIV or normal controls.
Suppression of SIV replication by unstimulated CD81 T
lymphocytes is MHC restricted.In the next series of experi-ments, we examined whether inhibition of SIV replication by CD81T cells from vaccinated animals in direct contact with
CD41T cells was MHC restricted. Acutely infected CD41T
cells from SIVmac239Dnef- or SIVmac239D3-immunized ani-mals were incubated in direct contact with either autologous or allogeneic CD81T cells. Allogeneic donors were prescreened
by isoelectric focusing of MHC class I alleles so as to minimize class I homology (22). As shown in Fig. 2, allogeneic CD81T
lymphocytes (MHC mismatched) from each of three different immunized macaques were not effective in suppressing SIV replication in CD41T lymphocytes, whereas autologous CD81
T cells efficiently suppressed viral replication in each instance. Therefore, suppression of SIV replication by CD81T
lympho-cytes in direct contact with CD41cells was MHC restricted.
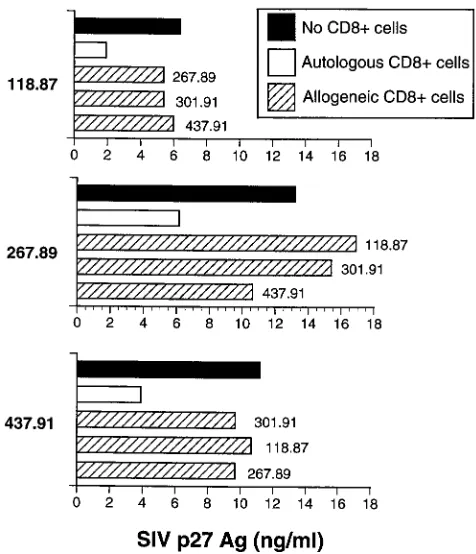
T cells was mediated, at least in part, by CTL that required MHC-restricted Ag presentation in order to be activated. Pre-vious analysis of SIVmac239Dnef-immunized macaques has demonstrated that these animals develop a strong CTL re-sponse against gag- and env-expressing target cells (22). Recent mapping of CTL epitopes recognized by these animals has identified several distinct epitopes (23a), including a SIV Gag peptide previously shown to be presented by the Mamu-A*01 MHC class I allele (42). To determine whether peptide-specific stimulation of CTL could elicit production of soluble factors able to inhibit SIV replication, CD81T lymphocytes from
SIVmac239Dnef-immunized macaques (animals 267.89 and 118.87) were stimulated with autologous B-LCL sensitized with either an immunodominant CTL epitope (7G for animal 267.89; 11C for animal 118.87) or, as a control, a previously identified SIV envelope CTL epitope (16). Following stimula-tion with the cognate CTL epitope but not the irrelevant con-trol peptide 12K, CD81T cells from animals immunized with
SIVmac239Dnef were able to secrete soluble factors that in-hibited SIV replication 3- to 10-fold (Fig. 4) (3). Since these experiments were performed, Allen et al. (3) have revised the optimal Gag epitope presented by Mamu-A*01 to include one additional NH2-terminal amino acid (CTPYDINQML instead of TPYNINQML). However, since the concentration of pep-tide used for these experiments (10mg/ml) results in maximal levels of target cell lysis when bulk effector cells are used, we feel it unlikely that use of the optimal peptide would alter these results. In subsequent experiments, autologous CD81T cells
[image:4.612.50.288.68.345.2]from either SIVmac239Dnef- or SIVmac239D3-immunized
FIG. 2. Suppression of SIV replication by unstimulated CD81T lymphocytes
is MHC restricted. Acutely SIV infected CD41T cells were cocultured in direct
contact with MHC class I-mismatched allogeneic CD81T lymphocytes isolated
from uninfected animals or animals infected with pathogenic or live attenuated strains of SIV. Inhibition of SIV replication by allogeneic CD81T cells was
assessed by measuring the production of SIV p27 Ag for 10 days.
FIG. 3. Suppression of SIV replication by soluble factors secreted from CD3-activated CD81T lymphocytes is not MHC restricted. CD41T cells from
animals (118.87 and 267.89) immunized with SIVmac239Dnef were infected with SIVmac239 and then cultured (lower well) with autologous (E) and allogeneic
(Œ) CD81T cells (mismatched; 118.87 and 267.89) stimulated with CD3-specific
beads. Similar data were obtained in two independent experiments.■, no CD81
cells.
on November 9, 2019 by guest
macaques were stimulated with autologous B-LCL infected with a recombinant vaccinia virus expressing gag, pol, and env and then inactivated with UV-psoralen (22). Following stimu-lation with B-LCL expressing SIV Ag, CD81T cells from both
animals were also able to inhibit SIV replication three- to five-fold compared with cells stimulated with control vaccinia virus-infected B-LCL (data not shown). Thus, SIV-specific CTL are able to produce soluble factors which inhibit SIV replication. Stimulated CD81T cells from macaques immunized with
live attenuated SIV strains produce increased concentrations of RANTES, MIP-1a, and MIP-1b.The recent demonstration that chemokine receptors serve as coreceptors for entry of HIV and SIV (6, 12, 14, 17, 18, 33, 39) has helped to elucidate the role of chemokines in inhibiting viral replication. In par-ticular, the chemokines RANTES, MIP-1a, and MIP-1bhave been identified as being in part responsible for the inhibition of CCR5-dependent HIV strains by soluble factors produced by CD81T cells (10). We therefore assessed whether CD81T
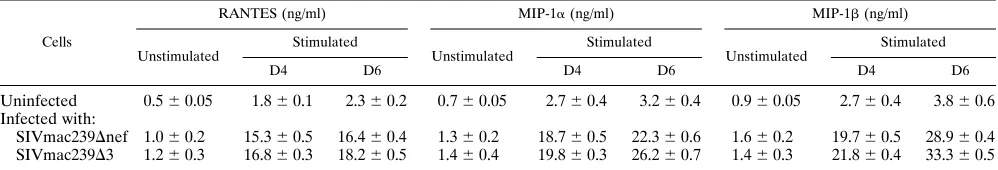
lymphocytes from macaques immunized with live attenuated SIV strains were able to produce these soluble factors after CD3 stimulation. The levels of RANTES, MIP-1a, and MIP-1b were measured in supernatants from unstimulated and CD3-stimulated CD81T cells from normal and immunized
macaques (Table 1). Four to six days after stimulation, peak levels of the threeb-chemokines were six- to ninefold higher in CD81T cells from animals vaccinated with SIVmac239Dnef or
SIVmac239D3 compared with normal controls (Table 1). Sim-ilar increase ofb-chemokines were observed at days 8 and 10 after stimulation (data not shown).
RANTES, MIP-1a, and MIP-1bmediate only low-level in-hibition of SIV replication in the C8166 and CEMx174 cell lines compared to soluble factors produced by CD81T cells
from SIVmac239Dnef- and SIVmac239D3-infected macaques. To facilitate characterization of soluble factors responsible for inhibition of virus replication, we wanted to establish a repro-ducible and quantitative system that provided a relatively rapid assessment of inhibition of viral replication. To ac-complish this, we used two cell lines (C8166-45 and CEMx174) that stably express SEAP under the control of the SIV LTR (41). Infection of these cell lines with SIV results in a dose-dependent release of SEAP over a 2- to 3-log range, allowing rapid quantitation of the level of SIV infection 72 to 96 h after infection (41). We first evaluated the ability of soluble factors from CD3-stimulated CD81T cells from macaques immunized
with SIVmac239Dnef or SIVmac239D3 to inhibit replication in this system. C8166-45-SEAP cells were preincubated for 1 h at 37°C with soluble factors secreted by CD3-stimulated CD81T
lymphocytes from either SIVmac239Dnef- or SIVmac239D 3-immunized macaques. SIV replication, as assessed by SEAP release, was inhibited up to 10-fold by the addition of soluble factors secreted by activated CD81T lymphocytes from ma-FIG. 4. Stimulation of CD81T cells with cognate CTL epitopes results in production of soluble factors that suppress SIV replication. Autologous CD41T cells
were acutely infected with SIVmac239 (lower well) and cocultured with stimulated CD81T cells from animals (267.89 and 118.87) immunized with SIVmac239Dnef
(upper well). Stimulation of CD81T cells included either CD3-specific immunomagnetic beads or specific peptides (7G for animal 267.89; 11C for animal 118.87)
[image:5.612.138.465.68.247.2]previously shown to represent CTL epitopes for these animals. A previously reported SIV CTL epitope (12K) not recognized by these animals was used as a negative control. All assays were performed in triplicate.
TABLE 1. Production of RANTES, MIP-1aand MIP-1bby CD81T cellsa
Cells
RANTES (ng/ml) MIP-1a(ng/ml) MIP-1b(ng/ml)
Unstimulated Stimulated Unstimulated Stimulated Unstimulated Stimulated
D4 D6 D4 D6 D4 D6
Uninfected 0.560.05 1.860.1 2.360.2 0.760.05 2.760.4 3.260.4 0.960.05 2.760.4 3.860.6 Infected with:
SIVmac239Dnef 1.060.2 15.360.5 16.460.4 1.360.2 18.760.5 22.360.6 1.660.2 19.760.5 28.960.4 SIVmac239D3 1.260.3 16.860.3 18.260.5 1.460.4 19.860.3 26.260.7 1.460.3 21.860.4 33.360.5 aCD81T lymphocytes at 2.53106to 33106cells/well were activated by CD3 cross-linking. Supernatants from stimulated or unstimulated CD81cells were
harvested after 4 and 6 days (D4 and D6) for ELISA quantitation of RANTES, MIP-1a, and MIP-1b. Supernatants from unstimulated were pooled. Results are expressed as the means6standard deviations of triplicate wells.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.50.549.615.700.2]caques immunized with SIVmac239Dnef or SIVmac239D3 (Fig. 5A).
We next examined the ability of RANTES, MIP-1a, and MIP-1b to inhibit SIV replication in infected C8166-45 and CEMx174 cells. The C8166-45 and CEMx174 cell lines were cultured in the presence of RANTES, MIP-a, or MIP-b(each at 500 ng/ml) for 1 h at 37°C and then infected with the SIVmac251 and SIVmac239, respectively. Even at low viral inocula, no more that twofold inhibition of SIV replication was observed, even when RANTES, MIP-a, and MIP-bwere com-bined together (data not shown). We then evaluated the ability of a range of chemokine concentrations to inhibit viral repli-cation following a relative low viral inoculum. SIV replirepli-cation was inhibited in a dose-dependent manner in both cell lines by addition of RANTES, MIP-1a, or MIP-1b(Fig. 5B). However, in contrast to the 10-fold inhibition observed with soluble fac-tors produced by activated CD81T cells, recombinant
chemo-kines inhibited viral replication only twofold even at the high-est chemokine concentration thigh-ested (500 ng/ml).
Soluble factors from stimulated CD81T cells mediate
po-tent suppression of SIV replication in C8166 and CEMx174 cell lines that is not neutralized by antibodies to b -chemo-kines. We next examined if inhibition of SIV replication by soluble factors from activated CD81T cells from macaques
immunized with either SIVmac239Dnef or SIVmac239D3 could be blocked by antibodies tob-chemokines. Neutralizing
anti-bodies to RANTES, MIP-1a, and MIP-1bwere added to the CD81 T-cell supernatants in concentrations sufficient to
neutralize at least 100 ng of each chemokine per ml. These anti-b-chemokine antibodies consisted of goat polyclonal neu-tralizing antibodies used at a final concentration sufficient to neutralize at least 100 ng of each chemokine per ml. As trols, neutralizing antibodies were also added to known con-centrations of recombinant RANTES, MIP-1a, and MIP-1b (each at 100 ng/ml) either alone or in combination. Both mix-tures were incubated for 30 min at room temperature before addition of SIVmac251-infected C8166-45-SEAP cells. SEAP activity was measured in culture supernatants on day 4 after virus inoculation.
As shown in Fig. 6, the ability of supernatants from stimu-lated CD81T cells to inhibit SIVmac251 replication in the
[image:6.612.57.539.70.372.2]C8166-45-SEAP cell line was not neutralized by adding these antibodies individually or in combination. In contrast, when C8166-45-SEAP cells were incubated with the mixture of re-combinant RANTES, MIP-1a, and MIP-1b 1 h prior to SIVmac251 infection, SIV replication was inhibited 2- to 2.5-fold. However, when neutralizing antibodies tob -chemo-kines were added to the mixture, the inhibition of viral repli-cation was reversed (Fig. 6). These results indicate thatb -che-mokines do not account for the majority of anti-SIV activity observed with CD81T-cell supernatants.
FIG. 5. SIV replication is inhibited by soluble factors from activated CD81T cells from immunized macaques and byb-chemokines RANTES, MIP-1a, and
MIP-1b. (A) Inhibition of SIV replication in SIV-infected C8166-45 cells (SEAP activity) by soluble factors secreted by CD81T cells from macaques immunized with either
SIVmac239Dnef (118.87) or SIVmac239D3 (437.91). C8166-45-SEAP cells were acutely infected with SIVmac251 at decreasing concentrations (from 5.9 to 0.3 ng/ml) and then incubated in the presence of soluble factors (1:2 dilution) secreted by CD3-activated CD81T cells from macaques immunized with either SIVmac239Dnef
(118.87) or SIVmac239D3 (437.91). SEAP activity in cell-free supernatants (Sup.) was measured on day 4 after infection. (B) Inhibition of SIV replication (SEAP activity) in infected C8166-45 and CEMx174 cells byb-chemokines. C8166-45-SEAP and CEMx174-SEAP cell lines were acutely infected with SIVmac251 (0.3 ng/ml) and SIVmac239 (1.47 ng/ml), respectively. Infected cells were then incubated with decreasing concentrations (fivefold dilutions starting from 500 ng/ml) of RANTES, MIP-1a, or MIP-1b. SEAP activity was measured from cell-free medium collected on day 4. All SEAP assays were performed in quadruplicate.
on November 9, 2019 by guest
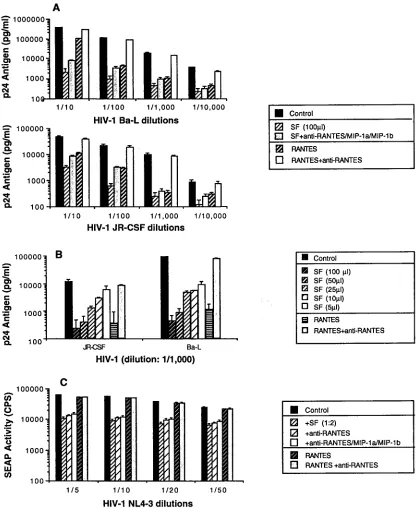
Soluble suppressive factors secreted by CD3-stimulated CD81T cells inhibit HIV-1 replication.We next evaluated the
ability of soluble factors produced from macaque CD81T cells
to inhibit HIV-1 replication. The PM1 cell line described by Cocchi et al. (10), which expresses both CD4 and CCR5, was acutely infected with either HIV-1Ba-L or HIV-1JR-CSF and then cultured in the presence of either soluble factors (1:2 dilution) or RANTES (500 ng/ml). Both CD81T-cell
super-natant and RANTES significantly inhibited HIV-1 replication (Fig. 7A). The ability of soluble factors to inhibit HIV-1 rep-lication was dose dependent (Fig. 7B). In addition, HIV-1Ba-L and HIV-1JR-CSFwere both suppressed by soluble factors pro-duced by activated CD81T cells, even in the presence of a
combination of neutralizing antibodies to the three chemo-kines (RANTES, MIP-1a, and MIP-1b) (Fig. 7A). The small reversion of suppressing activity observed after treatment with theb-chemokine antibodies suggests that elevated concentra-tions ofb-chemokines present in the supernatant contribute to some suppression of HIV-1 replication but do not constitute the major factors involved in the viral inhibition seen.
In addition, the ability of soluble factors produced from macaque CD81T cells to inhibit replication of the
CXCR4-dependent strain HIV-1NL4-3was assessed by using the SEAP assay system. CEMx174-SEAP cells were cultured in the pres-ence of either soluble factors (1:2 dilution) or RANTES (500 ng/ml). Addition of supernatant from stimulated CD81T cells
resulted in a significant inhibition of HIV-1NL4-3replication. As expected, recombinant RANTES (500 ng/ml) had no effect on replication of HIV-1NL4-3and no reversal of inhibition was observed when supernatants from CD81 T cells were
incu-bated in the presence of a combination of neutralizing anti-bodies to the three chemokines (RANTES, MIP-a, and MIP-1b) (Fig. 7C). Thus, CD81 T cells from immunized animals
produced factors distinct from RANTES, MIP-1a, and MIP-1b that are able to inhibit replication of both CXCR4- and CCR5-dependent HIV-1 strains.
DISCUSSION
In this study, we analyzed the ability of CD81T lymphocytes
from rhesus macaques immunized with live attenuated strains of SIV to suppress SIV infection and performed an initial evaluation of the soluble factors produced by these cells. Our data demonstrate that CD81 T lymphocytes from macaques
immunized with live attenuated SIV strains are able to inhibit SIV replication and suggest the existence of both MHC-re-stricted and -unreMHC-re-stricted mechanisms of inhibition. Although a subset of these animals had been challenged with wild-type SIV prior to these studies, no difference in CD81
T-cell-me-diated antiviral activity between challenged and unchallenged animals was observed (data not shown). In addition, there has been no evidence of wild-type SIV infection of these animals more than 3 years after challenge. It is therefore likely that the antiviral CD81T-cell activity was induced by infection with live
attenuated SIV and not the challenge. In contrast to a previous report (47), we did not observe any significant inhibition of SIV replication by CD81T lymphocytes from animals infected with
pathogenic SIV when these cells were cultured in direct con-tact with infected CD41T lymphocytes. Our failure to observe
antiviral activity in wild-type-infected animals may reflect the fact that these animals had relatively advanced disease with high virus load, low CD41T-cell counts (#300 mm3), and poor CTL responses (data not shown). Alternatively, our assay for analyzing the ability of CD81T cells to suppress SIV
replica-tion may be less sensitive than that used by Tsubota et al. (47), a finding that could be due to technical differences between our assays. For instance, we did not stimulate CD81T cells
prior to their incubation in direct contact with CD41T cells,
whereas the previous report used ConA-stimulated CD81cells
(47). In addition, we superinfected CD41T cells with SIV.
Since the initial description of the ability of CD81T
lym-phocytes to suppress HIV replication (49), there has been considerable debate over the relative contribution of cytolytic and noncytolytic mechanisms of suppression. Early reports suggested that this inhibition was primarily mediated by non-cytolytic mechanisms involving the production of soluble fac-tors (35, 38, 49). However, a recent report has demonstrated that HIV-specific CTL clones can inhibit viral replication and, for cells in direct contact, that MHC-restricted mechanisms account for most of the observed inhibition of replication (53). Our results are comparable with those of Yang et al. (53) and suggest that SIV-specific CTL account, at least in part, for the ability of CD81T cells from immunized animals to inhibit viral
replication. This conclusion is supported by the observation that for unstimulated CD81 T cells, optimal inhibition was
MHC restricted and required direct contact. Further demon-stration of the ability of SIV-specific CTL to release soluble factors that inhibit SIV replication comes from our experi-ments in which stimulation of CD81T lymphocytes from
im-munized animals with the cognate CTL epitope results in pro-duction of soluble factors able to inhibit SIV replication. These observations suggest that inhibition of SIV replication is likely to be mediated by CTL that require an MHC-restricted anti-gen presentation in order to be activated. However, once ac-tivated, these CD81 T cells can release soluble factors that
[image:7.612.53.288.66.316.2]inhibit SIV replication in an MHC-unrestricted fashion. At
FIG. 6. Neutralizing antibodies tob-chemokines do not block the ability of soluble factors from CD81T cells to suppress SIV replication. C8166-45-SEAP
cells were infected with SIVmac251 (0.3 ng/ml) and cultured either with super-natants (1:2 dilution) from CD3-activated CD81T cells from a macaque
immu-nized with SIVmac239Dnef or with RANTES, MIP-1a, or MIP-1b(500 ng/ ml) added individually or in combination. In addition, in some cases, neutralizing antisera to human RANTES, MIP-1a, and MIP-1bwere preincubated at 200, 80, and 100mg/ml, respectively, withb-chemokines and then were added to the indicated cultures (cross-hatched bars). Similar data were obtained in two
inde-pendent experiments.
on November 9, 2019 by guest
http://jvi.asm.org/
present, we cannot exclude the existence of noncytolytic cells that produce soluble factors able to inhibit viral replication. Although inhibition of HIV replication by soluble factors pro-duced by CD81T cells has been well documented, the identity
[image:8.612.90.510.67.578.2]of these factors remains controversial. The initial report from Cocchi et al. suggested that RANTES, MIP-1a, and MIP-1b were largely responsible for the ability of supernatants from CD81T cells to inhibit replication of CCR5-dependent HIV-1 FIG. 7. Soluble factors from CD3-activated CD81T cells from SIVmac239Dnef-infected macaques inhibit HIV-1 replication. (A) Replication of both HIV-1
Ba-L and HIV-1JR-CSFin PM1 cells is inhibited by SIVmac239Dnef-infected CD81T-cell-mediated soluble factors. PM1 cells were acutely infected with various dilutions of primary isolates HIV-1Ba-Land HIV-1JR-CSFand then incubated in the presence of either soluble factors (1:2 dilution) or RANTES (500 ng/ml). In addition, in some cases, neutralizing antisera to human RANTES, MIP-1a, and MIP-1bwere preincubated at 200, 80, and 100mg/ml, respectively, with soluble factors or RANTES and then added to the indicated cultures. (B) Soluble factors inhibited HIV-1 replication in a dose-dependent manner. In the same conditions, PM1 cells were acutely infected with primary isolates HIV-1Ba-Land HIV-1JR-CSF(dilution of 1/1,000) and then incubated in the presence of soluble factors at various concentrations. RANTES (500 ng/ml) was used as a control. (C) HIV-1NL4-3replication in CEMx174-SEAP cells is inhibited by SIVmac239Dnef-infected CD81T-cell-mediated soluble factors. CEMx-174-SEAP cells were infected with various dilutions of HIV-1NL4-3and cultured either with supernatants (1:2 dilution) from CD3-activated CD81T cells from a macaque immunized with SIVmac239Dnef or with RANTES (500 ng/ml). In addition, in some cases, neutralizing antisera to human RANTES, MIP-1a, and MIP-1bwere preincubated at 200, 80, and 100mg/ml, respectively, either alone or in combination with soluble factors or RANTES, and then were added to the indicated cultures. Similar data were obtained in two independent experiments.
on November 9, 2019 by guest
strains (10). However, subsequent reports have suggested that CD81T cells are likely to produce other soluble factors able to
inhibit HIV-1 replication (32, 43, 54). We found that both SIVmac239 and SIVmac251 were inhibited by RANTES, MIP-1a, and MIP-1b, a finding consistent with the identification of CCR5 as a coreceptor for several SIV strains, including SIVmac251 (6). In addition, we observed that production of these b-chemokines by CD3-stimulated CD81 T cells from
animals vaccinated with live attenuated SIV strains is 8- to 10-fold greater than in uninfected controls. A similar finding was reported by Lehner et al., who analyzed production of RANTES, MIP-1a, and MIP-1bby stimulated CD81T cells
from animals vaccinated with a subunit SIV vaccine and dem-onstrated greater production of b-chemokines in protected than in unprotected vaccinees (31).
However, our results suggest that theb-chemokines RANTES, MIP-1a, and MIP-1bare not the dominant soluble mediators of suppression of SIV replication by CD81T cells from
ani-mals vaccinated with live, attenuated SIV strains. First, we observed no significant blocking of the ability of supernatant from activated CD81T cells from SIV239Dnef-infected
ani-mals to inhibit SIV replication following the addition of a combination of polyclonal antibodies to RANTES, MIP-1a, and MIP-1b. Although neutralizing antibodies specific for rhesus macaque chemokines are not available, the amino acid sequences of rhesus RANTES and MIP-1a are 100% conserved with their human homologs, and MIP-1b is 95% identical (47b). Therefore, these human-specific antisera are likely to neutralize the appropriate rhesus molecules. Second, we observed a significantly greater inhibition of SIV replication by soluble factors produced from stimulated CD81T cells than
by recombinantb-chemokines. We demonstrated a 10- to 30-fold inhibition of SIV replication by soluble factors from stim-ulated CD81T cells, whereas only 2- to 3-fold inhibition was
observed with the recombinantb-chemokines RANTES, MIP-a, and MIP--b, even using concentrations as high as 500 ng/ml and combinations of these molecules. Measured levels of b-chemokines in these supernatants were generally 20 to 30 ng/ml, a level at which we observed only minimal inhibition of SIV replication in assays using recombinant chemokines. Third, soluble factors from stimulated CD81T cells were able
to induce up to 30-fold inhibition of SIV replication in the CEMx174 cell line, which does not express CCR5 (27). Finally, soluble factors from activated CD81T cells were also able to
inhibit replication of CXCR4-dependent HIV-1 strains. Thus, our data suggest that CD81T lymphocytes from
ma-caques vaccinated with live attenuated SIV strains produce factors other than RANTES, MIP-1a, and MIP-1b that are able to inhibit SIV and HIV replication and that these alter-native factors are likely to play a dominant role in mediating suppression of SIV replication. However, these findings do not exclude a potential role for theb-chemokines in suppressing SIV replication. Identification of the molecules responsible for inhibition of SIV replication may facilitate identification of ligands for the recently described additional SIV coreceptors BOB (Gpr15) (8, 12) and Bonzo (STRL33) (2, 33) and to a better understanding of the role of these coreceptors in infec-tion.
Since macaques immunized with SIV239Dnef or SIV239D3 are known to be protected against challenge with pathogenic SIVmac251 (11, 51), characterization of soluble factors able to inhibit SIV replication produced by CD81T lymphocytes from
these animals may prove relevant to defining the correlates of immune protection. Future studies will be necessary to define the role of soluble factors in protective immunity and whether this property is independent of SIV-specific CTL activity. Our
data suggest that the ability of CD81T cells to suppress SIV
replication is the result of a specific immune response rather than the result of viral interference. CD41T cells from animals
immunized with SIVmac239Dnef or SIVmac239D3 were easily infected with SIV in vitro, resulting in levels of viral replication similar to those observed in CD41T cells from uninfected
con-trols. Although we cannot exclude the possibility of viral inter-ference in a reservoir other than CD41T lymphocytes, since
CD41 T lymphocytes are the dominant reservoir for HIV-1
replication in vivo (21, 46) and the frequency of SIV-infected cells in animals infected with live attenuated SIV strains is relatively low (26, 51), this possibility appears unlikely. Inves-tigation of the ability of CD81T lymphocytes to inhibit SIV
replication in vitro, and the identification of components in-volved in the protective immunity in vivo, may lead to a better understanding of AIDS pathogenesis and to new preventative strategies.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grants RR 00168 and AI35365.
We thank Ronald C. Desrosiers for many helpful discussions and providing samples from SIV-infected animals, Bruce Walker for en-couragement and support, Ronald C. Desrosiers and Otto Yang for review of the manuscript, Otto Yang for providing the PM1 cell line and the HIV-1Ba-Land HIV-1JR-CSFisolates, Andrew Luster for pro-viding recombinant chemokines, and Michael Rosenzweig and Mary-Ann DeMaria for assistance with flow cytometry.
REFERENCES
1. Alkhatib, G., C. Combadiere, C. C. Broder, Y. Feng, P. E. Kennedy, P. M. Murphy, and E. A. Berger.1996. CC CKR5: a RANTES, MIP-1a, MIP-1b
receptor as a fusion cofactor for macrophage-tropic HIV-1. Science 272: 1955–1958.
2. Alkhatib, G., F. Liao, K. W. C. Peden, E. A. Berger, and J. M. Farber. 1997. A new SIV co-receptor, STRL33. Nature 388:238.
3. Allen, T. M., J. Sidney, M.-F. Del Guercio, R. Glickman, G. Lensmeyer, D. Wiebe, C. Pauza, R. P. Johnson, A. Sette, and D. I. Watkins. Characteriza-tion of the peptide binding motif of a rhesus MHC Class I molecule (Mamu-A*01) that binds an immunodominant CTL epitope from SIV. J. Immunol., in press.
4. Baier, M., A. Werner, N. Bannert, K. Metzner, and R. Kurth. 1995. HIV suppression by interleukin-16. Nature 378:563.
5. Brinchmann, J. E., G. Gaudernack, and F. Vartdal. 1990. CD81T cells inhibit HIV replication in naturally infected CD41T cells. Evidence for a soluble inhibitor. J. Immunol. 144:2961–2966.
6. Chen, Z., P. Zhou, D. D. Ho, N. R. Landau, and P. A. Marx. 1997. Genetically divergent strains of simian immunodeficiency virus use CCR5 as a corecep-tor for entry. J. Virol. 71:2705–2714.
7. Choe, H., M. Farzan, Y. Sun, N. Sullivan, B. Rollins, P. P. D., L. Wu, C. R. Mackay, G. LaRosa, W. Newman, N. Gerard, C. Gerard, and J. Sodroski. 1996. The beta-chemokine receptors CCR3 and CCR5 facilitate infection by primary HIV-1 isolates. Cell 85:1135–1148.
8. Clapham, P. R., and R. A. Weiss. 1997. Spoilt for choice of co-receptors. Nature 388:230–231.
9. Clerici, M., C. Balotta, and M. Galli. 1996. Soluble HIV suppressive factors: more than one Holy Grail? Immunol. Today 17:297.
10. Cocchi, F., A. L. DeVico, A. Garzino-Demo, S. K. Arya, R. C. Gallo, and P. Lusso.1995. Identification of RANTES, MIP-1a, and MIP-1bas the major HIV-suppressive factors produced by CD81T cells. Science 270:1811–1815. 11. Daniel, M. D., F. Kirchhoff, S. C. Czajak, P. K. Sehgal, and R. C. Desrosiers. 1992. Protective effects of a live attenuated SIV vaccine with a deletion in the nef gene. Science 258:1938–1941.
12. Deng, H., D. Unutmaz, V. N. KewalRamani, and D. R. Littman. 1997. Expression cloning of new receptors used by simian and human immunode-ficiency viruses. Nature 388:296–300.
13. Dittmer, U., T. Nisslein, W. Bodemer, H. Petry, U. Sauermann, H. C. Stahl, and G. Hunsmann. 1995. Cellular immune response of rhesus monkeys infected with a partially attenuated nef deletion mutant of the simian im-munodeficiency virus. Virology 212:392–397.
14. Dragic, T., V. Litwin, G. P. Allaway, S. R. Martin, Y. Huang, K. S. Na-gashima, C. Cayanan, P. J. Maddon, R. A. Koup, J. P. Moore, and W. A. Paxon.1996. HIV-1 entry into CD41cells is mediated by the chemokine receptor CC-CKR-5. Nature 381:667–673.
15. Dulbecco, R. 1988. Virology, p. 22–25. J. P. Lippincott, Philadelphia, Pa.
on November 9, 2019 by guest
http://jvi.asm.org/
HIV-1 infection. Nature 373:123–126.
22. Johnson, R. P., R. L. Glickman, J. Q. Yang, A. Kaur, J. T. Dion, M. J. Mulligan, and R. C. Desrosiers.1997. Induction of vigorous cytotoxic T-lymphocyte responses by live attenuated simian immunodeficiency virus. J. Virol. 71:7711–7718.
23. Kannagi, M., T. Masuda, T. Hattori, T. Kanoh, K. Nasu, N. Yamamoto, and S. Harada.1990. Interference with human immunodeficiency virus (HIV) replication by CD81T cells in peripheral blood leukocytes of asymptomatic HIV carriers in vitro. J. Virol. 64:3399–3406.
23a.Kaur, A., R. Glickman, and R. P. Johnson. Unpublished data.
24. Kawai, T., J. Wong, J. MacLean, A. B. Cosimi, and S. Wee. 1994. Charac-terization of a monoclonal antibody (6G12) recognizing the cynomolgus monkey CD3 antigen. Transplant. Proc. 26:1845–1846.
25. Kestler, H., T. Kodama, D. Ringler, M. Marthas, N. Pedersen, A. Lackner, D. Regier, P. Sehgal, M. Daniel, N. King, and R. Desrosiers.1990. Induction of AIDS in rhesus monkeys by molecularly cloned simian immunodeficiency virus. Science 248:1109–1112.
26. Kestler, H. W., D. J. Ringler, K. Mori, D. L. Panicali, P. D. Sehgal, M. D. Daniel, and R. C. Desrosiers.1991. Importance of the nef gene for mainte-nance of high virus loads and for development of AIDS. Cell 65:651–662. 27. Kirchhoff, F., S. Pohlmann, M. Hamacher, R. E. Means, T. Kraus, K.
Uberla, and P. D. Marzio.1997. Simian immunodeficiency virus variants with differential T-cell and macrophage tropism use CCR5 and an unidentified cofactor expressed in CEMx174 cells for efficient entry. J. Virol. 71:6509– 6516.
28. Koup, R. A., and D. D. Ho. 1990. Quantitative culture assay for HIV-1 in peripheral blood, p. 107–112. In A. Aldovini and B. D. Walker (ed.), Tech-niques in HIV research. Stockton Press, New York, N.Y.
29. Koyanagi, Y., S. Miles, R. T. Mitsuyasu, J. E. Merrill, H. V. Vinters, and I. S. Chen.1987. Dual infection of the central nervous system by AIDS viruses with distinct cellular tropisms. Science 236:819–822.
30. Landay, A. L., C. E. Mackewicz, and J. A. Levy. 1993. An activated CD81T cell phenotype correlates with anti-HIV activity and asymptomatic clinical status. Clin. Immunol. Immunopathol. 69:106–116.
31. Lehner, T., Y. Wang, M. Cranage, L. A. Bergmeier, E. Mitchell, L. Tao, G. Hall, M. Dennis, N. Cook, R. Brookes, L. Klavinskis, I. Jones, C. Doyle, and R. Ward.1996. Protective mucosal immunity elicited by targeted iliac lymph node immunization with subunit SIV envelope and core vaccine in ma-caques. Nat. Med. 2:767–775.
32. Levy, J. A., C. E. Mackewicz, and E. Barker. 1996. Controlling HIV patho-genesis: the role of noncytotoxic anti-HIV activity of CD81T cells. Immu-nol. Today 17:217–224.
33. Liao, F., G. Alkhatib, K. W. C. Peden, G. Sharma, E. A. Berger, and J. M. Farber.1997. STRL33, a novel chemokine receptor-like protein, functions as a fusion cofactor for both macrophage-tropic and T cell line-tropic HIV-1. J. Exp. Med. 185:2015–2023.
34. Lusso, P., F. Cocchi, C. Balotta, P. D. Markham, A. Louie, P. Farci, R. Pal, R. C. Gallo, and M. S. Reitz. 1995. Growth of macrophage-tropic and primary human immunodeficiency virus type 1 (HIV-1) isolates in a unique
42. Miller, M. D., C. I. Lord, V. Stallard, G. P. Mazzara, and N. L. Letvin. 1990. The gag-specific cytotoxic T lymphocytes in rhesus monkeys infected with the simian immunodeficiency virus of macaques. J. Immunol. 144:122–128. 43. Pal, R., A. Garzino-Demo, P. D. Markham, J. Burns, M. Brown, R. C. Gallo,
and A. L. DeVico.1997. Inhibition of HIV-1 infection by the beta-chemokine MDC. Science 278:695–698.
44. Paliard, X., A. Y. Lee, and C. M. Walker. 1996. RANTES, MIP-1aand MIP-1bare not involved in the inhibition of HIV-1SF33 replication mediated by CD81T-cell clones. AIDS 10:1317–1321.
44a.Reed, L. J., and H. Muench. 1938. A simple method of estimating fifty per cent endpoints. Am. J. Hyg. 27:493–497.
45. Scala, E., G. D’Offizi, R. Rosso, O. Turriziana, R. Ferrara, A. M. Mazzone, G. Antonelli, F. Aiuti, and R. Paganelli.1997. C-C chemokines, IL-16, and soluble antiviral factor activity are increased in cloned T cells from subjects with long-term nonprogressive HIV infection. J. Immunol. 158:4485–4492. 46. Schnittman, S. M., M. C. Psallidopoulos, H. C. Lane, T. Thompson, M.
Baseler, F. Massari, C. H. Fox, N. P. Salzman, and A. S. Fauci.1989. The reservoir for HIV-1 in human peripheral blood is a T cell that maintains expression of CD4. Science 245:305–308.
47. Tsubota, H., C. I. Lord, D. I. Watkins, C. Morimoto, and N. L. Letvin. 1989. A cytotoxic T lymphocyte inhibits AIDS virus replication in peripheral blood lymphocytes. J. Exp. Med. 169:1421–1434.
47a.U.S. Department of Health and Human Services. 1985. Guide for the care and use of laboratory animals. DHHS publication no. (NIH) 85-23. U.S. Government Printing Office, Washington, D.C.
47b.Villinger, F. Personal communication.
48. Walker, C. M., A. L. Erickson, F. C. Hsueh, and J. A. Levy. 1991. Inhibition of human immunodeficiency virus replication in acutely infected CD41cells by CD81cells involves a noncytolytic mechanism. J. Virol. 65:5921–5927. 49. Walker, C. M., D. J. Moody, D. P. Stites, and J. A. Levy. 1986. CD81
lymphocytes can control HIV replication in vitro by suppressing virus repli-cation. Science 234:1563–1566.
50. Walker, M. C., and J. A. Levy. 1989. A diffusible lymphokine produced by CD81T lymphocytes suppresses HIV replication. Immunology 66:62–630. 51. Wyand, M. S., K. H. Manson, M. Garcia-Moll, D. Montefiori, and R. C. Desrosiers.1996. Vaccine protection by a triple deletion mutant of simian immunodeficiency virus. J. Virol. 70:3724–3733.
52. Xu, X.-N., G. R. Screaton, F. M. Gotch, T. Dong, R. Tan, N. Almond, B. Walker, R. Stebbings, K. Kent, S. Nagata, J. E. Stott, and A. J. McMichael. 1997. Evasion of CTL responses by nef-dependent induction of fas ligand (CD95L) expression on simian immunodeficiency virus-infected cells. J. Exp. Med. 186:7–16.
53. Yang, O. O., S. A. Kalams, A. Trocha, H. Cao, A. Luster, R. P. Johnson, and B. D. Walker.1997. Suppression of human immunodeficiency virus type 1 replication by CD81cells: evidence for HLA class I-restricted triggering of cytolytic and noncytolytic mechanisms. J. Virol. 71:3120–3128.
54. Yang, O. O., and B. D. Walker. 1997. CD81cells in human immunodefi-ciency virus type 1 pathogenesis: cytolytic and noncytolytic inhibition of viral replication. Adv. Immunol. 66:273–311.