JOURNALOFVIROLOGY, June 1979,p.805-820
0022-538X/79/06-0805/16$02.00/0 Vol.30, No.
3
Isolation
and
Localization
of
Herpes Simplex
Virus
Type
1
mRNA
KEVIN P. ANDERSON, JAMES R. STRINGER,t LOUIS E. HOLLAND,AND EDWARDK. WAGNER*
Departmentof Molecular Biology and Biochemistry, UniversityofCaliforniaatIrvine, Irvine,
California
92717Received for publication4December 1978
Herpes simplex virus (HSV) DNA boundtocellulose has been usedas areagent
to isolate viral mRNA for size analysison denaturingagarose gels. Totalviral polysomal polyadenylated RNAwasisolated from cellslate afterinfectionwhen
such RNA hassequencesencodedbyapproximately 45%of theHSV DNA.This RNA hasasizerangeof from 1.5to -8kilobases, withcertainsizes, suchas 1.7
to 1.9 kilobases, being favored. We have used the restriction endonucleases HindIIIandXbaIsinglyandtogethertogeneratevarious sizedfragmentscovering
the entire HSV-1genome.Thesefragmentshave been bound to cellulose toallow isolation of HSV-1 mRNA annealing to different regions of the viral genome.
Discrete sizes of viral mRNAareassociated with certainregionsof thegenome,
but the mRNApopulationhybridizingtoeventhesmallest restrictionfragments is complex. We used hybridization of size-fractionatedRNA toSouthernblotsof restriction fragments of HSV-1 DNAgeneratedbytheBglIIaswellasHindIII
andXbaI endonucleases to confirm the preparative hybridization data and to provide some overlap data for positioning transcripts. The data of blot and
preparative hybridization agreed very well and were combined to construct a
preliminarytranscriptionmapof HSV-1. Suchamaprevealed at least twoareas
ofthelong unique regionofthe HSV-1genomewhich annealed toalargenumber
of HSV-1 transcripts. Furthermore, discrete-sized mRNA specieslargerthan 5 kilobases inlengthwerefoundonlyin themiddle of thelong unique region.The
implications of thesedataarediscussed.
Herpessimplex virustype 1(HSV-1)isalarge DNA virus which replicates in the nucleus of infected cells. At least 48 HSV-1-specific pro-teins, atleast half of which arestructural
pro-teins, have been identified in infected cellsby using sodium dodecyl sulfate-polyacrylamide gels (11). The virus particle contains double-stranded DNA having a guanine-plus-cytosine
content of 67% (23). Themolecular weights of various strains of HSV-1 DNA have been
re-portedtobe between95x
106
and100x 106(8, 32), correspondingtoanapproximate length of 140,000 to150,000basepairs (140to 150kilobase [kb] pairs). The particular strain of HSV-1 DNA used in this study(KOS) is28.1times thelengthof the relaxed replicative form of
4X174
(34), correspondingto alengthof 150 kbpairs based on the known length of this phage DNA (3). HSV DNAisstructurallycomplex and has been shown to consist of a long region and a short region, each composed of unique sequences bounded by different inversely repeated se-quences(24). Each of thesetworegionsmay betPresent address: Cold Spring HarborLaboratory, Cold Spring Harbor, NY 11724.
linked to the otherby either end, resulting in fourequimolarconfigurations of the viral DNA (9, 39).
Transcription of thisgenomehasbeenstudied in our laboratory and in several others, with
general agreement on the following points. In cells infected at moderate multiplicity, HSV DNA synthesis begins about3 hpostinfection (p.i.) and reaches a maximum rate by 6 h p.i. Before theonsetofviral DNA synthesis, abun-dantRNAtranscripts from20 to25% ofthe viral genome arefound associated with polyribosomes (15, 28, 31). Both"Southern blot" hybridization and RNA displacement loop hybridization of HSV RNA have shown that these early HSV mRNAspeciesmap throughout the HSV-1 ge-nome (7, 29). The abundant, early HSV-1 mRNA species can be differentiated into two classes. One class consists of the
immediate-early or a mRNA's, which are found in high abundanceinthecytoplasmofcells treated with cycloheximide fromthe time ofinfection (15). TheamRNAspecies have been shown,by both blothybridizationandsolution hybridizationto isolated restrictionfragmentsof HSV-1DNA, to betranscribed from defined, noncontiguous
re-805
on November 10, 2019 by guest
http://jvi.asm.org/
806
gions of DNA comprising 10 to 12% of the viral genome(7, 14). The other major abundant class of mRNA species found prior to viral DNA replication is the /3 messages. These mRNA's encodeproteinswhich aresynthesized onlyafter proteinsencodedbythe a mRNAspecieshave beensynthesized (12).
Attimesafter infection when viral DNA syn-thesis is maximal (late), the majority ofearly RNAsequences continuestobesynthesized (7, 33),andtranscriptsfromanadditional 15to20% of the viralgenome are also foundon polyribo-somes in good yield. These are the y, orlate, mRNA species. At least a portion of these -y mRNA species has been detected even at the earliest timesafterinfection,butat a concentra-tion10- to 20-fold lower than thea
and,8
mRNA classes (15, 28, 30). At this time after infection (late), as much as 90% of newly synthesized polyadenylated [poly(A)] RNA onpolyribo-somesin the infected cellis viral(28). Further-more,it has beenextensivelyshown that all the sequences in viral mRNA can be isolated as poly(A) RNA (25, 28).
mRNA molecules of HSV-1 associated with the polyribosomes of infected cells resemble mRNA molecules of other DNA viruses and eucaryotic cells. HSV-1 mRNA, in addition to beingpolyadenylated,iscappedatthe 5' end(4,
20),andwehavefound itsaveragesizetobeno less than2kb(28).However,due inparttothe large size and complexity of the HSV genome, little has beenreportedconcerningthe proper-ties and origins of individual viral mRNA spe-cies.
In thisstudy, wehave usedtwoindependent
methodstodetermine the sizerangesofspecific HSV-1mRNAspeciessynthesizedlate after in-fection and tolocalize individual transcriptsto
specific regions of the viral genome. We have used restriction fragments of HSV-1 DNA boundtocelluloseto
purify
viral mRNA homol-ogous to these restrictionfragments. Size frac-tionation on denaturing agarose gels of viral mRNA obtained in thismannerrevealsa char-acteristicpattem ofspecific-sizemRNAspecies homologoustoeach restrictionfragment.In ad-ditiontosuch characteristicspecies,allregions of the viralgenomehybridizeto acomplex pat-tern of minorspecies of viralmRNA. The size distributions of viralmRNAencodedby differ-entregions of the HSV-1genomealso have been determined by first fractionating infected-cell mRNA on denaturing agarose gels and then hybridizing size fractions to Southem blots of HSV-1restrictionfragments.Themajor species and less prominent discrete species of viral mRNAseeninRNApreparatively hybridizedtorestrictionfragmentsarereadilyidentifiableby
using this procedure. The results of both types of experiments have been used to construct a preliminarytranscription map of the HSV-1 ge-nome.This mapshows theminimumnumber of discrete mRNA species consistent withour data.
MATERIALS AND METHODS Celisand virus.Monolayercultures of HeLa cells
weregrown at37°C inEagle minimal essential
me-dium containing 10% calf serum and no antibiotics.
Plaque-purifiedvirusof the KOS strain of HSV-1 was
used forall infections.
Preparationof RNA. Monolayercell cultures (2
x 107cells per T-150flask) were infected for 30min
with 10PFU of virus per cell and overlaid with
modi-fied medium 199 containingEarlesalts (Flow
Labo-ratories)supplemented with 5% fetalcalf serum. For
3H-labeledRNA, thecellswerelabeled from 5.25 to 6
h p.i. with 15 MCi of [3H]uridine (28 Ci/mmol;
Schwarz/Mann)perml inmedium 199 containing 5%
dialyzed fetal calf serum. For 3P-labeled RNA, the
cellswerelabeled from2 to 6 hp.i. with 200,uCi of
carrier-free [32P]orthophosphate (New England
Nu-clear)perml inEagle minimalessential medium
con-taining 1/10 thenormalphosphate concentration and
5%dialyzedfetal calf serum.
Polyribosome-associated RNA was isolated by the
magnesium precipitationmethod ofPalmiter (22) as
describedbyStringeretal. (28). After phenol
extrac-tion and ethanolprecipitation,poly(A) RNA was
pu-rified from the total polyribosome-associated RNA,
usingoligodeoxythymidylic acid-cellulose
(Collabora-tiveResearch). Poly(A) RNA was bound to the
cellu-lose in 0.5 MNaCl-0.1MTris (pH7.6)-imM EDTA,
rinsedthoroughly,and eluted in asolution containing
0.01M Tris(pH 7.6) and1mMEDTA. This procedure
results in 50% of the3Hradioactivity and 15% of the
32P radioactivity in polyribosome-associated RNA
beingrecovered aspoly(A)RNA. Asingle T-150 flask
typically yielded4 to 5ygofpoly(A) RNA with specific activities of 120,000 and 200,000 cpm/,ug, respectively,
when 3H labeled and32plabeled. Radioactivity was
measuredasa1%solution inAquasolII scintillation
fluid (NewEngland Nuclear) in a Beckman LS-230
scintillationcounter.
Thispoly(A)RNA has been shown to be intact by the following criteria. Recovery of 5' cap structure from poly(A) RNA isolated in this manner is virtually
quantitative(20),demonstratingnoappreciable
break-age between the 3'poly(A) tail and the 5' cap.
Fur-thermore, this poly(A) RNA serves as an extremely
efficient template for in vitro translation, and the
polypeptide productsmigrate in sodium dodecyl
sul-fate-polyacrylamide gelsassharp bands corresponding
toproteins synthesized in vivo, withlittleor no
back-ground of partial translation products as would be
seenwithnickedordegradedtemplate RNA
(Ander-son etal.,unpublished data).
PreparationofHSV DNA.Cellsgrown inplastic
rollerbottles (108 cellsperbottle) were infected with
10PFU of virus percelland incubated in medium 199
containing5% fetal calfserum.For32P-labeledDNA, thecellswereoverlaid at4hp.i. with phosphate-free Eagle minimal essential medium containing 5%
di-alyzedfetal calfserumand25to 50,uCi of
on November 10, 2019 by guest
http://jvi.asm.org/
HSV mRNA 807 phosphate per ml. The cells were scraped into the
mediumafter24h,collected by low-speed
centrifuga-tion (2,000xgfor 5 min), and resuspended in 0.01 M
NaCl-0.01 M Tris (pH 7.4)-3 mM MgCl2-0.5%
Noni-det P-40. Thecells were lysed by 6 to 10 strokes in a
tight-fitting Dounce homogenizer, and nuclei were
re-moved by low-speedcentrifugation. Virus waspelleted
from thecytoplasmic supernatant by centrifugation at
27,000xgfor 20 min. The viruspellet was solubilized
in 0.1 M NaCl-0.02M EDTA-0.01 MHEPES
(N-2-hydroxyethyl piperazine-N'-2-ethanesulfonic acid) (pH7.8)-1%Sarkosyl and digested with proteinase K (250,ug/ml)for3h at 55°C. The DNA was banded by
CsCl-isopycnic centrifugationaspreviouslydescribed
byWagner et al. (35), and DNA at 1.725 g/ml (>90%
of DNA ingradient)wascollected and dialyzed against
anappropriate restriction endonuclease buffer. A total
of 25 to50pgofpurified HSV DNA is recovered from
108 cells by this procedure.
Restriction endonuclease digestion of HSV-1
DNA and fractionation of resulting fragments.
Restriction endonucleases HindIII, XbaI, andBglII
werepurchased from New England Biolabs.All
diges-tionswereperformedunder conditions recommended
by thesupplier. Enzyme, at 1 U/4 pugof DNA, was
incubated with DNA for 2 h at 37°C. Double digestion
with HindIII and XbaI enzymes was performed by
digestingfirstwith HindIII and then adjusting the salt
concentration and addingB-mercaptoethanol before
digestingwithXbaIenzyme.
Restricted DNA (30 to 100pg)wasfractionated by
electrophoresisintwo 7.5-cmtracks in 0.5%agarose,
horizontal slab gels (42 by 20 by 1.2 cm) for 48hat 60
V (80 mA). Electrophoresis buffer was 0.04 M Tris
(pH7.8)-0.005 M sodium acetate-2 mM EDTA-0.5%
ethidium bromide. Bands were visualized by using
long-wave UV light.
DNAwasisolated from excised bands by dissolving
the agarose in 5 M NaClO4 at 60°C and absorbing
DNA to hydroxyapatite as described by Wilkie and
Cortini (39).After elution in 0.5 Msodium phosphate
(pH6.8), the DNAwasextracted withisopentylalcohol
toremoveanyremaining ethidium bromide, desalted on aSephadexG-50 column, and ethanol precipitated.
Binding of DNA to diazotized cellulose. Total
HSVDNA,aswell as restriction fragment DNA, was
boundtodiazotizedcelluloseby amodification of the
procedureofNoyes and Stark (21).
m-Aminobenzylox-ymethyl cellulose(Miles Laboratories) was
reprecipi-tated and diazotizedasdescribed in reference21.
How-ever,for rinses andcoupling, 10 mM potassium
phos-phate (pH 6.5) was used as a buffer instead of 0.2M
sodiumborate (pH 8.0). DNA, in 80% dimethyl
sulf-oxide and2mMpotassium phosphate at
concentra-tions of 100to200,ug/milfor total HSV DNA and 25
to 50
jg/ml
forrestriction fragment DNA, was heatedto780Cfor 5 min, cooled to 4°C, and then mixed with
thediazotizedcellulose so that thecellulose
concen-trationwas10 mg(dryweight)/ml.When lessthan 10
pgofDNA wasbeing manipulated, Escherichia coli
DNA wasadded as carrier tobring theconcentration
to 10pg.Thecellulosesuspension was mixed
contin-uouslyat40Cfor 48hand thenrinsed twice with 80%
dimethyl sulfoxide and 2 mM potassium phosphate at
50°C, four times with 0.lx SSC (0.15 M NaCl plus
0.015Msodium citrate) at250C,three times with 98%
formamide and 0.01 M HEPES (pH 8.0), and three times with hybridization buffer (80% formamide, 0.4
MNa+, 0.1 M HEPES [pH8.0],0.005MEDTA).
32P-labeled DNA was included in each reaction so that the efficiency of coupling could be monitored. A total of 35 to45%ofinput radioactivity remained bound to the
celluloseafter rinsing.DNAcellulosecontained 5 to 10
pgof DNA per mg ofcellulosefor total DNA and 1 to
3 pug/mg of cellulose for restriction fragment DNA.
DNAcoupled tocellulosewasstored inhybridization
buffer under nitrogen at40C.
Hybridization of RNA to cellulose-bound
DNA. 3H-labeled, poly(A) RNA was hybridized to
cellulose-boundDNAfor4hat57°Cinhybridization
buffer containingrecrystallized formamide. For total
HSV DNA boundtocellulose, 1to2pgof RNA (1 x
105to 2 x 105cpm)washybridizedwith5 to 10pgof
cellulose-bound DNA. For restriction fragments
boundtocellulose,10 to20pugof RNAwashybridized
to15pg-equivalentsof DNAboundtocellulose(1
pg-equivalentis theamountofrestrictionfragment
cor-respondingtodigestionof1pigof intact HSVDNA).
Thehybridizationmixturewasagitatedeveryhourto
avoid settlingof thecellulose.Afterhybridization,the
cellulosewasrinsedfour times with2xSSCat250C,
twice withhybridizationbufferatroomtemperature,
and five times withhybridizationbufferat600C.
Hy-bridizedRNA waseluted in 98% formamide and 10 mMHEPES (pH8.0)at600C.Theeluantwasdiluted
to a final formamide concentration of 22% and
ad-justedto0.1 Msodium acetate, and the eluted RNA wasethanolprecipitated.
Methylmercuryagarosegels.RNAwas fraction-atedbyelectrophoresison1.4% agarosegels(15by0.6 cm)containing10mMmethylmercuryhydroxide
(Al-pha) as described by Bailey and Davidson (2). Gels
wererunfor4 to 6hat3mA/gel (2.5 V/cm). After
electrophoresis, the gels were soaked in 50 mM
,B-mercaptoethanolfor 20 min andslicedat2- or3-mm
intervals.The3H radioactivityineach slicewas meas-uredby dissolvingslices in0.3ml of7%perchloricacid
at 72°C andmixingwith4 ml ofAquasol. 32P
radio-activity of each slice was determined by Cerenkov
radiationcountingof intact slices.
Preparationandhybridizationof DNA
South-ernblots. Slabgelsof restrictionendonucleasedigests
were blotted onto nitrocellulose paper (BA 85,
Schleicher &Schuell) inlOx SSC, usingthe method
ofSouthern (27).TheDNA-containingblotswerecut into4-mm stripsfor hybridization to the RNA
con-tained inindividual slices ofmethylmercury agarose
gels. Agarosegels containing10to15
pg
of32P-labeledpoly(A)RNAweresoaked in15mM
,B-mercaptoeth-anolfor 30 min andcutinto 2-or3-mmslices. Slices
containing32P-labeledRNAweredissolvedby heating
to
720C
in asolution containing 0.4 M Na+, 0.1 MHEPES(pH 7.8)-0.005MEDTA,and 65%formamide.
Thedissolved32P-labeled RNA thenwashybridized
for24to36 hat570Cinsealedplasticbagscontaining
individualnitrocellulose strips.Stripsthenwererinsed
forthree 2-hperiodsat
480C
inhybridizationbufferand 65%formamide, arranged intheir originalorder
from the blot, wrapped in Saran wrap, and pressed
next toKodakX-omat-RX-rayfilm for24to72h.
VOL. 30,1979
on November 10, 2019 by guest
http://jvi.asm.org/
RESULTS
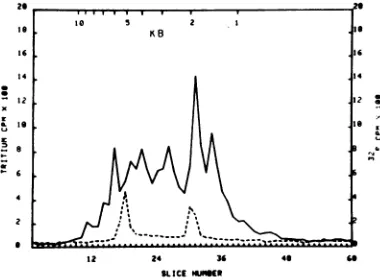
Sizerangeof HSV-1mRNA.P poly(A) RNA, labeled in HSV-1-ir
cellswith a45-min [3H]uridine pL afterinfection, washybridized to boundtocelluloseasdescribed in I Methods. The hybridized RNA(15 input radioactivity) was eluted fr(
lose and subjectedto electrophor turing agarose gels containing m hydroxide. Such gels exhibitalinea between the log size of RNA and migrated (2). The size, in kb, ofH species was determined by compa tancemigrated by HSV-1 mRNA,,c distance migrated by 3P-labeled
species, which were added to tI
internal size markers. Figure 1 sh tinct size species of HSV-1 mRNA >8 to -1 kbarereadily distinguis isaratherlarge proportion of vira size range of 1.9 to 1.7 kb, a de mRNAthat is2.1to 2.7 kb in size significantamountofviralmRNA kb. Inalarge number ofpreparati tions, the sizerangeand discretesj
20
to 18 16 1 4
x
a 1@
.8
--6
2
12 24 16
SLICECtNE*6
FIG. 1. Size distributionofHSV-1 beledpolysomal poly(A) mRNAfrom
infection was hybridized to 15pg o)
bound to cellulose, and the HSV-sl
eluted (see text). This viralmRNA w
32P-labeled HeLa cell rRNA andfracti
trophoresis on a 0.6- by 15-cm gel
agarosecontaining10 mMmethylmer(
(2). Electrophoresiswasfor4 hat 3n
The gel was fractionated in 2-mm s
radioactivitywasdetermined. The siz
wastakentobe0.67±0.07x106daltoi
2kb)andthatof28SrRNA,1.76±0.1L
(5.2 kb) (17, 37).Sizeextrapolationswei
factthat RNAmigrationisalogarith,
RNA sizeasshown byBaileyandDa discussed in Holmetal.(10).
RNA detected were invariant, but there was
'olyribosomal
somevariability
in theamountofradioactivity
nfected HeLa recovered in the size ranges shown. Theamount
alse
at 5.25 h of viral RNA between 10 and 2.5 kb varies HSV-1 DNA between 55 and 65% of the total viral RNAMaterials
and recovered. Such variation isnotcorrelated withto20% of the the
specific preparation
of DNAcellulose
used om the cellu- in a givenexperiment.esis on dena- To establish that the viral RNA purifiedby
ethylmercury
hybridization
tocellulose-boundHSV-1 DNA is rrelationship an accuraterepresentation
of the actualpopu-Lthe distance lation of HSV-1 RNA labeled
during
thepulse
[SV-1 mRNA
period,
we tested ourhybridization
procedure
iring
the dis- forspecificity
and foritsability
to leave RNAspeciesto the
undegraded.
Twoexperiments
established the HeLa rRNAspecificity
ofcellulose-bound
HSV-1 DNA for ie sample as viral sequences. When[3H]uridine-labeled,
un-ows
that dis- infectedHeLa cellpoly(A) polyribosomal
RNA ranging from was incubated withcellulose-bound
HSV-15hable. There
DNA,
only
0.05to0.10% of theradioactive HeLa 1 RNA in the RNAwasboundtothecellulose
afterourstan-barth
of viral dardrinsing procedure.
Equivalent
amounts
of , and quite apoly(A) polyribosomal
RNA frominfectedcells larger than5 yielded from 15 to 20%hybrids.
Analysis
ofive
hybridiza-nonspecifically
bound uninfected cell RNA onpeciesof viral
methylmercury
agarose gels showed nosignifi-cant peaks of radioactivity migrating in any
given sizerange.
In asecondexperimenttotestthe specificity of hybridization to cellulose-bound DNA,
poly(A)polyribosomal RNA from infected cells
.14 washybridizedtocellulose-boundE. coliDNA.
12 .Again, less than 0.1% of inputamounts of ma-terial bound to the cellulose. That which did bind showednopeaks of radioactivityon meth-ylmercury gels.
Two control experiments demonstrated that there is no appreciable degradation of HSV-1 mRNAduringthe hybridization and elution pro-cedure. First, 32P-labeled HeLa cellrRNA was
40 60 incubated under hybridization conditions and
thenrun ondenaturing gel electrophoresis.No
RA3H_
I evidence of degradation of RNA couldbe seen mRNA.lat
by either an alteration in the ratio ofthe 28Scells lateafter
f HSV-l DNA rRNA to 18S rRNA or the appearance of in-pecific- mRNA creased RNA
migrating heterogeneously.
Intheas mixed with second control experiment, 3H-labeled poly(A)
tonated by elec- polyribosomalRNAwasisolated from infected made of 1.4% cells and directly sized on a methylmercury aga-cury hydroxide rose gel. The gel wassliced,and each slice was
nA (2.5 V/cm)- assayedfor viral RNAby hybridizationto
HSV-1lices,
andthe
1DNA ina mannersimilarto that described byes
of 000rRNas
Weiss etal.(36).Detailed resultsof thisexperi-n~s(2,0(X bases,
5x106daltons ment
will
bepresented
elsewhere(Holland
etre based onthe al., submitted for publication). It suffices here to micfunction of
report
that the distribution ofHSV-specific
ividson (2)and RNA detected by hybridization acrossthe gel
was essentially identical to that of HSV RNA
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.505.64.254.361.501.2]HSV mRNA 809
isolated by hybridization to cellulose-bound HSV-1 DNA.
We concludefrom these controlexperiments
that preparative hybridization of viral mRNA with HSV DNA bound to cellulose under our conditions gives an accurate and reproducible
representation of thepopulationofviral mRNA species present in the isolated RNA
population.
Ashasbeen discussed inMaterials and
Methods,
our evidence that the RNA is not
degraded
during our isolation is excellent; therefore, we conclude that the DNA-cellulosehybridization
gives us a valid representation of viral RNA
labeled during the pulse period. This is
con-firmed byourfindings(Andersonetal.,
unpub-lished data) that DNA-cellulose-hybridized
RNA isanefficienttemplatefor in vitro trans-lation, andthe proteinproductsshow no indi-cation of mRNA
degradation.
Identification of HSV-1 mRNA
species
encoded by differentregions
of the viral genome. TheHindIII, XbaI,
BglIl,
andHindIII/XbaIdouble-digestrestriction endonu-clease sites in the
prototypical
arrangement of the DNA of our strain(KOS)
of HSV-1 are shown inFig.2.Theseenzymes cleave the viral DNA molecule into a number offragments
LR Lu
Bgl J K IOIPINIMI I
which migrate as individual bands on agarose
gels. Gel bands were shown to contain
single
DNAfragments by blotting thegelsonto
nitro-cellulose (27) and hybridizing with 32P-labeled overlapping fragments from different restriction
endonuclease digests. Sites generated by the otherthree arrangements of the viralDNA are described in thelegendtoFig.2.
The restriction fragments of HSV-1 DNA, shown in Table 1, were isolated from agarose
gelsand boundtocellulose(Materialsand Meth-ods). Individual cellulose-bound HSV-1 DNA restrictionfragments then were used toisolate viral RNA molecules containing sequences ho-mologous to that DNAfragment. We have used
15
,ug-equivalents
of restrictionfragmentboundtocelluloseas astandardamountof DNA inour
hybridizations. We define the amount of any givenrestriction fragment derived from1,ug of total HSV-1 DNA as 1 ,ug-equivalent of that fragment.Inthismanner, the concentrations of DNA sequences in the cellulose-bound restric-tionfragmentswerekeptinthesameproportion
aswouldoccurinahybridization using15,ug of totalHSV-1 DNA boundtocellulose.
Three controls accompanied each
hybridiza-tiontocellulose-bound HSV-1 DNA restriction
LR SR SU SR
D | G | F H IL
Hind H 10 J A I K L I D
Xba G I C F E
H/X XG II 10 J 1.I XF AE I K I L 11
AC
D (SHORT) D 'M
INI
GA
A
A80 90 100
GENOME LENGTH (%)
FIG. 2. Map of restriction endonucleasefragments of the KOSstrain of HSV-1. Thesemapsarebasedon
publishedmapsforthese sites(18,26,38). We have confirmedthemforourstrain (KOS) ofHSV-1asdescribed
previously (29). The fragmentsgenerated bycleavage by the restriction endonucleasesBglII,HindIII,XbaI,
andHindIII/XbaI doubledigestsareshown fortheprototypicalarrangement(P) ofourHSV-1 strain. The
solidboxesatthe endsof the longsegment representthe longrepeat(Lit)sequences;theopen boxesonthe
short segment represent the shortrepeat(Sit)sequences.Three otherarrangementsoccur(9,39). TheseareII
where thelongsegmentisinvertedrelativetothe short; Is,where the shortsegmentisinvertedrelativetothe
long; and ILs, where both the long and short segments are inverted relative to the prototype. These
arrangementshavenoeffectonthe yield offragmentswhicharetotally contained by cleavagesites in either the longorshortsegment, but do leadtosubmolar bands for fragments coming fromthe ends of molecules and thosebridgingthejunction between the long and shortsegments.Submolarfragments for the
endonucle-asesshown are asfollows.Half-molar fragments arise from ends of molecules: BglIIL andHindIIIGare
from P and IL. BglIIJ,HindIIIH, XbaI A (XbaI D+short region),andXbaI Garefrom P and Is. BglII F,
HindIII D,XbaI B(XbaI G+short region), and XbaI Darefrom IL and Its. BglII HandHindIIIMarefrom
Is andILs. Quarter-molarfragments arise asthe sumof the half-molar fragmentsfrom either sideof the
junction foreacharrangement:BglII A is F+H,andHindIIIC is D+M. These regionsarecontiguous in
P. BglIIB is J+H, andHindIII FisH+M. These regionsarecontiguous in IL. BglII C isF+ L, and
HindIII B is D+G. These regions arecontiguousin Is. BglII E is J+ L, andHindIIIE is H+ G. These regions arecontiguous in I-s. XbaIhas no cleavagesites in the short segment; therefore, the half-molar
fragmentsXbaI Aand XbaI B have botharrangementsof the shortregion.
MINI G
VOL. 30,1979
AO
A
A
A
A
A
10
.0\
/.o
40 50 60 70on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.108.393.359.482.2]810 ANDERSON ET AL.
TABLE 1. HSV-1 endonucleasefragments isolated
for DNA-cellulosebinding
Fragmenta Mol wt
Size'
Molar- Region of ge-(source) (x1lo6) pairs)(kb ity nomesented'repre-(X)G 6.9 10.5 0.5e Longrepeat
(H)IO 9.5 14.4 1
(H)J 7.5 11.4 1
(H/X)AC 3 4.5 1
(X)F 15.5 23.5 1 Long unique
(H/X)AE 7.3 11.1 1
(H)K 6 9.1 1
(H)L 5.4 8.2 1
Long unique (H)C 22.2 33.6 0.25 jLongrepeat
Shortrepeat
(H)M 4.7 7.1 0.5e Shortrepeat
(H)N 3.1 4.7 1 Shortunique
(H)G} 8.9 13.5 0.5e ShortShortuniquerepeat
H(F) } 12.8 19.4 0.25e Long repeatShortrepeat
a (X) isderived froman
XbaI
digest; (H) is fromanHindIII digest; (H/X) is fromaHindIII/XbaIdouble
digest.
bBasedon1 kbpair havingamolecularweight of 6.6x 105.
' BasedonthefourfoldarrangementoftheHSV-1
genomeasdescribedinthelegendtoFig.2.
dSee
Fig.2.'Submolar fragments are derived from the four
,ossiblearrangementsof theHSV- 1genomedescribed
inthelegendto Fig.2.Thus, X(G) is fromPandIs,
H(C) is from P,H(M)isfromIs andus,H(G) isfrom
PandIL, and H(F)isfromIL.
fragments. (i) Asmallamount of the RNA
sam-pleto behybridizedwasfractionatedinparallel with the preparatively hybridized RNA on de-naturingagarosegels; (ii) asample of the RNA
to be hybridized to cellulose-bound restriction fragment DNA was hybridized to total HSV-1 DNA bound to cellulose and the hybrids were fractionated in parallel with the hybridstothe restriction fragment; (iii) 32P-labeled HeLa rRNA wasaddedto the hybridization mixture, and radioactivity which did not bind to the cellulose also was fractionated in parallel with thehybridized RNA at the end of the hybridi-zation period. The quality of each sample of RNA obtained by each preparative hybridiza-tion was evaluated on the basis of the control
experiments. The RNA wasdeemed intact and representativeifnodegradationof the unhybrid-ized RNA and 32P-labeled rRNA was apparent and ifthe RNApurifiedbyhybridizationtototal cellulose-boundHSV-1 DNA lookedtypical(see Fig. 1). All DNA fragmentsshow hybridization efficienciesroughlycomparabletofragmentsize (Table2).
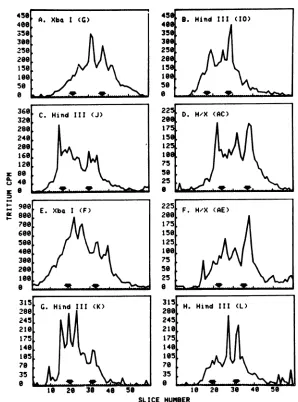
The size distributions of RNA purified by
hybridizationtodifferentHSV-1 restriction frag-ments are shown in Fig. 3. The panels are ar-ranged to represent regions of the HSV-1 ge-nomestarting withthe leftend of the L region through the S region of the P arrangement. Fromthisfigure, it is evident that certain viral mRNA species are associated with certain re-striction fragments. However, in all cases, the mRNA population homologous to a given re-striction fragment is complex. Some RNA spe-cies migrate as major sharp bands, suggesting very defined sizes for these mRNA species, which are synthesized inhigh, or at least mod-erate, abundance. RNA migrating sharply at a size of 6 kb hybridizes to HindIII fragment J and HindIII/XbaI double-digest fragment AC (Fig. 3C and D). A discrete 9.6-kb species is foundinRNAhybridizedtoHindIII/XbaI dou-ble-digest fragmentAE (Fig. 3F). HindIII frag-mentKhybridizesefficientlywith threediscrete RNA species, 7.2, 5.2, and 3.8 kb in size (Fig. 3G). Sharply migrating RNA species 3 and 1.9 kb insizehybridize toHindIIIfragmentL (Fig. 3H). Other RNA bands migrate broadly, sug-gesting a heterogeneity of sizes. A relatively large amount of such material is seen in RNA that hybridizes to XbaI fragment F (Fig. 3E). The RNA species that hybridizes to HindIII fragmentG (Fig. 3K) also isillustrative of such sizeheterogeneity.Note inparticularthe
broad-nessof thepeakcenteredat 1.9kb,which covers
arangeofsizesfrom 1.6to2.2kb.Otherdiscrete andheterogeneoussizeclassesof RNA homolo-gous to HSV-1 restriction fragments are indi-catedinTable 2.
Certain trends can be discerned from exami-nation of Fig. 3and Table2.Forexample,RNA ranging in size from 1.6 to 2 kb is a major component of RNA hybridizing to total HSV DNA and hybridizes to all of the restriction fragments shown. Such RNA is a major
com-ponent ofthe RNAhybridizing toall but three of these restriction fragments (HindIII
frag-mentsJ and K and HindIII/XbaI double-digest
fragment AE). Conversely, only one fragment (HindIII fragment IO) has sequences homolo-gous to a well-resolved RNA species (2.6 kb) between2.1 and2.7kb. Discrete peaks of RNA
>5.5 kb are found to hybridize to only four
fragments (HindIII fragments J and K and
HindIII/XbaI double-digest fragments AE and AC). It is clear, however, that heterodisperse RNA, upto about8kb in size, can be detected in RNAhybridizing to most fragments.
Some of the restriction fragments examined have commonDNAsequences. This is a conse-quence of the rearrangement of the long and shortsegmentsgenerating the four isomeric con-figurations of the genome and the presence of
on November 10, 2019 by guest
http://jvi.asm.org/
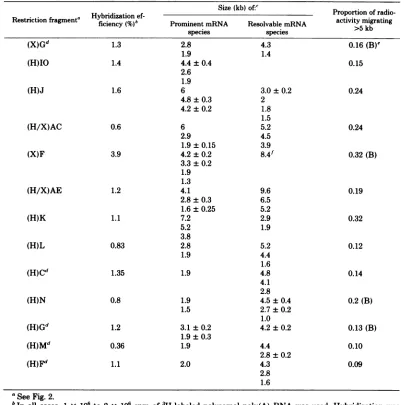
TABLE 2. HSV mRNA species hybridizing to restriction fragment DNA-cellulose
Size(kb)of' Proportionof
radio-Restrictionfragment" ficiency (%)b Prominent mRNA Resolvable mRNA activitymigrating
species species >5kb
(X)Gd
(H)IO
(H)J
(H/X)AC
(X)F
(H/X)AE
(H)K
(H)L
(H)Cd
(H)N
(H)Gd
(H)Md
(H)Fd
1.3
1.4
1.6
0.6
3.9
1.2
1.1
0.83
1.35
0.8
1.2
0.36
1.1
2.8 1.9
4.4±0.4
2.6 1.9 6
4.8±0.3
4.2±0.2
6 2.9
1.9±0.15
4.2±0.2
3.3±0.2 1.9 1.3 4.1
2.8±0.3
1.6±0.25 7.2 5.2 3.8 2.8 1.9
1.9
1.9 1.5
3.1±0.2
1.9±0.3
1.9
2.0
4.3 1.4
0.16(B)e
0.15
3.0±0.2 2 1.8 1.5 5.2 4.5 3.9 8.4f
9.6 6.5 5.2 2.9 1.9
5.2 4.4 1.6 4.8 4.1 2.8
4.5±0.4
2.7±0.2 1.0 4.2±0.2
4.4
2.8±0.2
4.3 2.8 1.6
0.24
0.24
0.32(B)
0.19
0.32
0.12
0.14
0.2(B)
0.13(B)
0.10
0.09
aSeeFig.2.
bIn
all
cases, 1 x 106 to 2 x106
cpm of'H-labeled
polysomal poly(A) RNA was used. Hybridization wascarriedoutwith15,ig-equivalentsof the fragment DNA bound tocelluloseand RNA fractionated as described
inFig.3.
'Size of RNAspecies is basedonthe positionof 28S and 18S rRNA markers (see Fig. 1). When a region of
migratingRNA wasbroad enough to indicate size heterogeneity, the range of size values is indicated.
Fragments (X)G, (H)C, (H)G, (H)M, and (H)F are generated by only certain arrangements of the HSV-1
genomeasdescribed inthelegendtoFig.2and inTable1.
'(B)indicates that the RNA species>5kb in size migrate as a polydisperse range.
fThis band isonly partially resolved.
the invertedrepeat sequences. Forexample,the entiresequence of HindIIIfragmentM is con-tained withinHindIII fragments C and F, and the short repeat sequence whichcomprises bet-ter than 90% of HindlIl fragment M also is contained in HindIlI fragment G. All three of thesefragmentsexhibithomologousRNA of the samesizes found forHindIIIfragmentM. The sizes of RNA hybridizing toXbaI fragment G
and HindIlI fragments C and F are similarly
consistent with the sequences shared by these fragments.
Localization ofspecific viral transcripts on the HSV-1 genome. As an independent
check for the DNA-cellulose fractionation of HSVmRNA,aswellas toprovidesomelimited overlapdata for mappingHSV mRNAspecies, wecarriedout anumber of DNAhybridizations
VOL. 30,1979 HSV mRNA 811
on November 10, 2019 by guest
http://jvi.asm.org/
812 ANDERSON ET AL.
456 400 350 308
251
200 1Se 16i 50 0K U K
362
C.Hiind
III (J)280
246, 206, 166, 120
46
6
w---4501 4660 351 361 250 206 150
50 0
980 E. Xba I (F) 5F. HX (AE)
206
70e, ~~~~~~~175
69e, ~~~~~~~158,
509, ~~~~~~~125,
l O 20 30 40 f O 75 t 0 28 30 XO
lee ~~~~~~~25
315
G.Hin~d
III (K) 315 H.Hivnd
III (L)2818 C.288
245 245
216, 216,
1-5, 175,
146, 146,
185, 185,
78 78
35 35
16 28 36 46 56 10 26 38 48 58
SLICE NUMBER
FIG. 3. Size distributionofHSV-1 mRNAhybridizedtorestrictionfragmentsof viral DNA. (A) through
(M) are denaturing agarose gels ofviral RNA hybridized to HSV-1 restriction fragments fromdifferent
regionsof the genome (see Table 1). Thefragmentsarearrangedso as torepresentregions beginning from the
left ofthe Lregion throughthe shortregion ofthe P arrangement. In eachpanel, polysomal poly(A) RNA
from 4 x 107 to8 x 107 cells washybridized to 15pg-equivalents of restriction fragment DNA cellulose.
SpecificamountsofDNAcelluloseused andhybridizationefficienciesareshown in Table2.Detailsof RNA
labeling and gelfractionationare as described in thelegendtoFig. 1 and in the text. The arrows on each
panel show thepositionof the32P-labeledHeLa rRNA includedas aninternal-size marker.
of size-fractionated RNA to Southern blots of restricted DNA.Polysomalpoly(A) mRNA was labeled for 4 h with 32P, isolated from infected
cells, and fractionated on methylmercury aga-rosegels.Thelongerlabelingtimewas necessary toget RNAofsufficientspecific activityforblot hybridization. Gelswere sliced into 2- or 3-mm
slices, and Cerenkov radiationwasdetermined. Radioactivityprofiles obtainedwerequite repro-ducible, and a typical one is shown in Fig. 4.
Parallel gels with a small amount of the 32P-labeled RNA and 3H-labeled HeLa cell rRNA showed that the two peaks of indicated radio-activity migrated with sizes of 6.0 and 1.9 kb. These two peaks were used as internal size markersin all blotting experiments. Theslices weredissolvedinhybridizationbuffercontaining 65% formamide (see Materials and Methods), andRNA from individualgel sliceswas hybrid-ized tonitrocellulose stripscontainingrestriction
A. Xb@ I (C)
_ ._ . _..
9. Hind III (10)
.A i.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.505.109.407.75.477.2]HSV mRNA 813
360 K. Hind III (G)
315
278,
225,
135
96
45
l-360 320 280 240 200 160 126
88
40 0
369 326 28 240 286 166 126
86
48 0
135 129 105 90 75
60 45 38 15
IM. Hind III (F)
I
16 20 30 48 56
SLICE NUMBER
FIG. 3-continued
FIG. 4.SzedsriuinpbAKBfplyo
9S
66 56
40
3. 26
le
0
is Is 26 25 36 33 46
SL ICEMUSSEl
FIG. 4. Size distribution of polysomal poly(A)
RNAfromHSV-1-infectedHeLa cells.Inthe
experi-ment shown, [32PJRNA from 8 x 107 cells labeled
from2to6h p.i. with[32P]orthophosphatewas
frac-tionatedon adenaturingagarosegelasdescribed in
the legendtoFig. 1. Thegelwasdivided into 2-mm
slices,andradioactivitywasdeterminedby counting
Cerenkov radiation. Thesizerangeofthe RNA was
determinedasdescribedin Fig. 1,using 6- and
1.9-kb peaks (arrows) described in the text as size
markers. Only the top two-thirds oftheprofile is shown.
1 20 30 45 56
SLICE NUMBER
fragments of HSV-1 DNA. After hybridization, the strips were rinsed, assembled in order of
RNA size, and autoradiographed. Representa-tive blothybridizationstoHindIII/XbaI double-digest fragments and BglII fragmentsareshown
in Fig. 5. Hybridizations to blots of HindIII fragments alsowereperfonned (not shown).
Ta-ble 3 summarizes theresults ofourblot
hybrid-ization data. We have shown the size and loca-tion of the RNA species which hybridized to
defined fragments of the HSV-1 genome. The qualitative correlation between preparatively hybridized RNA and blot-hybridized RNA is excellent(cf. Tables 2 and 3). For example, RNA migratingat6kb,whichhybridizedto
cellulose-bound HindIII fragments J and HindIII/XbaI fragment AC (Fig. 3C and D), appearsashighly
radioactive spots in blots of those same
frag-ments (Fig. 5A) and in the overlapping BglII
fragments N and P (Fig. 5B). The discretely migrating RNA species isolated by hybridization
toHindIIIfragment K (Fig. 3G) alsoappearas
discrete spotsonblotsof thisfragment (Fig. 5A).
Therare 2.6-kb size mRNA species inHindIII fragmentIOis found inblotstothisfragmentas
I J. Hind III (N)
0L
6-;~
L. Hind III (N)
I
VOL. 30,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.505.58.452.62.552.2]A 10 6 5 1.9 1.6 I RNASize (KB)
-~-- D
- E X)F
-F
-_10
-G
-J
_- (H/X)AE
_(X)G
-K
-N
- -(H/X)AC
78r 6.0 4.1 2.8 1.9 RNA Size
(KB)
-A
-B)c
-D
-E -F -G,H
-I
=J
-K
-L
M
--N
l
0I-p
B
3.5
on November 10, 2019 by guest
http://jvi.asm.org/
HSV mRNA 815
wellas toBglII fragment 0, which is contained in it. The congruence betweenthe DNA-cellu-lose data and the blot hybridization data also holds forthebroadly migrating viral RNA spe-cies. Examination of the RNA homologous to HindIIIfragment G (Fig.3K) shows a large and broad peakof radioactivity that is2.2 to 1.6kb insize. In the blots,there is extensive
hybridi-zation of RNA in this size range to HindlIl fragment G and BglII fragment L. Inaddition to the 6-kb species, the RNA hybridizing to HindIllfragmentJ boundtocellulosemigrates
as acomplex mixture ofpartiallyresolved sizes thatrangefrom5 to 3and2 to 1.5kb (Fig.3C).
RNA hybridizing to blots of this fragment, as wellas tothecomponentBglIIfragments P and N, show a similar range of sizes. A significant portion of the RNA hybridizing toXbaI
frag-ment FDNA-cellulose migratesbroadlyin the size range of 5 to 3 kb (Fig. 3E); and RNA
hybridizingtoblots ofBglIIfragments I andD,
which overlap XbaI fragment F, also migrates broadlyin this sizerange.
Quantitative comparisonbetweentheamount of RNA ofaparticularsizehybridizingtoagiven
blot ofarestriction fragmentand that isolated from the same fragment bound to cellulose is difficult. This is because the RNA for blot
hy-bridization waslabeled for 4 hcomparedto45 min for the preparative hybridization, and be-causeexactquantitation of theamountof radio-activityhybridizing to ablot is difficult due to differential efficiency of transfer of large and small DNAfragments. However,itis clearthat,
at afirstlevel ofapproximation,the two meth-odsgivesimilar results.
There is, however, one notable quantitative
differencebetween the methods. Inexperiments
withblotted DNA,verylittle RNAhybridizesto the long repeated sequences present in XbaI
fragment G and
BglII fragment
J. In contrast, RNApulsedfrom5.25 to 6hp.i.hybridizedwell tocellulose-bound XbaIfragment G.Inexperi-mentsusing labeling times of0to 2 hp.i.,
de-scribed elsewhere (Holland et al., submitted),
several discrete sizes of RNA hybridized effi-cientlytoblots ofXbaIfragmentG. The result is consistentwith there being reduced transcrip-tion from theseregionslate, comparedtoearly, after infection.Asecondpossibility is that RNA
transcribed fromthis region from 2 to 6h p.i. hasarapid turnover rate, resulting in alower steady-state concentration of this RNA in the infected-cell compared to other RNA species. This RNA would be represented to a greater extent in short pulse-labeled RNA (used for preparativehybridization) than in long-term la-beled RNA (used in blot hybridization).
Clem-ents etal. (7) also describedareduced efficiency ofhybridizationtoblots of the longrepeatregion with RNAlabeled from0to 7hp.i.
DISCUSSION
Complexity of HSV-1 mRNA. We have measured the size of HSV-1 RNA species syn-thesized during a time late in infection when viral DNA synthesis is proceedingat arapid rate and when80 to90% of the single-strand equiv-alent of the viralgenomeis expressedasmRNA inatleast moderate abundance. There are few
significantlylabeledspecies smaller than 1.5kb and little viral mRNA which migrates as a dis-crete sizespecieslargerthan 8 to 9kb (Fig. 1). Within thisrange,therearecertain favoredsizes, suchas6, 5.4,3,andespecially1.8 to 2.2and1.5 to 1.6kb.Also,onesizeclass, that between2and 3 kb, is rare for HSV-1 mRNA. The lack of HSV-1 mRNA 2to 3 kb in size appears to be
peculiar to viral RNA since uninfected HeLa cells donotlack mRNA in this sizerange (data notshown). HSV proteins have been reportedto range in size from 15,000 to at least 250,000 daltons (11). The largestsize of HSV-1 mRNA isolated in reasonable yield (8 to 9 kb) could
readily encode thelargestpolypeptides. On the other end of thespectrum, it isinterestingthat the smallest HSV mRNA species seen in any
quantityare nosmaller than 1.2kb,a sizewith
FIG. 5. Hybridization of size-fractionatedHSV-1 mRNA toDNA restriction fragment blots. Total
32P-labeledpolysomalpoly(A) RNA from infected cells -was fractionated as described in the legend to Fig. 4.
Individual slices were dissolved in 0.4 MNa',0.1M HEPES (pH7.8),5mMEDTA, and 65%formamide(see
text) andhybridizedtostripsof Southern blots ofHSV-1 DNA, either double-digested with HindIII and XbaI
endonucleases (A) or digested withBglII endonuclease (B). The locations of the cleavage sites for these
enzymesonHSV-1 DNA are shown in Fig. 2. After hybridization and rinsing (see text), strips were assembled
inorderof size of RNA used and radioautographed with Kodak X-omat-R film. Both radioautographs were
exposedfor24hat-70°C withaKodak intensifying screen, and contact prints of the X-ray film are shown.
SizeofRNAinthe slices wasdeterminedfrom the migration of the 6- and1.9-kb RNA species as described
in thelegendtoFig.4.Positions shown for restriction fragments in the blot weredeterminedbyvisualization
of the fractionated fragments of DNA before blotting. Resolution after blotting was checked by incubating a
strip with total32P-labeledRNA (not shown). In the case of thedouble-digest blot, the bands are as described
inthefootnotes to Table1;allbands areHindIII fragments except when indicated as an Xba fragment (X)
or adouble-digest fragment(HIX).
VOL. 30,1979
on November 10, 2019 by guest
http://jvi.asm.org/
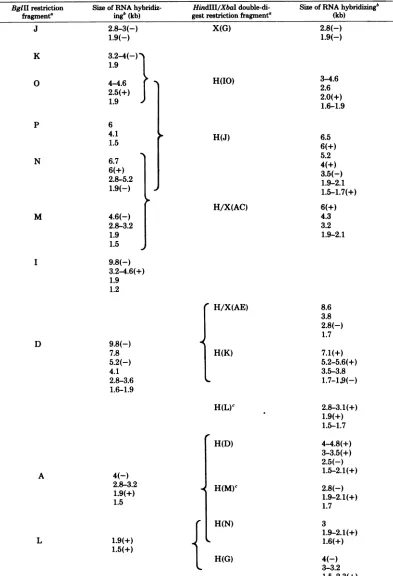
TABLE 3. LocalizationofHSV-1 mRNAbyblot
hybridization
Size of RNAhybridiz-ingb(kb)
2.8-3(-)
1.9(-)
3.2-4(-)
1.9
4-4.6 2.5(+)
1.9
6
4.1
1.5 6.7
6(+) 2.8-5.2
1.9(-)
4.6(-)
2.8-3.2
1.9
1.5
9.8(-)
3.2-4.6(+)
1.9
1.2
9.8(-)
7.8
5.2(-)
4.1
2.8-3.6
1.6-1.9
HindIII/XbaI
double-di-gestrestrictionfragmente
X(G)
H(IO)
H(J)
H/X(AC)
H/X(AE)
H(K)
H(L)C
4(-) 2.8-3.2 1.9(+) 1.5
1.9(+)
1.5(+)
H(D)
H(M)c
{ L H(N)
H(G)
Size of RNAhybridizing'
(kb) 2.8(-) 1.9(-)
3-4.6 2.6 2.0(+)
1.6-1.9
6.5 6(+) 5.2 4(+) 3.5(-) 1.9-2.1 1.5-1.7(+)
6(+) 4.3 3.2 1.9-2.1
8.6 3.8 2.8(-) 1.7
7.1(+) 5.2-5.6(+) 3.5-3.8
1.7-1.,q(-)
2.8-3.1(+) 1.9(+) 1.5-1.7
4-4.8(+) 3-3.5(+) 2.5(-) 1.5-2.1(+)
2.8(-) 1.9-2.1(+)
1.7
3 1.9-2.1(+) 1.6(+)
4(-) 3-3.2 1.5-2.3(+)
aFragments arearrangedin relativeorderontheprototype arrangement(seeFig.2).Data areshownonly
for thosefragmentswhichareclearlyresolved.
bBasedonthepositionof 6-and 1.9-kbRNA ingels(seetext). Slicesyieldasize±7%ofthe statedvalues.
Veryintensespotsareindicatedwitha(+)and faintones are indicatedwitha(-).
'BasedonblotsofHindIIIdigestswhich resolvefragmentsL andM.
816 BglIIrestriction
fragmente J
K
0
p
N
M
I
D
A
L
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.505.68.461.73.649.2]VOL. 30, 1979
atleast twice the coding capacitynecessaryfor translation ofa 15,000-dalton protein. Whereas about half of the identified viral polypeptides are smaller than 60,000 daltons, most mRNA species are 1.8 to 1.9 kb and greater in size,
providing more coding capacity than required. Since littleposttranslational processinghas been observed (11), this suggests that manyHSV-1 molecules can contain significant amounts of
untranslatedsequences.
Theviral mRNA whichhybridizestoeventhe smallest restriction fragments of HSV DNA isa complex mixture of sizes which, in some cases, exceeds the asymmetric coding capacityofthe fragment evenwhen only the mostprominent speciesareconsidered. HindIll fragment K(9.1 kb pairs in length) hybridizes efficiently with threehighlylabeled viral mRNAspecieswhose aggregatesize isover16kb.HindIII/XbaI
frag-ment AC is only 4.5 kb pairs in length, yet
hybridizeswith mRNAspecies totalingwell over 10 kb. Also, there are at least four species of viral RNA >3.5 kb in size hybridizing to the regionencompassed by BglII fragments Pand N, whosetotallengthisapproximately9kb.One explanationforcomplexpatternsof RNAspecies
bindingtoagiven region is thatsomespecies of mRNA hybridizingtospecific DNArestriction
fragments are encoded by a DNA sequence which bridges a restriction endonuclease site. Thisappears tobe thecasefor the 6-kb mRNA which hybridizes to both HindIII fragment J and HindIII/XbaI fragment AC (see below).
However, most cases cannot be explained this
simply. Forexample,withHindIIIfragment K,
we know from the hybridization patterns of neighboring DNA fragments thatatleasttwoof the large RNA molecules which hybridize to HindIIIfragmentKdonotsignificantlyextend into the sequencesadjacent tothissmallpiece
of DNA (Table 3). The aggregate size of these two species is 12 kb, which is 3 kb more than could be asymmetrically transcribed from HindIII fragmentK. Again, the localization of the RNAspecies>3.5kbhybridizingtoHindIII fragment J andBglII fragments P andN cannot be explained by RNA overlapping into
neigh-boring fragments. Suchanapparent "transcrip-tional overload" in a given region can be ex-plained by a combination of the following: (i)
symmetric transcription, (ii) asymmetric tran-scriptionandprocessing ofRNA-producing spe-cies which have overlapping nucleotide se-quences, (iii) RNA splicing eventsresulting in themRNAmoleculesbeinghomologousto more thanonelocaleonthegenome.Symmetric tran-scription probably doesnotsignificantly contrib-utesince <2% of1-hpulse-labeled poly(A) pol-yribosomalRNA frominfected cellsis
self-com-HSV mRNA 817
plementary (13; Stringer and Wagner, unpub-lished data).Since both RNA species containing shared sequences and spliced transcripts have been observed in papova- and adenoviruses (1,
5,6),it is reasonabletoexpectherpesvirusesto exhibit similar complexity in transcription and processing of viral RNA. Overlapping mRNA sequenceswould be indicatedexperimentally if 3'ends ofseveral specific mRNA sizes could be localized intoasmallregion of thegenome con-tiguouswith the restrictionfragment containing the bulk of the sequences homologous to the RNA.Preliminary results of such studiessuggest
overlappingsequences of HSV-1 mRNA do, in fact,occur(Hollandetal., submitted).
In addition to the large number of specific-sized mRNA speciesfound in localized regions of theHSV-1genome,therearealsoappreciable amounts of heterogeneously migrating RNA found hybridizing to all restriction fragments. This RNA isviral since it isnotfound in control
experiments using uninfected cells or E. coli DNAboundtocellulose. Thereare anumber of
possiblesourcesof this heterogeneously migrat-ing viral RNA. Certainly, some could arise by
degradationof RNA during workupor hybridi-zation. We have already shown that such deg-radation isquite limited, however. Furthermore, inseveralcases,similarpatternsof such
heter-ogeneously migrating RNA are seenno matter whether the RNA is size fractionated beforeor afterhybridization. Therefore,we feel this pre-cludesdegradationbeing the onlysourceofsuch
heterogeneously migrating RNA. Some RNA species may well be present in small amounts andwouldnotresolvewell frommoreabundant
species. Smallamountsofspecific-sized mRNA species hybridizing to a given restriction frag-mentcould also arise from low-efficiency hybrid-ization of RNA whosesequencesbarelyoverlap
into the fragment. Another possible source of small amounts of RNA of heterogeneous size would be thepresenceinthepolysomalpoly(A)
RNA ofpartially processed viral RNA species derived from a larger precursor. Such species have beenfound for both adenovirus and simian virus40 (5, 16).Afifthsourceofsmallamounts ofspecificsizes of RNA couldresult from
tran-scription of all four arrangements of the HSV genome (see Fig. 2). It has been reported that onlyone or twoof the arrangements of HSV-1 DNA areabletoyield viableprogenyvirus(18); however, other arrangements could be tran-scribed to produce viral RNA species whose processing could be abortive. Therelative con-tribution ofanyof thesepossiblesourcesofthe viral RNA speciespresent in lowabundancewill
beresolvedbycontinuedanalysisof HSV-1
tran-scription.
on November 10, 2019 by guest
http://jvi.asm.org/
818 ANDERSON ET AL.
Preliminarytranscriptionmap of HSV-1. Thecombined DNA-cellulose and blot
hybridi-zation data allow ustolocalizeanumber of viral mRNA sizespecies in restrictionfragments.We have constructed a preliminary transcription
mapbasedon ourdataasshown inFig.6.This map is asimplification of the data summarized inTables2 and3 in that it showsonly readily
identifiable mRNA species. We have included those mRNA's of discrete sizes whichwere ob-served from both DNA-cellulose hybridization
and blot hybridization data. Because of the greaterresolution of RNAprepared by
hybridi-zationtoDNA-cellulose,wehave used thesizes ofTable2for individual RNA
species.
Wehaveindicatedsinglespeciesof RNA whichwere re-solved after DNA-cellulosehybridizationevenif such couldnotbe resolvedbyusing blot
hybrid-ization. When RNAmigratedas arange of sizes inbothDNA-celluloseand blot
experiments,
we have shownonespecies,with bracketsindicatingthe extremes invalues.
Inmostcases, viraltranscriptsshown inFig.
6 are aligned in the middle of the restriction fragment containing homologoussequences. In
some cases, however, enough information is available to localize mRNA species more pre-cisely. The rationale forlocalization of the indi-vidual RNAspecies shown isasfollows. (i) The low-efficiency hybridization of both XbaI frag-mentG andBgIII fragmentJ madeconfirmation of the results of DNA-cellulose hybridization difficult. We have indicatedonlythetwospecies detectedbyblothybridizationonthemap. It is clear from both theDNA-cellulose hybridization and RNA displacement loop hybridization of early RNA (29) that there is considerable RNA hybridizing to this region in addition to the two species shown. We have rather arbitrarily lo-cated these twospecies tothelongrepeat, be-causetheunusually lowefficiencyof RNAblot hybridization suggests an unusual feature of the hybridization. (ii) RNA hybridizing toHindIII
fragment 10cellulose hybridizes mainly toBglII fragment0,which maps tothe right of most of this region. (iii) There are a large number of transcripts hybridizing to the region 20 to 30% of thetotalgenomelength in from the left end of the prototype arrangement. The size distri-butions of RNAhybridizing to BglII fragments
(M/X)
XG HIO H) AC _LL I I I
I0 9
71
6
5 a] 4 '14
LO 3
z
E2 I
(H/X)
XF AE HN NL HD (4M)KN HG
IJ
IEJ-I IEJ-I _ }J
BJ OK BOBPBN BM SI BD 8L
t II~~~~~
(25%) (50%) (75%)
FIG. 6. Preliminary minimaltranscriptionmap ofHSV-1 RNA. The data of Tables 2 and 3 and Fig. 3 and
5have been reduced to a minimal transcription map as described in the text. Only prominentlylabeledRNA
species areincluded. The location of the XbaI, BglII, andHindIIIfragmentsscored are shown for theP
arrangementof HSV-I DNA. The percentlength fromtheleftend of the genome is shown with arrows. When
RNAof thesamesize was found in neighboring restrictionfragments,it is shown as asingle speciesbridging
therestriction cleavage site, provided the radioactivity in bothfragmentsisequivalent,such as for the6-kb
RNAspecies in HindIII fragment J andHindIII/XbaIdouble-digestfragment AC. The length of the linesis
proportional to the relative length of the RNA in question. The RNA species with bracketed sizes indicate
RNA which migrates broadly in the range shown. The RNA species shown under the region spanned by
HindIIIfragmentD, which arisesfromthe IL andILsconformationsof the viral DNA, are based on blotdata
andareshown in the unique region due to the low hybridization of the long repeat region found in XbaI
fragmentG andBglIIfragmentJ.
I i
6 - - Ir 4
on November 10, 2019 by guest
http://jvi.asm.org/
[image:14.505.120.406.345.558.2]HSV mRNA 819
0, P, andN,which subdivide thisregion, indi-catethatmostofthespecies of RNAhybridize to the region on either side of the right-hand
HindIlI site generating fragment J. (iv) The RNA species of size around 8 to 9 kb found hybridizingtoXbaIfragment F cellulose corre-sponds to RNA of this size found in blots of BglII fragments I and D and HindIII/XbaI
dou-ble-digest fragment AE. For clarity, we have shown thisas a single species, since one RNA molecule of this size islarge enoughtobridge all the restriction sites generating the fragments.
However,preliminary experimentssuggestthat twodiscrete 3'ends for RNA of this size occur inthisregion(datanotshown). We have shown RNA species on either side of thejunction be-tweenXbaIfragmentF andHindIII/XbaI dou-ble-digest fragment AE, dependingon whether RNA of this size alsohybridizestoblots ofBglII
fragment I. (v) Three RNA species have been shownmapping in HindIIIfragmentK because of the resolution of these after preparative hy-bridization (Fig. 3G). However, the midsized RNA hybridizing toblots of HindIII fragment
Kconsistentlyshowsabroader size distribution than the7.4-and 3.8-kbspecies, suggestingthat more than one RNA species may befound in this size range. (vi) The RNA hybridizing
effi-cientlytoblots ofHindIlI fragmentDhave been localized totheunique regiontothe left of the long repeat sequences since RNA hybridizes verypoorlytoboth XbaI fragment G andBglII fragment J, which also contain the longrepeat sequence. Finally, (vii) the 4-kb species,aswell as arangeofsmaller species of viral RNA species foundhybridizingtoHindIIIfragmentG cellu-lose and blots, are shown in the short unique region of thisfragmentsincethey donot
hybrid-ize to blots of HindIII fragment M or BglII
fragment L, which, like HindIII fragment G,
contains theshortrepeatregion.
Thetranscriptionmapshown inFig.6 under-estimates the number of significantly labeled HSV-1 mRNA species. This is especially true forsmaller RNA species found in large restric-tionfragmentssuchasXbaIfragmentF. Butin
spite of this and otherlimitations, certain pat-terns in the location of HSV-1 mRNA on the viral genome can be seen. The region of the genome from 10to 15% in from the left end of theprototypearrangement (BglIIfragment K) hybridizes mainly to RNA of size 1.9 to 2 kb. RNA displacement loop hybridization of early HSV RNAalsoshowslowhybridizationinthis area (29). At least two regions ofthe genome, theregion20 to30%infrom the left end(HindIII fragment J) and the region 50 to 60% in from that end(HindIII fragmentsKandL),hybridize to numerous RNA species. Higher-resolution
mapping ofRNAspeciesin theregion covered byXbaIfragment F mayreveal other such re-gions. Whereas RNA of 1.6 to 2 kb in size is
found throughout the genome, a significant
amountof RNA smaller than 1.5kb (1.2 kb) is found only in BglII fragment I. Well-resolved RNA larger than 5 kb is found in significant
amounts only toward the center of the long uniqueregionof thegenome(25to60% from the left end); furthermore, little viral mRNA>4 kb in size isfound in the shortregionand in either thelongorshortrepeatsequencesof the HSV-1genome.Specific features of HSV-1 RNA
me-tabolism which result in thesepatternsare un-known.
Correlation of the transcription map with the HSV-1 genetic map. Of the nearly 50proteins whichhave beenidentifiedasviral, about half have been localized on the HSV-1 genome by using intertypic recombinants be-tween HSV-1 and HSV-2 (19). Itis interesting to note that structural (y) polypeptides map throughout the long unique region,with a large number mapping between 20 and 30% and an-otherlargenumber between50and 60% in from the left end of the genome in the prototype arrangement. The very large structural poly-peptide, ICP-2, which would require a large mRNA speciestoencode it, has beenlocalized
inthis latter region.These areas of the genome correspondtoregions contained by HindIII frag-ment J and HindIII fragments K and L; both regions encode large mRNA species. Itshould be noted that thesetworegionsaresymmetrical around the rotation axis ofthe longsequenceof theHSV-1genome(see reference 29)sothat the question of whether the prototype arrangement or onewiththe long region invertedis
biologi-callyactive doesnotinterfere with the correla-tion.Further correlation of the HSV-1 transcrip-tion map with polypeptide maps must await studies on the biological activity of the RNA
specieswehaveidentified in thisreport.
ACKNOWLEDGMENTS
Wethank B. Gaylord and L. Tribble for excellent technical help.We also thank J. Holland for the gift of HeLa cells.
This workwassupportedby Public Health Service grant CA 11861 from the National Cancer Institute. K.P.A. was supported byPublic Health Servicepredoctoraltraining grant GM07311 from the National Institutes of Health.
LITERATURE CITED
1. Aloni, Y., R. Dhar, 0.Laub, M. Horowitz, and G. Khoury.1977.Novel mechanism for RNA maturation: theleader sequences of simian virus40mRNA are not transcribed adjacent to the coding sequences. Proc. Natl. Acad. Sci. U.S.A.74:3686-3690.
2. Bailey,J.M., and N. Davidson.1976.Methylmercury asareversibledenaturing agent for agarose gel electro-phoresis. Anal.Biochem.70:75-85.
3. Barrell, B. G., G. M. Air, and C. A. Hutchison III.
VOL. 30,1979
on November 10, 2019 by guest
http://jvi.asm.org/
820 ANDERSON ET AL.
1976.Overlapping genes inbacteriophageOX174. Na-ture(London) 264:3441.
4.Bartkoski, M.,and B. Roizman. 1976. RNAsynthesis in cells infected with herpessimplex virus. XIII. Differ-encesin themethylation patternsofviralRNAduring thereproductivecycle. J. Virol. 20:583-588.
5. Berget,S., C. Moore, andP. A.Sharp.1977.Spliced segments atthe 5' terminus of adenovirus 2latemRNA. Proc.Natl.Acad.Sci.U.S.A.74:3171-3175.
6. Chow, L T.,R. E.Gelinas, T. R.Broker, andR. J. Roberts. 1977. Anamazingsequence arrangement at the 5' end ofadenovirus2messengerRNA.Cell12:1-8. 7. Clements, J. B., R. J.Watson,and N.ML Wilkie.1977. Temporal regulation ofherpessimplex virus type 1 transcription: location oftranscripts on the viral ge-nome.Cell 12:275-285.
8. Grafstrom,R.H.,J.C.Alwine,W.L.Steinhart,C. W.Hill,and R. W.Hyman.1975.The terminal repe-tition ofherpessimplexvirus DNA.Virology 67:144-157.
9. Hayward, G. S.,R. J.Jacob, S.C.Wadsworth,and B.Roizman. 1975.Anatomyofherpes simplexvirus DNA:evidencefor fourpopulationsofmoleculesthat differ in the relative orientations of theirlongandshort components. Proc. Natl. Acad. Sci. U.S.A. 72:4243-4247.
10.Holm, C. S., S. G.Oliver,A. M.Newman, L.E. Hol-land, C.S.McLaughlin,E. K.Wagner,and R.C. Warner. 1978. The molecularweightofyeastP1 dou-ble-strandedRNA. J.Biol. Chem. 253:8332-8336. 11.Honess, R. W., and B. Roizman. 1973. Proteinsspecified
byherpes simplexvirus.XI.Identificationandrelative molar rates ofsynthesisof structural and nonstructural herpes viruspolypeptidesinthe infected cell. J. Virol. 12:1347-1365.
12.Honess, R. W., and B. Roizman. 1974.Regulationof herpesvirus macromolecular synthesis.I. Cascade reg-ulation ofthesynthesisof three groups of viralproteins. J. Virol. 14:8-19.
13. Jacquemont,B.,and B.Roizman.1975.RNAsynthesis incellsinfected withherpes simplexvirus. X.Properties ofviral symmetrictranscriptsand ofdouble-stranded RNApreparedfrom them. J. Virol. 15:707-713. 14.Jones, P.C., G. S. Hayward,and B.Roizman.1977.
Anatomyofherpessimplexvirus DNA. VII. a RNA is homologoustononcontiguoussites in both the L and S componentsof viral DNA. J. Virol. 21:268-276. 15. Kozak,M., andB.Roizman. 1974.Regulationof
her-pesvirus macromolecularsynthesis:nuclearretention of nontranslated viral RNAsequences. Proc. Natl. Acad. Sci. U.S.A. 71:43224326.
16. Lai,C-J.,R.Dhar,and G.Khoury.1978.Mappingthe splicedandunsplicedlatelyticSV40RNAs. Cell14: 971-982.
17. McMaster,G.K.,andG. C. Carmichael.1977.Analysis ofsingleand double stranded nucleic acidson polyacryl-amide andagarose gelsbyusingglyoxaland acridine orange. Proc.Natl. Acad.Sci. U.S.A.74:4835-4838. 18.Morse,LS.,T. G.Buchman,B.Roizman,and P. A.
Schaffer.1977.Anatomyofherpes simplexvirusDNA. IX.Apparent exclusion ofsomeparentalDNA arrange-ments inthegeneration ofintertypic(HSV-1xHSV-2) recombinants. J. Virol.24:231-248.
19. Morse,L.S.,LPereira,B.Roizman,and P. A. Schaf-fer. 1978. Anatomy of herpessimplex virus (HSV) DNA. X.Mappingof viralgenesby analysisof polypep-tidesand functionsspecifiedbyHSV-1x HSV-2 recom-binants. J.Virol. 26:389-410.
20. Moss,B.,A.Gershowitz,J.R.Stringer,L.E.Hoiland,
and E. K. Wagner. 1977. 5'-Terminal andinternal
methylated nucleosides in herpes simplex virus type 1 mRNA. J. Virol. 23:234-239.
21. Noyes, B. E., andG. R. Stark. 1975. Nucleic acid hy-bridization using DNA covalently coupled to cellulose. Cell 5:301-310.
22. Palmiter,R. D. 1974. Mg++ precipitation of ribonucleo-protein complexes. Expedient techniques for the isola-tion ofundegraded polysomes and messenger ribonu-cleic acid.Biochemistry13:3606-3614.
23. Plummer,G., C.R.Goodheart,D.Henson, and C. P. Bowling.1969.Acomparativestudyof the DNA den-sity and behavior in tissue cultures of fourteen different herpesviruses. Virology 39:134-137.
24. Sheldrick, P., and N. Berthelot. 1974. Inverted repeti-tions in the chromosome ofherpes simplex virus. Cold SpringHarborSymp.Quant. Biol. 39:667-678. 25. Silverstein, S., R. Millette, P. Jones, and B. Roizman.
1976. RNAsynthesis in cells infected withherpes sim-plex virus. XII.Sequence complexity and properties of RNA differing in extent ofadenylation. J. Virol. 18: 977-991.
26. Skare, J., and W. C.Summers. 1977. Structure and function ofherpesvirusgenomes.II.Eco RI, Xba I, and Hind III endonucleasecleavagesites on herpes simplex virus type 1 DNA. Virology 76:581-595.
27. Southern,E. 1975.Detectionofspecific sequences among DNA fragments separated by gel electrophoresis. J. Mol.Biol. 98:503-517.
28. Stringer, J., L. Holland,R.Swanstrom,K.Pivo, and E.Wagner. 1977. Quantitation of herpes simplex virus type 1 RNA ininfectedHeLa cells. J.Virol. 21:889-901. 29. Stringer,J.R.,L E.Holland,and E. K. Wagner. 1978. Mappingearly transcripts of herpes simplex virus type 1byelectronmicroscopy. J. Virol. 27:56-73.
30. Swanstrom, R., K.Pivo, and E. Wagner. 1975. Re-stricted transcription of the herpes simplex virus ge-nomeoccurring earlyafter infection and in the presence of metabolic inhibitors. Virology 66:140-150. 31. Swanstrom,R., and E. Wagner. 1974. Regulation of
synthesis of herpes simplex type 1 virus mRNA during productive infection. Virology 60:522-533.
32. Wadsworth, S.,R. J. Jacob, and B. Roizman. 1975. AnatomyofherpessimplexvirusDNA.II. Size, com-position,and arrangement ofinverted terminal repeti-tions. J. Virol. 15:1487-1497.
33. Wagner, E., R.Swanstrom, and M.Stafford. 1972. Transcription oftheherpes simplex virus genome in human cells. J.Virol.10:675-682.
34. Wagner,E. K., R.L.Swanstrom, M. Rice, L. Howell, and J. Lane. 1976.Variationin the molecular size of theDNA fromcloselyrelated strains of type 1 herpes simplexvirus. Biochim. Biophys. Acta 435:192-205. 35. Wagner,E.K.,K. K.Tewari, R. Kolodner, and R. C.
Warner.1974.The molecular size of theherpes simplex virus type 1 genome. Virology 57:436-447.
36. Weiss, S.R., H. E. Varmus, and J. M. Bishop. 1977. The size and geneticcomposition ofvirus-specific RNAs inthecytoplasm of cells producing avian sarcoma-leu-kosis viruses.Cell 12:983-992.
37. Wellauer, P. K., and I. B. Dawid. 1973. Secondary structuremaps of RNA: processing of HeLaribosomal RNA. Proc.Natl.Acad.Sci. U.S.A. 70:2827-2831. 38. Wilkie,N. M. 1976.Physical maps for herpessimplex
virustype 1 DNA forrestriction endonucleases Hind III,Hpa-1, and X. bad.J.Virol. 20:222-233.
39. Wikie,N.M.,and R.Cortini.1976.Sequence arrange-ment inherpes simplex virus type 1 DNA: identification ofterminal fragmentsinrestriction endonuclease di-gests and evidence for inversions in redundant and unique sequences. J. Virol. 20:211-221.
on November 10, 2019 by guest
http://jvi.asm.org/