0022-538X/79/01-0161/09$02.00/0 No. 1
Effect of Actinomycin
D
onthe Expression
of Herpes Simplex
Virus-Common Surface
Antigen in Cells
Transformed by
Herpes Simplex
Virus Type
2
SUSUMU KIMURA,'* KATSUICHIROOKAZAKI,2 NAGAYUKIYOSHIDA2AND
YOSHINARI OHNISHI'
Department of Bacteriology, Schoolof Medicine, TokushimaUniversity,1 andDepartment ofMicrobiology,
Schoolof Pharmacy, TokushimaUniversity of ArtsandScience,2 Tokushima770,Japan
Received forpublication7April1978
Using rabbit antiserum hyperimmune toherpes simplex virus (HSV) type 1,
the expression of HSV-common surface antigen(s) was studied by indirect
im-munofluorescencetestsin cells transformedby HSVtype2and in derivedtumor
cells. The following resultswere obtained. (i) AntiserumtoHSV type 1 reacted
specifically with surface antigenpresent onthe plasma membrane of both HSV type2-infected and HSVtype2-transformed hamstercells.(ii)The expression of
this antigen was enhanced in the absence ofactive protein synthesis in
trans-formedcells, but notin tumorcells, after culture for3to5 h at37°C. (iii) This
enhancement ofexpressionwasmaintained for20h in thepresenceof actinomycin
D, but this prolonged expression required active protein synthesis. (iv) The
enhancing effect observed in thepresenceof actinomycin D continued forsome
timeafter removal of the drug, for example, for20h after5h oftreatmentwith
2 ,ug/ml of actinomycin D perml. Actinomycin D hadno detectable effect on
antigen expression in tumor cells. (v) Theprotease inhibitor antipaininhibited the actinomycin D-enhanced expression withoutcausing significantcell damage
but didnotmodify the transient enhanced expression of antigen when cellswere
seeded in theabsence ofactinomycin D. These results indicate that in transformed cells antigen expression canbe enhanced inatleasttwoways.
It is well known that mammalian cells are
transformed
oncogenically
in vitro by herpes simplex virus(HSV)
type1(HSV-1)
andtype 2(HSV-2) (1, 5, 6, 13-16, 28). Viral DNA
se-quences (4, 10, 19),
HSV-specific
RNA (2), andHSV-specific
antigen (1,5, 13-16, 28) have been demonstrated in cells transformed by HSV-2. Moreover, viral genetic information has beenconcluded to be present in HSV-transformed cells fromthefollowing findings. (i) Acquisition of
HSV-specific
thymidine kinasehasbeendem-onstratedinthymidine kinase-negativemouseL
cells after in vitro transformation with HSV-1
(20). (ii) Complementation between
tempera-ture-sensitive (ts) mutants ofHSV-2 and
resi-dent, functional HSV genes in HSV-2-trans-formedcells has been demonstrated atthe
non-permissivetemperature (12, 17).
Recently,ithas been demonstrated that
sev-eral type-common and type-specific HSV anti-genscanbeidentifiedby
immunological analysis
of HSV-1- and HSV-2-induced glycoproteins
(30) and that type-common antigen(s) is also
detectableonthesurface ofcells transformed
by
HSV(21, 22).
In an attempt to
identify
theHSV-specific
geneproducts
expressed
intransformedcells,weobservedanHSVtype-commonsurface
antigen
(CSA) which increased after
actinomycin
D(ActD) treatment of cultures transformed by a ts
mutantofHSV-2. Thispaper reports studieson
theexpression of CSAintransformedandtumor
cells, the doses ofAct D which enhance
CSA
expression and the kinetics ofits
effect,
andthe effect of a proteaseinhibitor,antipain,
onActD-enhancedCSA
expression.
MATERIALS AND METHODS
Cell lines. HSV-2-transformed cells and the
de-rivedtumorcellsused in thisstudywereall isolated
by the routine method fromhamster (inbredstrain
LSH)embryo fibroblast cellcultures,asreported
pre-viously by Kimura etal. (13). Transformed cell line
155-4(passages130to180)wasobtainedafter infection ofcellswith theDNA-negative,mutantB5 of HSV-2
strain 186 at the nonpermissive temperature (38°C)
and then passagedserially at37°C,because thiscell line wasnotstabletomaintenanceat38°C.Celllines
U-15, U-26, and U6V-7 (passages 131 to 135) were
obtained after infection with UV-inactivated strains
186 (U-15 andU-26) and333 (U6V-7) ofHSV-2,
re-161
on November 10, 2019 by guest
http://jvi.asm.org/
spectively.Two otherhamstertumorcelllines,
TSV-5 (passages232 to 249) andBAT-6 (passage> 100),
induced by simian virus40andbovineadenovirus type
3,werekindly suppliedby T. Kurimura(Tottori
Uni-versity,Tottori, Japan) and Y. K.Inoue(Kyoto
Uni-versity, Kyoto, Japan),respectively.Primaryhamster
embryofibroblasts(HEF)wereobtained from
13-day-old hamster embryos (inbred National Institutes of
Healthstrain). All cellsweregrownat37°CinEagle
minimum essential medium (MEM) supplemented
with 10%fetal calf serum(FCS) and 0.075% NaHCO3
for cultures in closed vessels and 0.225%NaHCO3for
cultures in open vessels.
Virus. HSV-1 (strain KOS), HSV-2 (strain 186),
and the DNA-negative ts mutant (ts B5) of HSV-2
(strain186) used in thisstudywerekindly providedby
P. A. Schaffer (SidneyFarber CancerInstitute,
Har-vard Medical School, Boston, Mass.). Virus stocks
were grown in human embryonic lung cell cultures
withEagle MEMcontaining 5%FCS,andvirus assays
wereperformedasdescribedpreviously (8, 25).Virus
stocks had the following titers inhuman embryonic
lung cell cultures: HSV-1, 6 x 107 PFU/ml at 37°C;
HSV-2, 2 x 107 PFU/ml at 37°C; ts B5, 2 x 107
PFU/mlat340C.
Antisera. Two hyperimmune rabbit antisera to
HSV-1(strainKOS) werepreparedindependently by
immunizing albino rabbits intraperitoneally with
HSV-1 in Freundcomplete adjuvant (Difco
Labora-tories,Detroit, Mich.)onceaweek for10weeks. The
animalwasbled 1 week after the finalinjection.The
neutralizingantibody titerstoHSV-1 of thetwo
anti-serawere1:1,280 and1:320,respectively,in 50%plaque
reductiontests.Anti-HSV-1serum(serumA in Table
1) possessing a higher neutralization titer was used
mainly in thisstudy. Additionalhyperimmunerabbit
antiserumtostrain HF of HSV-1waskindly provided
by Y.K.Inoue and usedas areference antiserum with
aneutralization titer of1:640.Fluorescein-conjugated
goatantiserumtorabbit 7Simmunoglobulin (FGAR)
waspurchased from HylandDivision,Travenol
Lab-oratories,Inc., CostaMesa,Calif.Nonspecific
reactiv-ity was removed from these antiserabytheabsorption
technique described below.
Absorption technique.Theabsorption technique
describedbyFlanneryetal.(9)wasused,with minor
modifications. Four milliliters of undiluted rabbit
anti-HSV-1serumwasmixed with1gof rabbit liverpowder
suspended in4ml of Trisbuffer(pH 7.4). The mixture
wassonically treated inanicebathat 10kcyclesfor
two 1-minperiods,shakenat370 for 1 hand at40C
overnight,and thencentrifugedat100,000xgfor 1 h.
The supernatant fluid was mixed with the pellet of
human embryonic lung cells (108 cells), sonically
treated, incubated at 370C for 1 h and at4°C
over-night,and thencentrifugedat100,000xgfor 1h. The
supernatantwasabsorbedathirdtimewith108intact
HEF cells for 1 h at 370C and subjected to sonic
treatment. Thesuspensionwasincubatedat37°C for
1 h and at 4°C overnight and then centrifuged at
100,000 xg for 1h.Absorbed anti-HSV-1 serum was
used todetectHSV-CSA.Fortestingthespecificityof
thisantiserum,fourdifferentcelltypes(155-4,TSV-5,
and HSV-2- and ts B5-infected HEF) were used in
absorptionexperiments.
Celllines 155-4andTSV-5wereseeded in150-mm
petridishes(FalconPlastics,Oxnard, Calif.)atinocula
ofabout1x 107cells perdish andharvested 3 h later
by gentle
trypsinization. HEF cellswereinfectedatamultiplicity of 3PFU/cellwith HSV-2at37°C orts
B5at34°C. The HEF cellsinfected with HSV-2 and
ts B5were harvested with a rubberpoliceman 10 h
and 20 h later, respectively, when the numbers of
HSV-CSA-positivecellsweremaximal. The antiserum
(0.7 ml)wasabsorbedagaintwice,with each cellpellet
containing6x107cells,bythe abovemethod,andthe
absorbedserum was testedforHSV-CSA reactivity.
FGAR serum (4 ml) was also absorbed with 1 g of
rabbit liverpowderand108HEF cells.
Detection ofHSV-CSA. Indirect
immunofluores-cence tests were used for detection of HSV-CSA.
Before experiments, the test cells were grown and
passagedatleast three times understrictly controlled
conditions: thatis, 10 ml ofcellsuspension (105cells
perml) wasinoculated intoaculture bottle(50 cm2)
andincubated at37°C for48h.After thistime, the
culturewasalmost confluent(about 90%)andthecell
number had increased four fold (i.e., 4 x 105 ml).
Conditioned cellsuspensions (105 cells perml) were
seeded into 5 ml ofmedium in 60-mm petri dishes
(Falcon Plastics) and incubated at 37°C in a CO2
(5%)incubator. Atdesignated times,cellsweretreated
with 0.25% trypsin (1:250; Difco Laboratories) for 1
min andwashed three times withbarbital buffer(pH
7.4) containingbarbital(3x
1i-'
M), sodium barbital(1.8x 10-3M),0.85% NaCl, Mg2+ (5x 10-4 M),Ca2+
(1.5x 10-4 M),and 0.1% gelatin (GBB) (31) by
cen-trifugationat800xgfor10min.Samplesof1dropof
antiserum absorbed with various cellsornormalrabbit
serum at a final dilution of 1:4 were added to cell
suspensions (6 x 105 to 10 x 105 cells) incentrifuge
tubes. Thesuspensionswereincubatedat37°C for30
min,washed threetimes withGBB, and then treated
for30 min at 37°C with 1 drop of absorbed FGAR
serumdiluted1:8andwashedthreetimes with GBB.
Washed suspensions were mounted in 50% glycerin
solution,and 103 cells persample were observed with
a reflected light fluorescence microscope (Olympus
OpticalCo.Ltd., Tokyo, Japan). The number of viable
cellswasdeterminedby trypan blue exclusion.
To test the effects ofvarious conditions on the
expression of HSV-CSA in 155-4 cells (see Table2),
cellsdetachedby pipettingweresuspendedin1ml of
phosphate-buffered saline withoutCa2" and Mg2' and
treated with 1 ml oftrypsin (0.5%) or 1 ml of
phos-phate-buffered saline without Ca2' andMg2"for 1 and
3 min, respectively. Trypsinization was stopped by
addition of15ml ofEagle MEM containing 10% FCS
andbycentrifugation. Some cell suspensions withor
without trypsinization wereimmediately assayed for
indirectimmunofluorescence. Other suspensions (1 x
105to25 x 105 cells per sample) wereincubated with
5ml ofEagle MEM containing 10% FCS, in 60-mm
dishes asmonolayers orintubes (15 by 150mm) as
suspensions, for3h at4or37°C and then assayed for
indirectimmunofluorescence.
Inhibitors. Act D and puromycin-dihydrochloride
(PM) were obtained from Boehringer Mannheim,
Mannheim,West Germany, and Nutritional
Biochem-icalsCo., Cleveland, Ohio, respectively. The protease
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
SURFACE ANTIGEN IN HSV-2-TRANSFORMED CELLS 163
inhibitor antipain [(1-carboxy-2-phenylethyl)
car-bamoyl-L-arginyl-L-valyl-argininal] (26) was kindly
providedby T. Uchida (Osaka University, Osaka,
Ja-pan). Ten times-concentrated stock solutions were
prepared, passed through 450-nm membrane filters
(Millipore Corp., Bedford,Mass.) and storedat-20°C.
Routinely, cells were treated with Act D (0.5 to 4
,ug/ml)whentheywereseeded into60-mmdishesand
then incubated for the desired time at 37°C. For
further incubation withoutAct D,cells werewashed
twice with Tris bufferandreincubatedinfresh Eagle
MEM containing10%FCSat37°C.Antipain (0.5 mM)
wasalsoaddedto ActD-treated and untreated cells
at the time ofseeding, and the cells wereincubated
with antipain for30 h, even when the medium was
changed.
Incorporation of[3H]leucine.A5-,uCiportion of
L-[4, 5-3H]leucine (40 to 60 Ci/mmol; New England
Nuclear, Boston, Mass.) was added to 60-mm petri
dishes for10h. Thecells werethenwashedoncewith
cold(4°C) Trisbuffer, harvested, and centrifuged at
800xgfor10min. Pelletssuspended in1ml ofcold
Tris bufferweresonicallytreated inanice bathat10
kcycles for20s,treated with1mlof20%trichloroacetic
acid for 20min at 0°C, and then boiled for 15 min.
The suspensionswereappliedtoWhatman GF/Cfilter
disks, usinganaspirator. The diskswerewashed three
timeswith 5%trichloroacetic acid andoncewith
ethyl-alcohol, dried, and placed in scintillation vials. Ten
milliliters ofscintillationfluidwasaddedtoeach vial,
andradioactivitywasmeasured inaliquid scintillation
counter(Packardmodel 3385). RESULTS
Specificity of anti-HSV-1 serum. To test
the specificity of the reaction of anti-HSV-1
serum with HSV-CSA on HSV-2-transformed
cells, three HSV-2-infected, five HSV-2-trans-formed, and two other DNA virus-induced
tu-morcellsanduninfected HEFcellsweretested
for reactivitytothree anti-HSV-i seraabsorbed
withHEF cells (Table 1). HSV-2at370C andts
B5atthe permissivetemperature (340C) caused
anincrease in the number ofCSA-positive cells
with timeafter infection,tomaximaof 10 h(58% positive cells) and 20 h (48% positive cells),
respectively,after infection; however,atthe
non-permissivetemperature (38°C),tsB5induceda
maximum of only 13% positivecellsat10hafter infection, and the number of positivecellsthen
rapidly decreased. A positive reaction (ranging from4 to36%CSA-positive cells) wasobserved
in thefive HSV-2-transformed cell lines, regard-less of the strain ofHSV-1 used for the
antise-rumpreparation; themostpositive reactionwas
found in ts B5-transformed (155-4) cells (23to
36%CSA-positive cells).No specific reactionwas
observed in tumorigenic transformed cells
in-ducedwith simian virus 40orbovineadenovirus
type 3 or in control HEF cells ('0.7%
CSA-positive cells). In the experiments carried out
below, antiserum to strain KOS of HSV-1
(se-rum A in Table 1), which has ahigher titer of
neutralizing antibody, wasused to detect HSV-CSA. Both HSV-2 at 370C and ts B5 at 340C
(Fig. 1A) induced predominantly ringlike
fluo-rescence, withafew dotted and arc-shaped
re-gions of fluorescenceonthe surface of HEFcells.
Although the intensityofstaining of 155-4cells
TABLE 1. DetectionofHSV-CSA inHSV-2-infectedand-transfornedhamster cellsby indirect
immunofluorescencea
No.ofceH
~~~~~~~CSA-positive
cellsCell type No. ofcell Virus used(strain)
SerumA Serum B Serum C
HSV-2 infectedHEF,at37oCc HSV-2(186) 58.0
tsB5 infectedHEF,at
34oC'
HSV-2(tsB5of strain186) 48.0tsB5infectedHEF,at380Cd HSV-2(tsB5ofstrain186) 13.0
U6V-7 135 HSV-2(333) 10.4 4.2 8.4
U-15 Te 86 HSV-2(186) 8.0 7.9 8.1
U-26 T 126 HSV-2(186) 5.2 5.5 5.9
155-4 138 HSV-2(tsB5 ofstrain 186) 36.1 23.4 35.0
155-4T 114 HSV-2(tsB5 of strain 186) 7.0 9.0 7.0
TSV-5 246 SV40 0.5 0.7 0.6
BAT-6 >100 BAD-3f 0.3 0.6 0.6
HEF 5 None 0.4 0.5 0.5
a
All cells
exceptinfectedcells
wereharvested3to5 h afterseedingat37°C and stained withanti-HSV-1serun absorbed with HEFcells.
bAtotal of
103 cells
werecounted.SeraAandBwererabbit antiserapreparedindependently againststrainKOS of HSV-1 andserumCwasrabbitantiserumtostrain HFofHSV-1.Neutralizingantibodytitersto
HSV-1of seraA,B, and Cwere1:1,280,1:320,and1:640,respectively,in50%plaquereductiontests.
cHEF
cells
infectedwith HSV-2(3PFU/cell)wereharvested10hafterinfection.d HEF
cells
infected withtsB5(3PFU/cell) at34and380C
wereharvested20and 10h,respectively,
afterinfection when thenumbers ofpositivecellsweremaximal.
'T representscultured
cells
derived fromtumorspassedoncein vivo.fBAD-3,Bovine adenovirustype3.
VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
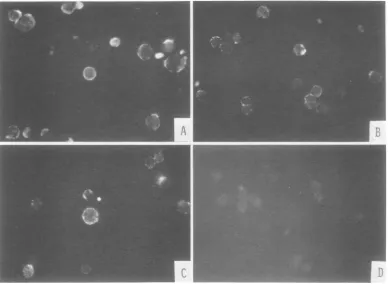
FIG. 1. SurfacefluorescenceobservedinHSV-2-infectedandtsB5-transformed (155-4)cellsaftertreatment
with anti-HSV-I serum absorbed with various cells in indirect immunofluorescence tests. (A) HEFcell
infectedwithtsB5(20hafterinfectionat
34QC)
stainedwith anti-HSV-IserumabsorbedwithHEFcells. (E155-4cells3 hafterseedingwithanti-HSV-Iserumabsorbedwith HEF cells. (C)155-4cells 3hafterseeding stained with anti-HSV-I serum absorbed with TSV-5cells. (D) 155-4 cells 3 h afterseedingstained with normalrabbitserum orwithanti-HSV-I serumabsorbed withHSV-2-infected, tsB5-infected,or155-4cells. was slightly less than that of the two types of
HSV-infectedcells,distinctring-shapedand dot-ted regions offluorescence were observed
pre-dominantly, with a few arc-shaped regions of fluorescence onthe surface oftransformedcells
(Fig. 1B).
To confirm thespecificityof thereaction, anti-HSV-1serum wasabsorbed with TSV-5(simian
virus 40-induced tumor cells), HSV-2-infected,
ts B5-infected, or the transformed 155-4 cells
andthen tested forHSV-specificreactivity with
155-4 cells. Reactivity with 155-4 cells was
re-tained after absorption with TSV-5 cells (Fig. 1C); however, afterabsorption with HSV-2- or
ts B5-infected or 155-4 cells, the reactivity of this antiserumwasreducedtothe level of that of normal rabbitserum (Fig. 1D). These results indicate that the surfacefluorescence observed with transformed 155-4 cells is HSVspecific.
Expression ofHSV-CSA in transformed
and tumor cells. Preliminary experiments
showed that 155-4cellsdoubled every 25 hand that when5 x 105cellswereinoculated into
60-mm dishes at 37°C, CSA-positive cells were
more numerous 3 to 5 haftertrypsinization and
seeding (30to 40% positive) than after 15 to48
h (8 to 20%positive). To testwhether trypsini-zation itself induces activation of CSA, it was
carried out atvarying times, with varying sizes of cellinoculum, and under varying incubation conditions (Table2). No activation of CSA was observed in 155-4 cells that weresimply trypsin-ized for 1 or 3 min or after addition of fresh medium. PartialactivationofCSA wasobserved when: (i) cells were not trypsinized; (ii) cells
were trypsinized for 1 min and then incubated at4°C for 3 h; (iii) large inocula (2.5x 106cells) or small inocula (105 cells) were used; and (iv) cells were incubated for 3 h at 4 or 37°C after trypsinization for 3 min. Activation of CSA in 155-4cells was markedonly when the cellswere
trypsinized for 1 min andincubated at an appro-priate inoculum (5 x 105 cells per dish ortube) for 3 h at 37°C. Activation was observed with cell suspensions and even moremarkedly with monolayers. Thus, trypsinization was necessary
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.501.67.456.64.348.2]SURFACE ANTIGEN IN
TABLE 2. Various conditionsaffecting theexpression of HSV-CSAin155-4 cells
a
.. Cellinoculum/ Incubation conditions CSA-positive
Trypsinization
sample(x105)
Stateb Time (h) Temp(°C) cells(%)c
None 5 Noincubation 12.2
5 Suspension 3 4 13.1
5 Suspension 3 37 15.5
5 Monolayer 3 37 14.4
Medium
changed
25 Monolayer 3 37 8.41min 5 Noincubation 6.2
25 Noincubation 9.6
5 Suspension 3 4 18.2
5 Suspension 3 37 30.5
1 Monolayer 3 37 13.8
5 Monolayer 3 37 37.5
25 Monolayer 3 37 20.6
3mm 5 Noincubation 5.0
5 Suspension 3 4 12.2
5 Suspension 3 37 18.6
5 Monolayer 3 37 18.9
aTrypsinization wasperformedasdescribed in the text.
b
Cells
wereinoculated into dishes(monolayer)orintubes (suspension) with gentleshaking. No incubation,Cell suspensionswereimmediately assayed for indirect immunofluorescence.
cAtotal of103
cells
werecounted.dTheculture medium was replaced by fresh medium2days after seeding.
butnotsufficient for activation.
Next, the relationship between CSA expres-sion and cell growth was examined in trans-formed(155-4) andtumor (155-4T) cellsduring
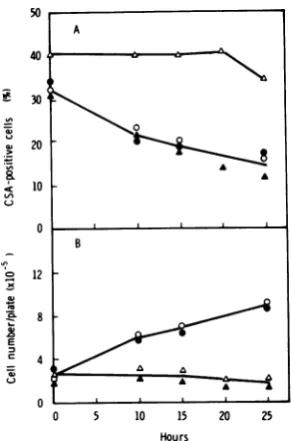
incubation at37°C (Fig. 2). In 155-4 cells a peak of CSA-positive cells (36% positive) was ob-served 3 to 5 h after seeding, and then the
number ofpositivecellsdecreasedwithincrease
in cellnumber.Only 15%ofthecellswere
posi-tive 25 h after seeding, when the cell number hadalmostdoubled, andonly 8%wereobserved 48 h after seeding, when the cell number had
increased fourfold (data not shown). On the otherhand, although thedoublingtime of 155-4Tcellswassimilartothatof155-4cells(about
25h), they didnotshowapeak of
CSA-positive
cells within 25 h after seeding, although a low but constant level of5 to8%positive cells was
observed
during
thisperiod.
Effect of Act D on expression of HSV-CSA in transformed and tumor cells. Pre-liminary experiments with 155-4 cells showed
thata
high
percentageofCSA-positive
cells wasinduced by treatmentwith 1 to 2
jig
ofAct D per ml for 15 h. To confirmthiseffect,155-4and155-4T cellsweretreated withActD (2,ig/ml)
for 25 h from the time of cell
seeding,
and atintervalsCSA-positivecells and viable cellswere
counted(Fig. 2).Inuntreated155-4cell
cultures,
the percentage of
CSA-positive
cells decreased from 3 hafterseeding;but thepercentage in Act D-treated cell cultures remainedhigh
for20h.50
if-ae
CD
._e
-W
v
E
m
c
0 10 20 0 10
Hours after inoculation
FIG. 2. Relationship between expressionof HSV-CSA and cell growth in transformed (155-4) and tumor (155-4T) cells in thepresenceand absenceof Act D (2tLg/ml). Symbols: 0, absence ofAct D; *,
presenceofAct D.
20 30
VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.265.434.335.600.2]166
Thus, the initial increase ofCSA-positive cells afterseedingwasmaintainedbytreatmentwith Act D. Fluorescence staining in Act D-treated cellcultureswasalso more intense than that in untreated cell cultures (not shown). The viable cells in ActD-treatedcellcultures decreasedto
four-tenths of the cell inoculum (5 x 105 cells perdish) 10h afterseeding. On the otherhand,
treatment with Act D didnotaffectCSA
expres-sion in 155-4T cells for 25 h after seeding,
al-though theproportion of viable cells decreased to the same extent in 155-4 cells treated with ActD.
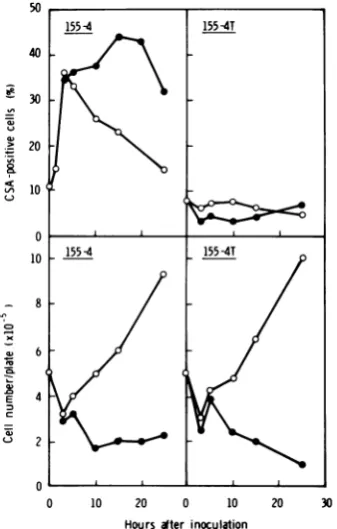
Effective time oftreatment and dose of actinomycinD. Totry to find out the time at which Act D treatmentaffectedCSAexpression,
155-4cells were treatedwithAct D(2
[ig/ml)
for3 to 15 hand then incubated infresh medium without Act D foratotal of 30 h. As shown in
Fig.3, 155-4cell culturestreated with -Act Dfor
5, 10,and15h retainedabout40%CSA-positive cells foratleast20hafter removal ofthe ActD,
but incell cultures treated with Act D for3 h and in control cultures the numbers of CSA-positive cells decreased with cell growth; the number ofpositivecells decreased morerapidly
in control cultures than in those treated with Act Dfor3h.
2
E
c
0 10 20 30 Hours ater removal d drug
FIG. 3. Effect oftimeofAct Dtreatment on
expres-sionofHSV-CSA and cell growth in 155-4 cells. After treatmentwith Act D(2
tog/ml)
for various times (3 to 15 h), cells were washed (O h in this figure) and reincubated infreshmedium without Act D at 37°C.(A) Expression of CSA; (B) cell growth. Symbols:0,
control cells in medium changed3 h after seeding;
0, 3-h treatment with ActD; A, 5-h treatment with ActD;A, 10-h treatment with Act D; O, 15-h treat-mentwith Act D.
Next, to examine the effect of the concentra-tion of Act D onCSA expression in 155-4cells, cellsweretreatedwith various concentrations of Act D(0.5 to 4
tog/ml)
for 10 h(Fig. 4). A linearrelationship wasobserved between the
concen-tration of Act D (0.5 to 2 ,ig/ml) and
CSA-positive cells. Althoughthe viable cell number wasthesameincells treated with 0.5 to4
tog
of Act D perml,addition of 2,ug/mlresulted in the greatestCSA expression;ahigher dose(4,tg/ml)
inhibited itsexpression.
EffectofpuromycinonHSV-CSA expres-sion in transformed cells. To determine
whether new protein synthesis is required for CSAexpressionin 155-4
cells,
PMwasaddedtocultures,andCSA-positivecells, viable cells, and theincorporation of[3H]leucineintocells were
then scored.Theinitialincrease ofCSA-positive cells observed in untreated 155-4 cells was in-hibited only partially by 70
jig
of PM per ml (Fig. 5A), a level which almost completelyin-hibited theincorporation of[3H]leucine into vi-able cells for20h (Fig.
5G).
TheresponseofActD-affected, CSA-positive cells was greatly
in-hibited by the same concentration of PM. In
controls, the number ofviable cellswassimilar
inculturestreated with Act Dalone, PMalone, or ActDandPM (Fig. 5B).
Toconfirm theinhibitoryeffectof PM on Act
D-affected CSA expression, various concentra-tions (1 to 50,ug/ml) of PM wereadded to 155-4 cells treated with Act D for 10 h (Table 3). The Act D effect was inhibited partially by 10 and 20
jig
of PM per mland completely by 50,tg/ml,
but itwas notaffectedwith theaddition of 1,Ag
of PM per ml. Similar results wereobtained withexperimentson theeffectof PM on incorporation of [3H]leucine into the cells.
Theseresultsindicate that newproteinsynthesis
8
2
6
4
2 (
0 0.5 1 2 4
Concentrationodactinomycin D
lpg/mlI)
FIG. 4. Effect of concentration of Act D on HSV-CSAexpression in 155-4 cells. Act D (0.5 to 4 ,ug/ml)-treated and un,ug/ml)-treated cells were incubated for 10 h
at37°C. Points in this figure represent the average
valueoftwo separate tests. Symbols:0,cellnumber;
0,CSA-positivecells.
W
-,
:1
0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.97.219.352.562.2] [image:6.501.303.423.472.596.2]SURFACE ANTIGEN IN HSV-2-TRANSFORMED CELLS 167
20 / °
-4U-A
> 20
-~~
ur
O
CD
-,
-!E
E
:3
1-z
un
1-E
0
10
5
0
30
20
10
0
0 5 10 15
[image:7.501.75.221.59.378.2]Hours after seeding
FIG. 5. Effect ofPMonHSV-CSA expression,cell
growth, and incorporation of['Hileucine in 155-4
cells treated with andwithout Act D.ActD(2 ug/ml)-treated and unug/ml)-treated(control) cellswereincubated with and without PM(70,ug/ml)at37°C for20h. (A)
Expression of CSA; (B)cellgrowth; (C) incorporation
of ['H]leucineinto105viablecells.Symbols:0,
con-trol cells withoutPM; 0, control cells with PM; A,
Act D-treated cells without PM; A, Act D-treated
cells with PM.
is required for Act D-affected CSA expression, but itishardly required for the initialincrease
of CSA in untreated cells.
Effect of antipain on HSV-CSA
expres-sionintransformed cells.Anapproachtothe studyof themechanism(s) ofCSA expressionis
provided by the protease inhibitor antipain,
which prevents A phage induction in
Esche-richiacolibyUV irradiationataconcentration
of 0.5 mM (18).NeithergrowthnorCSA
expres-sion in 155-4 cells was affected by addition of
antipain (0.5 mM) (Fig. 6). However, theboost
in CSAexpressionin thepresenceof Act Dwas
abrogated by0.5 mMantipain (0.05mMhadno
effect and isnotshown);theviablecellnumber
in Act D-treated cultures wasnotdecreasedby
this concentration (0.5 mM) ofantipain. When
antipain (0.5 mM) was added to the cellsafter
removal of Act D, CSA-positive cells also de-creased to 16.3 and 13.2% by 10 and 20 h, re-spectively, after addition of antipain.
DISCUSSION
The specificity of the reaction between anti-HSV-1 serum and HSV-CSA present on the
plasmamembrane of cells transformed by HSV-2 was demonstrated in the present study. The evidence was as follows: (i) HSV-2-transformed cellsand the derivedtumorcellsgave apositive
reactionwithHSV-1antiserum, but otherDNA virus-induced tumor cells did not; and (ii) the
reactivity ofthis antiserum in ts B5-transformed
cells (155-4) was significantly decreased after
absorption withHEFcells infectedwith HSV-2 (wild type or ts B5) or with homologous 155-4
cellsbut not with normal HEF cells or simian virus 40-induced tumor hamster cells. These
findings show thatthe antiserum reactedwith a geneproduct(s), detected asCSA, that was
ex-pressedspecificallyinHSV-2-transformedcells;
however, it isnotknownwhether the CSA
ex-pressed in 155-4 cells is identical to CSA ob-served in theinfected cells.
The present study also demonstrated that
CSA expression was enhanced in transformed
155-4cells3to 5hafter cell seedingat37°C,but not inthe derivedtumor(155-4T)cells,and that
the increase of CSA in 155-4 cells was not an immediate consequenceof trypsinizationbut
re-quired incubation ofthe cells at
37°C
for 3 h. Theexpression ofCSAis not temperaturesen-sitiveinthesecellsat37°C,sinceapeakof CSA-positive cells
(34%
positive) was also found in 155-4cells 10hafter seedingat34°C,
andonlya low level (1 to 7%) of the positive cells was
detected in 155-4 T cells at
34°C
as well as at37°C throughout70h afterseeding (manuscript
in preparation). Furthermore, the finding that
CSA was not enhanced in tumor cells could indicate that either CSA expressionismore
re-pressed in tumor cells or the viral gene(s)
re-sponsible forCSA expression is reduced in the tumor cells. The latter
possibility
is consistent with thereport that the sequencecomplexity
ofHSV-2DNA intumorcellsis lower than that in theoriginal HSV-2-transformedcells
(10).
It iswellknownthatAct DbindstoDNA and inhibits DNA, RNA, andprotein syntheses (3,
23). Thisdrugisalso
tumorigenic (27)
andmu-tagenic (7, 11) in animals. The present
study
demonstratedthatincreased CSA
expression
in transformed cells after cellseeding
could be maintained bytreatmentwithActD,
and that new proteinsynthesis
wasrequired
for CSAexpression affected by Act D but not for the initial increase of CSA
expression
in the absenceVOL. 29,1979
An
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 3. Effectof PMonHSV-CSA in 155-4 cells treated with Act D
Incorporationof
Treatment (10 h) CSA-positivecells(%)b Viablecells/plate(x 104) [BH]leucine (cpm/10' viablecells)
Untreated 23.5 50.0 8,038
Act D 42.4 13.5 6,779
PM (50,ug/ml) 16.7 5.9 761
Act D+PM(1,ug/nl) 43.9(1.04)c 8.0(0.59)d 4,786(0.71)e
ActD+PM(10yg/ml) 10.7(0.25) 6.8(0.50) 1,179(0.18)
Act D+PM(20,g/ml) 10.1 (0.24) 6.4(0.47) 983(0.15)
Act D+PM
(50,ug/ml)
2.0(0.05) 5.6(0.41) 478(0.07)a 155-4
cells
weretreatedsimultaneouslywith Act D (2iog/ml)
and various concentrations of PM and thenincubated for10hat37°C.
b Atotal of
103 cells
werecounted.'Values in parentheses represent the ratios ofCSA-positivecells in cultures treated with ActDand PMto
those incultures treated with Act Donly.
dValues in parentheses represent the ratios of cell numbers in cultures treated withAct Dand PMtothose
incultures treated with ActDonly.
eValues inparentheses represent the ratios ofincorporationof
[3H]leucine
intocells
treated with Act D andPMtothat intocells treated withActDonly.
50
30
20
10
0 B
12
8
C4
u~~~~
0
0 5 10 15 20 25
Hours
FIG. 6. Effect of antipainonHSV-CSAexpression in 155-4 cells. Act D(2,ug/ml)-treatedanduntreated (control) cells incubated for 5 h at 37°C with or
withoutantipain (0.5 mM) werewashed(0h in this
figure) and reincubated in fresh medium with or
without antipain (0.5 mM). (A)Expression of CSA; (B) cell growth. Symbols: 0, control cells without
antipain; 0, control cellswith antipain; A,Act
D-treated cells withoutantipain;A,ActD-treated cells
withantipain.
of Act D.Thiseffectof Act DonCSA expression
continued for 20 h after removal of the drug,
once cellshad been treated with2
,ug/ml
for5h. However,Act Ddid notaffect CSAexpression inthederivedtumorcells. Perhaps inananalo-gous way, Epstein-Barr virus-associated
mem-braneantigenin atested Burkitt
lymphoma
celllineshowednoresponse to anymetabolic inhib-itor(includingActD)or toanychangeinculture conditions (32).
Theproteaseinhibitorantipain isarelatively nontoxic (29),
low-molecular-weight
compound,
which inhibitstrypsin- andpapain-likeproteases in particular (26). It is very interesting that
antipain inhibitedActD-enhancedCSA expres-sionwithoutcausingsignificant cell damage but didnotinhibit theinitial increase ofCSA
expres-sionin theabsence ofAct D.Roberts and Rob-erts (24) have shown thatAprophage inE. coli is induced by proteolytic cleavage of the A re-pressor. Recently, Meynetal. (18) have shown thatantipain inhibitsAphage induction (oneof the SOS functions) in E. coli by blocking
pro-teolytic inactivation of therepressor.Therefore,
itisconceivablethat theputative repressor(s) of the CSA
function(s)
is inactivated by apro-tease(s) inducedoractivatedbyActD, leading
to the appearance of CSA. Another possible explanation ofthe effect ofAct D would be an
inhibition ofa repressor at the transcriptional level. Of course, these possibilities are highly
speculativeatpresent.
In any case, our findings indicate that two kinds of CSA expression occur in transformed
155-4
cells.
One type is expressed soon afterseeding, without the need for further protein
synthesis, and is insensitive to antipain. The other isexpressed in response to Act D, requires
protein synthesis, and is sensitive to antipain. Protease(s), therefore, seems to play an impor-tant role in the enhanced level of expression.
Sinceit seems that cellline 155-4 respondsin a
heterogeneousway with respect toCSA
expres-168
KIMURA ET AL.on November 10, 2019 by guest
http://jvi.asm.org/
SURFACE ANTIGEN IN HSV-2-TRANSFORMED 169
sion, clonal analysis of cells from the line isnow
inprogress.
ACKNOWLEDGMENTS
WearegratefultoDavid Schlessinger for critical reading of themanuscript.
LITEIATURE CITED
1. Boyd, A. L., and T. W. Orme.1975.Transformationof
mousecellsafter infection withultraviolet
irradiation-inactivatedherpessimplexvirustype2.Int. J. Cancer 16:526-538.
2. Collard, W., H. Thorton, and M. Green. 1973. Cells transformed by human herpesvirustype 2 transcribe virus-specific RNA sequences shared by herpesvirus types1and2.Nature(London)New Biol. 243:264-266. 3.Cooper,H. L., andR. Braverman. 1977. The mecha-nismby which actinomycinDinhibitsproteinsynthesis inanimal cells. Nature(London)269:527-529. 4. Davis,D.B., and D. T.Kingsbury,1976.Quantitation
of theviralDNA present incells transformedby UV-irradiated herpes simplexvirus.J. Virol. 17:788-793. 5. Duff, R., and F. Rapp. 1971. Properties of hamster
embryo fibroblasts transformedin vitroafterexposure
toultraviolet-irradiatedherpes simplexvirustype2.J. Virol.8:469-477.
6. Duff, R.,and F.Rapp.1973.Oncogenictransformation of hamsterembryocellsafterexposure toinactivated herpes simplex virustype 1.J. Virol. 12:209-217. 7. Epstein, S., E. Arnold,J. Andrea,W.Bass,and Y.
Bishop.1972.Detectionofchemicalmutagensbythe dominant lethal assay in the mouse. Toxicol. Appl. Pharmacol. 23:288-325.
8. Esparza, J.,D. J.M.Purifoy,P.A.Schaffer,and M.
Benyesh-Melnick. 1974. Isolation, complementation andpreliminary phenotypiccharacterizationof temper-ature-sensitivemutantsofherpessimplexvirustype2. Virology57:554-565.
9. Flannery,V.L.,R. J.Courtney,andP. A. Schaffer.
1977.Expressionofanearly,nonstructural antigenof herpes simplex virus incells transformed in vitro by herpes simplexvirus. J. Virol.21:284-291.
10.Frenkel, N.,H.Locker,B.Cox,B.Roizman,and F.
Rapp.1976.Herpes simplexvirus DNAintransformed
cells:sequencecomplexityin fivehamstercelllinesand
onederived hamstertumor.J.Virol. 18:885-893. 11.Haruta, M. 1968.Teratogenic effects ofactinomycin-D
onddOmouseembryos.Acta Pathol.Jpn.18:267-286.
12. Kimura, S.,J.Esparza,M.Benyesh-Melnick,and P.
A. Schaffer. 1974. Enhancedreplicationof tempera-ture-sensitive mutantsofherpes simplexvirustype2
(HSV-2) at the nonpermissive temperature in cells
transformedbyHSV-2.Intervirology3:162-169. 13. Kimura, S.,V.L.Flannery,B.Levy,and P. A.
Schaf-fer. 1975.Oncogenictransformationofprimaryhamster
cells by herpes simplexvirustype2 (HSV-2) andan
HSV-2temperature-sensitivemutant.Int. J. Cancer15:
786-798.
14. Kucera,L.S.,and J. P. Gusdon.1976.Transformation of humanembryonicfibroblastsbyphotodynamically inactivatedherpes simplexvirustype2 atsupraoptimal
temperature.J. Gen.Virol. 30:257-261.
15. Kutinova L., V. Vonka, and J. Broucek. 1973.
In-creasedoncogenicitv and synthesis of herpesvirus
anti-gensinhamstercellsexposedtoherpes simplextype2
virus. J.Natl.Cancer Inst. 50:759-766.
16. Macnab,J. C. M. 1974. Transformationofrat embryo
cells bytemperaturesensitivemutantsof herpes
sim-plex virus.J. Gen. Virol. 24:143-153.
17. Macnab, J. C. M., andM. C. Timbury.1976. Comple-mentationofts mutantsbyaherpes simplexvirus
ts-transformed cell line. Nature(London) 262:233-235. 18. Meyn, M. S., T. Rossman,and W. Troll.1977. A
pro-tease inhibitor blocks SOS functions in Escherichia coli:antipain preventsArepressorinactivation,
ultra-violetmutagenesis, andfilamentous growth.Proc. Natl.
Acad. Sci.U.S.A.74:1152-1156.
19.Minson, A. C., M. E. Thouless, R. P. Eglin, and G. Darby.1976.The detection of virus DNAsequencesin
aherpestype2transformed hamster cell line(333-8-9). Int.J. Cancer17:493-500.
20. Munyon, W., E. Kraiselburd,D. Davis, and J. Mann. 1971.Transfer ofthymidine kinasetothymidine kinase-less L cells by infection with ultraviolet-irradiated herpessimplex virus. J. Virol.7:813-820.
21. Rapp,F.,and R. Duff.1972.Invitrocelltransformation byherpesviruses. Fed. Proc.31:1660-1668.
22. Reed, C. L., G.H.Cohen, and F.Rapp. 1975. Detection ofa virus-specific antigen on the surface of herpes simplex virus-transformed cells. J. Virol. 15:668-670. 23. Reich, E., A. Cerami, and D.Ward. 1967. Actinomycin,
p.714-725. InD.Gottlieb andP.Shaw (ed.), Antibiot-ics, vol.1.Springer-Verlag,NewYork.
24. Roberts, J. W., and C. W. Roberts. 1975.Proteolytic cleavage of bacteriophage Lamdarepressor in
induc-tion. Proc.Natl. Acad. Sci. U.S.A.72:147-151. 25. Schaffer,P.A., G. M. Aron, N. Biswal, and M.
Ben-yesh-Melnick.1973.Temperature-senstivemutantsof herpes simplex virustype1:isolation, complementation andpartial characterization. Virology 52:57-71. 26. Suda, H.,T.Aoyagi,M. Hamada, T. Takeuchi, and
H.Umezawa. 1972.Antipain,a newproteaseinhibitor isolated from actinomycetes. J. Antibiot. 25:263-266. 27. Svoboda, D., J. Reddy, and C.Harris. 1970. Invasive
tumorsinduced inratswithactinomycinD.Cancer Res. 30:2271-2279.
28. Takahashi, M.,and K. Yamanishi. 1974. Transforma-tion of hamsterembryo and humanembryo cellsby temperaturesensitivemutantsofherpes simplexvirus
type2.Virology61:306-311.
29. Umezawa,H. 1972.Enzymeinhibitors of microbial ori-gin,p.29-32.UniversityofTokyoPress, Tokyo. 30. Vestergaad,B.F.,and P. C.Grauballe.1977.Crossed
immunoelectrophoreticidentification ofpartially puri-fied typecommonandtypespecific herpes simplexvirus
glycoprotein antigens.Proc. Soc.Exp.Biol. Med. 156: 349-353.
31. Williams,C.A.,and M. W. Chase(ed.).1968.Appendix II.Buffer.Methods Immunol. Immunochem. 2:388. 32. Yata, J.,G.Klein,J.Hewetson,andL.Gergely.1970.
Effect ofmetabolic inhibitorsonmembrane
immunoflu-orescencereactivityof established Burkittlymphoma celllines.Int. J. Cancer 5:394-403.

![FIG. 5.growth,ActExpressiontreatedwithcellsofcellstrol ['H]leucine Effect ofPM on HSV-CSA expression, cell and incorporation of ['Hileucine in 155-4 treated with and without Act D](https://thumb-us.123doks.com/thumbv2/123dok_us/1520125.104609/7.501.75.221.59.378/growth-actexpressiontreatedwithcellsofcellstrol-leucine-effect-expression-incorporation-hileucine-treated.webp)