JOURNAL OF VIROLOGY,May 1981,p.420-429 0022-538X/81/050420-10$02.00/0
Mechanisms of
Expression
of
Herpes
Simplex Virus-Common
Surface
Antigens in
Clonal Cells of
a
Herpes Simplex Virus
Type
2-Transformed Line
KATSUICHIROOKAZAKI,' YOSHINARIOHNISHI,' NAGAYUKI YOSHIDA,2ANDSUSUMU
KIMURA3*
Department of Bacteriology, School of Medicine, University of
Tokushima,'
andDepartmentof Microbiology, School of Pharmacy, Tokushima University ofArts and Science,2 Tokushima 770, andDepartmentofMicrobiology, Kochi MedicalSchool, Oko, Nangoku, Kochi 781-51,3Japan
Received24September 1980/Accepted 19 January 1981
Rabbit antiserum hyperimmune to herpes simplex virus type 1 was used to
studytheexpression of herpessimplex virus type-commonsurfaceantigens(CSA)
byindirectimmunofluorescencetestsinthreerepresentativecell clonesisolated
from a herpes simplex virus type 2-transformed hamster line, 155-4. These three clones showed differentphenotypeswith respect to CSAexpression: (i) a CSA-positivetype (clone 155-4-213), in which the antigensincreased soon (5 h) after seedingat370C,but notaftertreatmentwithactinomycinD; (ii)aCSA-inducible type (clone 155-4-03), in which the antigens increased after tretment with acti-nomycin D (2,ug/ml) for20h, butnotafter seeding only;and(iii)aCSA-negative type (clone 155-4-16),in whichthe antigensdid not increase after seeding orafter actinomycin Dtreatment.CSA expressionintheCSA-positivetypewasinhibited by 2-deoxy-D-glucose, but not by puromycin, suggesting that the expression required glycosylation, but notactive proteinsynthesis. CSA expressionin this
type was insensitive to the protease inhibitors antipain and
p-nitrophenyl-p'-guanidinobenzoate. On theotherhand, actinomycinD-induced CSA expression in the CSA-inducible type wasinhibited by both 2-deoxy-D-glucose and puro-mycin, suggesting thatthe induced expression required both glycosylation and protein synthesis. CSAexpression inducedin this type wassensitive to the two protease inhibitorsatconcentrations havinglittleeffect onoverallcellular metab-olismor cell viability.These resultsindicate thatCSA expressionsin the CSA-positivetypeand theCSA-inducibletype areenhnaced bydifferent mechanisms.
Various mammalian cells have been
trans-formedoncogenicallyinvitrobyherpessimplex virustypes 1 and2 (HSV-1 and HSV-2, respec-tively) (2, 12, 13, 25, 27, 29, 44). Viral DNA
sequences (11, 15, 33), HSV-specific RNA (7),
andHSV-specific antigens(2,12,25,27,44) have beendemonstratedincells transformed by
HSV-2.Moreover, viralgenetic information has been
concluded tobe present inHSV-2-transformed
cells from reports that complementation
be-tween temperature-sensitivemutantsof HSV-2
andresident HSV genes intransformedcells has
been demonstrated at the nonpermissive
tem-perature (24, 30).
SeveralHSV type-specificand type-common
antigens havebeenidentifiedrecently in
HSV-infectedcellsby immunological analysis (16, 34,
46), and type-common antigens have also been
detectedonthesurface ofcells transformed by
HSV (4, 18, 38, 39).
In an attempt to identify the HSV-specific
geneproducts
expressed
inHSV-2-transformed hamstercells,
wedemonstratedthatHSVtype-commonsurfaceantigens
(CSA)
canbedetectedspecifically
in transformed and derived tumorcelllinesby immunofluorescencetestsand that
HSVCSAexpressioncanbeenhancedin
trans-formed cellline 155-4 inatleasttwoways: one
way is after seeding at 37°C, without active
protein synthesis, and the other way is in
re-sponseto actinomycin D (ActD) and involves
protein synthesis (26).We have shownrecently
byclonal analysisof the cellsthat the cellline
consists of at least three different phenotypes
with respect to the expression of HSV CSA,
namely, aCSA-positive type, a CSA-inducible
type, and a CSA-negativetype. This paper
de-scribes a study of the mechanisms of CSA
expressionintwodifferentcelltypes
(CSA-pos-itive type andCSA-inducibletype).
420
on November 10, 2019 by guest
http://jvi.asm.org/
MATERIALS AND METHODS Cell lines. Transformed cell line 155-4 (passages 165 to 185) was obtained byaroutine methodafter theinfection of hamster (strainLSH) embryo fibro-blast (HEF) cellswith the DNA-negative tempera-ture-sensitive mutant (tsB5) of HSV-2 strain 186 at the nonpermissive temperature (38°C), as reported previously by Kimura et al. (25). The cell line was passagedseriallyat37°Cbecause it was not stable at 38°C. Clones from cell line 155-4 were isolated as follows. 155-4cells (approximately 30 cells per dish), which hadbeen subcultured for 180 passages in vitro, wereseeded into60-mmpetri dishes (Falcon Plastics, Oxnard, Calif.) and incubated in Eagle minimum es-sentialmedium (MEM) containing 20% fetal calf
se-rum (FCS) at 37°C in a CO2 (5%) incubator. At 10
days afterseeding, when there were4 to8cell colonies per dish, a total of30colonies were collected from several dishes, using a steel cylinder and
typsiniza-tion. Cellcloneswere passaged in Eagle MEM con-taining 10% FCSat37°C. Allclones were used between passages 9 and 24. All cells used in this study were grown in Eagle MEM supplemented with 10% FCS and 0.075%NaHCO3 for cultures in closed vessels and 0.225% NaHCO3 for cultures in open vessels.
Antisera. Rabbit antiserumhyperimmuneto HSV-1 (strain KOS) was prepared as reported previously (26). Theneutralizingantibody titer to HSV-1 of this anti-HSV-1 serum was 1:1,280 in 50% plaque reduction tests.Fluorescein-conjugatedgoatantiserum to rabbit
7Simmunoglobulin (FGAR)waspurchased from
Hy-landLaboratories, Inc., Costa Mesa,Calif.Nonspecific reactivity was removed from the two antisera by the absorption technique described previously (26). In brief,4ml ofundiluted rabbit anti-HSV-1 serum was absorbed with 1 gof rabbitliver powder, 108human embryoniclung cells,and10'HEFcells. FGAR (4 ml) wasalso absorbed with1gofrabbit liver powder and 108 HEFcells. The absorbed anti-HSV-1 serum used
in thisstudyreacted specificallywithHSV CSAon
HSV-2-transformedandHSV-2-infectedcells,butnot
withtumorigenictransformedcellsinduced with sim-ian virus 40 or bovine adenovirus type 3 orcontrol HEFcells(26).
Detection of HSV CSA The procedure of the indirectimmunofluorescence testfor thedetection of HSV CSA has been describedindetailbyKimuraet al. (26). The cells weregrown andpassagedat least three timesbeforetests.Cells (5x
105)
weretreated withAct D(0.5to 4ug/ml)atthe time ofseeding with 5 ml of medium into60-mm petri dishes. Inhibitors werealsoaddedtoActD-treatedcellsatthe time of seeding.Atthe indicatedtime, cellsweredispersedby treatmentwith 0.25% trypsin for 1 min and washed threetimes with barbital buffer(pH 7.4)containing3X10-3 Mbarbital, 1.8x10-3Msodiumbarbital,0.85% NaCl,5x 10-4MMg2+, 1.5x 10-4MCa2,and0.1% gelatin(GBB).Samplesof1dropofabsorbed antise-rum at a final dilution of 1:4 were added to cell suspensions (6x105to 10x105cells).After30minof incubationat37°C,cellswerewashedthreetimeswith GBB and then incubated with 1 drop of absorbed FGARdiluted1:8for30minat37°C.Finally,the cells werewashed threetimes withGBB and mountedina 50%oglycerol solution, and 103cells persamplewere
observed with a reflected-light fluorescence micro-scope(Olympus OpticalCo.Ltd., Tokyo,Japan). The number of viable cellswasdeterminedby trypanblue exclusion.
For testsontheeffects ofvarious conditions on the
expressionofHSV CSAinclone 155-4-213 (see Table
1), cells detached by pipetting were suspended in 1 ml ofphosphate-buffered saline withoutCa2" and
Mg2e
[PBS(-)]and treatedwith1ml of trypsin(0.5%) or 1 ml of PBS(-) for 1 to 3 min. Trypsinization was
stopped by the addition of 15 ml of Eagle MEM
containing 10% FCS and centrifugation. Some cell
suspensions with or without trypsinization were
im-mediately assayed for indirect immunofluorescence.
Other suspensions(1x105 to25 x105cells per sample) were incubated with 5 ml of Eagle MEM containing 10%FCS in 60-mm dishes as monolayers or in tubes (15 by 150mm) assuspensions for5h at 4 or37°C andthen assayed for indirectimmunofluorescence.
Inhibitors. Act D andpuromycindihydrochloride (PM) were obtained from Boehringer Mannheim,
Mannheim, West Germany, and Marker Chemicals Ltd., Jerusalem, Israel, respectively. The protease in-hibitor antipain
[(l-carboxy-2-phenyl-ethyl)carbam-oyl-L-arginyl-L-valyl-argininal] (42) was kindly pro-videdby T. Uchida, Osaka University, Osaka,Japan,
andp-nitrophenyl-p'-guanidinobenzoate
dihydrochlo-ride(NPGB)wasobtained from ICNPharmaceuticals, Life SciencesGroup Inc., Cleveland, Ohio. 2-Deoxy-D-glucose was purchased from Sigma ChemicalCo., St. Louis, Mo. Stock solutions concentrated 10 times wereprepared, passedthrough 450-nm membrane fil-ters (Millipore Corp.,Bedford, Mass.),andstored at -200C.
Incorporations of[3Hlthymidine, [3H]uridine,
and[3H]leucine.The radioactiveprecursors
[methyl-3H]thymidine (40to60Ci/mmol),[5-3H]uridine (>25 Ci/mmol), and L-[4,5-3H]leucine (40to60Ci/mmol) wereobtainedfrom NewEngland NuclearCorp., Bos-ton, Mass. For the examination of DNA and RNA syntheses, cultureswerelabeled with either [3H]thy-midine (0.5,Ci/ml) or[3H]uridine (0.1 XCi/ml) con-taining 0.1 ,ug of thymidine per mlor0.1
,Ag
of uridine per ml, respectively, for 8 h afterseeding. For the examination ofproteinsynthesis, cultureswerelabeled with[3H]leucine (1.6t,Ci/ml) for20h afterseeding. The cellswerethen washedoncewith cold(4°C) Tris buffer, harvested, andcentrifuged at 800 xg for 10 min.Pelletssuspendedin1ml ofcold Tris bufferwere sonicated inanice bathat10 kc for 20sand treated with1ml of 20% trichloroacetic acid for20minat0°C. The[3H]leucine-labeledsampleswerethenboiled for 15 min. The suspensionswere applied to Whatman GF/C filterdisks,usinganaspirator. The diskswere washed three times withcold 5% trichloroacetic acid and once with ethyl alcohol, dried, and placed in scintillation vials. A 10-mlamountofscintillation fluid was then added to each vial, andradioactivity was measured inaliquidscintillation spectrometer(model3385;Packard InstrumentCo., Inc.,Rockville,Md.).
RESULTS
Expressionof HSV CSA in variousclones
of transformed line 155-4. We isolated three
on November 10, 2019 by guest
http://jvi.asm.org/
422 OKAZAKI ET AL.
representative clones, 155-4-213, 155-4-03, and
155-4-16, from cell line 155-4 transforned by
HSV-2 (tsB5 of strain186). These clones showed
different phenotypes with respect to the
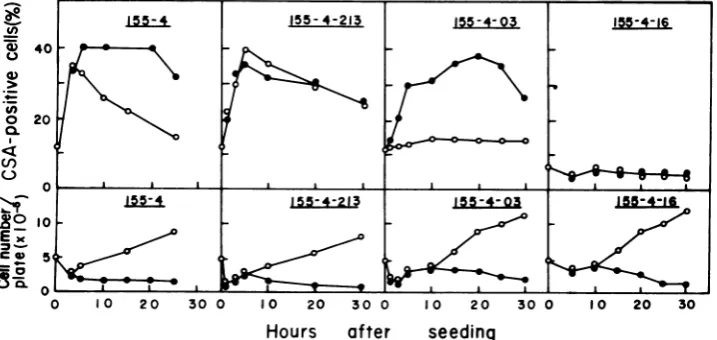
expres-sion ofHSV CSA (Fig. 1). In clone 155-4-213
(CSA-positive type), CSA expression was
en-hanced (about 40% positive) soon (5 h) after
seeding at 37°C; thereafter, the expression de-creased with an increase in the cell number, returningto theoriginallevel (about 12% posi-tive) 48hafterseeding.Act D(2
,ug/ml)
didnotaffect this CSA expression. In clone 155-4-03 (CSA-inducible type), CSA expressionwas low
(11 to 15%positive) for30hafterseeding.
How-ever, theexpression oftheseantigensgradually
increased after treatment with Act D(2
I.g/ml),
reachinga maximum (35 to 40% positive) 20h
after treatment. Inclone155-4-16(CSA-negative
type),CSA expression didnotincrease within 30
hafter seeding or after Act D treatment. The number of viable cells decreased to a similar
extentin these three cell clonesupon treatment
with Act D.
Conditions
affecting expression
of HSV CSA in theCSA-positive
type. The condi-tions inducing CSA expression in clone 155-4-213wereexamined byvarying thetimeof tryp-sinization, the size of the cell inoculum, theincubationconditions, andthemedia(Table1).
No increase inCSA was observed in 155-4-213
cells thatwere immediately assayed forthe de-tection of CSA without trypsinization or after trypsinization for 1 or 3min or that were
incu-bated at 4°C for 5 h after trypsinization for 1
min. A slight increase in CSA was observed
when: (i) cells were incubated at 37°C for 5 h without trypsinization; (ii) smaller inocula
(105
'd
155-4 155-4-21*~40
0 20 U)
o 20
0.
;W°O IS15-4 155-4-21
EX)
9
0loft fc 5
cells) or larger inocula (2.5 x
106
cells) wereused; (iii) cells were incubated at 37°C for5 h
aftertrypsinization for3min;and(iv) cellswere
incubated with PBS(-), MEM without serum,
or MEMcontaining a low concentration (5%) of
serum. The increase in CSA in 155-4-213 cells
wasmarked only when the cells were trypsinized
for 1min and incubated at an appropriate
inoc-ulum (5 x 105 cells per dish ortube) for 5 h at
37°C with MEM containing 10 or 20% serum. Anincrease inCSAwasobservedwithcell sus-pensions andeven morewithmonolayers.Thus,
incubation of cells at37°C after trypsinization
was necessaryfor an increase in CSA in clone
155-4-213, andtrypsinization itself was not
suf-ficient.
Conditions inducing expression of HSV CSA in theCSA-inducible type. The effectof
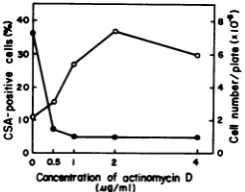
the concentrationofAct D onthe expression of
CSAinclone155-4-03 wasexaminedby treating the cells with various concentrations of Act D
(0.5 to 4
tig/ml)
for 20 h(Fig. 2). A highcorre-lationwas observed between the concentration
of Act D (0.5 to 2 ,ig/ml) and the number of
CSA-positive cells. Although the viable cell numbers were almost the same in cultures
treated with 0.5 to 4
,tg
of Act D per ml, theadditionof 2
,ig
of Act Dresultedinthe greatest CSA expression, whereas a higher dose (4 ,ig/ ml) slightly inhibited its expression. Therefore,in theexperiments describedbelow,2,ug of Act
D per ml was used for the induction of CSA
expressionin 155-4-03 cells.
For thedeterminationof theeffective time of
additionof Act D on the induction ofCSA, 155-4-03cellsweretreated with Act D (2,Lg/ml) at various times (0, 3, 5, and 10 h) after seeding
0 10 20 30 0 10 20 30 0 10 20 30 0
Hours
after
seeding
FIG. 1. Relationshipbetween the expression of HSV CSA and cell growth in HSV-2-transformed cell line 155-4andthree clonesfrom the line at 37°C in the presence and absence of Act D (2,ug/ml). Symbols: 0, absenceofActD;0,presenceof Act D.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.500.76.435.459.629.2]TABLE 1. Various conditionsaffecting the expression of HSV CSA in clone 155-4-213
Cell inoculumper Incubation conditions SA-positive
Exptno.andtreatment sample(X105) Sa l (h) Temp(°C) cells (%)b
1. Trypsinizationc
None 5 Noincubation 9.5
5 Monolayer 5 37 16.7
1min 5 Noincubation 6.8
5 Suspension 5 4 10.8
5 Suspension 5 37 32.4
1 Monolayer 5 37 22.3
5 Monolayer 5 37 38.3
25 Monolayer 5 37 13.8
3min 5 Noincubation 7.0
5 Monolayer 5 37 21.0
2. Medium andserumd
PBS(-) without serum 5 Monolayer 5 37 15.4
MEM without serum 5 Monolayer 5 37 20.8
MEM with 5%serum 5 Monolayer 5 37 24.8
MEM with 10%serum 5 Monolayer 5 37 38.2
MEM with20% serum 5 Monolayer 5 37 34.0
a
Celis
wereinoculated into dishes (monolayer) or tubes (suspension) with gentle shaking. No incubation,Cellsuspensionswereimmediately assayed for indirectimmunofluorescence.
'Atotal of103
cells
werecounted.Trypsinizationwasperformedasdescribed in the text.
d
Cells
weretreatedwith trypsin (0.25%) for1min andsuspendedin5mlofPBS(-) or Eagle MEM containing various concentrations of FCS.3o o
E
.~0
o o5 2 4 Cancmnitonof oinonycinD
[image:4.500.59.450.88.313.2](Mg/mi)
FIG. 2. Effect ofthe concentration ofAct D on
HSV CSAexpression in clone 155-4-03. Act D(0.5to
4pg/ml)-treatedanduntreated cellswereincubated for20hat37°C. Symbols: 0, CSA-positive cells;0,
cell number.
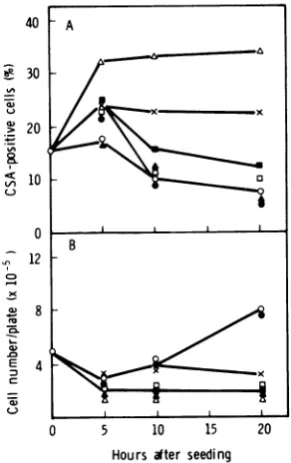
(Fig. 3). When ActD wasaddedtothe cellsat
thetime ofseeding,HSVCSAincreasedgreatly by20 h afterseeding.The addition of Act Dat
3 h after seeding induced CSA afteralong lag
(about 17h). NoincreaseinCSAwasobserved
uponthe addition of thedrugat5or 10h after
seeding. Thesefindings indicate that the
addi-tion of Act D within 3 h after seeding was
necessaryforthe induction ofCSAin clone
155-4-03.
Forthe determination of the effective period
of treatment with Act D, 155-4-03 cells were
treated with Act D (2 ag/rnl) for3, 5, and 10 h
fromthe time of seeding and then incubated in
freshmedium without Act D foratotal of 30h.
In cells treated with Act D for 5 and 10 h,CSA
expression wasinduced and a high percentage
(35to40%) ofCSA-positivecellspersistedfor 20
h after the removal of Act D. However,in cells
treated with Act D for 3h, thenumber of
CSA-positive cells did not increase greatly (data not shown).
Effect of PM on the expression ofHSV CSA in the CSA-positivetypeand CSA-in-ducible type.To determine whether new
pro-tein synthesis is required for CSAexpression in
clones 155-4-213 and 155-4-03, we added PM to
cultures with or without Act D (2 ,ug/ml) and
determined the numbers and percentages of
CSA-positive cells and viable cells and the in-corporation of [3H]leucine into the cells. As
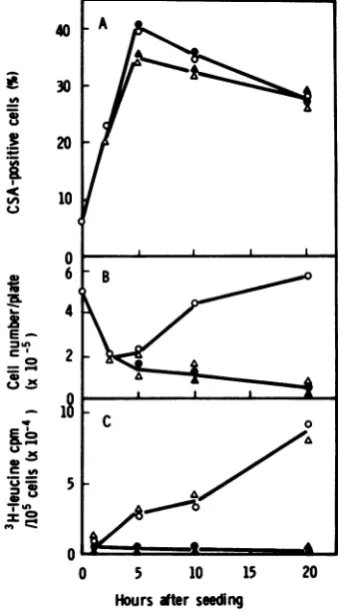
shown in Fig.4A,CSAexpression inclone
155-4-213 with or without Act D was notsuppressed
by PM (70,ug/ml),whereas theincorporationof
[3H]leucine into viable cells was almost com-pletely inhibited by this concentration of PM (Fig. 4G).
On the other hand, the induction of CSAby
Act D in clone 155-4-03 was suppressedto the
level of that in untreated cultures by 70 tg of
PM per ml (Fig. 5A), a concentration which
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.500.90.213.379.475.2]40
- 30
2
i
204A 10
0
15
12
9
6
1
3
0
10 15 20
[image:5.500.58.252.61.339.2]Hours ftersuing
FIG. 3. Effect ofthe timeofadditionofActDafter
cellseedingontheexpression ofHSVCSAinclone 155-4-03. At the indicated times (0, 3, 5, and 10h) after seeding,cellsweretreated with ActD(2 pg/ml) and reincubatedat37°C. (A) Expression of CSA; (B)
cellgrowth. Symbols:0,control(untreated) cells;0,
additionofAct Dat0 hafter seeding; A,additionof Act Dat3 hafter seeding;A,additionofAct Dat5
h after seeding; O, addition ofAct D at 10h after seeding.
greatly inhibited the incorporation of
['H]leu-cine into the cells (Fig. 5G). CSA expressionin
this clone in the absence of Act D was not
suppressed by the same concentration of PM.
Thenumbers of viable cells in clones 155-4-213
and 155-4-03 were similar in cultures treated
with Act Dalone, PM alone, and Act D plus PM
(Fig. 4B and 5B). These results indicate that
new protein synthesis is required for Act
D-inducedCSA expression inclone155-4-03,but is
notrequired forCSA expression in clone
155-4-213afterseeding.
Effect of 2-deoxy-D-glucose on the
ex-pression of HSV CSA in the CSA-positive
type and CSA-inducible type. We next
ex-amined whether glycosylation is necessaryfor
CSA expression in clones155-4-213and155-4-03
by treating them with various concentrations (1,
5, 10, and 20 mM) of2-deoxy-D-glucose, which
inhibits glycosylation of viral glycoproteins (9,
10,22). CSA expression in clone 155-4-213after
seedingwasinhibited inadose-dependent
man-nerby2-deoxy-D-glucose (Fig. 6A).The number
of viable cells in this clone also decreasedslightly
upon treatmentwith thedrug.
CSA inductionbyAct D in clone 155-4-03was
inhibited greatly bythe addition of
2-deoxy-D-glucose (more than 5 mM), although CSA
expression in the clone without Act D was not
affected by 2-deoxy-D-glucose (Fig. 6B). CSA
inductionbyAct D in 155-4-03 cells was more
sensitive to 2-deoxy-D-glucose than was CSA
expression in 155-4-213 cells afterseeding. The
number of viable cells in clone 155-4-03 de-creasedgreatlyathigherconcentrations(10and 20mM) of2-deoxy-D-glucose in the absence of
ActD,but not in the presence of Act D. These
findings suggest thatglycosylationisnecessary forCSAexpressionboth in 155-4-213 cells after
e 30
i
20I^ 10
u
E^
=
-C0
_ _
u
C
0 5 10 15 20
Hoursafterseeding
FIG. 4. Effect ofPMonHSVCSAexpression,cell
growth, and incorporation of [3H]leucine in clone 155-4-213 incubated with andwithout Act D. Act D (2pg/ml)-treatedand untreated(control)cellswere
incubatedwith andwithout PM(70ug/ml) at37°C for20 h.(A) Expression of CSA; (B)cellgrowth; (C)
incorporation of [3H]leucine into 105 viable cells. Symbols:0,control cellswithoutPM; *, control cells
withPM; A,ActD-treated cellswithoutPM;A,Act
D-treated cellswith PM.
C
P-E
.5
U)1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.500.274.445.262.569.2]40
:-,
-.Z
'a
=
In
30
20
10
0
8
4
0
16
8
0
0 5 10 15 20
Hours after seeding
FIG. 5. Effect ofPMonHSV CSAexpression,cell growth, and incorporation of[3Hileucine in clone 155-4-03 incubated with andwithout Act D. Act D(2
Pg/ml)-treatedanduntreated(control)cellswere
in-cubated with andwithout PM(701tg/ml)at37°Cfor 20 h. (A) Expression of CSA; (B) cellgrowth; (C) incorporation of[3Hileucine into 105 viable cells. Symbols:0,control cells withoutPM; *,control cells
withPM; A,ActD-treatedcells withoutPM;A,Act
D-treated cells with PM.
seeding and in 155-4-03 cells after treatment
withAct D.
Effects of protease inhibitors on the
expression of HSV CSAintheCSA-positive
type and CSA-inducible type. Our previous
study (26) demonstrated that theprotease
inhib-itorantipain (0.5 mM) inhibited Act D-enhanced
CSA expression in the parent cell line 155-4
withoutcausing significantcelldamagebut that
it didnotinhibitanincreaseinCSA expression
in the absence of Act D in thesame cellline. In
this study, the effects of antipain and another
proteaseinhibitor, NPGB,onCSA expressionin
clones 155-4-213 and 155-4-03 were examined.
As shown in Fig. 7, CSA expression and the
number ofviablecellsin clone 155-4-213 in the
presence and absence of Act D were hardly
affected by antipain (0.5 or 1.0 mM).
Further-more, CSA expression in this clone 5 h after
seedingwas not affected by NPGB, even at a high concentration(0.1mM;seeFig. 9A).
On the other hand, Act D-induced CSA
expression in clone 155-4-03 was inhibited in a dose-dependentmannerby antipain(Fig. 8) and NPGB (Fig. 9B) without significant cell damage. Higher doses of antipain (0.5 and 1.0mM) and NPGB (0.1 mM) suppressed theinduced expres-sion to the level of the control culture (not treated with Act D), but these high doses of antipain and NPGB didnotinhibit CSA expres-sion in this clone in the absence ofAct D. To determine whether the doses of antipain and NPGB used in this study affect DNA,RNA, or proteinsynthesis in155-4-03cells,wemeasured the effects of thesecompounds onthe incorpo-rations of [3H]thymidine,
[3H]uridine,
and[3H]leucine
intomacromolecules in thecellsinthe presence and
abs9nce
of Act D (Table 2).Higher doses of antipain (0.5 and1.0 mM) and NPGB (0.05 and 0.1 mM) did not inhibit the incorporation of any precursors into 155-4-03 cellswithorwithoutActDtreatment, indicating that theseproteaseinhibitors,at concentrations
40
130
120
uo
40
o
(0
20
I0
0
o1 5 10 20
Conbniof of2-doxy-D-gkucose
(mm)
I
IE
[image:6.500.60.233.60.371.2]i
FIG. 6. Effect of2-deoxy- D-glucoseon HSV CSA
expression inclones 155-4-213 and 155-4-03.(A)
155-4-213cellswereincubated with andwithout 2-deoxy-D-glucose (1to20mM)at37°C for5h.Symbols:0,
CSA-positive cells;0, cellnumber. (B)Act D(2pg/
ml)-treated and untreated (control) 155-4-03 cells
wereincubatedwith and without2-deoxy- D-glucose (1 to 20mM) at 37°C for20 h. Symbols: 0,
CSA-positivecells in controlcultures;0, ceUnumberin
control cultures; A, CSA-positive cells in Act
D-treated cultures; A, cell number in Act D-treated
cultures.
. A.155-4-213
.S
- 4
''^ v ^ *~~~~~~~21
I.. a 2
B.155-4-03 0
4
r ,-- .
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.500.272.422.338.545.2]426 OKAZAKI ET AL.
40-S30
.~20
10
0
B
_6
04
4
0 5 10 15 20
[image:7.500.78.230.60.322.2]Hoursafterseeding
FIG. 7. EffectofantipainonHSV CSAexpression
and cellgrowth in clone 155-4-213 incubated with and without Act D. Act D (2
pg/ml)-treated
and untreated (control) cells were incubated with and withoutantipain at37'Cfor20h. (A) ExpressionofCSA; (B) cellgrowth.Symbols: 0,control cells with-outantipain; *, control cells withantipain (1 mM); A, Act D-treated cells without antipain; U, Act D-treated cells withantipain (0.5 mM); A,ActD-treated cellswithantipain (1mM).
that suppress Act D-induced CSA
expression,
didnotaffect overall cellular metabolism.
DISCUSSION
We have recentlyfound that the anti-HSV-1
serumused in this study reacts with both early
and late HSV CSA induced in HSV-2-infected
HEFcellsand that the HSVCSA detected with
thisantiserum reaches a maximum before virus
infectivityreaches amaximum in infectedcells (S. Kimura, K. Okazaki, and N. Yoshida,
sub-mitted forpublication). Our previous study (26)
alsodemonstrated that the reactivity of the
an-tiserum in HSV-2-transformed cell line 155-4
wassignificantlydecreased after absorption with HEFcellsinfected with HSV-2 or with
homol-ogous 155-4cells,but not with uninfected HEF
cellsorsimian virus 40-induced tumor cells.
Sincepreviousfindings suggested thatcellline
155-4 consisted of several different cell types
with respect to CSA expression (26), 30 clones
wereisolated from this line and tested forCSA
expression. These isolated clones were classified
into three different phenotypes withregard to
CSAexpression,namely,aCSA-positivetype,a
CSA-inducible
type, and a CSA-negative type (Kimuraetal., submittedforpublication). CSA expression by the CSA-positive type occurred soonafterseedingat37°C.Oftheclonesisolated, 20% belonged tothis type,and clone 155-4-213 wasusedas arepresentative. CSA expressionin thisrepresentative clonerequired incubationof thecellsfor5hat37°Caftertrypsinization andwas not affected by Act D treatment. CSA
expression in the
CSA-inducible
type occurredafter treatment of thecellswith Act Dfor 20h,
but notafterseeding. Oftheclonesisolated,
33%
belonged to this type, and clone 155-4-03 was
used as a representative. Act D-induced CSA expressionin the
representative
clonewasgreat-estwhen2
jig
ofAct D permlwasaddedtothecellsatthetimeofseeding.In theCSA-negative
type,CSAexpressiondidnotincrease after
seed-ingorafter Act Dtreatment.Of theclones,47%
belonged to this type, and clone 155-4-16 was
usedas arepresentative.
40
Be 30
@ 20
LPa- 10
0
12
I?
0
E
.-8
4
Hoursater seeding
FIG. 8. Effectofantipain on HSVCSAexpression and cellgrowthin clone 155-4-03 incubated with and without Act D.Act D(2
lg/ml)-treated
and untreated (control)cells were incubated with and without anti-painat37°C for20h.(A)ExpressionofCSA; (B) cell growth. Symbols: 0, control cells without antipain;0, control cells with antipain (1 mM); A, Act D-treated cells withoutantipain;x,ActD-treatedcells withantipain (0.01 mM);*, ActD-treatedcells with antipain (0.1 mM); O,ActD-treated cells with anti-pain (0.5 mM); A,ActD-treated cells with antipain
(1 mM).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.500.288.434.315.547.2]This study also demonstrated that CSA expressions inthe CSA-positivetype(clone
155-4-213) and theCSA-inducibletype (clone
155-4-A.
155-4-213
I-L
-...-...
B.
155-4-03
W___ ____ __--_
~
-,_
o0.01 0.5 0.1
Concentration of NPGB(mM)
FIG. 9. Effect ofNPGBonHSV CSAexpression in
clones155-4-213and 155-4-03. (A)NPGB(0.01 to0.1 mM)-treated and untreated 155-4-213 cells were
in-cubatedat 37°C for5h. Symbols: 0, CSA-positive cells;0,cellnumber.(B)Act D(2
pg/ml)-treated
anduntreated(control)155-4-03 cellswereincubated with
and without NPGB(0.01to0.1mM)at37°C for20h. Symbols: 0, CSA-positive cells in controlcultures;
0, cellnumber in controlcultures; A, CSA-positive
cells in Act D-treatedcultures;A,cellnumber in Act D-treated cultures.
03) were enhanced by different mechanisms.
CSAexpressionin clone155-4-213 wasinhibited
by2-deoxy-D-glucose,but not byPM, suggesting
that theexpression required glycosylation, but
not activeprotein synthesis. This expression in
the clonewasinsensitiveto theprotease
inhibi-tors antipain and NPGB. On the other hand,
ActD-inducedCSAexpression inclone 155-4-03
was inhibited by both 2-deoxy-D-glucose and
PM, suggesting that the induced expression
re-quired bothglycosylationandprotein synthesis. Thisinducedexpression in the clone was
sensi-tive to antipain and NPGB at concentrations
that had littleeffectonoverall cellular
metabo-lism (DNA, RNA,and proteinsyntheses)orcell viability.
It iswell known that Act D binds to native
DNA and inhibits RNA, DNA, and protein
syntheses (8,40). Thisdrug alsoinduces muta-tions (14) and chromosome abnormalities (32,
36) in eucaryotic cells and has tumorigenicity
(43) andteratogenicity (20) inanimals. Itisnot clear how Act D induces CSA expression in clone 155-4-03. We recently found that CSA expression inclone 155-4-03 (about 40% ofthe cells) is also induced by treatment with the anthracycline antibiotics adriamycin (0.25 ,ug/ ml) and daunomycin (0.25 ,ug/ml), which are
known to bind to DNA (1, 3, 37) and inhibit
both RNA and DNAsyntheses (19,23), asdoes Act D; however, no CSA was induced in this clonebytreatmentwith
5-iododeoxyuridine,
cy-tosinearabinoside,
ormitomycin
C(D. Kako,
Y.Ohnishi,
and S.Kimura, manuscript
in prepa-ration).Furthermore,wefound that in this clonethemaximallevel ofCSA
expression
(about
40%of the
cells)
could be inducedby the simultane-ousadditionof lower doses ofAct D(0.5 ug/ml)
TABLE 2. Effectsof antipainandNPGBonincorporationsof[3H]thymnidine, [3H]uridine,and[3H]leucine into clone 155-4-03 in the presenceandabsenceof ActD
Incorporation (per 105 cells)of:
Group Treatment
[3H]thymidine
[3H]uridine [3H]leucine(cpmx10-3) (cpmx10-3) (cpmx10-4)
A Untreated 1.45 3.55 4.90
Antipain(1.0mM) 1.86(1.28)a 4.55(1.28) 5.72(1.17)
NPGB(0.1mM) 1.45(1.00) 3.38(0.95) 6.03(1.23)
B Act D(2,g/ml) 1.40 0.23 2.67
Act Dplus antipain (0.5 mM) 1.48(1.06)b 0.25(1.09) 2.84(1.06) Act Dplus antipain (1.0 mM) 1.36(0.97) 0.28(1.22) 2.73(1.02)
Act Dplus NPGB (0.05mM) 1.58(1.13) 0.23(1.00) 3.34(1.25)
Act Dplus NPGB(0.1mM) 1.51 (1.08) 0.28(1.22) 2.60(0.97) aValues withinparentheses ingroup A represent the ratio ofcounts per minute incorporated into cells
treated withantipainorNPGBtothatincorporatedinto untreated cells.
bValues withinparentheses in group B represent the ratioofcounts per minuteincorporated into cells
treated withAct Dandantipainorwith Act Dand NPGBtothatincorporatedintoceUlstreated with Act D only.
40
. 20
- 10
0 0
040
& 30
cn
() 20
90
0
0
_6%fr
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.500.63.234.114.351.2] [image:8.500.60.446.486.609.2]and
adriamycin
(0.1
,ug/ml),
each of which alone inducedonly low levels ofexpression (each
about 20% of thecells),
and that the simultaneous addition of themosteffective doses of Act D(2
,ug/ml)
andadriamycin
(0.25Ag/ml)
did notcause CSA induction in morethan 40% of the
cells.Thus, these doses of the
drugs
donotshow additive orsynergic
effects(Kako
etal.,
manu-script in
preparation).
Thesefindings
suggest that CSA induction in clone 155-4-03is dueto acommon
effect(s)
of Act D andanthracycline
derivatives andnot to aside action ofa contam-inant(s) in Act D.
Antipain, a protease inhibitor of microbial origin, isa
relatively
nontoxic(45), low-molecu-lar-weight compound, which inhibits trypsin-and papain-like proteases inparticular
(42).NPGBis oneof several activesite-specific
inhib-itorsof serineproteases, such astrypsin,
throm-bin, plasminogen activator, and
plasmin
(5, 6, 17). Asdescribedabove, thesetwo protease in-hibitors reducedActD-inducedCSAexpression inclone 155-4-03 (CSA-inducible type), indicat-ing that protease(s) plays animportant
role in CSA inductionbyActD. It has beenshownthat A prophage in Escherichia coli is induced by proteolyticcleavage of the Arepressor (41). An-tipain inhibits A phage induction (one of the SOSfunctions) inE.colibyblocking proteolytic
inactivation ofthe repressor (31). Therefore, it is conceivable that aputative repressor(s) ofa
CSA function(s) isinactivated
by
aprotease(s) induced or activatedin the cellsrespondingto ActD,leadingtotheappearance ofCSA. Simi-larly, it wasreportedrecently
that protease in-hibitors,includingantipain,
canblock the chem-ical (28) or physical (35) induction of anen-dogenous
xenotropic
typeCvirus from Kirstensarcomavirus-transformedmouse
celLs,
suggest-ingarole of
proteolysis
invirusinduction. On the otherhand, theincreased expression ofCSA in clone 155-4-213 (CSA-positive type) afterseeding maybe due toa translocation or modificationof apreexisting antigen(s) (possibly protein). Furthermore, CSA expression in thisclone was not affectedby treatment with Act D,
PM, or the two protease inhibitors. Although
the mechanism of this phenomenon is not
known, it might be explained by supposing that
arepressor(s) reponsible for CSA expression is
notproduced or is produced in aninactiveform
inthecellsorthat arepressor-binding site(s) of
DNA is altered so that it is insensitive to the
repressor present in the cells. In a
repressor-controlledsystem, such as the A phage and Lac
system of E. coli, the presence ofconstitutive
regulator and operator mutations has been es-tablishedpreviously (21).
Furtherexperiments are in progress to
deter-mine whether the antigens expressed by the
CSA-positivetype are identical to those induced intheCSA-inducibletype.
LITERATURE CITED
1. Blake, A.,and A. R. Peacocke.1968.The interaction of aminoacridines with nucleicacids.Biopolymers 6:1225-1253.
2. Boyd,A.L.,and T. W. Orme. 1975.Transformationof mousecells after infection withultraviolet irradiation-inactivated herpessimplex virus type 2. Int. J. Cancer 16:526-538.
3. Calendi, E., A.DiMarco, M.Reggiani,B.Scarpinato, and L. Valentini.1965.Onphysico-chemical interac-tions betweendaunomycinandnucleicacids. Biochim. Biophys. Acta103:25-49.
4. Camacho,A.,and P.G.Spear.1978.Transformation of hamster fibroblasts by aspecific fragment of the herpes simplex virus genome. Cell 15:993-1002.
5. Chase, T.,Jr., and E. Shaw. 1967. p-Nitrophenyl-p'-guanidinobenzoate HCl: a new activesite titrant for trypsin. Biochem. Biophys.Res. Commun. 29:508-514. 6. Chase, T.,Jr.,and E. Shaw. 1969.Comparison ofthe
esterase activities oftrypsin, plasmin,and thrombin on
guanidinobenzoate esters. Titration of the enzymes. Biochemistry8:2212-2224.
7. Collard, W., H.Thorton, and M. Green. 1973. Cells transformed byhuman herpesvirus type2 transcribe virus-specific RNA sequences shared by herpesvirus types 1 and2.Nature(London) New Biol. 243:264-266. 8. Cooper,H.L.,and R. Braverman. 1977.The mecha-nismby whichactinomycin D inhibits protein synthesis in animalcells. Nature(London)269:527-529. 9. Courtney, R. J.1976.Herpessimplexvirusprotein
syn-thesisinthepresence of2-deoxy-D-glucose. Virology 73:286-294.
10. Courtney,R.J.,and M.Benyesh-Melnick.1973/1974. Differential effect of2-deoxy-D-glucose on glycopro-teinssynthesized byherpes simplex virus type1and type2.Intervirology 2:120-127.
11. Davis,D.B.,and D. T.Kingsbury.1976.Quantitation ofthe viralDNApresent incells transforned by UV-irradiatedherpes simplex virus. J. Virol. 17:788-793. 12. Duff, R., and F. Rapp. 1971. Properties of hamster
embryo fibroblasts transformedinvitroafter exposure toultraviolet-irradiatedherpes simplex virus type2.J. Virol.8:469-477.
13. Duff,R., and F.Rapp.1973.Oncogenic transformation ofhamsterembryocells after exposure to inactivated herpessimplex virus type1.J. Virol.12:209-217. 14.Fisher,C.R.,H. V.Malling,F. J. DeSerres,andS.
Snyder.1975.Mutagenicity of actinomycin D in Neu-rospora crassa.Mutat. Res. 33:187-192.
15.Frenkel, N.,H.Locker,B.Cox,B.Roizman,and F. Rapp.1976.Herpes simplexvirus DNA intransformed cells:sequence complexity in five hamster cell lines and onederivedtumor.J. Virol. 18:885-893.
16.Glorioso, J. C., and J. W. Smith. 1977. Immune
inter-action withcells infected with herpes simplex virus:
antibodies toradioiodinatedsurfaceantigen. J.
Immu-nol.118:114-121.
17.Goldberg,A.R.1974.Plasminogenactivators ofnormal
andRous sarcomavirus-transformedcells, p. 347-359.
InW.S. Robinson andG. F. Fox(ed.), Mechanismsof virusdiseases, vol. 1. W. A. Benjamin, Inc., Menlo Park, Calif.
18.Gupta, P., and F. Rapp. 1977. Identification of virion polypeptides in hamster cells transformed by herpes simplexvirustype1.Proc.Natl.Acad. Sci.U.S.A. 74: 372-374.
J.
on November 10, 2019 by guest
http://jvi.asm.org/
19.Hartmann, G., H. Goller, K. Koschel, W.Kersten, and H. Kersten. 1964. Hemmung der DNA Abhangi-genRNA-undDNA-Synthese durch Antibiotica. Bio-chem. Z. 341:126-128.
20. Haruta, M. 1968.Teratogenic effectsofactinomycin-D onddO mouse embryos. Acta Pathol. Jpn. 18:267-286. 21. Jacob, F., andJ.Monod. 1961. Genetic regulatory mech-anisms in thesynthesis of proteins. J. Mol. Biol. 3:318-356.
22. Kaluza, G., M. F. G. Schmidt, and C. Scholtissek. 1973.Effect of2-deoxy-D-glucose on the multiplication of Semliki forest virus and the reversal of block by mannose.Virology 54:179-189.
23. Kersten, W., H. Kersten, and W. Szybalski. 1966. Physicochemical properties of complexes between de-oxyribonucleic acid and antibiotics which affect ribo-nucleicacid synthesis (actinomycin, daunomycin, cine-rubin, nogalamycin, chromamycin, mithramycin, and olivomycin).Biochemistry5:236-244.
24. Kimura,S.,J.Esparza,M.Benyesh-Melnick,and P. A.Schaffer. 1974. Enhanced replication of tempera-ture-sensitive mutants ofherpessimplex virus type 2 (HSV-2) at the nonpermissive temperature in cells transformedby HSV-2.Intervirology3:162-169. 25. Kimura,S.,V.L.Flannery,B.Levy,andP.A.
Schaf-fer.1975.Oncogenic transformation of primary hamster
cellsbyherpes simplex virus type 2(HSV-2) andan HSV-2temperature-sensitivemutant.Int.J.Cancer 15: 786-798.
26.Kimura,S.,K.Okazaki,N.Yoshida,and Y.Ohnishi.
1979. Effect ofactinomycin D on the expression of herpes simplex virus-common surface antigen incells
transformedby herpessimplex virus type 2. J. Virol. 29:161-169.
27. Kucera, L.S., and J. P. Gusdon. 1976. Transformation of humanembryonic fibroblastsbyphotodynamically
inactivated herpessimplexvirustype 2 atsupraoptimal temperature. J.Gen. Virol. 30:257-261.
28. Long, C. W.,J. A.Bruszewski,W.L.Christensen,
and W. A. Suk.1979.Effects of protease inhibitorson chemical induction of type C virus. Cancer Res. 39: 2995-2999.
29. Macnab,J. C. M.1974. Transformation ofratembryo
cellsbytemperature sensitive mutants of herpes sim-plex virus. J. Gen. Virol. 24:143-153.
30. Macnab, J. C.M.,and M. C.Timbury.1976.
Comple-mentation ofts mutantsbyaherpessimplexvirus
ts-transformedcellline. Nature(London)261:233-235. 31. Meyn, M.S.,T.Rossman,and W. Troll. 1977.A
pro-teaseinhibitor blocks SOS functions in Escherichia coli:antipainpreventsA repressorinactivation,
ultra-violetmutagenesis, and filamentousgrowth. Proc. Natl. Acad. Sci. U.S.A. 74:1152-1156.
32. Miles, C. P. 1970. Labeling and other effects of actino-mycin D on human chromosomes. Proc. Natl. Acad. Sci. U.S.A.65:585-592.
33. Minson, A. C., M. E. Thouless, R. P.Eglin, and G. Darby. 1976. The detection of virus DNA sequences in aherpes type 2 transformed hamster cell line (333-8-9). Int. J.Cancer 17:493-500.
34. Nahmias, A.J., I.Pelbuno, K. E. Schnewis, A. S. Gordon, and D. Thies. 1971. Type-specific surface antigens of cells infected with herpes simplex virus (1 and 2). Proc. Soc.Exp. Biol. Med. 138:21-27. 35. Niwa, O., and T.Sugahara.1979.Effect ofantipain on
radiation induction of endogenous type-C virus from mousecells in vitro. Intervirology 12:120-123. 36. Osterag, W., and W. Kersten. 1965. The action of
proflavin and actinomycin D in causing chromatid breakage in humancells. Exp. Cell Res. 39:296-301. 37. Pigram, W. J., W.Fuller, and L.D.Hamilton. 1972.
Stereochemistryof intercalation: interaction of dauno-mycin with DNA. Nature (London) New Biol. 235:17-19.
38. Rapp,F., and R. Duff. 1972. In vitro cell transformation by herpesviruses. Fed. Proc. 31:1160-1168.
39. Reed, C.L., G. H. Cohen, and F. Rapp.1975.Detection of a virus-specific antigen on the surface of herpes simplex virus-transformed cells. J. Virol.15:668-670. 40.Reich, E., A. Cerami, and D. Ward. 1967. Actinomycin,
p. 714-725. In D. GottliebandP.Shaw (ed.), Antibiot-ics, vol.1.Springer-Verlag, New York.
41.Roberts, J. W., and C. W. Roberts. 1975. Proteolytic cleavage of bacteriophage lambda repressor in induc-tion.Proc. Natl. Acad.Sci. U.S.A. 72:147-151. 42. Suda, H.,T.Aoyagi, M.Hamada,T.Takeuchi, and
H. Umezawa. 1972.Antipain,a newproteaseinhibitor isolated from actinomycetes. J. Antibiot. 25:263-266. 43. Svoboda, D.,J.Reddy,andC.Harris.1970.Invasive
tumorsinduced inratswithactinomycin D. Cancer Res. 30:2271-2279.
44.Takahashi,M., and K. Yamanishi. 1974. Transforma-tion of hamsterembryo and human embryo cells by temperature sensitive mutants of herpessimplex virus type2.Virology61:306-311.
45. Umezawa,H.1972.Enzymeinhibitors of microbial ori-gin, p. 29-32. University of Tokyo Press,Tokyo. 46. Vestergaad, B.F., andP.C. Grauballe.1977.Crossed
immunoelectrophoretic identification of partially puri-fied typecommonand typespecific herpessimplex virus glycoproteinantigens. Proc. Soc. Exp. Biol. Med.156: 349-353.

![TABLE 2. Effects of antipain and NPGB on incorporations of[3H]thymnidine, [3H]uridine, and [3H]leucineinto clone 155-4-03 in the presence and absence ofAct DIncorporation of:](https://thumb-us.123doks.com/thumbv2/123dok_us/1479376.100574/8.500.63.234.114.351/effects-antipain-incorporations-thymnidine-uridine-leucineinto-presence-dincorporation.webp)
