Restriction of Viral Replication by Mutation of the Influenza Virus Matrix Protein
Full text
Figure
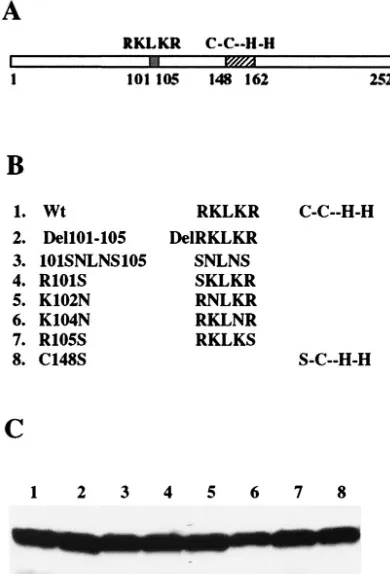
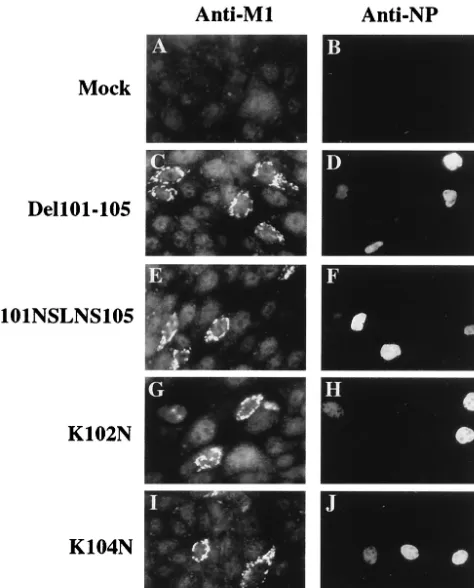
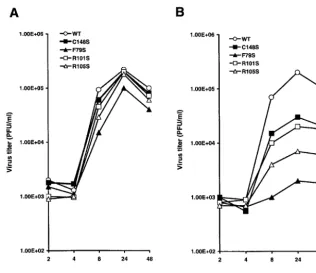
Related documents
ZAP, a CCCH-type zinc finger protein, inhibits porcine reproductive and respiratory syndrome virus replication and interacts with viral Nsp9.. All
Salinomycin Inhibits Influenza Virus Infection by Disrupting Endosomal Acidification and Viral Matrix Protein 2 Function.. Yejin Jang, a Jin Soo Shin, a,b Yi-Seul Yoon, a Yun Young
RNA interference (RNAi) silencing of NXF1 in human embryonic kidney 293 cells inhibited viral gene expression and overall replication (11) and was subsequently shown to reduce
In this study, we showed that deletion of 6K1 or a short motif/region of 6K1 in the full-length cDNA clones of plum pox virus abolishes viral replication and that mutation of the N-
Host Protein Moloney Leukemia Virus 10 (MOV10) Acts as a Restriction Factor of Influenza A Virus by Inhibiting the Nuclear Import of the Viral Nucleoprotein.. Junsong Zhang, a,b,d
The nuclear location of PML, a cellular member of the C3HC4 zinc-binding domain protein family, is rearranged during herpes simplex virus infection by the C3HC4 viral protein
The diffuse nuclear staining pattern of PrV UL3 did not change in the absence of several other viral proteins which are found in the nucleus during infection, in particular those
To address this issue, we used H7 as a biochemical tool to suppress late viral protein synthesis in influenza virus-infected cells and block vRNP export.. We then expressed
