Copyrighti 1974 AmericanSocietyforMicrobiology PrintedinU.S.A.
Structural
Components
of
Oriboca
Virus
ROBERT R. ROSATO, MARY LOUISE ROBBINS, AND GERALD A. EDDY
U.S. Army Medical ResearchInstituteof InfectiousDiseases, FortDetrick,Frederick,Maryland21701and Department ofMicrobiology, The George Washington University Medical Center,Washington, D.C. 20037
Received forpublication 15 November 1973
Analysis of purified Oriboca virions by neutral, sodium dodecyl sulfate
polyacrylamide-gel electrophoresis indicated the presence of three structural
polypeptides designated V-1, V-2, and V-3 on the basis of their relative electrophoretic mobilitiesin8%gels.PolypeptidesV-2 and V-3areglycopeptides associated with the virion envelope as demonstrated by the preferential
incorporation oflabeled glucosamine into thepolypeptides and by releaseofthe
polypeptides
fr6m
theintact virionby thenonionicdetergentNP-40.Polypeptide V-1 is the protein component of the nucleoprotein core of Oriboca virus as evidenced by the specific incorporation ofuridine into the nucleoprotein, its release from the intact virion by NP-40treatment, and its separation by both rate-zonal and isopycnic density gradient centrifugation from both the intact virion and envelope components. Molecular weights have been tentativelyassignedtothepolypeptides by extrapolationfromthe structuralpolypeptidesof
Sindbis viruswhen bothare runinthesamegel.PolypeptideV-1hasanapparent molecular weight of 20,000 to 23,000; V-2, 30,000 to 32,000; and V-3, 83,000 to 85,000.
The prototype strain BE An 17 of Oriboca
viruswasisolatedfrom asentinel
monkey
intheOribocaforest,
Belem,
Brazil in 1954(1,7).
Thisviruswas soonincorporated into thenew,
previ-ously undescribed arbovirus
serological
group Cwhich also contained
Apeu,
Caraparu,
Maritu-ba, and Murutuca viruses (6, 19).
Group
Cvirusesare nowclassifiedas aseparate
serologi-cally
defined group within theBunyamwera
supergroup (23, 24). Electron microscopy has
definedavirion90to 100nmindiameterwhich
consistsofa
central,
electron-dense "core"60to70 nmindiameter surrounded
by
a15- to 20-nmelectron-dense outer portion with
spikes
pro-truding5 to 6 nm from thesurface (2, 4, 10, 15,
16). Oriboca virus obtained from mouse
brain,
serumandliver, and cell culture consists of four
physically distinct components separable by
buoyant and rate-zonal density gradient
cen-trifugation. Two ofthe conponents, ofmedium
(MDCF)
andlight
(LDCF)densities,
exhibitcomplement-fixing activities, whereas the
re-maining two, HA-2 and VHA, have
hemag-glutinating characteristics, the latter being
associated also with virion infectivity (11).
Oriboca virus contains three structural
poly-peptides with tentatively assigned molecular
weights of 83,000 to 85,000; 30,000 to 33,000;
and20,000 to23,000. However, thesestructural
proteins have not been characterized as to
functional roles or locations within the virion. Similar structural proteins were also reported
forBunyamwera, BFS-283, Tahyna, and
Muru-tucavirions(Rosato etal., ActaVirol., inpress), thus further indicatingthe homogeneity ofthe
Bunyamwera supergroup viruses withrespect to
serological (6, 7, 19), morphological (2, 4, 10, 15), andmorphogenetic(15, 16)characteristics.
Our knowledge ofthe molecular structure of
Oriboca virus is limited, since little is known
concerning the structural composition of the maturevirion. Aninitial step inprovidingsuch
information isthedeterminationofthenumber,
size, gross composition, and location of the structural polypeptides in the virion and in subvirion particles. The present studies were undertaken to determine some physical and chemical properties of Oriboca virions.
MATERIALS AND METHODS
Virus. Oriboca virus, strain Be An 17, was obtained from the American Type CultureCollection (ATCC),
Rockville, Md., and carried the following designa-tions:ATCC no. VR 310,SM/8 lot no. 1, Nov. 1968. It waspassed once in 1- to 3-day-old suckling mice by intracerebral inoculation, the brains were harvested anda 20%suspensionwasprepared, lyophilized, and
designated as seed stock virus.Working virus stocks were prepared in a similar manner from the seed stocks butwere wet-frozenat -70 C. Working virus stocksweredeterminedtocontain 3.63 x 106PFU/ml, 780
on November 10, 2019 by guest
http://jvi.asm.org/
COMPONENTS
andwereverifiedtoconsist ofOribocavirusby plaque reduction-neutralization tests.
Preparation and purification of radioactive
viri-ons.Oribocaviruswaspropagatedin 64-oz.
roller-bot-tle cellcultures ofLLC-Mk, maintained in medium 199containing 10%fetal calfserum(FCS), penicillin
G (100 U/ml), streptomycin sulfate (100 sg/ml), and amphotericin B(fungizone, 50ug/ml). Priorto infec-tion, monolayerswerewashed twicewith 0.9% NaCl, oncewith Hanksbalanced salt solution (HBSS), and once with medium 199-deficient medium (Earle or
Hanks salts)containing 2%dialyzedFCS and antibi-otics. Medium 199 (Grand Island Biologicals, Grand Island, N.Y.) was deficient in either all amino acids
or all amino acids and glucose. Depending on the
radioactive compoundtobeincorporated, the
concen-tration oftheunlabeledaminoacidsorglucoseadded
was adjusted to 1/10 to 1/20 the normal concentra-tions. Virus wasdiluted indeficient medium, added at a multiplicity of inoculum of 0.01 to 0.1, and allowedtoadsorb for 60to90min at36C. After the adsorption period the inoculawere removed and the
infected monolayerswerewashedasdescribed above.
A 30-or40-mlamountof deficient mediumwasadded
and the cultures were returned to 36C. Labeled substances were added 24 h after the adsorption periodatJCi10 of3H(G)-L-aminoacidmixtureperml
containing 15 amino acids (specific activity 23.6 Ci/mmol), 3H(6)-D-glucosamine-hydrochloride (spe-cific activity 7.3 Ci/mmol), and at 3H(4,5)-L-lysine (specific activity20to40Ci/mmol) insingly labeled experiments, and at 10 gCi/ml for tritiated
com-pounds and2ACiof "4C(U)-L-amino acid mixtureper
ml (specific activity 3.5 Ci/mmol) indoubly labeled experiments. All labeled substances were obtained
from New England Nuclear Corp., Boston, Mass. Cultures requiring pH adjustment were returned to pH7.0to7.2by the additionof 7.5%NaHCO orby
directgassingwithCO2.Culture fluidswereharvested
2 to 3 days after infection when cytopathic effects
weredistinctly visible. The infected fluidwasclarified
by centrifugation at 380 x g for 10 min at 4C (International PR-2, no. 253 rotor) and the superna-tantwascentrifuged at 12,000 x gfor 30 minat4C (Sorvall RC2-B, SS-34 rotor).The finalsupernatant was mixed with cold, saturated(NH4)2S04 solution,
previously bufferedtopH7.2to7.4by the additionof 1 M Tris. The buffered (NH4)2S04 wasadded
drop-wise to the virussuspensions (final concentration of 60%
[vol/voll
ammonium sulfate) to precipitate the virus. The suspension was held at 4C for 30 min, duringwhich time itwasswirledat 10-minintervals. Thesuspension was centrifuged at 10,400 x gfor 30 min at 4 C (RC2-B, GSA rotor) and the resulting pelletwasresuspendedin 1.0to 1.5ml ofTSE buffer (0.01 M Tris-hydrochloride, 0.15 M NaCl, 0.001 M EDTA), pH 7.2. Purified virus was obtained byprocessingthissuspension byrate-zonaldensity
gra-dientcentrifugationinwhich virionswereobtainedas
asingle peakin linearsucrosegradientsformedovera
70%sucrose cushion. All sedimentation patterns are
presentedfromrighttoleftinthe Results section. The first fraction isatthe bottom of the tube.
Plaqueassay of Oriboca virus. Confluent
mono-layers of LLC-MK2 cells were infected with 10-fold
serial dilutions ofOriboca virus prepared in normal growth medium. After adsorption for 60to 90 min, they were overlaid with 5 ml of medium containing finalconcentrations of 1.0%agarose(SigmaChemical
Co., St.Louis, Mo.), 1 x M-199medium, 10.0%
heat-inactivated fetal calfserum, 0.3% sodium bicarbon-ate,0.02%DEAE, and antibioticsatpreviously stated concentrations. On thesecondorthirdday of
incuba-tion at 36C, 5 ml of a second overlay of the same
compositim astheprimary plus4.0%final concentra-tion ofa 1:300 dilution ofneutral red in waterwas
added. After incubation for2to4hat36C,thecells
wereleftatroomtemperatureovernight; plaqueswere
counted thenextday.
Sucrosedensity gradient centrifugation. Linear 15 to 50% (wt/vol) sucrose gradients prepared with
ribonuclease-free sucrose (Schwarz/Mann,
Orange-burg, N.Y.) in TSE buffer were formed in mixing
chambers ofthetypeoriginally described by Britten and Roberts (3). Large gradients of 28 or 29 ml,
required for initial viruspurification,werecentrifuged at64,000 x g in the Spinco SW25.1 rotorfor 3 hat 4C. Small volumes were applied to 4.4- or 4.8-ml
gradientsofidenticalcompositionand the tubeswere
centrifugedat 204,000x gin theSpincoSW50Lrotor for60minat4C.
Forisopycnic experiments, sampleswereappliedto gradients of20to70%sucrose (wt/vol) indeuterium oxide (Isomet Corp., Oakland, N.J.)andcentrifuged at 204,000 x g in the Spinco SW50L rotor for the indicated time intervals. Density wasdetermined by
the direct weighing of ice-cold 50-uliter portions of each 0.20-ml fraction. All gradients were collected
from the tube bottom in an apparatus designed to allow accurate displacement of a set volume of the gradient by the addition of an equal volume of mineral oil overlay. Usually 0.20-ml fractions were
collected from small gradients and 1.0-ml volumes fromlarge gradients.
Polyacrylamide gel electrophoresis. Polyacryla-mide gel electrophoresis was performed essentially
accordingtoMaizel(13)asmodifiedby Shapiroetal. (17) with the following exceptions. Recrystallized, electrophoresis-grade acrylamide and N,N'-bis-methylene acrylamide (Bio-Rad Laboratories, Rich-mond, Calif.) wereused without furtherpurification.
Samples, whichnormallycontainedsucroseand
var-iedinvolume from 25to100uliters,weredegraded by
the addition of both SDS and 2-mercaptoethanol (2-ME), each to a final concentration of 1%, then
heated in sealedglasstubes at100 C for10min.After the addition ofbromophenolbluetoafinal concentra-tion of 0.05%, samples were applied to the gels, overlaid with 1:5 diluted electrophoresis buffer, and electrophoresed.Electrophoresiswasperformedat20 Vuntil thesampleentered thegel,thenat40Vfor 10 min,andat60V until thedyefrontmigratedabout 5 to 6 cm, which usually required90to 120min.The gels were then removed, placed in a metal trough (Scoopula, Fisher Scientific Co., Pittsburg, Pa.)
fro-zen on a blockof dry ice, and pressed while frozen against atransverseslicercontainingrazorblades at
1-mm intervals. The slices were removed with a
VOL. 13, 1974
781
on November 10, 2019 by guest
http://jvi.asm.org/
dissecting needle, placed in scintillation vials, and counted as described in the section on the countingof radioactive samples. All the electropherograms are presentedwith migrationfromcathode(left)toanode (right).
Degradation of virions. Purified virions were de-graded by the addition of thedetegentNonidet P-40 (NP-40, Shell Oil Co., Tulsa, Okla.) to a final concentration of 1%. Samples were mixed on a Vortex mixer and held at 4C for 20 min, after which the degradedvirionswereseparated into their component parts by rate-zonalcentrifugation on small gradients.
Counting ofradioactive samples. Liquid 25- to
50-Mliter samples were assayed in scintillation fluid
(10ml) consisting of 150 mlofLiquifluor and 100 ml of Protosol (both from New England Nuclear Corp., Boston, Mass.) per 3 liters ofspectral grade toluene. Gel slicesweretreated identically and thenincubated overnight at 36 C to allow for swellingof the slices before counting. Radioactivesampleswerecounted in eithera Packard no. 3375 (3H efficiency, 40%) or a Nuclear-Chicago, Mark II liquidscintillationcounter
(single-label 3Hefficiency 61%; double-label 3H effi-ciency 52% with 11% "4C to 3Hspillover). Afterthe start ofthe project acomputer program was devel-opedfordirect correction of the data from the tapefor
background, spilloverfrom "C to 3H channels, and conversion ofcountsper minutetodisintegrations per minute;therefore resultsaregivenineithercountsper minute ordisintegrations per minute.
RESULTS
Preparation of radiolabeled Oriboca
viri-ons. When Oriboca was grown and labeled in
LLC-MK2
cellculture,
concentratedby
ammo-niumsulfate
precipitation,
andcentrifuged
on arate-zonal,
15 to 50%(wt/vol)
linear sucrosegradient,one
principal
areaofradioactivity
wasobserved which
appeared
in the center ofthegradient and
corresponded
to thearea ofmax-imuminfectivity.
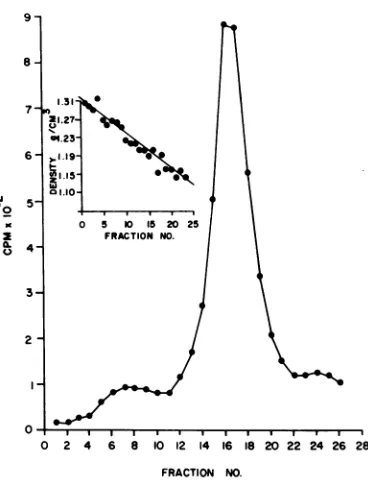
Isopycnic centrifugation of
purified
labeledvirions indicated an
equilibrium density
of 1.187 to 1.190g/cm3
for Oriboca virus(Fig.
1inset, fractions 16 and
17).
Polyacrylamide gel
analysis
ofthe virions atdensity
1.187to 1.190g/cm
3(fractions 16, 17) indicatedacharacteris-tic polypeptide profile for Oriboca virus as
illustrated by the
"4C-amino
acid-labeledpor-tion of the
double-labeled
preparation
inFig.
4A. Onoccasion, thedensity ofOribocavirions varied from a minimum of 1.185
g/cm3
to amaximum of 1.221
g/cm3.
Electropherogramsof the more dense samples indicated that the virions contained a slightly larger proportion(10%) of V-1 polypeptide than is found in virions from the middle density range. There
was also a comparable decrease in the amount
ofV-3polypeptide.
Polypeptides ofOriboca virus. Purified
vi-0
5-K 0 5 I520 25 0 4_ FRACTION NO.
3-2
O-
I4 I6 2 II*I0 2 4 6 8 0 12 14 16 18 2022242628
FRACTION NO.
FIG. 1. Isopycniccentrifugation ofrate-zonal puri-fied Oriboca virions. Centrifugation was for 18 h at 204,000 x g at 4 C inan SW50L rotor. The linear gradient was 20 to 70%(wt/vol)sucrose indeuterium oxide. Sedimentation is from right to left. Symbol 0, 3H-aminoacids.
rions,labeled with glucosamine, lysine,uridine,
or an amino acid mixture, were degraded and
analyzed by electrophoresis on polyacrylamide
gels. The
electropherogram
(Fig. 4A) depictingthe
'"C-amino
acid-labeled portion of intactvirions shows that Oriboca virus contains three
polypeptides designated V-1, V-2, andV-3.
On the assumption that all structural
poly-peptides are synthesized at the same rate
dur-ing thelabeling period, andthat incorporation
of isotope is uniform, the distribution of the
radioactivity between the polypeptides might
beusedtoestimatethemassdistributionofthe
three
polypeptides
in the purified virion whenlabeled with a mixture containing 15 amino
acids. Analysis of electropherograms of
poly-peptides labeled with the
3H-amino
acidmix-ture is given in Table 1.
Comparable
experi-ments in which
3H-lysine
and 3H-glucosamine were also used to determine the distribution ofthepolypeptides arealsogiven. The amino acid
distribution provides a base line from which
comparisons with the lysine and glucosamine
distributions could be made. There is a slight
increase in the lysine content of the V-1
poly-peptide above base-line levels which, when
coupledwiththe very low glucosamine content
ofthe samepolypeptide, indicates thatV-1may
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.494.266.450.69.310.2]be the polypeptide component of the nucleo-protein core.
The data for percent distribution of the amino acid mixture were also used to approxi-mate the molar ratios of the polypeptides in
which the ratios were determined by dividing
the estimated molecular weight of the
polypep-tide into the total disintegrations per minute
under thepolypeptide peakasvisualized by the
electropherograms. The estimated molar ratios were V-3, 1.00; V-2, 1.09 + 0.06standard
error;
V-1, 2.42 + 0.08 standard error.In an attempt todetermine the location of the
polypeptides within thevirion, purified labeled
virions were degraded with NP-40 and the components were separated by rate-zonal
cen-TABLE 1. Distribution of the structural polypeptides inOribocavirusa
Distribution(%7c) Polypeptide
V-3 V-2 V-1
3H-amino acid mixture .... 45-50 18-25 22-32
3H-lysine ... 42-48 20-22 32-36 3H-glucosamine ... 58-65 35-36 <5
aDetermined as the totalcounts under a specific
polypeptide peak divided bythe total counts under the three polypeptide peaks as observed in electro-pherograms.
81 7
-
6-
5-32
0
,I
-I
A
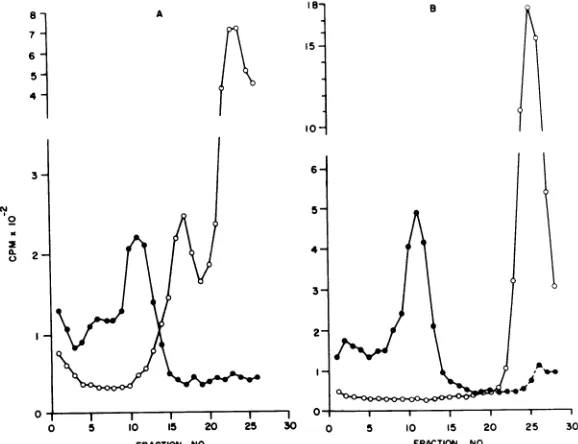
trifugation. We found that dissociation ofthe
virionenvelopefrom the core was complete in 20 min at 4 C. Degraded virions, labeled with
3H-lysine or 3H-amino acids, sedimented as a
clearly defined peak 5 to 6 fractions higher in
the gradients than the undegraded control
vi-rions (Fig. 2A). This will be referred to as the
"peak" component. A new area ofradioactivity
also appeared at the top ofthe gradient to be
referred to as "top component." Degraded
3H-glucosamine-labeled virions did not produce a
peak; thebulkofthe radioactivitywasfoundat
thetop ofthe gradient (Fig. 2B).
Polyacrylam-idegelelectrophoresisoftheundegradedcontrol
virions gave the expected structural profile.
Similar analysisofthepeak andtop component
ofdegraded 3H-labeled virions and the
calcula-tion of percent distribution from the
electro-pherogramsdemonstratedthatthe peak compo-nentcontain
'-85%
of theincorporated 3H-aminoacid label and -75 to 100% of the 3H-lysine
label.
ThetopcomponentcontainedV-3plusV-2 at
base-line or slightly increased levels, and a
decreasedamount of V-1 polypeptide.
A 31-h isopycnic centrifugation of pooled
3H-lysine-labeled peak component showed an
equilibrium
density of >1.310g/cm3,
i.e., thematerial passed
through
the 20 to 70%deuter-iumoxidegradient.Polyacrylamide gel analysis
0B-15
10
-
6-
5-
4-3-I
B
A
25 30
FIG. 2. Rate-zonalcentrifugation of NP-40-degraded purifiedOribocauirions. Centrifugation wasat204,000
x gfor60mininanSW50Lrotoron 15 to50%(wt/vol)lineargradients. Symbols:*,Undegradedvirions;0,
NP-40-degradedvirions.A,Sedimentationpattern ischaracteristic for 3H-lysine-and3H-aminoacid-labeled virions.B, 3H-glucosamine-labeled virions. Sedimentation isfrom right to left.
I --'--I-- l l l
5 10 I5 20 25 30 0 5 10 15 20
FRACTION NO. FRACTION NO.
783
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.494.42.238.262.333.2] [image:4.494.100.389.386.608.2]confirmed that this material was V-1 polypep-tide. Pooled, NP-40-released, 3H-lysine-labeled top component did not band but was found
throughout the top two-thirds of the gradient,
emphasizing the heterogeneity of
NP-40-released top components.
Although the data derivedfromthepreceding
experimentsare indicative of thepossible
loca-tion of the polypeptides in the virion, they
cannot be -considered conclusive since the
nu-cleoprotein core material has not been
specifi-cally labeled.
To specifically label both the RNA and
pro-tein portions of the nucleoprotein component,
virions were prepared, doubly labeled with
3H-uridine and a "C-L-amino acid mixture,
concentrated, and purifiedasdescribedin
Ma-terials and Methods. The relative amounts of
label incorporated intothe virion are shown in
Fig. 3A. Infectivity asPFU was determined for
odd-numbered fractions and corresponded to the area ofradioactivity. An electropherogram
of the "4C-amino acid-labeled polypeptides of virions from the major area (Fig. 3A, fraction 13) showed a characteristic profile. The
3H-uri-dine-labeled RNA did not significantly enter
the gel, and more than 80% was found in the
first two slices.
The purified virions from fraction 13 (Fig.
3A) when degradedby NP-40 gave
characteris-ticsedimentationpatterns on sucrosegradients
(Fig. 3B). The percentage of 3H-uridine in the
peak comparedtothepercentage in the control
in which both were approximately 80 to 85%,
and the low value in the region of top
compo-nentsupported theassumptionthatpeak
mate-TABLE 2. Distribution of thedoubly labeled OribocapolypeptidesreleasedbyNP-40degradationa
Distribution(%)
Labeled material
3H-uridine 14C-amino acids
Polypeptide Topb V-3 V-2 V-i Top V-3 V-2 V-i
Control (undegraded) ... 77-90 6-21c 0 0 0 50-51 17-18 22-28
Peak component ... 90-98 0-8c 0 0 0 0-4c 0 83-92
Top component ... 4 0-10 0-7 0 0 64-70 19-24 0
aDetermined aspercentage of the totaldisintegrations per minute recoveredinthepolyacrylamidegel.
bActivityinthe first 1 to2slicesofthegel.
cActivitywas 1 to 2slicestotheleft of the V-3peak andare notconsideredtobe partofV-3butaseparate entity.
A 9
dt
30-
20-
10-0
i
B
5 l0 I5 2a
FRACTION NO.
FIG. 3. Rate-zonal centrifugation of doubly labeled Oriboca virions. A, Primary rate-zonal purification; centrifugation was at64,000 x gfor 180 min inanSW25.1 rotor at4 C. Symbols: 0, 14C-amino acids; 0,
3H-uridine;0,PFU. B, NP-40-degraded purified virions,centrifugationasin Fig.2;(A) "4C-amino acid-and(0)
9H-uridine-labeled degraded virions; (0) "4C-amino acid- and(0) 3H-uridine-labeled undegraded control virions.
784
50
-40 30
-20
12-1
10
'o
0
0 5 10 15 20 25
FRACTION NO.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.494.114.405.418.610.2]rial was in fact nucleoprotein and therefore contained the V-1 polypeptide.
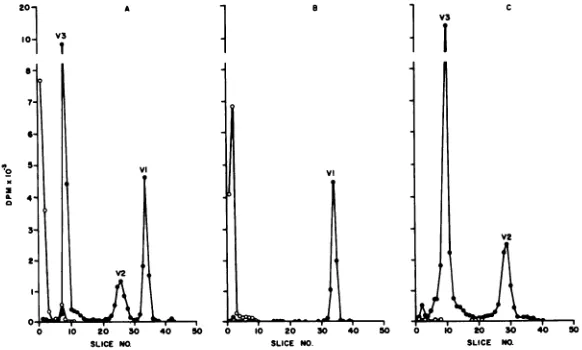
Electropherograms of the polypeptides
re-leased by NP-40 treatment and separated by
rate-zonal density gradient centrifugation are
presented in Fig. 4A-C. The percent
distribu-tionscalculated from theelectropherogramsare
summarizedinTable 2. In the untreated control
(Fig. 4A) more than 85% of the 3H-uridine
remained on top ofthe gel and a minorpeak
representing about 10% of the total activity
appeared in slices 6 to 8. The "C-amino acid
polypeptide profile of the control was normal
(Fig. 4A). Analysis of the peak component
showed more than 95% of the
3H-uridine-labeled portion on top ofthegel andmorethan
85% of the total activity of the "C-amino
acid-labeled protein portioninthe V-1
polypep-tide (Table 2). The calculations ofthe percent
distribution
ofthepeakcomponentclearly
showits purity. Essentially all of the 3H-uridine
remains on top of the gel, and the protein
moiety is not contaminated with V-3 and V-2
components (Fig.4B). Thetopcomponent(Fig.
4C)
containedrelatively
little 3H-uridine, andthe bulk of the
"IC-amino
acid label wasinV-3and V-2; V-1 contained very little of the total
4C-amino acid activity.
DISCUSSION
Our data indicate that Oriboca virions
ob-tained from
LLC-MK2
cell cultures have anequilibrium
density insucrose-D20
of 1.187 to1.190
g/cm3,
a value which differsslightly fromthatobtained withvirions from mouse brainor
liver
(intracellular
virus) and virions derived20-10- V3
0
1
from BHK-21 cell culture or mouse serum
(extracellular virus)(11).Aswehaveindicated,
virions from the more dense samples (1.221
g/cm3)
contained aslightly largerproportion ofthe V-1 polypeptide than is found in virions
from the middle density range. There is also a
comparable decrease in the amount of V-3
polypeptide. This might be expected if, in fact,
the virion had lost a portion of the outer
membraneorpossibly the spike-likeprojections
reported to be present on Bunyamwera
super-groupviruses(10, 15, 16).V-1 isthe polypeptide
portion of the nucleoprotein core of Oriboca
virus; therefore, an increasein the ratio ofV-1
tothe otherpolypeptidesinapartiallydegraded
virion could lead to an apparent increase in
viriondensity. The dataonpercentdistribution
ofradioactive lysine and glucosamine
incorpo-ration indicate that the V-1 polypeptide is
associated with the nucleoprotein portion and
V-2 and V-3 with the envelope components of
the virion. Normally, a high lysine content is
indicative of the presence ofthe protein moiety
ofthe nucleoprotein core (12, 20) and
glucosa-mine ofenvelopecomponents (5,21). Thesame
distribution data when used to calculate the
molar ratios of the polypeptides correspond
reasonably with those reported (1:1:2) for La
Crosse virus, a member ofthe Californiagroup
ofthe Bunyamwerasupergroup (14).
The
previously
reported (R. Rosato et al., inpress) molecular
weight
estimates ofthestruc-tural
polypeptides
of Oriboca virions of V-3,83,000to85,000;
V-2,
30,000to33,000; andV-1,
20,000to23,000,correspondtothosereportedby
McLarran etal.
(14)
for La Crosse virusofV-3,
1
10 20 30 40 SLICENO.
V3
V2
[image:6.494.98.389.441.616.2]50 0 10 20 30 40 SO SLICE NR
FIG. 4. Polyacrylamide gel electropherograms of NP-40-degraded purified Oriboca virions. Samples were obtainedfrom the rate-zonalcentrifugationrunof Fig. 3.A, Undegradedcontrol; B,peak component; C, top component; *, 'IC-aminoacids;0, 3H-uridine.
785
VI
0
on November 10, 2019 by guest
http://jvi.asm.org/
85,000; V-2, 45,000; andV-1, 25,000.We have on occasion assigned a molecular weight of 34,000 to 35,000 to V-2 when extrapolated from the
standard curve based on the molecular weights
of the polypeptides of Sindbis virus (21) but
have not observed a molecular weight greater than 35,000. If, in fact, the inability to detect
the 45,000-dalton polypeptide is due to
propa-gation of virions in amino acid-deficient
me-dium as suggested by McLarren et al. (14) for
Uukuniemi virus, it does not explainthe pres-ence ofthe30,000- to32,000-dalton glycosylated
V-2 polypeptide reportedhere.
PolypeptidesV-3 and V-2 areassociatedwith
the envelope structure ofthe virion as
demon-strated by the preferential incorporation of
labeled glucosamine into the polypeptides and
by the release ofenvelope components from the
virion byNP-40. PolypeptideV-1 isthe protein
moiety of the nucleoprotein core of Oriboca virus as evidenced by the incorporation of
uridine, its release from the intact virion by
NP-40in arelativelypure state, and its
separa-tion by both rate-zonal and isopycnic density
gradient centrifugationfrom both intactvirions andenvelope constituents.
Similar results as to the number and
rela-tive sizes of the structural
polypeptides
havebeen obtained for Kaeng Khoi virus (W. A.
Neill, personal communication). Kaeng Khoi
virus is an ungrouped bat virus,
morphologi-cally similartoBunyamwerasupergroup viruses
but asyet not shown tobe relatedserologically
tothe supergroup. Preliminary polyacrylamide
gel analysis of Pacui virus, an ungrouped
ar-bovirus morphologically indistinguishable from
Bunyamwera supergroup viruses, indicated a
similar polypeptide structural profile as tothe
number and relative sizes of its structural
polypeptides (unpublished data).
The dataconcerningLaCrosse virus
polypep-tides when added to those previously reported
forBunyamwera, BFS-283, Tahyna, and
Muru-tucu virions (Rosato et al., in press) and Ori-bocavirusreported heresupport thecontention thatarboviruses classed together on the basis of
serological relationships (1, 6, 7, 9, 23, 24) also
tend tobe alike ingeneralmorphology (2, 4, 10, 15, 16), mode of morphogenesis (15, 16) and the number, site of virion origin, and apparent molecular weights of their structural polypep-tides (8, 12, 18, 22).
Furtherstudy of many viruses of the various serogroups of the current arbovirus classifica-tion will be necessary to determine in a finite way if, in fact, the antigenic configuration,
which is abiologicexpression of chemical struc-ture, correlates also with the similarities in
structural polypeptides, morphology, and mode
ofmorphogenesis.
ACKNOWLEDGMENTS
Wewish toacknowledge the technical assistance of Mi-chael D. Douglas and Keith M. Hendrix. R. R. R. submitted portionsofthis work inpartialfulfillment of therequirements for the degree of Doctor of Philosophy at The George WashingtonUniversity.
LITERATURE CITED
1. Berge,T.O., R. E. Shope, T. H. Work, and R. M.Taylor. 1970. Catalogue of arthropod-borne viruses of the world. Amer.J. Trop. Med. Hyg. 19:1082-1084. 2. Bergold, G.H.,T.H. Graf, and K. Munz. 1969.
Struc-tural differences among arboviruses, p. 41-49. In V. Bard6s (ed.), Arboviruses of the California complex and Bunyamwera group. Publishing House of the SlovakAcademyofScience, Bratislava.
3. Britten, R. J., and R. B. Roberts. 1960.High-resolution density gradient sedimentation analysis. Science 131:32-33.
4. Brummer-Korvenkontio, M. 1969. Arboviruses of the CaliforniacomplexandBunyamwera groupinFinland, p. 131-134. In V. Bard6s (ed.), Arboviruses of the California complex and Bunyamwera group. Publish-ingHouse of the Slovak Academy ofScience, Bratis-lava.
5. Burge, B. W., and A. S. Huang. 1970. Comparison of membrane glycopeptides of Sindbis virus and vesicu-lar stomatitis virus. J. Virol. 6:176-182.
6. Casals, J., and L. Whitman. 1961. Group C, a new serological group of hitherto undescribed arthropod-borne viruses. Immunological studies. Amer.J.Trop. Med. Hyg. 10:250-258.
7. Causey, 0. R., C. E. Causey,0. M. Maroja, and D. G. Macedo. 1961. The isolation of arthropod-borne vi-ruses, including members of two hitherto undescribed serological groups, in the Amazon region of Brazil. Amer.J.Trop. Med. Hyg. 101:227-249.
8. Dalrymple, J. M. 1972. Biochemical and biophysical characteristics of Venezuelanequineencephalitisvirus. p.56-64.InVenezuelan Encephalitis. Scientific Publi-cation No. 243, Pan American Health Organization, Washington, D.C.
9. Dalrymple,J. M.,A.Y.Teramoto, R. D. Cardiff, andP. K.Russell. 1972. Radioimmune precipitation of Group Aarboviruses. J. Immunol. 109:426-433.
10. Holmes, I. A. 1971. Morphological similarity of Bunyam-werasupergroup viruses. Virology 43:708-712. 11. Karabatsos, N. 1973. Physical properties and antigenic
components of Oriboca virus. Infect. Immunity 8:53-62.
12. Kennedy, S. I. T., and D. C. Burke. 1972. Studies on the structural proteins of Semliki Forest virus. J. Gen. Virol. 14:87-98.
13. Maizel, J. W. 1969. Acrylamide gel electrophoresis of proteins and nucleic acids, p. 35-48. In K. Habel and N. P. Salzman, (ed.), Fundamental techniques of virology.Academic Press Inc., New York.
14. McLarran, C. J., and R. A. Arlinghaus. 1973. Structural componentsof avirus of the California encephalitis complex: LaCrosse virus. Virology 53:247-257. 15. Murphy, F. A., A. K. Harrison, and T. Tzianabos. 1968.
Electron microscopic observations of mouse brain in-fectedwith Bunyamwera grouparboviruses. J. Virol. 2:1315-1325.
16. Murphy, F. A., S. G. Whitefield, P. H. Coleman, C. H. Calisher,E. R.Rabin,A.B.Jenson, J. L.Melnick, M.
on November 10, 2019 by guest
http://jvi.asm.org/
ORIBOCAVIRUS STRUCTURAL COMPONENTS R. Edwards, and E. Whitney. 1968. California group
arboviruses: electron microscopic studies. Exp. Mol. Pathol. 9:44-56.
17. Shapiro, D., W. E. Brandt, R. D. Cardiff, and P. K. Russell. 1971. The proteins of Japanese encephalitis virus. Virology44:108-124.
18. Shapiro,D.,D. Trent. W. E. Brandt, and P. K. Russell. 1972.ComparisonofthevirionpolypeptidesofgroupB arboviruses. Infect.Immunity6:206-209.
19. Shope, R. E., and0.R. Causey. 1962.Furtherstudieson
the serological relationships of group C
arthropod-borne viruses and theapplicationof theserelationships
torapid identification oftypes. Amer.J. Trop. Med. Hyg. 11:283-290.
20. Simons, K., andL.Kaariainen.1970.Characterizationof
the Semliki Forest virus coreand envelope proteins.
Biochem.Biophys. Res. Commun. 38:981-988. 21. Strauss, J. H., Jr., B. W.Burge, E. R. Pfefferkorn,andJ.
E. Darnell, Jr. 1968. Identificationofthe membrane proteinand"core"protein ofSindbis virus. Proc. Nat.
Acad. Sci. U.S.A.59:533-537.
22. Westaway, E. G. 1973. Proteins specified by Group B Togaviruses in mammalian cells during productive
infections.Virology 51:454-465.
23. Wildy, P. 1971. Classification andnomenclature of
vi-ruses.InJ. L. Melnick(ed.), Monographsinvirology. vol.5.S. Karger AG,Basel,Switzerland.
24. World Health Organization Scientific Group on Ar-boviruses. 1967. Arboviruses and human disease. WHO Tech. Rep. Ser.369:1-84.
VOL.13,1974