Copyright©1976 American Society for Microbiology Printed in USA.
Hin D
Restriction Mapping of Unpaired Regions
in
Simian
Virus 40 Superhelical DNA I: Considerations
Regarding
Structure-Function
Relationships
M. CHEN, J. LEBOWITZ,'* AND N. P. SALZMAN
Laboratory ofBiology of Viruses, National Institute ofAllergy and Infectious Diseases, National Institutes of Health, Bethesda, Maryland 20014, and Department of Microbiology, The Medical Center, University of
Alabama in Birmingham, Birmingham, Alabama 35294*
Received for publication 29 September 1975
Superhelical simian virus 40(SV40) DNA Iwasreactedwith N-cyclohexyl-N'-8-(4-methylmorpholinium)ethylcarbodiimide (CMC), and the location ofCMC
sites was mapped using the Hin D restriction endonuclease. The use of 14C0
labeledCMC allowsaquantitativeanalysisof thebindingto therespective Hin Drestriction endonuclease fragments. The percentage ofreactivity was 6.54%
for fragment A, 3.87% for fragment B, and 2.74% for fragment G. No CMC
radioactivity was detected in other fragments. This reactivity is in agreement
with the evaluation of binding by buoyantdensity measurements. The above
fragmentsalso contain the sitessusceptibleto
S,
endonucleaseaction.Thisaddsfurther support to the view that superhelical DNA can contain regions of
localized interrupted secondary structure which may be capable of forming
intrastrand hairpin structures if sequence relationships are favorable. The
possible structure-function relationships for this model arediscussed with the
emphasisontranscription.
In the previous paper (15) we showed that
superhelical simian virus 40(SV40)DNA I can
be modified with
N-cyclohexyl-N'-j8-(4-methyl-morpholinium)ethylcarbodiimide (CMG). CMCreacts preferentially with the imino sites of
unpaired thymineand guanine residues toform
a stable covalent product in the neutral pH
range (19, 20). Consequently, one can map the
locationsofCMConSV40DNA I. Thisinvolves
locating those Hin D restriction endonuclease
fragmentscontaining CMC. A comparison can
then be madebetween the CMCreactive sites
and those sites that are cleaved by
single-strand-specific S, endonucleaseaction.
If a correspondence exists between the re-gions ofreactivity of CMC,
single-strand-spe-cificendonucleases (3, 15),andgene 32 protein
(21, 22), this would addfurther support to the
view that superhelical DNA can contain
re-gions of localized interrupted secondary
struc-ture which may be capable of forming
intras-trand hairpin structures. An examination of
possible structure-function relationships for the
model proposed arediscussedinthe contextof
recentstudies, particularly transcription data.
MATERIALS AND METHODS
Cell culture, SV40, and DNA preparation. Full details for the above procedures have been described
IThis research wasconducted while on sabbatical leave
fromSyracuse University, Syracuse, N.Y. 13210.
previously (15).
Enzymes. The endonuclease R-Hin D was
puri-fiedby the method of Smith and Wilcox (25) andwas
storedat -20Cin0.2 MKCI-0.01 MTris(pH
7.5)-50%glycerol at 6 U/ml.
Chemicals.Cold CMCwasordered fromthe
Ald-rich Chemical Co., Milwaukee, Wis.; 14C-labeled
CMC was prepared by the New England Nuclear
Corp. (Boston, Mass.)custom synthesis laboratory
using1-cyclohexyl-3-[2-morpholino-(4)-ethyl]
carbo-diimide thatwasprepared byusfollowing previous
procedures (19). This intermediate carbodiimide
wasconverted to CMC by reaction with 14C-labeled
CH3Iinthe NewEnglandNuclearCorp.laboratory.
Electrophoresis. All of the electrophoretic runs
werecarried outinanECvertical-gel
electrophore-sis apparatus (model E-C 470). Samples (0.05 ml)
were applied to slab gels (17 by 12 by 0.30 cm)
consisting of 5%polyacrylamide and 0.5% agarose.
Electrophoresis was carried out for16h at20Cat 50
V ina buffer consisting of 40 mM
Tris-hydrochlo-ride, 20 mM sodium acetate, and 1 mM EDTA
ad-justed to pH 7.2 with acetic acid. To quantitate
radiolabeled bands, the gels were frozen and sliced
into 1-mm segments.Each segment was dissolvedin
0.2ml of30% H202 and counted with Triton X-100
toluene scintillation fluid.
Analysis of the binding of "4C-labeled CMC to
SV40 DNA. A defined amountofSV40 DNA I was
placedinthegel slab afterHin D digestion. Since
the weight fraction of eachfragmentisknown, the
calculation ofbindingwasperformed, e.g.,for
frag-mentA,asfollows. The Afragment is 22.5%of the
SV40 genome and consequently the gel contained
211
on November 10, 2019 by guest
http://jvi.asm.org/
212 CHEN ET AL.
2.25
jLgg
of A. Since only T and G are capable ofreacting we have 1.125,igor 0.00338 ,umol of
possi-ble bases as potential covalent binding sites. The
amount of"4C-labeledCMC bound afterbackground
subtraction was 292 counts/min, which was
con-verted into disintegrations per minute per
micro-mole using the specific activity for CMC, 2 mCi/
mmol, and the determined counting efficiency of
30%. In the case of the Afragment, 0.000212Mmol of
CMC wasbound, giving a reactivity of 6.54%.
Simi-lar calculations were performed for the B and G
fragments.
RESULTS
Electrophoretic analysis of a Hin D digest
ofSV40DNA Imodified with unlabeled CMC.
The first experiment utilized 3H-labeled SV40
DNA Itreated with unlabeled CMC (15). The
Hin Ddigestionwascarriedout inthe presence
of untreated "4C-labeled SV40 DNA after
re-moval of excess CMC. Figure 1shows that the
20[ | IX
16
o12
-a.
0U
X
8~
electrophoretic mobilities of the Aand B
frag-ments are clearly slowed by onefraction.There
is an indication that the G fragment is also
displaced by the CMC reaction. To confirm
these assignments, it was necessary to obtain
labeled CMC. This was provided to us as
14C-labeled CMC by custom synthesis from the
NewEngland Nuclear Corp. (see Materials and
Methods).
Electrophoretic analysis of theHinDdigest
of 3H-labeled SV40 DNA Imodified with
14C-labeledCMC. Thereactionof"4C-labeledCMC
was carried out under conditions similar to
those that wereused for the reaction between
cold reagent and 3H-labeled SV40 DNA. After theexcess reagent wasremoved, the 3H-labeled
DNA,with covalently bound"4C-labeledCMC,
was digested in the presence of 32P-labeled
SV40 DNA. The results of thistriple-label
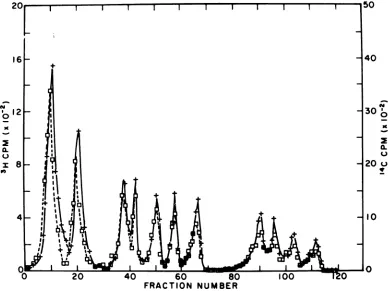
ex-perimentareshowninFig.2. Itcanreadilybe
X | | W l l 50
40
30;
0~
a.-U
20 u
-0 20 40 60 80 100 1
-FRACTION NUMBER
FIG. 1. 3H-labeledSV40 DNA I (specific activity, 5300 counts/min per
pg)
wasreacted with an800-foldexcessof cold carbodiimide (moles per mole of nucleotide) in a buffer containing 0.01 MNa2B4O7, 0.1 M
Na2SO4,0.25M NaCl, pH 8.0. The excess carbodiimide was removed by dialysis against the same buffer. The
carbodiimide-reacted 3H-labeled(0) SV40 DNA Iwasdialyzed against6.6mMTris, 50 mMNaCl(pH 7.4)
before being cleaved with endonuclease Hin D. "4C-labeled(+)SV40 DNA I markerdigest(specific activity
16,000 counts/min perMg)wascleavedsimultaneouslyinthesameHin Dbefore electrophoresisin 5%
acryl-amide-0.5% agarosein anECvertical-gel electrophoresisapparatusfor16h. Gelswerethenslicedinto 1-mm
segmentsand dissolvedin 0.3ml 30%H202 overnightat65C. Ten millilitersofscintillationfluidwasadded
and counted.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.509.56.445.280.571.2]. I i
0K
0. u
xI
6T
S-2
4x
25
20
Is
0
m
10 gona
5~ 5
10
FRACTION NUMBER
FIG. 2. The experimentwasessentially thesame asinFig.1,except'4C-labeled carbodiimide(A) (2mCi/
mmol) wasreactedwith3H-labeled (l) SV40DNAI.32P-labeled SV40DNAI(0) markerwascleavedinthe
sameHinDdigestwith 14C-labeledCMC-modified3H-labeledSV40DNA.
seen that no 14C counts are observed (closed
triangles)inanyofthefragments, exceptA,B,
and G. It is also apparentthat the 14C counts
follow the 3H counts in those fragments that
reactwith CMC. Itcanbe concluded fromthis
experiment thatA,B,and Garetheonly Hin D
fragments thatreact with CMC. It ispossible
fromthespecificactivityof14C-labeled CMCto
calculate theamountofbindinginthe
respec-tivefragments.The totalreactivitycanbe
com-paredtothebindingcalculated from the buoy-antshift measured in theprecedingpaper (15).
Binding analysis of[14C]CMCtotheHin D
A,B,and GfragmentsofSV40DNA. Basedon
thespecificactivity of"4C-labeled CMCand the
amountofSV40 DNA used intheexperiment,
we cancalculate theamount ofbindingtothe
respectivefragments. Theresultsarepresented
in Table 1. The tabulated reactivity for each
fragmentcanbe readilyconverted intoatotal
valueforthe entire SV40genomeby
multiply-ing the percentage of reactivity by the
frac-tional molecular weightofeach fragment and summing the total. This value is compared to
thevalue obtained from thepreviousstudy (15)
inwhichweemployedthebuoyant densityshift
produced by CMCand the partial specific
vol-TABLE 1. Percentage of reactivity of CMC to respective Hin D fragments and the number of CMC
molecules bound perfragment
HinD fragment No. of
and relative mol Reactivity with "'C-la- CMC wt(%ofSV40 beled CMC(%) mol
DNA) bound
A 22.5 6.54 80
B 15.0 3.87 31
G 7.0 2.74 10
Totals in terms of 2.24% of SV40 DNA 121 base pairs
Analysis bybuoyant 2.0%ofSV40 DNA 108 density shift and
densityof CMC from previous study(15)
ume of the reagent. The reaction conditions
were very similar except for the different
sourcesofCMC. Itcanbeseenthat the results
are in good agreementusingtwo independent
methods.
DISCUSSION
The results of the CMC bindinganalysis
us-ing theHin D restrictionendonuclease can be
best examined in thecontextofpreviousresults
on themapping of the single-strand-specific S,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.509.62.453.56.339.2] [image:3.509.260.455.390.538.2]214 CHEN ET AL.
endonuclease (3),gene 32 protein, andthe
cur-rentresults on thespecificity of theinitiationof
transcription of Escherichia coli polymerase
(14, 16,30).
Figure 3 summarizes thedata in theHin D
fragment map (6). It is apparent thatthere is
excellent agreement between the binding of
CMC tofragmentsA,B, andGand thesites cut
byS1 endonuclease onSV40 DNA I.
Inthe light ofthe aboveresults, we can
con-sider the possible structure-function
relation-ships for the sensitive regions ofsuperhelical
SV40DNA.The firstquestionthatisof
particu-larsignificance is thestructure ofthe regions containing unpaired bases in superhelical
DNA. The previous two papers (15, 29) have
presented evidencethatcanbeinterpreted with
themodelthatsupercoilingproducesregionsof
interruptedsecondarystructure. One can
envi-sion that different single-strand regions,
pro-duced by superhelical torsional forces, could
migrateuntiltheyreachedregionswhere
suffi-cientintrastrandcomplementarity, i.e., forma-tion ofhairpin structures atthese sites, would
lower the free energy and localize the
inter-rupted duplex structure. This localization of
interrupted secondary structure is dependent
onsequencerelationships. Thus, SV40 and
pol-yoma superhelicalDNAsappearto have a
lim-itednumberoflocalizedsitesfor
S,
cleavage(3,8) whereas 4X-RF is cleaved by Neurospora
crassa endonucleaseinanonpreferential man-ner (K. Bartok and D. T. Denhardt, personal communication).
We suggest that some superhelical DNAs will contain localized hairpin regions which 0
.8 D .2
-~~~~~~~~~
CMICBOUND
.7
\t.5 3/ E. coli RNA
Polymerase
Transcription
Early in vivo
Transcription
FIG. 3. Map of theHin DfragmentsofSV40DNA with the location of the following sites: dark regions
represent the area sensitive to the single-strand endonuclease S1 (3); the Hin D fragments binding the indicated number of CMC molecules determined in this study. Gene 32 protein binding sites (22, 23) are represented by arrows inside the circle. Three pppAp initiation sites determined according to the methods ofP.
Lebowitz and Zainet al. (15, 17, 30; manuscript in preparation) used by E. coli polymerase (see text) are
shown by the outside arrows with the direction of transcription. Early in vivo transcription is shown for
comparison (13).
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.509.114.396.259.585.2]may show fluctuations from the most stable
state (hairpin configurations). This introduces
amoredynamic view which allows for the
in-fluence of environmental factors. Beard et al.
(3)did observe that
S,
cuttingshiftedpreferen-tiallyto 0.45and 0.55 sites upon an increase in
NaCl concentration from 75 to 250 mM. The
additional consideration ofsequence
relation-ships suggest that some superhelical DNAs
could have many transient intrastrand
hair-pins or single-strand sites and, consequently, many sitesforsingle-strand nuclease attack. A
testfor thisview would be the precise locations
of CMC in order to sequence the regions
re-acted. Suchastudy is inprogess.
Sincegene 32proteinbinds cooperativelyto a
single-strand DNA (26), it is striking that it
binds only at one site, at 0.46, which
corre-sponds to one of the three sites at which
S,
cleaves SV40 DNA, whereas itfailstobind to
the B and G sites which are susceptible to
S,
cleavage. Inaddition, another gene 32protein
sitehas beendiscovered that doesnot mapwith
S,
orCMCsites(22).It has beenpointedout(26)thatthefreeenergyof binding ofaproteinto a
single strand ofDNA
(AGbind),
in aregionnor-mally double stranded under the prevailing
en-vironmentalconditions,must atleast offset the
free energy favoring the native over the
ran-dom coil conformation at that locus (AGCOflf).
These energetic considerations are applicable
tothemeltingoutof A-T-richregionsinduplex
DNA, aswell asthe meltingoutof short
hair-pinregions.Bothevents areneeded during
rep-lication(13, 26).
Inthecaseof SV40DNAI,itwould be
neces-sarythat
AGCOff
orhairpins<AGbhld
inordertomelt out the proposed hairpin structures. The
inabilitytosatisfy favorableenergeticsforgene
32proteinmelting of the0.55 site in Aand the
sites in B and G fragments suggests stable
hairpinsitesatthese locationsinSV40 DNA I.
Other structural featuresmay play an
impor-tantrole. The cooperativity ofhairpin melting
might bepartially controlled byprotein-protein
interactions,asboundgene 32proteinsinteract
favorably with oneanother (26). Steric factors
insupercoiled DNA could affect the binding of
gene 32protein.
A variety of in vitro transcription studies
have beenperformed on different superhelical
DNAs(4, 5, 11, 14, 17, 18, 23, 24, 27, 28, 30, 31).
Hayashi andHayashi(10)showed that UX-RFI
ismorerapidly transcribedthan
4X-RF
II. Thiskindof resulthasbeenobtained for
superheli-cal X (4) and PM2 DNA (23, 24, 27, 31). Itwas
proposed that supercoiling increased
nonspe-cific initiation(4, 27),sinceanincrease in
bind-ing is morefavorable inordertolower the free
energy by a coupled unwinding ofthe duplex
structure and supercoils. However, ifhairpin
regions exist,then binding ofRNApolymerase
could occur at these sites. Thiswould present
easily recognizable, specific regions for the
en-zyme aswell as promote more favorable
ener-geticsofbinding.
A number ofstudies (11, 14, 17, 18, 24) sug-gest that the above model may reflect the
en-hanced transcription seen for superhelical
DNA. Richardson (24) found that PM2 DNA I
forms 16 stable binary complexes with E. coli
RNApolymerase, whereasonly 2 are foundfor
closed, nonsupercoiledPM2 DNA. The reaction
of PM2 DNA Imodified with CMC,atabinding
ratio of0.01, eliminates 95% of the
transcrip-tion capacity relative to untreated form I (M.
Flashner and J. Lebowitz, unpublished data).
This hasalso been found for SV40 DNA I (P.
Haleand J. Lebowitz, unpublished data).
Recent studies(14, 16, 30) of thetranscription
of SV40 DNA with E. coli RNA polymerase
show that therearethreeprincipal y-[32P]ATP
initiation fragments: one, designated
A,,,
cop-ied fromthe Hin D-G fragment, and the other
twodesignated
A,
andA,,,,
each copiedfromtwoseparatesitesinthe Hin-A fragment(16, 30; P.
Lebowitz, unpublished data). It was also
re-ported thattwoadditionalpppAp andalimited
number ofpppGp sites (14) exist.
InFigure 3weshow the three pppAp
initia-tionsites onthe SV40map. Itcanbeseenthat
initiation occurs at 0.52and0.44 intheA
frag-ment,andat 0.17 inthe G fragment. The
direc-tionofE.coli RNApolymerasetranscription is
showninFig. 3,anditiscompletely
asymmet-ric (28). The strand copied corresponds to that
copied early in lytic infection and has been
designatedthe (-) orearly strand (12). An ex-aminationofFig. 3 shows anextremely
inter-esting structural feature: there is a potential
hairpin site beforeeach pppAp initiation site.
Consequently, hairpin regions in these cases
would promote considerable specificity in the
interactionofE.colipolymerase with
superhel-ical SV40DNA. The formation of RNA
polym-erase promoter complexes would be achieved
morereadilyathairpinsites, sincetheenergy
neededtoopentheDNAforaccess tothe
tem-plate bases should be less than that forafully
paired structure (5).Hence SV40 DNAIcould,
inthepppApinitiationsitesshown, bypassthe
activation step of opening an intact duplex
strand. This would allow for more initiation
events, which would explain the enhanced
transcription ofsupercoiledDNA.The
specific-ity of the RNApolymeraseinteractionwouldbe
dependentonwhetheropenregionsorhairpins
canbefixedonthe molecule. Further
on November 10, 2019 by guest
http://jvi.asm.org/
216 CHEN ET AL.
ingbythe formationofaDNA-RNA hybrid (14)
would be assisted by the free energy decrease
caused by acoupled unwinding of duplex and
superhelicalturns.
Recently, a series ofpapers was published
on the in vitro transcription of SV40 DNA I,
relative to the linear form, usingcalf thymus
and rat liver RNA polymerases (11, 17, 18).
These results are very similar to the
OX-RF
and PM2dataandshowenhanced transcription
for SV40 DNA I. They conclude that unpaired
basesareresponsible for theenhancedratedue
to an increased binding affinity for the RNA
polymeraseatunpairedsites(11).In contrastto
E. coli polymerase, the mammalian RNA
po-lymerasestestedtranscribed SV40 DNA I
sym-metrically. The incorporation of -y-[32P]ATP
and y-[32PIGTP suggests multiple initiation
sites,although thenumberand locationare not
known (17). It is important to note that an
initiationcomplexof theBRNApolymerase of calfthymuswith SV40 DNA I apparently pro-tectsagainstS1 cleavage (11).
An in vivocomparisonof SV40 mRNAinthe
nucleus and thecytoplasmsuggeststhat RNA
synthesis occurssymmetrically and that
anti-message isdegraded priortoitstransport tothe
cytoplasm(1,12).Thisproposalagreeswiththe
symmetrical synthesis observed in vitro for
mammalian RNA polymerases. The
hybridiza-tionpatternof cytoplasmic mRNA withHin D
fragments reveals that early message starts
almost attheend of the A fragment (12) (Fig.
3). It is difficult at this time to reach
conclu-sionssolely from thehybridizationdata. First,
wedonotknow thedetails of processing of viral
mRNA and second, weknow verylittle about
the transcription of SV40 DNA complexes with
histones(7, 9).It isentirely possible that
struc-turalvariationwill beproduced by the
forma-tionof nucleosomes (7, 9). For example, astudy
of the mapping of the SV40-specific sequences
transcribed in vitro from chromatin of
SV40-transformed SV 3T3 cells shows that the early
regions A throughB (Fig. 3) were transcribed
fiveto ten timesmore frequently than the
re-maining regions (2). In contrast, transcription
ofpurifiedSV3T3 DNAbyE. colipolymerase
produced equal frequencies of transcription fromall regionsof theintegratedDNA (2).
Thisbriefsurveyofaspectsoftranscription of
SV40clearly suggests thatfurther
experimen-tal exploration is required in order to
under-stand the structural relationship involved in
transcriptional events for SV40 DNA I. We,
alongwith other (3, 10, 11, 17, 18),suggestthat
there maybe an important role played by
re-gionsofinterruptedsecondary structure
form-ingpossiblelocalized hairpinsinthe
transcrip-tionalprocess.
ACKNOWLEDGMENTS
J. L. wassupported by Public HealthService Research Career AwardK04-CA00141-03 (formerly5K04-CA07514) from theNational Cancer Institute throughout theconduct and preparation ofthisstudy.J. L.thanksSyracuse Uni-versity for thenecessarysabbaticalleave to carry out the reportedresearch.
LITERATURE CITED
1. Aloni, Y. 1974. Biogenesisand characterization of SV40 and polyoma RNAs in productively infected cells. ColdSpringHarborSymp. Quant. Biol.39:165-177. 2. Astrin,S. M. 1975. Mapping of theSV40specific
se-quencestranscribedin vitrofromchromatinofSV40 transformedcells.Biochemistry 14:2700-2711. 3. Beard,P., J. F. Morrow,andP.Berg.1973.Cleavageof
circular, superhelical SV40 DNA toa linear-duplex byS,nuclease.J.Virol.12:1303-1313.
4. Botchan, P., J. Wang,andH.Echols.1973. Effect of circularity and superhelicity ontranscription from bacteriophageXDNA. Proc.Natl. Acad.Sci. U.S.A. 70:3077-3081.
5. Chamberlin, M. J. 1974.Theselectivity of transcrip-tion. Annu.Rev. Biochem. 43:721-775.
6. Danna, K.J.,G.H. Sack,Jr.,andD.Nathans. 1973. Studies ofSV40 DNA VII. Acleavage map ofthe SV40genome. J. Mol.Biol.78:363-376.
7. Germond, J.-E.,B.Hirt,P.Oudet,M.Gross-Bellard, andP. Chambon. 1975.Folding of the DNAdouble helix inchromatin-likestructures from simian virus 40.Proc.Natl. Acad.Sci.U.S.A.72:1843-1847. 8. Germond,J.-E., V. M. Vogt, and B. Hirt. 1974.
Charac-terizationof thesingle-strand-specificnuclease S, ac-tivity ondouble-stranded supercoiledpolyoma DNA. Eur. J.Biochem.43:591-600.
9. Griffith,J. D. 1975. Chromatin structure:deducedfrom aminichromosome. Science187:1202-1203.
10. Hayashi,Y., and M. Hayashi.1971. Template activities ofthe4bX-174replicative, allomorphic deoxyribonu-cleic acids. Biochemistry10:4212-4218.
11. Hossenlopp, P., P. Oudet, and P. Chambon. 1974. Ani-mal DNA-dependent RNApolymerases. Studies on thebindingofmammalianRNA polymerasesAland B to simianvirus 40DNA. Eur. J.Biochem. 41:397-411.
12. Khoury, G., P. Howley, D. Nathans, and M. Martin. 1975.Posttranscriptionalselection of simian virus 40-specificRNA. J. Virol. 15:433437.
13. Kornberg,A. 1974. DNA synthesis, p. 203-207. W. H. Freeman &Co., San Francisco.
14. Lebowitz, P., and R.Bloodgood.1975.Transcription of simian virus 40DNAby EscherichiacoliRNA polym-erase:synthesis of a DNA-RNA hybrid and discrete RNAsunder restrictive transcription conditions. J. Mol.Biol.94:183-201.
15. Lebowitz, J., C. F.Garon, M. Chen, and N. P. Salz-man.1976. Chemical modification ofsimian virus 40 DNA with awater-soluble carbodiimide. J. Virol. 18:205-210.
16. Lebowitz, P., W. Seigel, and J. Sklar. 1974. Hemophi-lusaegyptius restrictionendonucleasescleavage map ofthe simian virus 40genome and its colinear
rela-tionwith theHemophilus influenzae cleavage map of
SV40. J. Mol. Biol.88:105-124.
17. Mandel, J. L., and P.Chambon. 1974. Animal DNA-dependentRNApolymerases.Analysis of the RNAs --- ' I nsimianvirus 40superhelical DNA by
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
mammalian RNA polymerases Al and B. Eur. J. Biochem. 41:379-395.
18. Mandel, J. L., and P. Chambon. 1974.Animal DNA-dependent RNA polymerases.Studies on the reaction parametersof transcription in vitro of simian virus 40 DNA by mammalian RNA polymerases Al and B. Eur. J. Biochem.41:367-378.
19. Metz, D. H., and G. L.Brown. 1969. The investigation of nucleic acid secondary structure by means of chem-icalmodification with a carbodiimide reagent. I. The reaction between N-cyclohexyl-N'-P-(4-methyl mor-pholinium)ethylcarbodiimide and model nucleotides. Biochemistry 6:2312-2328.
20. Metz, D. H., and G. L.Brown. 1969. The investigation of nucleic acidsecondarystructurebymeansof chem-icalmodificationwith acarbodiimidereagent.II. The reaction between N-cyclohexyl-N'-P-(4-methylmor-pholinium) ethylcarbodiimide and transfer ribonu-cleic acid.Biochemistry 8:2329-2341.
21. Morrow, J. F., and P. Berg. 1973.Location of the T4 gene 32 proteinbindingsiteon simian virus 40 DNA. J.Virol. 12:1631-1632.
22. Reed, S. I., J. Ferguson, R. W.Davis, and G. R. Stark. 1975. T antigenbindstosimian virus 40 DNA at the origin ofreplication. Proc. Natl. Acad. Sci. U.S.A. 72:1605-1609.
23. Richardson, J. P. 1974.Effectsof supercoiling on tran-scription from bacteriophage PM2deoxyribonucleic acid.Biochemistry13:3164-3169.
24. Richardson, J. P. 1975. Initiation of transcriptionby
Escherichia coli RNA polymerase from supercoiled and non-supercoiled bacteriophage PM2 DNA. J. Mol. Biol. 91:477-487.
25. Smith, H. O., and K. W. Wilcox. 1970. A restriction enzyme from Hemophilus influenzae. I. Purification and general properties. J. Mol. Biol. 51:379-391.
26. von Hippel, P. H., and J. D. McGhee. 1972.
DNA-protein interactions. Annu. Rev. Biochem. 41:231-300.
27. Wang, J. C. 1974. Interactions between twisted DNAs andenzymes: theeffectsofsuperhelicalturns. J.Mol. Biol. 87:797-815.
28. Westphal, H. 1970. SV40 DNA strand selection byEsch-erichia coli RNA polymerase. J. Mol. Biol. 50:407-420.
29. Woodworth-Gutai, M., andJ. Lebowitz. 1976. Intro-duction of interruptedsecondarystructure in super-coiled DNA as a functionofsuperhelix density: con-sideration of hairpin structures insuperhelicalDNA. J. Virol. 18:195-204.
30. Zain, B. S., R. Dhar, S. M. Weissman, P. Lebowitz, and A. M.Lewis.1972.Preferredsiteforinitiationof RNA transcriptionby Escherichia coli RNA polymer-ase within the simian virus 40 DNA segmentofthe nondefective adenovirus-simian virus 40hybrid vi-rusesAD2+ND,and Ad2+ND3.J. Virol. 11:682-693. 31. Zimmer,S. G., and R. L.Millette.1975.
DNA-depend-ent RNA polymerase fromPseudomonasBal-31. II. Transcription of theallomorphic forms of bacterio-phagePM2DNA. Biochemistry14:300-306.
on November 10, 2019 by guest
http://jvi.asm.org/