0022-538X/92/127469-12$02.00/0
Copyright© 1992, American Society for Microbiology
Human
Immunodeficiency Virus Type 1 Rev Activation Can
Be
Achieved without Rev-Responsive Element RNA if Rev Is
Directed
to
the Target as a Rev/MS2 Fusion Protein Which
Tethers
the MS2 Operator RNA
SUNDARARAJAN VENKATESAN,1* SUSAN M. GERSTBERGER,1HEESUNG
PARK,'
STEVEN M. HOLLAND,2ANDYONG-SUK
NAM'Laboratoty of Molecular
Microbiology'
andLaboratory of Host Defenses,2 National Institute ofAllergy
and Infectious Diseases, Bethesda, Maryland 20892Received16July1992/Accepted 9 September 1992
The posttranscriptionaltinnsactivation ofunspliced or partially spliced human immunodeficiency virus RNAs by the Rev regulatory proteiniscrucial forvirusreplication and is dependent on sequence-specific RNA binding by Rev. The cognate RNA target of Rev is contained within a
highly
structured, 244-nucleotide Rev-responsive element(RRE) RNA in the viral env gene. Here, we show thatspecific interaction with the RRE is not an absolute requirement for Revfunction.When the RRE is replaced by aheterologousMS2 phage operator sequence, Rev willfacilitatethe cytoplasmic expression of human immunodeficiency virus mRNAs containing this sequence if directedtotheMS2 operator via the RNA binding motif of the MS2 phage coat protein (MS-C) as a Rev/MS-C fusionprotein. Rev/MS-Cefficiently
activated both RRE and MS2 targets. A mutation in the MS2 operator that abolishedthecoatprotein binding in vitro rendered the mutant RNA nonresponsive to the fusion protein in vivo. Notwithstanding that Rev canbe tethered to the viral RNAs via another RNA binding motif, the structuralintegrityof the N terminus of Revwasstillrequired for optimal trans activation. The life cycle of human immunodeficiency virus type 1
(HIV-1) includes several biochemical mechanisms that may also be general paradigms for eukaryotic gene regulation.
Sequence-specific interaction of viral RNAs with virus-encodedproteinsis acommon theme in manyfundamental
viral processes, such as RNA transcription and
posttran-scriptional activation and RNA encapsidation (13, 45).
HIV-1 transcription is activated by the viral Tat protein,
which recognizes a 22-nucleotide stem-loop nascent TAR RNA(7,9,21, 42,46).HIV Revactsposttranscriptionallyto modulate thesplicing (11, 30) and tofacilitate the extranu-clear transportand cytoplasmic utilization of viralmRNAs
by binding to a highly structured Rev-responsive element (RRE)sequencein theenvmRNA(15-17, 19, 20, 23,32, 35,
49). Rev isabasicprotein thatconcentrates inthe nucleoli (12, 14) and readily oligomerizes both in vitro and in vivo
(33, 36, 47, 50). Genetic studies have identified separate
nuclear localization-RNAbindingand activation domainsat the N- and C-terminalportions of the Revprotein,
respec-tively(31). Genetic analysesof theN-terminalregionof Rev have mapped a contiguous stretch of amino acid residues between positions 13 and 56 that confers RNA binding,
nucleolarlocalization,andoligomerization properties essen-tial forRevfunction
(5,
8, 27,33).Interestinthe
specific
sites of functional contact of Revwith the RRE has led to attempts to narrow the element down to a minimal
responsive
motif. The computer-pre-dictedsecondarystructureofRRE RNA(32)
iscomposed
offourstem-loops
(A, C,
D, aridE)
andabranchedstem-loop
(B/B1/B2) surrounding a central loop sequence
(Fig.
1).
Largeportionsofthis RREstructure aredispensablefor the Rev response (17, 22, 23,35),
and a 64-nucleotide subse-quence,predictedtofold into the branchedstem-loop B/B1/
*
Corresponding
author.B2, retains the Rev response both in vitro and in vivo (23,
35). A 9-nucleotide 5'-CACUAUGGG-3' sequence of the RRE corresponding to the B/B1 regions represents the minimal Revrecognitionsequence,both in vitro and invivo, if it ispresentedas astem-bulge-stemstructureandcontains atleasttwo G's,one of whichmustbeunpaired (24).
In spite of the accumulated evidence of the role of the RREas ananchor for Rev, thepossibilitythatthe RREalso recruits cellular factors that may be agonistsorantagonists of Rev function cannot be excluded. For instance, Rev cannotactivate the 5'-most132-nucleotide RREfragment in
vivo, despite having a greater in vitro affinity for this
fragment than the full RRE (23). It is possible that this
132-nucleotidefragmentlacksanessentialrecognitionmotif
for a putative cellular helper
factor(s).
In fact, a 56-kDaRRE-bindingcellular factor that mayassist in Rev function has been demonstrated previously (44). Also, both stably
and transiently expressed Rev functions poorly in murine cells, implying a requirementfor tissue- or
species-specific
Revhelperfactors(43).To explore the role of the RRE in the trans-activation
processes other than Revbinding and toexamine the con-tributions of thepeptidemotifs of Revtoactivation,wehave deviseda functional assayfor Revwithheterologous RNA targets. We show that the RRE may be
replaced by
theheterologous MS2 phage operator RNAwithout
losing
the Revresponse invivo if Rev is directedtothe operatoras a Rev/MS2coatprotein fusion.Using
bivalent targets recog-nized byboth Rev and the MS2 coatprotein,
we demon-stratethatthe in vivo responses of the targetsparallel
their in vitro affinities for therespective
RNAbinding
proteins.
These studies indicate that Revbinding
maybetheprimary
function of the RRE and thatRRE-binding
cellularfactors maynotbeanabsoluterequirement
for thetrans-activation effect of Rev.7469
on November 9, 2019 by guest
http://jvi.asm.org/
7470 VENKATESAN ET AL.
MATERIALS AND METHODS
gag expression plasmids. All of the recombinants were constructed
by
site-directedmutagenesis
of RRE DNA with a commercial M13-based protocol (Mutagene kit; Bio-RadLaboratories,
Richmond, Calif.), and the respectivemutant DNAs wereexchanged
for the RRE in the HIV-1 long terminal repeat(LTR)-linked
gagexpression
vectorcontain-ing
thewild-type
(wt)
RRE(23).
RREZ-MSwasconstructedby
firstdeleting
theRev-responsive B/B1/B2
subdomain(23) in the RRE(RRE
coordinates49 to113)andtheninsertinga modified version ofphage
MS2 translational operator se-quence(6).
RREZ-MS in M13 was further mutagenized to generateRREZ-MSAA,RREZ-MS(U/A),
and RREZ-MS(A/C),
which refer to the mutants which deleted the bulgedA residue(underlined
in the -GAGG-sequence) orsubstituted A forU(-AUIJA-)
orC for A(-AUUA-)
in theloopsequence of MS. Inaddition,
the RREwas alsocompletely exchangedforamonomeric MS sequence in thesense(MS)orantisense
(SM)
orientation and forahead-to-tail [(MS)(MS)] or head-to-head[(MS)(SM)]
dimer of the modified operator. (MS) wasconstructedby
inserting
themodified operatorflankedby
twoAresidues at the Asp-718(KpnI)
site ofM13mpl8,andtheMS monomerwas subsequently exchanged for the RRE in the wtgag RRE
plasmid.
To generate [(MS)(SM)]and
[(MS)(SM)],
the MS monomerwas recovered from the recombinant M13replicative
formby Asp-718 digestionandself-ligated.
TheMSdimer from this reactionwasrecoveredby polyacrylamide gel
electrophoresis and insertedinto the above gagexpression
plasmidinplaceof the RRE. (MS)+A wasconstructedby inserting
the RRE sequencecorrespond-ing
topositions
31 to49andpositions204to221, upstream anddownstream, respectively,
of the MS sequence in the M13 recombinant. In(MS)+a,
RRE sequence between po-sitions 39to49and 204to213flankedthe MS sequence. Thefollowing
two chimerasincorporating
the targets for both Rev and the MS coatprotein
were also engineered. (i) To construct(MS)+B/B1/B2,
theB/B1/B2
subdomain(positions49to
115)
of the RREwasinitially
clonedintoM13mp18.MS sequenceflankedby
three A'swastheninserted upstream of theB/B1/B2
motif.(ii)
M13 containingB/B1/B2
was alsomutagenized
toreplace
theBi
stem-loop(positions59to82) with the MS2 sequence. The resulting recombinant B/3G/MS2/B2
preserved the B stem and contained the RRErecognition
sequence -CACUAUGGG- upstream of the op-erator. Each ofthe bivalent chimeraswasindividuallymu-tagenized
tosubstitute the three G's in the RREBstemwith threeA's,
todelete thebulgedAresidue intheMS operator,ortodo both. All ofthe chimerasweretheninserted into the gag
expression plasmid
pRMK4RREinplace of the RRE.Expression plasmidsforRev/MS-C fusion proteins and their derivatives. HIV-1 Revwasexpressed either from the
Tat-responsive
HIV-1 LTR or from the constitutive Roussar-coma virus
(RSV)
LTR (29). MS-C and MS-N wereex-pressed
from thepSRa
promoter (6). Rev/MS-C denotes a tandem fusion of Rev and MS-C open reading frames(ORFs).
The fused gene was constructed by a two-steppolymerase
chain reaction(PCR)procedure. In the first step, the Rev ORF was PCR amplified with anXbaI restrictionsite-tagged
5'oligonucleotideprimer and a 3' primer that wastagged
with sequencescorresponding to codons 2 through 7 ofMS-C,
MS-D,orMS-N. TheMS2 coat protein expressionplasmids
were generously provided by Benjamin Berkhout of theUniversity
of Amsterdam, The Netherlands. The ORFs for the respective MS2 coat proteins were PCRamplified
with a 5' primer tagged with a sequencecorre-spondingtotheC-terminal 6codons of Rev and a3'
primer
tagged with anXbaI site. The PCR-amplified Rev and MS
DNAs were gel purified, annealed together, and
amplified
further with XbaI restrictionsite-tagged 5' primers comple-mentary to the codons at the N terminus of Rev and the C terminus ofMS,respectively. The resultingRev/MS
fusion genes werepurifiedfromgels, cleavedwithXbaI, and cloned into M13mp18. After the DNA sequence of theRev/MS-mpl8 recombinants was verified, the fusion genewas PCR
amplified byprimerpairs tagged with T7
phage
RNA poly-merase promoter and terminatorsequences at the 5' and 3' ends, respectively. The resulting T7 template was tran-scribed in vitrowith T7 RNApolymerase, and theRNAwas translated in a rabbit reticulocyte translation system with [35S]methionine. The translation yielded a fusionprotein
with the expected molecular mass that was readily im-munoprecipitable with anti-Rev antiserum. All other Rev/MS-Cderivativescarryingthe indicated mutationswere constructed by site-directed mutagenesis of Rev/MS-CM13mpl8 phage DNA according to a commercial protocol
(Mutagene kit, Bio-Rad Laboratories). All of the Rev/MS
chimeraswereclonedattheXbaI site ofanRSV LTR-linked
eukaryotic expression plasmid, pRSV.5 (29).
Escherichia coli expression of Rev/MS-C fusion proteins. Rev/MS-C fusion protein and the indicated mutants were expressed in E. coli as fusion proteins linked to the C
terminus of E. coli maltose-binding protein (MBP) from an isopropyl-,-D-thiogalactopyranoside (IPTG)-inducible
,-ga-lactosidase promoter with a commercial kit (New England Biolabs, Beverly,Mass.). Rev/MS-C andRev(LE/PE)/MS-C mutant ORFs were recovered from the respective RSVLTR-linked eukaryotic expression plasmids byXbaI diges-tion and cloned downstreamof the lone XbaI sitein the MBP vector. As a negative control, an Rev/MS-C coding sequence was alsoclonedin the antisense polarity with respect to the MBP ORF. Biosynthesis of the MBP-fused proteins was inducedby IPTG(1 mM) at an optical density at 600 nmof 0.3 followedby 2 hof incubation in at 37°C. Bacterialcells were harvested, rinsed with phosphate-buffered saline (PBS), and resuspended in lysis buffer containing 50 mM Tris-HCl (pH 7.8), 50 mM NaCl, 1 mM EDTA, 1 mM ethylene glycol-bis(,B-aminoethyl ether)-N,N,N',N'-tetra-acetic acid(EGTA), 1 mM dithiothreitol, and 1 mM phenyl-methylsulfonyl fluoride. Cells were disrupted with a French pressat 10,000lb/in2 and centrifuged at 25,000 x g for 15min
to collect the pellet containing the inclusion bodies. The pellet wascompletely dissolved in Tris buffer containing 50 mMTris-HCl, 50 mMKCI,and 5 mM EDTA supplemented with 6 M guanidine HCI and dialyzed successively against 200 volumes of the above buffer containing 1, 0.5, and 0.05 M guanidine HCI for 4 h at each step. Samples were centrifuged at 10,000 x g, and the supernatant was stored at -70°C or directly bound to amylose resin under conditions recommended by the manufacturer. The fusion protein was eluted with 20 mM maltose, concentrated by ultrafiltration, and used for binding studies.
RNA synthesis and protein binding. DNA templates, tagged attheir 5' ends with theT7promoter, were generated by PCR with primer pairs corresponding to the ends of the desired RNA (23). T7 promoter-tagged, PCR-amplified DNA fragments (0.5 pmol) were used as templates to generate unlabeled or uniformly labeled ([a-32P]UMP) RRE tran-scripts with a commercial kit from Promega Biotec (Milwau-kee, Wis.). All of the RNAs were purified by gel filtration on Sephadex G-100 columns and then electrophoresed on urea-acrylamide gel (10%), if necessary. RNA-protein binding J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
was evaluated by RNA gel mobility shift experiments. A reaction mixture containing protein samples and heparin (5 ,ug per reaction mixture) in HEPES binding buffer (20 mM HEPES [N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid]-KOH, pH 7.9, 62 mM KCI, 0.15 mM dithiothreitol, 6% glycerol) was preincubated at 30°C for 10 min before the labeled RNA (5 x
103
cpm) was added and further incubated for 10 min at 30°C. Samples were loaded onto 5% native polyacrylamide gel in 0.5x Tris-borate-EDTA andelectro-phoresed at30 mA at 4°C. Radioactivity was visualized by
autoradiography of dried gels. Rev used in these experi-ments was expressed in E. coli and purified to near
homo-geneity (47).
Transient expression assay.Although the results presented in this article were obtained with HeLa cells, all
transfec-tions were repeated four times with Cos-7 cells. Approxi-mately 2 x 106 HeLa orCos-7 cells were electroporated at 300 V and 250 ,iF in a Bio-Rad electroporator with the individual gag plasmids (5
jig)
and 2jigof pHIV-Tat(pSV40 TatforCos-7 cells) and with or without the indicated Rev or theRev/MScoatprotein fusion protein expression plasmids. For the expression of Rev, 2.5 or 5 ,ug of an HIV-1 LTR-linked (23) or an RSV LTR-linked plasmid was used. The Rev/MS fusion protein plasmids (5 jigperexperiment) were expressed from the RSV LTR. All transfections also included HIV-1 LTR-linkedchloramphenicol acetyltransfer-ase (CAT) (2 ,ug) plasmid to monitor the transfection effi-ciency. Aliquotsof transfected cellswereroutinely removed at 48 hand used forimmunodetection of Rev and Rev/MSfusion proteins (see below). Alternatively, aliquots of cells from each transfection were plated on coverslips
immedi-ately after electroporation and processed for immunofluo-rescenceof Rev and Rev/MSfusionproteins.Transfections were routinely terminated at 48 to 60 h, the cells were disrupted bythree freeze-thawcycles,and theextracts were clarified by low-speed centrifugation.gag
expression
was quantitated by a p24 enzyme-linked immunosorbent assay(ELISA) of cell extracts (Coulter Diagnostics).
p249a&
expression levels were normalized to constant values of CAT activity
expressed
from HIV LTR-linked CAT(23).
Each gag ELISA value from HeLa cell transfections repre-senteda meanfromatleast eight independenttransfections withmultiplepreparationsofplasmidDNA.CATexpression
was quantitated by a commercial CAT ELISA kit
(Boehr-ingerMannheim).
Immunoblotting. After electroporation, cells were
plated
on60-mm-diameter dishes and harvested 48to72 h later. A one-thirdequivalentof cells from each dishwascollected
by
centrifugation, rinsed twice in
PBS,
andlysed by
two freeze-thaw cycles in 50 ,ul of a buffercontaining
50 mMTris-HCl, pH 7.4, 0.5% Triton X-100, and 0.5%
3-[(3-cholamidopropyl)
-dimethyl
-ammoniol
-1-propanesulfonate.
Thecelllysateswere thenadjusted
to 100 jiMphenylmeth-ylsulfonyl fluoride-1
jig
ofaprotinin
perml-1jig
ofleupeptin
per ml and digested at37°C
for 15 min with a mixture ofpancreatic
RNase A(2
jig/ml)
and 10 U ofRQ
DNase(Promega
Biotec).
Thedigestion
wasstopped by
theadditionofanequalvolume ofabuffer
containing
50mMTris-HCl,
pH 7.4, 4% sodium
dodecyl
sulfate(SDS),
12%(wt/vol)
glycerol, and 5%(vol/vol)
2-mercaptoethanol
which washeated at95°Cfor 5 min.
Aliquots
wereelectrophoresed
on12.5%polyacrylamide gelsinSDS. The
proteins
were blot-ted to0.2-jim-pore-size
Immobilon filters(Millipore
Corp.)
and reacted first with rabbit
polyclonal
anti-Revantiserumormousemonoclonal antibodies
against
Revand thenwith theappropriatesecond
antibody
tagged
with horseradishperox-idase.Theimmunoreactive bandswerethen visualizedbya commercial chemiluminescence
protocol
(Amersham Corp.)
and
quantified by scanning.
Immunoprecipitation. HeLaorCos-7 cellswereroutinely
labeled 48 h posttransfection. For labeling, a one-third
equivalent
ofcells from each confluent 60-mm-diameter dish wasstarved for 15 min in RPMI mediumlackingmethionine andcysteine
andcontaining
2.5%dialyzed
fetal bovine serum. The cells were thenmetabolically
labeled with[35S]methionine
and[35S]cysteine (2 mCi/ml)
inatotal vol-umeof 0.1 ml for 1 h at37°C.The cellswereharvested andlysed as described above for immunoblotting, except that RNaseand DNasetreatments werein atotal volume of 0.2 ml.Followingthedigestion,thesamplesweredilutedwith 5 volumes of a buffer
containing
50 mMTris-HCl,
pH 7.4, 0.25% Triton X-100, 0.25%3-[(3-cholamidopropyl)-dimeth-yl-ammonio]-1-propanesulfonate,
0.15%bovine serumalbu-min,
60 mMNaCl,
100jiM
boundphenylmethylsulfonyl
fluoride,
1jig
ofaprotinin
perml,
and 1jig
ofleupeptin
per ml. Thelysates
werethenprecleared by being
boundat4°C
for 1 h toStaphylococcus protein
A beads that had beenpreloadedwith nonimmune
immunoglobulin
G. The super-natantsfrom this stepwere then bound toprotein
Abeads coated with rabbitpolyclonal
or mousemonoclonal anti-Rev antibodies for 2 hat4°C.
Theprotein
Abeadswerecollected bycentrifugation, rinsed,
andprocessed
forelectrophoresis
through
10%acrylamide gels
inSDS.Immunofluorescence microscopy.
Following
electropora-tion,
2 x104
cellswereplated
on7-mm-diametercoverslips
in 24-wellplates.
At48 to 72 hposttransfection,
themono-layers
wererinsed twice with PBS andfixedfor 15 min in4%paraformaldehyde
in PBS. The cellswererinsed three times with PBS andpermeabilized
with 1% Triton X-100 in PBS for 3 min.They
wererinsed four times in PBS and incubated witheitherrabbitpolyclonal
anti-Rev antiserum(1:200
dilu-tion)
oranti-Revmousemonoclonalantibody
(1:500 dilution)
in 0.2% bovine serum albumin in PBS for 2 to 4 h. After
being
washedextensively,
themonolayers
were reacted for 1 h with fluorescein-labeled goat anti-rabbit Fabfragment
(1:500)
ordonkey
anti-mouse antiserum(1:200)
in 0.2% bovineserumalbumin in PBS. After several washes inPBS,
the
coverslips
wererinsed in deionized waterand mounted in MOWIOL(Calbiochem
Corp.)
in 15%glycerol
supple-mented with5%(wtlvol)
DABCO(Sigma
ChemicalCo.)
to preventfading.
The cellswereexaminedby
regular
interfer-ence or Nomarskioptics
andby
dark-field immunofluores-cencewith aZeissAxiophot
microscope.
RESULTS
Rev/MS2
coatprotein
fusion activatesHIV-1 gagexpression
from a
hybrid
RNAlacking
theRev-responsive
domain. Toinvestigate
whether the RRE can bereplaced by
anotherRNA, the RRE sequence in the gag
expression
vectorpRMK4RRE
(23)
wasmutatedtoexchange
the64-nucleotideRev-responsive
domain for a modifiedMS2
phage
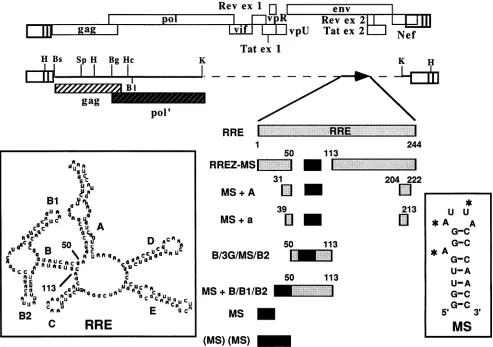
transla-tion operator sequence of 19nucleotides(Fig. 1,
RREZ-MS).
TheMS2 operator hadbeen modifiedtointroduce twoG-C base
pairs
to stabilize the stemregion
withoutovertly
affecting
the MS2 coatprotein
binding
(6).
To direct thebinding
of Rev to RREZ-MSRNA,
MS2
coatprotein
wasappended
to the C-terminal end of the Rev. Asingle-nucleotide deletion atthe seventh codon of the MS-C
ORF,
which introduced a frameshift and
early
termination of the coatprotein
(Rev/MS-D),
served as anegative
control. Anothermutantthatdeleted 48 residuesatthe Nterminus ofon November 9, 2019 by guest
http://jvi.asm.org/
7472 VENKATESAN ET AL.
Rev
ex
1
Nef
H Bs SpH BgHc K
I I
lIII
I.
gYag
I
pol
IRRE
A
B c a cu URU
a1uCa C G e u RA I 13
c
C RRE
1 244
50
RREZ-MS ...I-ZZ
31
MS+A
El
MS
+aB/3G/MS/B2
MS
+B/B1/B2
39
El
[image:4.612.62.555.71.418.2]113
.........
... ...
...
204 222
213 El
50 113
.... ...
....
50 113
m..zzi
MS
-(MS) -(MS)
FIG. 1. Schematicdiagram ofHIV-1 LTR-linkedgag expressionplasmids containing RRE and RRE-MSchimeric inserts. The basic
structureofthepHIVLTR-gag-RRE plasmid (previouslyreferredtoaspRMK4RRE)has been described before(23) and isdrawntoscale
under the linear genomicmapof the HIV-1provirusNL4-3(1).ThepolORF in theexpression plasmidwastruncatedattheIjpnI(Asp-718)
site and is denotedby pol'.Theletter codes refertoafew of the relevantrestriction sites. The 244-nucleotide RREsequencein thisvector,
indicatedby the lightly shaded rectangle,hadbeeninserted between the
YpnI
(Asp-718)site inthepolORF and theKpnIsite in thenefORF. Thepredicted secondarystructureof the RREis shown in the boxatleft, and theindividualstem-loopsarelabeled. The MSsequenceisdenotedby aheavilyshaded block with the modified MS RNAsequence (6)illustrated as astem-loopstructure in the boxat right. The
asterisksontheMSRNAstructurerefertothenucleotides critical for MS2phagecoatproteinbinding. TheMSmonomeranddimerare
denotedby singleand dualheavilyshaded boxes. Thestructuresof the different chimerasareindicatedbycombinations oflightlyandheavily
shadedboxes. The numbersbythese boxes refertothe RRE coordinates. These chimerasare morefullydescribed inMaterials and Methods.
Revnecessaryfornuclear-nucleolartargeting,RNAbinding, andoligomerizationwasalso constructed(A3-5ORev/MS-C).
Toensurethe nucleartargetingofA3-5ORev/MS-C, a-GRK
KRR- sequence, corresponding to the nuclear localization signalof Tatprotein,wasinsertedatthethirdpositionin the MS-C ORF of the above deletion mutant to generate
A3-5ORev/MS-N(Fig. 3).
HeLa cellsweretransiently cotransfected with Revorthe
respective fusion protein expression plasmids and the indi-catedgag expression vector. Aliquots of transfected cells
were taken formetabolic labeling, and the labeledextracts
wereimmunoprecipitated with anti-Rev antiserum.
Alterna-tively, total cellular extractswere electrophoresed through
SDS-polyacrylamide gels, and the fusion proteins were
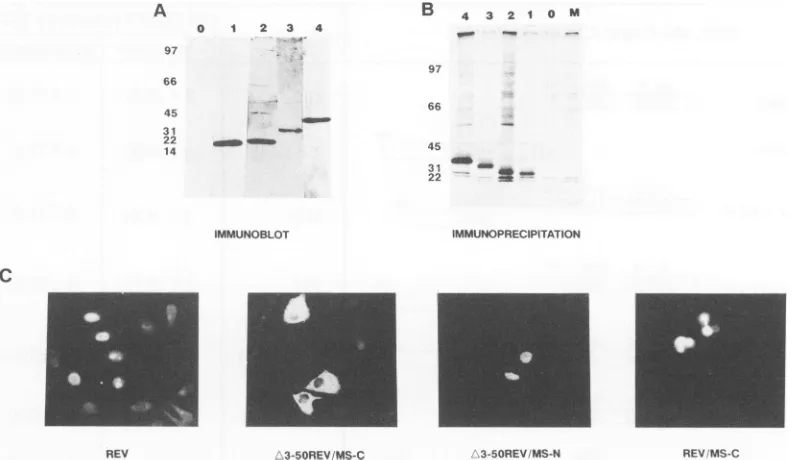
detected byimmunoblotting. As shown in Fig. 2A and B, fusionproteins of the predictedsizeswereeasilydetected by immunoblotting or immunoprecipitation with the anti-Rev
antiserum. Thesubcellular distribution ofthe various fusion proteinswasinvestigated by indirectimmunofluorescence of
fixed, permeabilized cells with anti-Rev antiserum. As
ex-pected, Revwasconcentrated inthenucleoli oftransfected
cells (Fig. 2C). Although the Rev/MS-C fusion protein
ap-peared concentrated in the nuclei, the protein was also
uniformly expressed throughout the cytoplasm andthe
nu-clei ofmanytransfectants. A3-5ORev/MS-C,whichlackeda
nuclearhoming signal, accumulated exclusively in the cyto-plasm.A3-5ORev/MS-N,withtheTatnucleartargetingsignal at the third codon ofMS-C, had adiffuse nuclear
accumu-lation with some hint of condensation, as well as diffuse
cytoplasmicstaining (Fig. 2C).
gagexpressionfrom plasmids containing thewtRREwas
compared with that from plasmids containing RREZ-MS following transfection of HeLa or Cos-7 cells. In thegag
ELISA,all of the cellextractswereserially dilutedtoobtain opticaldensityvalues of between 1 and 2 for RREactivation by Rev. Under these conditions, the ELISA values for a
positiveRevresponsevariedbetween 100 and500 timesthat ofgagexpression in the absence of Rev. The cutoffvalue
B2
* U U
*A
AG-C G-C
* A G-C U-A U-A G-C G-C
5'
3'
MS
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
0 1 2 3 4
---Gomm
B 4 3 2 tO M
97
66
45
31 _
22 -
-IMMUNOPRECIPITATION
[image:5.612.118.514.72.302.2]REV 3-50REVMS-C 3-50REViMS-N REVMS-C
FIG. 2. Eukaryotic expression and subcellular localization of Rev and selected Rev/MSfusion proteins. HeLacellsweretransfected with
therespectiveplasmids in thecontextoftransientgag expression fromanHIV-1LTR-linked, RREZ-MS-containinggag expressionplasmid.
(A) Immunoblotting. After electroporation, HeLa cellswereplatedon60-mm-diameter dishes and harvested 48to72 h later. Aone-third equivalent of cells from each dishwasused forimmunoblotting with polyclonal rabbit anti-Rev antiserum asdescribed in Materials and Methods. The results fromimmunoblotting of 12.5%polyacrylamidegelsareillustrated. Lanes: 0, notransactivator; 1, Rev; 2, Rev/MS-D;
3, A3-5ORev/MS-N; 4,Rev/MS-C. Molecular sizes (in kilodaltons)areindicatedatthe left. (B) Immunoprecipitation. Total cell lysateswere
prepared and processedforradioimmunoprecipitation with polyclonal rabbit anti-Rev antiserum. The immunoprecipitateswereresolved by SDS-polyacrylamide gel electrophoresis with 10% gels, and the radioactive proteinswerevisualized byfluorography. Lanes: M, mock transfection; 0, transfection without RevorRev/MS expression plasmids; 1, Rev; 2, A3-5ORev/MS-C; 3, A3-5ORev/MS-N; 4, Rev/MS-C.
Molecular sizes (in kilodaltons) are indicated at the left. (C) Indirect immunofluorescence assay. After DNA was electroporated, approximately2x 104HeLacellswereplatedoncoverslips in 12-or24-well plates and harvested 48to72 h later forimmunofluorescence
assay.The resultsobtained with four differentproteinsareillustrated.
(recommended by the manufacturer) for the negative
re-sponse was at ELISA readings that fell below 10% of RRE-Revvalues afterdilution. Valuesthatrangedbetween 10to 20% of those with the RRE and Revwere scored as
marginal responders. gag expression from the RREZ-MS
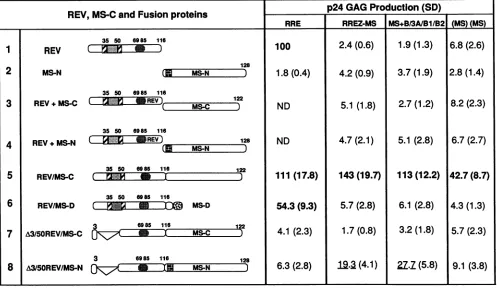
plasmidwasinducedbytheRev/MS-Cfusionproteinbutnot by Revorthe MS-C protein, alone orincombination (Fig. 3). TheRev/MS-Cfusionwas equallyefficacious with both
theRRE and theRREZ-MSplasmids (Fig. 3,row5).When
theMS ORF inRev/MS-Cwasinterrupted byasingle-base
deletion, causinga frameshift, theresultingfusion protein, Rev/MS-D, lost all activity with the RREZ-MS chimera while itpreservedthe activation of the RREplasmid (Fig.3,
row6). Thedeletion of 48 residuesnear the Nterminus of Revgenerated a fusion protein (A3-5ORev/MS-C) thatwas
predominantlylocalizedtothecytoplasmandwasinert with both the RRE and the RREZ-MS targets (Fig. 3, row 7). Insertion of the Tat nucleartargeting signal into A3-5ORev/ MS-C allowed the resulting mutant, A3-5ORev/MS-N, to accumulate in thenucleus but did not restore the activation for the RRE.A3-5ORev/MS-Nwasmarginallyactive with the RREZ-MS target (Fig. 3, row 8), which suggested that although A3-50Rev/MS-N most likelybound RREZ-MSvia the MS2 coat protein end, the N terminus of Rev was
requiredfor the activation oftheheterologous target. MS operator mutants with reduced in vitro coat protein affinities have corrsponding decreases in the in vivo Rev
response. To examine whether trans activation of the RREZ-MS chimerabythefusionproteinwasdictatedbythe
binding of the MS coat protein, the MS2 operator was
mutagenized to introduce changes known to reduce coat protein bindinginvitro. Deletion of the entireMSsequence
(RREZ) resulted in the lossof the Revresponse (Table 1).
Removalof thesinglebulgedAresiduein thestem structure of theMS2 operator has been showntoreduce the in vitro bindingaffinityof thecoatprotein bygreaterthan 2 orders of magnitude (10, 40).AnRREZ-MS chimera with this deletion (RREZ-MSAA) was defective for trans activation by Rev/ MS-C (Table 1). Changes at two other positions in the -AUUA-loopsequence [RREZ-MS(U/A) andRREZ-MS(A/
C)]knowntodiminish in vitrobindingbyatleast 10-fold(10,
40) ledto amoderate decrease or no change in the in vivo
responsetothe fusionprotein (Table 1).
To demonstrate directly that the Rev/MS-C fusion
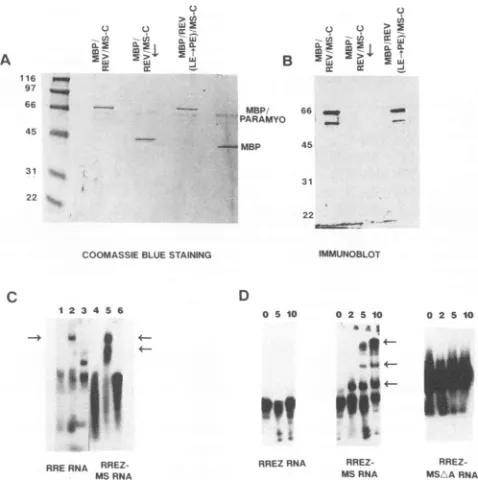
re-tained the authenticbinding properties of the Revand MS coat proteins, the Rev/MS-C protein wasoverexpressed in
E. coli as a fusion protein tagged to the C-terminal end of the MBP in the sense (MBP/Rev/MS-C) or the antisense (MBP/Rev/MS-9 orientation. Additionally, a derivative of
Rev/MS-C carrying a defective interfering mutation in theactivation domain of Rev[MBP/Rev(LE/PE)/MS-C]was
alsoexpressed andpurified. TheMBP fusionproteinswere
induced with IPTG and purified from inclusion bodies by affinity chromatography on amylose resin. Asingle major band of ca. 68 kDa was visualized in the affinity-purified fractions from the MBP/Rev/MS-C and the MBP/Rev(LE/ PE)/MS-C recombinants (Fig. 4A). The (MBP/Rev/MS-C) recombinantsynthesized anelongatedMBPtaggedattheC A
97 66 45 31 22 14
C
IMMUNOBLOT
on November 9, 2019 by guest
http://jvi.asm.org/
7474 VENKATESAN ET AL.
REV,
MS-C
and Fusion
proteins
p24 GAG Production (SD)
l
RRE RREZ-MS MS+BI3AIB1/B2 (MS) (MS)
1 |
695sREV
3 s 100 2.4 (0.6)1.9
(1.3) 6.8 (2-6)2
MS-N ( MS-N 1.8(0.4) 4.2(0.9) 3.7(1.9) 2.8(1.4)35 50 6985 116
R=EV12
3
REV+MS-C ( MS-C ND5.1
(1.8)
2.7(1.2)
8.2(2.3)
35 so 6985 116
4
|REV+ MS-N
(
REV)
M28
ND
4.7
(2.1)
5.1
(2.8)
6.7
(2.7)
35 50 685N 11612
5
REV/MS-C111
(17.8)
143(19.7)
113
(12.2) 42.7 (8.7)
6
REV/MS-D
C
MS-D
54.3
(9.3)
5.7 (2.8)
6.1 (2.8)
4.3 (1.3)
7 |A3I5OREVIMS-C |Z7chZZE
ieXZ
s-ZZ 4.1(2.3)
1.7(0.8)
3.2(1.8)
5.7(2.3)
8
A3/5OREV/MS-N|
9ZZ
lie6.3
(2.8)
|1..a
(4.1)
27.Z (5.8)
9.1
(3.8)
FIG. 3. trans activation of HIV-1gag mRNAs containing RRE, RREZIMS, (MS)+B/3A/MS/B2, and
[(MS)(MS)]
targets to Revandselected Rev/MS fusion proteins under transient conditions in HeLa cells. The ORFs of the various transactivators are schematically illustrated. In eachcase, the shaded square nearthe Nterminus ofRev representstheArg-rich domain requiredfor RNAbinding and nuclear-nucleolarlocalization, and therounded, shadedrectanglecorresponds totheleucine-rich activation domain. The hatchedregions flanking the RNAbinding motif representthe putative oligomerization domains. InMS-N,sixamino acids from thenuclear localization domain ofTathavebeeninsertedatthe thirdpositionofMS-C(6).MS-D refersto asingle-nucleotidedeletionatthe sixth codon of MS-C which resultsinaframeshift andearly termination(6).gagexpression values,normalizedfor transfectionefficiency,aretabulated with the respective standard deviations(SD) derived fromatleasteightindependentexperiments.Thepositivevaluesaredenotedbyboldtypeface, andintermediatevaluesareunderlined.Rev-dependentgagexpressionfroma wtRRE-containing plasmidisarbitrarily assignedavalue of 100. Theresults of thisandall othersubsequentexperimentswerealsoduplicatedwith Cos-7 cells. ND,notdone.
terminusby 12residuesencodedbytheantisense strandof
the Rev/MS-Cgene. Themajor 68-kDa band inthe lysates
from theMBP/Rev/MS-CandMBP/Rev(LE/PE)/MS-C cells
[image:6.612.63.560.75.363.2]immunoreacted with murine monoclonal anti-Rev antibody
TABLE 1. Invivoresponsesof mutationsinthe MSsequenceof RREZ-MStoRevandRev/MS-C
p24019production(± SD) relativetoRREand
RRE orRRE-MS Rev in the presence of:
RNAchimerae
No transactivator Rev Rev/MS-C
wtRRE 2.2(1.5) 100 111(17.8)
ERR(reverse RRE) 1.8(1.1) 3(1.3) 2(1.4)
RREZ 1.8(1.4) 3.1(1.7) 6.3(2.7)
RREZ-MS 4.8(2.4) 2.8(1.4) 143(19.7) RREZ-MSAA 3.7(1.8) 3.7(1.8) 12.3(6.8) RREZ-MS(U/A) 7.8(3.2) 1.8(0.9) 49(7.8) RREZ-MS(A/C) 10.7(4.3) 4.7(1.3) 87(14.6)
a In addition tothe deletion of theentire MSsequence(RREZ), threepoint
mutationswereintroduced in theMS stem-loopsequenceinthe context of the
RREZ-MS target: (i) deletion of the single bulged A in the MS stem
(RREZ-MSA&A), (ii) substitution ofAforUin the -AUUA-loop sequence
[RREZ-MS(U/A)], and (iii) substitution of Cfor A in the -AUUA- loop sequence[RREZ-MS(A/C)].NormalizedGAG-expression valuesareshown.
(Fig. 4B).Thefaster-migrating immunoreactive bandsinFig.
4B probably represent a protein
breakdown product(s).
Whenthepurified MBP
fusionproteins
wereincubated with[a32P]UMP-labeled
RRE or RREZ-MSRNA,
ribonucleo-protein (RNP) complexeswereformed only with theMBP/ Rev/MS-C protein and not with the fusion containing the
antisense Rev/MS-C(Fig. 4C). Under the sameconditions, MBP/Rev(LE/PE)/MS-C fusion protein bound both RRE andRREZ-MSRNAs,whilepurified RevboundonlyRRE RNA (data not shown). When purified
MBP/Rev/MS-C
protein was incubated with RREZ, RREZ-MS, or RREZ-MSAARNAs,
RNP
complexeswere formed onlywith theRREZ-MS RNA and not with the RREZ or the
RREZ-MSAA chimera withadeletedbulgedAresidue(Fig. 4D).
Definition ofthe minimalRRE/MSchimera that is respon-sive toRev/MS-Cfusionprotein in vivo. To examine whether the RRE sequences in the RREZ-MS target contribute to
activationby Rev/MS-Cfusionprotein(by recruitingcellular
factors, etc.), the stem-loops C, D, and E
and
the centralloopof the RRE were excised from RREZ-MS to generate
(MS)+A
and
(MS)+a. In (MS)+A, RREstem Asequencescorrespondingtopositions31 to50 andpositions 204 to 222 were
placed
5' and 3', respectively, of the MS sequence.RREsequences between positions 39 and 50 and between 1.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.62.303.568.673.2]>ux
A~~~~~~~~~~>(
~ ~ ~ ~ ~ ~
y L) cC
V) Uln -_
-A :;> E >1 m
cc cc X4
116 _m
97 _
66 _ .
45 _"
um
U
li c; a
L~2
;I tmB 2 i
cc --j
MBP 66 _
PARAMYO
-MBP 45
31
22
22
IMMUNOBLOT
C
12 3 4 S 6
I
4
t
;* t
RRE RNA
RREZ-MS RNA
D
0510 025 10
4.
4-h*
+-"I
i.
[image:7.612.63.302.74.314.2]02 510
TABLE 2. Minimalheterologous RNA recognized by Rev/MS-C in vivo
p241a production (- SD)relative to RRE and RRE or RRE-MS Rev in the presenceof":
RNAchimerae
Notransactivator Rev Rev/MS-C
wt RRE 2.0 (0.4) 100 111 (17.8)
ERR(reverse RRE) 1(0.7) 3(1.3) 2(1.4)
RREZ-MS 2.3 (0.9) 2.8 (1.4) 143 (19.7)
(MS)+A 2.8 (1.5) 1.8(0.6) 32.8 (6.8)
(MS)+a 0.8(0.5) 0.8(0.6) 45.8(9.8)
(MS) 4.8(1.9) 3.7(1.3) 17.1(6.2)
(SM) 5.1(1.3) NDc 4.1(2.2)
MSAA) 6.1(4.4) 3.7(1.3) 3.1(1.2)
(MS)(MS) 9.1(5.4) 3.9(2.2) 42.7(8.7)
(MS)(SM) 8.4(4.7) ND 29.7(9.2)
aThreetypesof minimaltargets were analyzed: (i) MS sequence flanked by
the 5' and 3' strands ofstem Aof RRE[(MS)+Aand(MS)+ainFig.1], (ii)
complete exchange of theRREwiththe MS sequence in the sense (MS) or
antisense (SM) orientationorwithanMSmutantwhichdeleted thesingle
bulgedA residue(MSAA), and (iii)ahead-to-tail [(MS)(MS)]or head-to-head
[(MS)(SM)]dimer oftheMSsequence.
bgagexpressionfromthe respectivetargets in response to Rev or Rev/
MS-Corthat in theirabsence is tabulatedas described in the legend to Fig. 3. cND, notdone.
RREZRNA RREZ-
RREZ-MS RNA MS. A RNA
FIG. 4. Characterizationof Rev/MS fusion proteins expressed in E. coli. Rev/MS-C and Rev(LE/PE)/MS-Cproteins were expressed in E. coli as MBP fusion proteins and purified as described in Materials and Methods. (A) Coomassieblue staining of the respec-tive fusion proteins purified by affinity chromatography and resolved by SDS-polyacrylamide gel electrophoresis with12% gels. Results obtained with Rev/MS-C fusedto MBPin the sense and antisense (underlined by arrow) orientations and MBP/Rev(LE/PE)/MS-C fusionareshown. LE- PEreferstothe substitution ofleucine for proline in the -LE- sequence in the activation domain of Rev. Molecular mass markerswere run on the left lane, and sizes (in kilodaltons) are indicated. Purified MBP and paramyosin-MBP fusionproteinswereobtainedcommerciallyandelectrophoresed as indicated on the right. (B) Immunoblot of the respective fusion proteins. Inclusion bodies fromE. coli expressing the variousfusion proteinsweresolubilizedand electrophoresedon12% gelsin SDS. Theproteinswereelectroblotted and reactedwithmurine anti-Rev monoclonal antibodies. Immunoreactive bandswerevisualized bya commercial chemiluminescence protocol (Amersham Corp.). The faster-migrating band probablyrepresentsabreakdownproduct(s). (C) Gel mobility shift analysis ofRNAbinding by Rev/MS-C fusion proteins.Theaffinity-purified fusion proteins described for panelA wereincubated with[a-32P]UMP-labeled RRE RNA orRREZ-MS RNA, and the formation ofRNA-protein complexes (RNPs) was evaluated by polyacrylamide gel electrophoresis as described in Materials and Methods.Lanes: 1and4, control lanes withoutE.coli proteins; 2 and 5, results obtained with MBP/Rev/MS-C fusion protein; 3 and 6, MBP ORF fused to the antisense strand of Rev/MS-C. Thearrowsdenotethe RNPs. Theslow-migratingband in lane 3represents aconformational isomer of theRREandnotthe RNP. (D) Sequence-specific binding by MBP/Rev/MS-C fusion protein. Increasing concentrations (top) of affinity-purified MBP/ Rev/MS-C proteinwereincubatedwithconstantmolarequivalents of [a-32P]UMP-labeled RREZ, RREZ-MS, and RREZ-MSAA RNAs, and theRNPcomplexeswereanalyzed bypolyacrylamide gelelectrophoresis.
positions 204 and 213 flanked the MS sequence in (MS)+a (Fig. 1). Both(MS)+A and(MS)+aweretrans-activatedat levels somewhat lower than the RREZ-MS activation(Table 2). As expected, neither chimeraresponded to Rev alone. This led us topursue whetherMS sequence alone may be sufficienttotargetthefusionproteinin vivo.Arecombinant
that completely exchanged the RRE for the MS sequence
(MS) was poorly activated by Rev/MS-C, while the MS sequence inreverseorientation (SM) and the MS sequence with the deleted Abulge (MSAA) were not responsive to Rev orRev/MS-C(Table 2).However,when a head-to-tail dimer
[(MS)(MS)] linked by a 6-nucleotide palindromic Asp-718
site (-GGTACC-) was inserted in place of the RRE, the resulting RNA had a modest response compared with that of RREZ-MS(Table 2). Ahead-to-head dimer[(MS)(SM)]was less efficientfor the Rev/MS-C-dependent response (Table 2). These resultsimplythat the MS operator sequence alone is sufficient to tether the fusion protein if the operator structureisexposed for protein binding.
Invivo responsesofthe bivalent RNAs correlate with their predicted binding affinities for Rev and MS-C. Rev/MS-C trans-activatedboth RRE and RREZ-MSRNA,presumably
by its abilitytobindthe RREby the N-terminal end of Rev and the MS operatorby thecoatprotein moiety. An RNA moleculecontainingboth ofthesetargets maybeinactivated at one orboth ofthe sites ofprotein binding and used to uncouple the bindingand theactivationeventsforthe fusion
protein.Twobivalentchimerasweredesigned:(i) B/3G/MS/ B2, whichreplaced theB1stem-loopin the
Rev-responsive
B/3G/B1/B2RREsubdomainwith the operator sequence but leftthecriticalthree-Gsequenceintact, and (ii)
(MS)+B/B1/
B2, with tandemly placed MS and B/B1/B2 sequence ele-ments(Fig. 1). Neither of theseplasmids expressedgagin
the absence of Rev or Rev/MS-C. B/3G/MS/B2 was acti-vated by Rev/MS-C at 40% of the levels observed with
RREZ-MS
(Table 3).
Inthe presenceofRev,gagexpression
fromB/3G/MS/B2wasabout24% of thatfromanequivalent
RRE plasmid (Table 3). Arecombinant that hadtandemly
placedMS and B/B1/B2 targets
[(MS)+B/B1/B2]
was acti-vatedby bothRevandRev/MS-Ctolevelsexceeding
thoseobtained with the RRE and Rev or
RRE/MS-C
andRev/
MS-C,respectively(Table
3).
The above bivalent chimerasweremutatedtorestrict their
targeting for the Rev or MS-C component of the fusion
protein. When the three G's in the
Rev-responsive
subdo-mainwerereplaced bythreeA's,Revwas nolonger
abletoon November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.315.561.96.231.2]7476 VENKATESAN ET AL.
TABLE 3. In vivo Rev responses ofwtandmutant
bivalent chimeras
p240agproduction (± SD) relativetoRRE and
RRE or RRE-MS RNA Rev in the presenceofb: chimerasa
Notransactivator Rev Rev/MS-C
wtRRE 2.0(0.4) 100 111(17.8)
RREZ-MS 2.3(0.9) 2.8(1.4) 143(19.7)
B/3G/MS/B2 7.8(3.8) 24(6.5) 59.7(11.3) B/3A/MS/B2 7.8(3.8) 5.1(2.5) 43.7(9.3) B/3G/MSAA/B2 5.2(2.2) 22(8.2) 37.7(8.3) (MS)+B/3G/B1/B2 4.3(1.2) 137(21.8) 173(18.4) (MS)+B/3A/B1/B2 5.5(3.2) 7.2(1.8) 94.3(12.4) (MSAA)+B/3G/B1/B2 3.8 (1.8) 96.7(9.8) 87.3(6.7) (MSAA)+B/3A/B1/B2 6.3 (2.8) 2.7(0.8) 9.3(4.9)
aTwoplasmids containingtargets forbothRevandthecoatproteinwere
used.In B/3G/MS/B2,the MSsequence replacedtheBi stem-loop in the
Rev-responsiveRREsubdomainB/B1/B2butleft the three G's ofthe RRE
intact.In(MS)+B/B1/B2, the MS sequencewaslinkedtothe downstream
Rev-responsiveB/B1/B2subdomain of theRREbythree A's.Thechimeras
weremutagenizedtoreplace the three G's intheRev-responsivesubdomain
with three A's,todelete thesinglebulgedAintheMSoperator, or todo both.
bgagexpressionvalues in thepresenceof RevorRev/MS-Corthose in
their absenceareshown.
activate theresultingchimera (Table 3).Thechangeof three G'stothreeA'sin thecontextof the full RRE haspreviously
beendemonstratedtolose the Rev response withoutovertly affectingthefolding dynamicsof the RREstructure(23, 24).
The three-G-to-three-Atransition mutantresulted onlyin a slight to moderate diminution in the activation responseto Rev/MS-C(Table3), since thesemutants retainthebinding
potential for MS-C. In contrast,amutationwhichdeletes the
bulgedAresidue maylosecoatproteinbindingbutcanstill beboundbyRev. Accordingly, deletion of thebulgedAin MS induced only a modest diminution of the in vivo re-sponse toboth Rev and Rev/MS-C (Table 3). Mutatingthe
(MS)+B/B1/B2RNAatboth the three G's and the Abulgein the MS operator causeda severeimpairmentof activationby Rev/MS-C andacomplete loss of the Rev response (Table 3).
A comparison of the in vivo responses of [(MS)(MS)],
(MS)+B/3A/MS/B2,
and RREZ-MS to Rev and combina-tions of the Rev and Rev/MS fusionproteinsis shown inFig.3. All three chimeras lack Rev recognition sites and are exclusively activatedby theRev/MS-C fusionprotein (Fig.
3, row 5) but not by Rev, MS-C, or MS-N, alone or in combination(Fig.3,rows1to4).All threeunivalenttargets also failed to respond to the Rev/MS-D fusion protein containingaprematurely terminated MScoatprotein(Fig.3, row6). The differential responses of the three chimeras to
Rev/MS-C probably reflect the intrinsic stability of the MS RNA secondary structure in the various molecules rather than the requirements for additional sequence to facilitate
protein binding. Therefore, ifthe RNA can bind the fusion protein, Rev moiety will activate that RNA. A large deletion at the Nterminus of Rev in the fusion protein (A3-5ORev/ MS-C)abolished trans activation(Fig. 3, row 7), most likely because of the loss of nuclear targeting (Fig. 2C). Insertion ofthe HIV-1 Tat nuclear homing signal in the coat protein
moiety
ofthe above deletion mutant generatedA3-50Rev/
MS-N,whichwasdistributed in both the nuclear and
cyto-plasmic compartments (Fig. 2C). However, with the
A3-5ORev/MS-N
protein, onlyafraction of the trans-activationpotentialofRev/MS-Cwasrealized(Fig.3, row 8), implying that foroptimaltransactivation of evenaheterologous RNA
TABLE 4. Repressionof MSoperatoractivationbyMS-N coatprotein'
p2401 productionb
Type oftransfection
- MS-N + MS-N
RRE+ Rev 100 89
RRE + Rev/MS-C 117 84
RREZ-MS + Rev/MS-C 135 23
[(MS)+B/3G/B1/B2] + Rev/MS-C 144 87
[(MSAA)+B/3G/B1/B2] + Rev/MS-C 87 73
[(MS)+B/3A/B1/B2] + Rev/MS-C 82 17
aAnMS-Nexpressionplasmidwascotransfected intoHeLacells withRev
orRev/MS-C transactivatorplasmidswith therespectivegagtargets.Ineach
experiment,theMS-Nplasmidwastransfectedat asixfoldmolarexcess over
Rev orRev/MS-CDNA.
bNormalized gagexpressionvaluesareshown. Dataareaveragevalues
from two orthreeexperiments.
target,thestructural
integrity
of the N terminus of Rev has tobepreserved.Coatproteininhibits thetransactivation of theMSoperator by Rev/MS-C. To examine whether the MS operatorwas boundby the coat
protein
invivo,
we cotransfectedHeLa cells with the MS-Nexpression plasmidunder conditions of trans activationby
RevorRev/MS-C.
MS-Nprotein
con-tained the HIV-1 Tat nuclear localizationsignals
and was distributedin both the nuclear and thecytoplasmic
compart-ments. MS-N would be expectedto completedirectly
with the fusionproteinfor the targetscontaining
the MS operator butnotfor theRev-responsive
target. Asshownin Table4,
asixfold molar excess of MS-Nover
Rev/MS-C
induced a sevenfold decrease in the trans activationof RREZ-MSby
the Rev/MS-C fusionprotein; activation of the RRE eitherby Rev or by the fusion
protein
was notsignificantly
af-fected. Activation of the bivalent RNA and the mutant chimera restricted to Revbinding
was alsonotsignificantly
influenced by MS-N coexpression(Table 4).
In contrast,MS-N caused a fivefold reduction in Rev/MS-C-dependent
gagexpression from achimera containingthe three-A mu-tation in the Rev-responsivemotif
(Table
4).Activationby the fusion protein requires the Rev activation domain. Genetic studies have identified an Leu-rich motif near the C terminus of Rev as the activation domain. Mutations inthis domain were not only inactive fortrans activation but alsodominantlyinterfered with the
function-ingofwtRev (31, 34).Toexaminewhethertransactivation
of the heterologous RNAbythe fusion protein was
depen-dent on the
integrity
of the activation domainof Rev, we designedtwosubstitutionmutantswithchanges
in the Leu-rich motifof Rev inthecontextofthefusionprotein. BothRev(78-79LE/EE)/MS-C
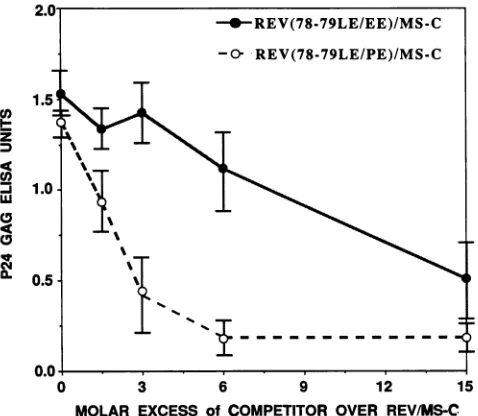
and Rev(78-79LE/PE)/MS-C pro-teinswereinactive for gagexpression from RREplasmids;theyweremarginallyactive(10to20% of the levelsobtained with Rev/MS-C) on the RREZ/MS-C plasmid (data not shown). However, when increasing amounts of either mu-tant werecotransfected with thewtRev/MS-C, therewas a dose-dependent repression oftrans activation (Fig. 5). Un-der similarconditions, cotransfection ofamolar excess of
A3-5ORev/MS-C was without an inhibitory effect (data not
shown).Themutant(-LE-to-PP-)wasmoreinhibitorythan the change (-LE- to -EE-). trans activation of the RREor
(MS)+B/3A/B1/B2
containinggagplasmids bythewtfusionprotein
was repressed more efficiently (90% inhibition at a fivefoldmolarexcess) bythesameLeumotifmutants(data not shown). These results, which parallel the previously J. VIROL.on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.61.302.95.222.2]Cn
z
4L
CD
0 3 6 9 12 15
MOLAR EXCESS of COMPETITOR OVER REV/MS-C
FIG. 5. Effects of dominant interfering mutations near the C
terminus of Rev on trans activation by Rev/MS-C. Increasing
amountsof RSV LTR-linkedRev/MS-C proteinswith substitutions
at the 78 and 79 residues of the RevORFwere cotransfected with fixed amounts ofRev/MS-C and RREZ-MSgagplasmid. Normal-ized values ofp24,a' production arepresentedas afunction of the
competitor concentration. Conditions are as described for other
experiments.
reported behavior of activation domain mutants of Rev on
the RRE (31, 34), imply common biochemical pathways of activation for both Rev and Rev/MS-Cfusion proteins.
DISCUSSION
We have demonstrated that direct interactionof Rev with the RRE RNA is not an absolute requirement for trans
activation. Rev will activate a heterologous RNA if it is directed to it by another RNA binding protein. When the Rev-binding domain of the RRE was replaced bythe MS2 phageoperatorsequence(RREZ-MS), Rev/MScoatprotein fusion activated therespectivegagmRNA at levels slightly greater than those for the corresponding activation of the RRE by Rev. Mutations in the MS operator known to abolish or reduce in vitro binding by the coat protein demonstratedacorrespondingloss of in vivoresponseto the fusionprotein. Thispoint was directlyconfirmed by the in vitrobinding preferences of thepurified fusionproteinwith the various RNAs. The purified fusion protein bound the RRE and RREZ-MS RNAs but not the RREZ or RREZ-MSAARNAs.
Althoughthe RREZ-MS chimerawas fullyfunctional for the Rev response, RNA with a complete exchange of the
RRE with the MS sequence was activated by the fusion
protein to less than 20% that of RRE activation by Rev.
Activation of the MS stemwas slightly improved when it
was extendedtoincludethesequence correspondingto the
RREstemA. This improvementwasnot attributable to the
sequence content of the RRE, since similar enhancement
was also observed by lengthening the MS stem by an
unrelated sequence (data not shown). A head-to-tail or a
head-to-head MS dimer also showed a modest increase in
response over the MS monomer response to the fusion
protein.A deletionmutantof the RRE(RREZ)that removed
the
Rev-responsive
domain(B/B1/B2)
wasactivated neitherby
Rev norby
the fusionprotein.
Therefore, the MS sequence alone is sufficient to tether the fusion proteinduring
trans activation if this sequence forms a stablesecondary
structurein thecontextof the ratherlarge
HIV-1gag-polmRNA,asinthecasesof RREZ-MS
and,
to alesserdegree,
theotherminimalMSchimeras.The fusion
protein
wasequally
effectiveagainst
both the RREandthe RREZ-MStargets.Cotransfection ofRREandRREZ-MS
expression plasmids
resulted in an additive re-sponse when the fusionprotein
was notlimiting (data
notshown).
Directproof
for the dual-RNA attachmentpotential
of the fusionprotein
was obtained witha bivalent chimera that contained the MS operator and the RREsubdomain in tandem[(MS)+B/3G/B1/B2].
As summarized inFig. 6,
thebivalent targetwas activated
by
both Rev andRev/MS-C.
Substitution of the three G's
by
three A's in theRev-responsive subdomain selectively abolishedthe responseto Rev but not that to
Rev/MS-C.
When the coatprotein
sequenceinRev/MS-C
wastruncated,
theresulting
mutant,Rev/MS-D,
failed to activatetheMS2-only
targets,empha-sizing
therequirement
of the coatprotein
end for RNAbinding.
Invivobinding
of the fusionprotein
to MS2RNA wasindirectly
demonstratedby
competition experiments
with excess
quantities
of the coatprotein,
MS-N. MS-Ninhibited the activationof
operator-only
targetsby
thefusionprotein
but hadnoeffectonRREactivation.Thus,
the RNAbinding
andactivationproperties
of Rev may bestructurally
andfunctionally
dissociatedwithout thefidelity
of the acti-vation processbeing
lost.The invivo reactivitiesof the bivalentchimerasreiterated that RNA attachment
by
eitherbinding
protein
ishighly
sequencespecific. Although
sequence-specific
interactionsof MS coat
protein
with the MS operator have been welldocumented,
the situationregarding RRE/Rev
interactionshasbeen
equivocal.
Recent reportshave in factclaimed thatRev
binding
of the RRE may bespecified
notby
aunique
sequence in theB/B1/B2
RREsubdomainbut ratherby
theunusual distortions in the sugar
phosphate
backboneim-posed by
noncanonicalbase-pairing
schemes(3, 18,
28).
We havedemonstratedpreviously
that Revbinding
and the Rev response arepreserved
in mutants thatexchanged
theBi
stem-loop
for aperfectly
Watson-Crickbase-paired
MS2
helixloop
both with Rev(24)
and withRev/MS-C
fusionproteins.
The loss of both the in vitro and in vivo Rev responses as aconsequence of the three G'sbeing
mutated upstream of the MS sequence in theseconstructsargues in favor ofsequence-specific binding
of Rev to theB/B1
stem-loop
region,
both in its native context and as theRev/MS-C
fusionprotein.
Severalstudies have
implied
thatforoptimal
activationby
Rev,hostcellular factorsbinding
toRRERNA, Rev,
orboth may berequired.
Several RRE RNAbinding
nuclear pro-teins were demonstrableby
a UVcross-linking
procedure
(36a), including
thepreviously reported
56-kDa RBFprotein
(44).
The case for theRRE-binding
Revhelper
factors issupported by
theinability
ofcertain RREmutantsthat bind Rev to be trans-activatedby
it(23)
andby
the restrictedtropism
of the Rev response(43).
Certain RREmutantsthat bound Revpoorly
andwerenotRevresponsive
in vivo stillfacilitatedgag
expression
inthe absence ofRev,
implying
the presenceofacellular
surrogate(s)
for Rev(24).
Our best non-RREconstructswere rescuedby
theRev/MS-C
fusionprotein
toonly
about 42% of the RREby Rev,
suggesting
that RRE sequence
recognition by
cellularfactors may alsoimprove
activationby
the fusionprotein.
However,
our1%
1%1.
- --- --- -- -- -
on November 9, 2019 by guest
http://jvi.asm.org/
[image:9.612.64.303.75.283.2]7478 VENKATESAN ET AL.
[image:10.612.91.521.73.296.2]MS/B/3G/B1 /B2
MS/B/3A/B1 /B2
MSAA/B/3G/B1 /B2
MSAA/B/3A/B1
/B2
FIG. 6. Schematicrepresentation ofRNAmotifsrecognized byRev and MScoatproteins.Achimeralinkingthe MSoperator sequence totheRev-responsiveB/B1/B2subdomain oftheRREwasmaximallyresponsivetobothRevandRev/MS-C.The cognate targetsforthe MS-C and Revproteinsaredenotedbythelightlyandheavilyshadedareas,respectively.Theasterisksrefertothechangeof threeG'sto threeA's intheRREsubdomainandtothe deletion that removed thebulgedAinthe MSoperator.The relativeRev responsesofthevarious mutants areshown.
substitution mutation of three G'stothree A's
(3G/3A)
inthe bivalent chimerawas notactivatedbyRevortheRev/MS-Dmutant that truncated the coat protein. Besides the MS2
RNA,the3G/3Amutantcontainedonlythe
non-Rev-respon-sive subdomain of the RRE. Furthermore, a
3G/3A
substi-tution mutant,B/3A/MS/B2, thatcontainedan evensmallerRev-responsive sequence (B and B2 stem-loops of the
B/B1/B2 subdomain) was also nonresponsive to Rev,
al-though not tothe fusion protein. While these observations underscore the fact that Rev binding to the target is the
primary event during activation, it is still possible that
RRE-binding cellular factors may regulate the overall effi-ciencyof the Rev response.
The amino-terminalpeptidedomain(s)of Rev is necessary for RNAbinding, nuclear-nucleolar localization, and
oligo-merizationproperties(8, 25, 27, 31).
A3-5ORev/MS-C
witha 48-amino-acid deletion at the N terminus of Rev accumu-lated preferentially in the cytoplasm and was therefore inactive with both the RRE and the MS chimeras. When the Tat nuclear localization sequence was inserted at the third codon of the MS-C ORF in the abovedeletion,theresultingfusion protein,
A3-5ORev/MS-N,
was distributed equallybetween thecytoplasm and the nucleus andpossibly in the nucleolus.
A3-5ORev/MS-N
failed to activate the RRE target andwasonly partiallyactive with the MS2 operator. Since A3-5ORev/MS-N lacks the RRE-binding site, its failure to activate the RRE is understandable. But its partial in vivo effect with the MS operator targets implies that the amino terminus of Rev may possess an additional function(s)re-quiredfor maximal activation.BindingtoheterologousRNA targets is probably not one of these properties, since the
Rev/MS-D fusion, with anintact Rev ORF but a truncated coat proteinORF, activated only the RRE but not the MS targets. Since
A3-5ORev/MS-N
is not excluded from thenucleus, a loss ofprotein oligomerization potential may be the critical defect in
A3-5ORev/MS-N.
However, theMScoatprotein itselfcandimerize and formhigher-orderstructures upon specific RNA binding (4, 48). It is likely that the A3-5ORev/MS-N fusion may have retained this property of theMS2coatprotein.Therefore, theA3-5ORev/MS-N dele-tionmutant mustlack yet another function encodedbythe N terminus of Rev. Even substitution mutants of Rev/MS-C
thatreplacedone or morearginineresidues in the N termi-nusof Revalso hadanegativephenotype withthe RRE and were only marginally active with the MS operator targets
(45a). Assuming that the various mutations in the Rev N terminus donotdrasticallyaffect fusionproteinfolding,we can speculateonthepossible roles of the Rev Nterminus. First,evenif RNAbindingoccursatthecoatproteinmoiety,
thefusionproteinmaystill havetooligomerize by usingthe N-terminal domain of Rev. Alternatively, activation may
require cooperative intramolecular interactionsbetween the
heterologousRNAbindingsite and theintactNterminus of
Revorintermolecular interactions between theNtermini of Revindifferent fusionproteinmolecules.Finally,an
authen-ticNterminus of Rev may berequiredforbindingof cellular factors.
In the life cycle of HIV, the Rev protein regulates the
temporal switch from the earlyregulatoryphase to the late
lytic phase (2, 13, 37,41). HIVreplicationmayberestricted atthe level of Rev from constraints of the threshold require-ments of Rev (33, 38, 39), by the requirement of species-specific cellular helper factors (43), or by both. Rev is
presumedtobea determinant of virallatencyand reactiva-tion during natural infection (38, 39) and is therefore an excellent candidate for targeted drug development. Like manyactivators, Revis a modularprotein, withthe N-ter-minal and theC-terminal halvessubservingthe RNA binding and the activation functions, respectively. Mutants with
changesin the leucine-rich activation domain bind the RRE, buttheyaredefectivefor activation anddominantly interfere with the Rev function (31), presumably by forming mixed J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
oligomers (26, 33). However, it is equally plausible that the activation domain interacts with specific cellular factors modulating the RNA spliceosome assembly or transnuclear RNA transport (11, 30, 31). The activation domain mutants also inhibited activation by the fusion protein, implying common biochemical pathways of activation for both Rev andRev/MS-C fusion proteins. While the activation domain is being explored as a potential target for drug development, the fusion protein system we have developed is useful for examining the various RNAs aspotential Rev decoys. Using adistant RNAbinding site in the fusion protein rather than the RRE-binding domain may uncover the in vivo RNA
bindingparameters and also identify cross talk between the heterologous RNA binding site and the Rev oligomerization domain at the N terminus. The need for an authentic
N-terminal domain of Rev for the activation of even the
heterologous targets prompts us to examine the contribu-tions of thedifferent residuesatthisdomain to the activation process. These studieswillallow us to synthesize "design-er" peptides that may interfere with the discrete steps leading to RNA binding, protein-protein interaction, and activation.
ACKNOWLEDGMENTS
We thank AliciaBuckler-White foroligonucleotide synthesis.We
aregratefultoPaulWingfield for the purifiedRevprotein expressed in E. coli.Wethank Barbara Felber ofNCI-FCRCand MarieLou Hammerskjold of SUNY, Buffalo, for anti-Rev antisera.Wethank Anthony S. Fauci, Bernard M. Moss, Damien Purcell, and Keith Peden ofNIAIDfor their criticalreviews andcomments.
This work was supported fully by a grant to S.V. from the intramural AIDS targeted anti-viral program administered by the OfficeoftheDirector, National Institutes of Health.
REFERENCES
1. Adachi, A.,H.Gendelman, S.Koenig, T.Folks, R.Willey, A. Rabson, and M. Martin. 1986.Production ofacquired immuno-deficiencysyndrome-associated retrovirus in human and
non-human cells transfected withan infectious molecular clone. J. Virol.59:284-291.
2. Ahmad, N., R. Maitra, and S. Venkatesan. 1989. Rev-induced modulation of Nef protein underlies temporal regulation of humanimmunodeficiency virus replication. Proc. Natl. Acad. Sci. USA86:6111-6115.
3. Bartel, D., M. Zapp, M.Green,andJ.W.SzostaL1991.HIV-1 Revregulation involves recognition of non-Watson-Crick base pairs in viralRNA.Cell 67:529-536.
4. Beckett, D.,H. N.Wu,and0. C. Uhlenbeck. 1988. Ribonucle-oprotein complexes of R17 coat protein and a translational operatoranalog.J. Mol. Biol. 204:927-948.
5. Berger, J., C. Aepinus, M. Dobrovnik, B. Fleckenstein, J. Hauber, and E. Bohnlein. 1991. Mutational analysis of func-tional domains in the HIV-1 Rev trans-regulatory protein. Virology 183:630-635.
6. Berkhout, B., and K. T. Jeang. 1990. Down modulation of HIV-1 gene expressionusinga procaryotic RNA-binding pro-tein.Nucleic AcidsRes.18:6903-6907.
7. Berkhout, B., R. H. Silverman, and K. T. Jeang. 1989. Tat trans-activates the human immunodeficiency virus through a
nascentRNA target.Cell59:273-282.
8. Bohnlein, E.,J. Berger, andJ. Hauber. 1991. Functional map-pingofthe human immunodeficiencyvirus type 1 Rev RNA binding domain:newinsightsintothe domain structureofRev andRex.J. Virol.65:7051-7055.
9. Calnan, B. J.,B. Tidor, S.Biancalana, D.Hudson, and A. D. Frankel. 1991.Arginine-mediated RNArecognition: the argin-inefork. Science 252:1167-1171.
10. Carey, J.,V.Cameron,P. deHasseth,and0.Uhlenbeck. 1983. Sequence specific interaction of R17 coat protein with its ribonucleic acidbindingsite.Biochemistry22:2601-2610.
11. Chang,D.D., andP.A. Sharp. 1989. Regulation byHIVRev dependsuponrecognitionofsplicesites. Cell 59:789-795. 12. Cochrane, A.,A.Perkins,andC.Rosen.1990.Identification of
sequences important in the nucleolar localization of human immunodeficiencyvirusRev:relevance of nucleolarlocalization
tofunction.J. Virol. 64:881-885.
13. Cullen, B. R. 1991. Regulation of HIV-1 gene expression. FASEBJ.5:2361-2368.
14. Cullen,B.R., J. Hauber, K.Campbell, J. G. Sodroski,W.A. Haseltine,andC. A. Rosen.1988. Subcellular localizationof the humanimmunodeficiencyvirustrans-actingartgeneproduct.J. Virol.62:2498-2501.
15. Daefler, S., M. E. Klotman, and S. F. Wong. 1990. Trans-activating revproteinof the human immunodeficiencyvirus 1 interacts directly and specifically with its target RNA. Proc. Natl. Acad. Sci. USA 87:4571-4575.
16. Daly,T.,KCook,G.Gray,T.Maione,andJ.R.Rusche.1989. Specific binding of HIV-1 recombinant Rev protein to the Rev-responsiveelementinvitro.Nature(London)342:816-819. 17. Dayton, E.,D.Powell,and A.Dayton. 1989. Functionalanalysis of CAR, the target sequence for the Rev protein ofHIV-1. Science 246:1625-1629.
18. Dayton, E.T.,D.A.Konings,D. M. Powell,B.A.Shapiro,L. Butini, J. V. Maizel, and A. I. Dayton. 1992. Extensive
se-quence-specificinformationthroughouttheCAR/RRE,the
tar-get sequenceof the humanimmunodeficiencyvirustype 1 Rev protein.J.Virol. 66:1139-1151.
19. Emerman, M., R. Vazeur, and K. Peden. 1989. The revgene productof the humanimmunodeficiencyvirusaffects envelope-specificRNAlocalization. Cell 57:1155-1165.
20. Felber, B., M.Hadzopoulu-Cladaras, C.Cladaras,T.A. Atha-nasopoulos,andG. Pavlakis.1989. Revproteinof human immu-nodeficiencyvirus type 1affects the stability and transport of theviralmRNA. Proc. Natl. Acad.Sci. USA 86:1495-1499. 21. Feng, S., and E. Holland. 1988. HIV-1 tat trans-activation
requiresthe loop sequence within the TAR. Nature (London) 334:165-167.
22. Heaphy, S., C. Dingwall, M. Ernberg, M. Gait, S. Green, J. Karn,A.Lowe,M.Singh,andM.Skinner. 1990. HIV-1
regula-tor of virion expression (Rev) binds to an RNA stem-loop
structure located in the Rev response element region. Cell 60:685-693.
23. Holland, S.M.,N.Ahmad, R. K. Maitra,P.Wingfield,andS. Venkatesan.1990. Humanimmunodeficiencyvirus Revprotein recognizesa target sequencein Rev-responsiveelement RNA within the context of RNA secondary structure. J. Virol. 64:5966-5975.
24. Holland,S.M.,M.Chavez,S.Gerstberger,andS. Venkatesan. 1992.Aspecificsequencewithabulgedguanosineresidue(s)in
astem-bulge-stemstructureofRev-responsiveelementRNAis requiredfortransactivationbyhumanimmunodeficiencyvirus type1 Rev. J.Virol. 66:3699-3706.
25. Hope, T. J.,X. J. Huang, D. McDonald, and T. G. Parslow. 1990. Steroid-receptor fusion of the human immunodeficiency virus type 1 Rev transactivator: mappingcryptic functions of the arginine-rich motif. Proc. Natl. Acad. Sci. USA 87:7787-7791.
26. Hope,T.J.,N. P.Klein,M. E.Elder,and T.G. Parslow. 1992. trans-dominant inhibition of human immunodeficiency virus type 1 Revoccurs through formationofinactiveprotein
com-plexes.J. Virol. 66:1849-1855.
27. Hope, T. J., D. McDonald, X. J. Huang, J. Low, and T. G. Parslow. 1990. Mutational analysisof the human immunodefi-ciencyvirus type 1 Revtransactivator: essential residuesnear
theamino terminus.J. Virol.64:5360-5366.
28. Kjems, J.,B.J.Calnan,A. D.Frankel,and P. A.Sharp. 1992. Specific bindingofabasic
peptide
from HIV-1Rev. EMBO J. 11:1119-1129.29. Long,E.O.,B.S.Rosen,D. R.Karp,M.Malnati,R. P.
Sekaly,
and D.Jaraquemada. 1991.EfficientcDNAexpression
vectorsforstable and transientexpressionofHLA-DR in transfected fibroblast andlymphoidcells.Hum.Immunol.31:229-235. 30. Lu, X. B., J. Heimer, D. Rekosh, and M. L.
Hammarskjold.
on November 9, 2019 by guest
http://jvi.asm.org/
7480 VENKATESAN ET AL.
1990.Ul small nuclearRNAplaysadirect roleintheformation ofaRevregulated human immunodeficiency virusenvmRNA that remains unspliced. Proc. Natl. Acad. Sci. USA 87:7598-7602.
31. Malim, M., S. Bohnlein, R. Fenrick, S. Le, J. Maizel, and B. Cullen. 1989. Functional dissection of the HIV-1 Rev trans-activator-derivation ofatrans-dominantrepressorof Rev func-tion. Cell 58:205-214.
32. Malim, M., J. Hauber, S.Le,J. Maizel, and B.Cullen.1989. The HIV-1 rev trans-activator acts through a structured target sequence toactivate nuclearexportofunspliced viralmRNA. Nature (London) 338:254-257.
33. Malim, M. H., and B. R. Cullen. 1991. HIV-1 structuralgene expression requires the binding of multiple Rev monomers to the viralRRE:implications for HIV-1 latency. Cell 65:241-248. 34. Malim, M. H., D. F. McCarn, L. S.Tiley, and B. R. Cullen. 1991. Mutational definition of the human immunodeficiency virustype 1 Revactivation domain.J. Virol. 65:4248-4254. 35. Malim, M. H., L. S. Tiley, D. F. McCarn, J. R. Rusche, J.
Hauber,and B. R. Cullen. 1990.HIV-1structuralgene expres-sion requires binding of the Rev trans-activator to its RNA target sequence. Cell 60:675-683.
36. Nalin, C. M., R. D. Purcell, D. Antelman, D. Mueller, L. Tomchak, B. Wegrzynski, E.McCarney,V.Toome, R. Kramer, and M. C. Hsu. 1990. Purification and characterization of recombinant Rev protein of human immunodeficiency virus type1. Proc.Natl.Acad. Sci. USA 87:7593-7597.
36a.Park,H.Unpublished data.
37. Pavlakis,G. N., and B. K. Felber. 1990. Regulationof expres-sion of humanimmunodeficiency virus. NewBiol.2:20-31. 38. Pomerantz, R. J., T. Seshamma,and D. Trono. 1992. Efficient
replication of human immunodeficiency virustype 1requires a threshold level of Rev: potential implications for latency. J. Virol. 66:1809-1813.
39. Pomerantz, R. J., D. Trono, M. B. Feinberg, and D. Baltimore. 1990. Cells nonproductively infected with HIV-1 exhibit an aberrant patternofviralRNAexpression:amolecularmodel for
latency. Cell 61:1271-1276.
40. Romaniuk, P., P.Lowary,H.Wu,G.Stormo,and0.Uhlenbeck. 1987. RNA binding site of R17 coat protein. Biochemistry 26:1563-1568.
41. Rosen, C. A. 1991. Regulation of HIV gene expression by RNA-protein interactions. Trends Genet. 7:9-14.
42. Roy,S., U. Delling,C. H.Chen, C. A. Rosen, and N.Sonenberg. 1990. AbulgestructureinHIV-1 TAR RNAisrequired for Tat binding and Tat-mediated trans-activation. GenesDev. 4:1365-1373.
43. Trono, D., and D. Baltimore. 1990. A human cell factor is essential forHIV-1Rev action.EMBO J. 9:4155-4160. 44. Vaishnav,Y. N.,M.Vaishnav,andS.F.Wong.1991.
Identifi-cation and characterization ofanuclear factor thatspecifically bindstothe Revresponseelement(RRE) of human immunode-ficiency virustype 1 (HIV-1). New Biol. 3:142-150.
45. Vaishnav, Y. N., and S. F. Wong. 1991. The biochemistry of AIDS. Annu.Rev. Biochem. 60:577-630.
45a.Venkatesan, S.Unpublished observations.
46. Weeks, K.M.,and D. M.Crothers.1991. RNArecognition by Tat-derived peptides: interaction in the major groove? Cell 66:577-588.
47. Wingfield,P. T., S. J.Stahl,M.A. Payton, S. Venkatesan, M. Misra, andA. C. Steven.1991.HIV-1 Revexpressed in recom-binantEscherichia coli: purification, polymerization, and con-formationalproperties. Biochemistry 30:7527-7534.
48. Witherell, G. W., H. N. Wu, and 0. C. Uhlenbeck. 1990. Cooperative binding ofR17coatproteintoRNA.Biochemistry 29:11051-11057.
49. Zapp, M., and M.Green. 1989.Sequence-specificRNAbinding by theHIV-1 Revprotein. Nature (London)342:714-716. 50. Zapp, M. L., T. J. Hope, T.G.Parslow, and M. R. Green.1991.
Oligomerization andRNAbinding domains of thetype1human immunodeficiency virus Rev protein: a dual function for an arginine-rich binding motif. Proc. Natl. Acad. Sci. USA 88: 7734-7738.
J. VIROL.