0022-538X/95/$04.0010
Copyrightq1995, American Society for Microbiology
Asymmetric Replication In Vitro from a Human Sequence
Element Is Dependent on Adeno-Associated Virus Rep Protein
ELENA URCELAY,
1PETER WARD,
2STEPHEN M. WIENER,
1BRIAN SAFER,
1AND
ROBERT M. KOTIN
1*
Molecular Hematology Branch, National Heart, Lung, and Blood Institute, Bethesda, Maryland, 20892,
1and
Department of Microbiology, Cornell University Medical College, New York, New York 10021
2Received 11 October 1994/Accepted 3 January 1995
The DNA of human parvovirus adeno-associated virus type 2 (AAV) integrates preferentially into a defined
region of human chromosome 19. Southern blots of genomic DNA from latently infected cell lines revealed that
the provirus was not simply inserted into the cellular DNA. Both the proviral and adjoining cellular DNA
organization indicated that integration occurred by a complex, coordinated process involving limited DNA
replication and rearrangements. However, the mechanism for targeted integration has remained obscure. The
two larger nonstructural proteins (Rep68 and Rep78) of AAV bind to a sequence element that is present in both
the integration locus (P1) and the AAV inverted terminal repeat. This binding may be important for targeted
integration. To investigate the mechanism of targeted integration, we tested the cloned integration site
subfragment in a cell-free replication assay in the presence or absence of recombinant Rep proteins. Extensive,
asymmetric replication of linear or open-circular template DNA was dependent on the presence of P1 sequence
and Rep protein. The activities of Rep on the cloned P1 element are analogous to activities on the AAV inverted
terminal repeat. Replication apparently initiates from a 3
*
-OH generated by the sequence-specific nicking
activity of Rep. This results in a covalent attachment between Rep and the 5
*
-thymidine of the nick. The
complexity of proviral structures can be explained by the participation of limited DNA replication facilitated
by Rep during integration.
Mammalian chromosomal DNA replication initiates from
sites dispersed throughout the genome with cell cycle
regula-tion. An early step in replication is recognition of origin of
replication (ori) sequences by a DNA-binding protein that is
thought to function by nucleating polymerase (Pol) complexes
for leading- and lagging-strand DNA synthesis (for reviews, see
references 3 and 16). Cellular ori sequence elements and
cel-lular factors involved in initiation of replication have not been
characterized. Mammalian DNA viruses have been useful as
paradigms of cellular DNA replication. These viral ori
se-quences have been defined (for a review, see reference 5) for
simian virus 40 (SV40) (1, 14, 22) and herpes simplex virus
(34). Viral proteins required for initiation of replication that
function as origin-binding proteins have been characterized,
including, e.g., SV40 large-t antigen (T-Ag) (9, 36) and herpes
simplex virus UL-9 (25). The development of an in vitro
rep-lication system based on SV40 (21, 33, 43) has led to the
characterization of cellular proteins necessary for DNA
syn-thesis (for reviews, see references 15 and 32). T-Ag is the only
viral gene product necessary for SV40 DNA replication; all
other components are cellularly derived. T-Ag hexamers bind
to the ori and initiate replication by unwinding the DNA with
T-Ag helicase. DNA Pol
a
-primase complex associates with
T-Ag via the B subunit of Pol
a
(8). Pol
d
and proliferating cell
nuclear antigen displace Pol
a
-primase and processively extend
the leading strand (37–39). Pol
a
-primase remains associated
with T-Ag and catalyzes lagging-strand synthesis. The
symme-try of the SV40 ori and T-Ag double hexamer results in
bidi-rectional replication.
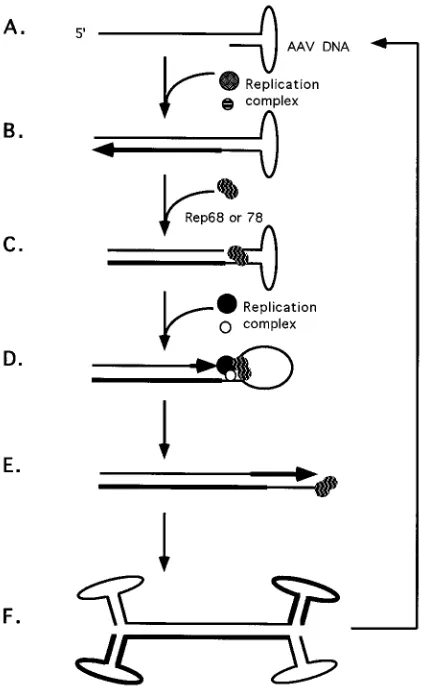
In contrast, adeno-associated virus type 2 (AAV) DNA
rep-licates only by leading-strand synthesis (for a review, see
ref-erence 2). The genome of AAV is single-stranded linear DNA,
and either the positive or negative strand is infectious (29).
Replication of AAV is dependent on the AAV nonstructural
proteins (Rep); either Rep68 or Rep78 is the only AAV
pro-tein required for replication in vitro (16, 24). Several activities
have been characterized for Rep68 and Rep78 that are
in-volved in viral DNA replication, including binding to the AAV
inverted terminal repeat (ITR) (12, 13, 26), sequence-specific
DNA binding (6, 42), sequence- and strand-specific
endonu-clease activity (12), and ATP-dependent DNA helicase activity
(12). Second-strand synthesis by cellular replication proteins
initiates from the 3
9
-OH of the hairpinned ITR (Fig. 1A) (for
reviews of AAV replication, see references 2, 4, and 23). A
duplex replication intermediate, which is covalently attached at
one end through the ITR (Fig. 1B), requires AAV Rep
tein(s) for terminal resolution. Four Rep proteins are
pro-duced in cells productively infected with AAV and helper
virus: Rep78, Rep68, Rep52, and Rep40. Either Rep68 or
Rep78 binds to a specific region within the ITR and nicks one
strand of the duplex at a unique site (Fig. 1C). The
endonu-clease reaction results in covalent attachment of Rep to the
newly generated 5
9
end. The 3
9
end of the nick serves as a
primer for extension of the ITR, possibly by the same type of
Pol complex involved in viral second-strand synthesis (Fig. 1D
and E). The helicase activity of the covalently attached Rep
molecule may unwind the secondary structure of the ITR. The
process of terminal resolution provides the means for
restora-tion of the termini and generarestora-tion of progeny viral genomes
(Fig. 1F). Thus, AAV DNA replication can be accounted for
entirely by leading-strand synthesis independent of de novo
synthesis. Recently, a truncated ITR (
D
ITR) incapable of
hair-pin formation has been shown to act as a Rep-responsive ori in
vitro (6).
AAV DNA has the unique property among animal DNA
* Corresponding author. Mailing address: Building 10, Room 7D18, Molecular Hematology Branch, National Heart, Lung, and Blood In-stitute, National Institutes of Health, Bethesda, MD 20892. Phone: (301) 496-1594. Fax: (301) 496-9985.
2038
on November 9, 2019 by guest
http://jvi.asm.org/
viruses of integration into a defined region of the human
ge-nome in chromosome 19 q.13.3 - q.ter (17–19, 30). An
expla-nation to account for this phenomenon has been elusive until
the recent finding that Rep bound specifically to a defined
region (P1) within the integration locus that has a sequence
element similar to that of the Rep-binding sequence in the
AAV ITR (see Fig. 2) (42). Because of these newly described
findings and to extend the model for targeted integration, we
tested the cloned chromosome 19 Rep-binding element
(pMAT50) in an in vitro replication assay.
MATERIALS AND METHODS
Plasmids.A SmaI subfragment derived from the human chromosome 19 integration site for AAV DNA (see Fig. 2) (18, 19) has previously been shown to bind Rep68 and Rep78 was inserted into the SmaI site of pUC19 (42). Cloned full-length, wild-type AAV DNA has been described elsewhere (20). Plasmid pSVoriAAV was produced by isolating a 326-bp AvrII-PvuII fragment corre-sponding to nucleotides (nt) 5187 to 270 of SV40 DNA. The overhangs of the restriction sites were blunted with T4 DNA polymerase and inserted into the EcoRV site of pBluescript SK(1). The 4.4-kb MscI fragment of pAV2 was inserted into the SmaI site of the plasmid containing the SV40 sequences. Plasmid pHisD is an 11,000-bp plasmid that contains the bacterial gene for histidinol dehydrogenase (HisD) and a portion of the genomic sequence for the mouse hypoxanthine phosphoribosyltransferase gene (28).
In vitro replication reactions.Cell extracts were prepared from HeLa S3 cells
as described elsewhere (40). The replication reactions were performed as de-scribed elsewhere (40), with the following modifications: the total dCTP concen-tration was increased to 0.1 mM, and the reaction mixtures were preincubated for 1.5 h prior to the addition of [a-32P]dCTP (specific activity,$5,000 Ci/mmol). The latter modification reduces the labeling from the putative repair activities to negligible levels. The concentration of protein in the cell extract was determined by the Bio-Rad colorimetric assay. Each assay was done with 0.1 mg of cellular protein in a final volume of 15ml that contained 7 mM MgCl2; 4 mM ATP; 200 mM each CTP, GTP, and UTP; 100mM each dATP, dGTP, and dTTP; 10mM dCTP; and 10mCi of [a-32P]dCTP (6,000 Ci/mmol; Amersham), 2 mM dithio-threitol (DTT), 4mg of bovine serum albumin, 40 mM creatine phosphate (pH 7.7), 2mg of creatine phosphokinase, 100mg of HeLa cell extract protein (40), 0.3mg of$90% supercoiled plasmid. When indicated, 1mg of the recombinant fusion protein was included (7).
Each reaction mixture was incubated at 348C for 18 h. Reactions were termi-nated by passing over a Sephadex G-50 spin column (5939, Inc.) and digestion for 2 h at 378C with proteinase K (200mg/ml) in 7 mM EDTA–0.2% Sodium dodecyl sulfate (SDS)–50 mM NaCl. The nucleic acids were extracted with phenol-chloroform, precipitated with ethanol, and dissolved in 2.5 mM Tris-Cl (pH 7.5)–0.25 mM EDTA. Products were fractionated by electrophoresis on 1% (wt/vol) Tris-borate-EDTA (TBE) agarose gels. The gels were dried and exposed to X-ray film (Kodak X-Omat).
DpnI andMboI digestions.DNA for DpnI sensitivity assays was prepared as described above, with the exception that no [a-32
P]dCTP was added. The DNA samples were digested with increasing amounts of DpnI for 3 h and fractionated on a 1% agarose TBE gel with 0.5mg of ethidium bromide per ml. The extent of cleavage was determined by UV light-induced fluoresence of the DNA bands in the gel.
Sensitivity to MboI was determined using conditions established for complete digestion of amethylated phage lambda DNA.
MBP-Rep proteins.The construction of the maltose-binding protein (MBP)-Rep expression vectors and production of the MBP-(MBP)-Rep proteins used for in vitro replication reactions have been described in detail elsewhere (6). Briefly, MBP-Rep68Dwas produced by PCR amplification of the open reading frame (ORF) of rep from codons 3 to 520. The PCR product was cloned in frame with the malE ORF into expression vector pPR997 (New England Biolabs). MBP-Rep78 was derived from MBP-Rep68Dby extension of the amino terminus of the ORF with an overlapping PCR product. MBP-Rep68DNTP contains a substitu-tion of lysine 340 with histidine. The activities of the fusion Rep proteins were determined and were found to be similar to those of wild-type Rep protein (6, 7).
[image:2.612.72.285.71.414.2]Covalent attachment of MBP-Rep78 to the 110-bp P1 element and flanking polylinker sequence.The 110-bp P1 sequence was removed from pMAT50 by EcoRI and HindIII digestion. The fragment was either 59 end labeled with polynucleotide kinase and [g-32P]ATP or 39end labeled with Klenow fragment
[image:2.612.314.554.452.650.2]FIG. 2. Sequence alignment of the P1 element derived from the chromosome 19 integration locus (AAVS1) with the AAV ITR. AAVS1 is represented sche-matically by the positions of the CpG island (filled box) (16), and sites of proviral integration are indicated (hatched region) (16, 28, 29). P1 was obtained as a SmaI subfragment of AAVS1 (nt 354 to 468 [16]; see also reference 42). The Rep-binding motifs of both P1 and AAV ITR are outlined and labeled. The sequences homologous to the TRS of AAV are outlined in both the ITR se-quence and P1. The endonuclease cleavage site is between the thymidines within the TRS box and is indicated by the arrow.
FIG. 1. Replication model for AAV DNA. Thicker lines represent newly synthesized DNA, and the arrowheads indicate directions of synthesis. (A) Input viral DNA; (B) elongation by a cellular replication complex; (C) Rep protein(s) binding to the recognition site within the ITR and nicking one strand of the duplex; (D and E) extension of the ITR by cell replication complex; (F) produc-tion of two strands of the viral genome. Each strand consists of newly synthesized and parental DNA. Terminal resolution is represented by steps C, D, and E.
on November 9, 2019 by guest
http://jvi.asm.org/
and [a-32
P]dCTP. Following polyacrylamide gel electrophoresis (PAGE) purifi-cation, the substrates were digested with either PstI or KpnI. Digestion with PstI resulted in EcoRI end-labeled products, while KpnI digestion yielded HindIII end-labeled probes. The end-labeled probes were purified by phenol-chloroform extraction and ethanol precipitation. MBP-Rep78 (1.4mg) was incubated with the probes in a final volume of 25ml of core buffer (12 mM Tris-Cl [pH 7.4], 40 mM NaCl, 1 mM EDTA, 0.1% Triton X-100 [wt/vol], 1 mM dithiothreitol, 4% glycerol [wt/vol]; 2.5 mM ATP and 25 mM MgCl2were added as indicated). The probes were added ('15,000 cpm) and incubated for 30 min at 308C. The reaction was stopped with an equal volume of 23SDS gel loading buffer.
The products of this reaction were run on a 10% denaturing SDS gel to detect covalent bond formation between protein and P1 probe.
[image:3.612.67.548.63.596.2]Synthetic oligonucleotides.Oligonucleotides containing 57 bp of P1 sequence FIG. 3. Rep requirement in trans for replication. Template requirement for
stimulation of [32
P]dCMP incorporation into replication products. Both func-tional Rep protein and a sequence element that functions as a Rep-dependent ori are necessary for specific radiolabeling of the template. (A) Replication assay mixtures containing 0.3mg of either supercoiled pAV2 (9.0 kb [lanes 1, 2, and 3]), pMAT50 (2.8 kb [lanes 4, 5, and 6]), pHisD (11.0 kb [lanes 7, 8, and 9]), or pUC19 (2.7 kb [lane 10]) as templates were processed as described in Materials and Methods. Rep proteins were included as indicated. N, MBP-Rep68DNTP; 68, MBP-Rep68D; 78, MBP-Rep78. Size markers are in kilobases (M). The open-circular (O) or linear (L) forms of pMAT50 are indicated. (B) Replication assay mixtures containing 0.3mg of pSVoriAAV (7.4 kb [lanes 1, 2, and 3]) or pAV2 (9.0 kb [lanes 4, 5, and 6]) were processed as described in Materials and Methods. The linear (L) and rescued (R) products of pAV2 are indicated. The 4.7-kb rescued product is derived from the AAV moiety of pAV2, and the 4.2-kb product corresponds to the vector moiety.
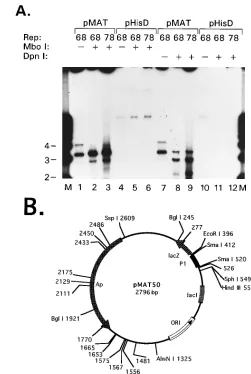
FIG. 4. (A) Extent of DNA synthesis determined by DpnI resistance. Repli-cation reactions were performed as described in Materials and Methods by using either pMAT50 or pHisD as the template. Replication reactions that included MBP-Rep68D(68) or MBP-Rep78 (78) are as indicated. Samples were digested with 60 U of DpnI (New England Biolabs) in 0.21 M NaCl for 12 h at 378C or 10 U of MboI (New England Biolabs) for 3 h in reaction buffer supplied by the manufacturer. Digested samples are indicated by1, no enzyme treatment is indicated by2. Lane M, 1-kb ladder. Sizes are indicated in kilobases. The appropriate enzyme concentrations were determined by analyzing the digestion products with increasing amounts of enzyme after ethidium bromide agarose gel electrophoresis (data not shown). pMAT50 processed from replication reactions without [a-32P]dCTP was used as the substrate for standardization of DpnI digestion. The substrate for MboI standardization was bacteriophage lambda DNA produced in dam mutant E. coli (data not shown). (B) Circular map of pMAT50 with the locations of DpnI-MboI sites indicated by numbers alone. The recognition sites for the following endonucleases are shown: AlwNI, BglI, HindIII, EcoRI, SphI, and SspI. The P1 element is represented by the filled thick line flanked by SmaI sites (nt 412 to 520). The positions ofb-lactamase gene (Ap), plasmid ori, and lacZ ORF are labeled.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.300.551.69.443.2]were synthesized and PAGE purified (Midland Certified Reagant Company). The P1 sequence contained both the Rep-binding motif (GCTC)3and the 5-bp sequence homologous to the AAV terminal resolution site (TRS) site. Three oligonucleotides were utilized for covalent attachment and endonuclease activity reactions. NP-1 is complementary to NP-3. NP-2 is a 39-terminal deletion of NP-3. The sequences are as follows: NP-1, 59-TACGTCCCGCCCGCCCAGCGAGC GAGCGAGCGCCGAGCCCCAACCGCCGCCACCAGTCATG; NP-2, 59-CA TGACTGGTGGCGGCGGTTGGGGCTCGGCGCTCGCTCGCTCGCTGG GCGGGCGGGA; NP-3, 59-CATGACTGGTGGCGGCGGTTGGGGCTCGG CGCTCGCTCGCTCGCTGGGCGGGCGGGACGTA.
The duplex oligonucleotide NP-1–NP-2 was uniquely 39end labeled by filling in a 4-base 59overhang with Klenow and [a-32P]dCTP. Unincorporated de-oxynucleoside triphosphates were removed by a G-50 Sephadex spin column, and the oligonucleotides were purified by phenol-chloroform extraction and ethanol precipitation. A total of 120,000 cpm of probe (specific activity, 2.2 3107 cpm/mg) per reaction was used. NP-3 (1mg) was 59end labeled with [g-32P]ATP (3,000 Ci/mmol) and T4 polynucleotide kinase (New England BioLabs) using the reaction buffer that was supplied. Unincorporated ATP was removed by a G-50 Sephadex spin column.
Covalent attachment of MBP-Rep78 to synthetic oligonucleotides.The cova-lent attachment of MBP-Rep78 to the oligonucleotides was performed as de-scribed elsewhere (21), except that the reaction was carried out at 378C for 10 min. The amounts of MBP-Rep78 ranged from 0.028 to 0.14mg.
Endonuclease cleavage site determination.The endonuclease cleavage site was mapped by methods previously described (6).32
P-59-end-labeled oligonu-cleotide NP-3 was annealed with unlabeled NP-1. A 20-ng amount of duplex oligonucleotides was used in each reaction. The reaction mixtures were incu-bated for 30 min at 378C. The reactions were terminated and deproteinated by the addition of 40ml of stop buffer (10 mM Tris-Cl [pH 7.9], 10 mM NaCl, 0.5% SDS, 0.2 mg of yeast tRNA per ml, 20 mM EDTA, 2 mg of proteinase K per ml). The reaction mixtures were incubated for 30 min at 378C. The nucleic acids were extracted with phenol-chloroform and ethanol precipated. The dried samples were resuspended in gel loading buffer (80% formamide, 0.025% bromophenol blue, 0.025% xylene cyanole FF) and were denatured by heating to 808C. The samples were fractionated electrophoretically on an 8% polyacrylamide–8 M urea sequencing gel.
RESULTS
Rep68 and HeLa cellular factors catalyze P1-dependent
DNA replication.
The 110-bp human chromosome 19 element,
P1 (Fig. 2), binds specifically to wild-type Rep68 (42) and to
recombinant Rep protein produced as a fusion protein in
Esch-erichia coli (MBP-Rep78 or MBP-Rep68
D
) (6). To test
[image:4.612.134.482.71.309.2]whether binding of Rep to human DNA may facilitate
integra-tion of AAV DNA into human chromosome 19, the P1
ele-ment cloned into pUC19, pMAT50 (42), was used as a
tem-plate in an in vitro replication assay. Cellular extract served as
the source of DNA polymerases and other replication factors
(40). Template pAV2 served as a positive control for
Rep-specific replication and rescue (6, 41). Using template
contain-ing the P1 element and either MBP-Rep68
D
or MBP-Rep78,
two radiolabeled bands comigrating with linear and open
cir-cular forms of pMAT50 were detected (Fig. 3A, lanes 5 and 6).
The differences in the amounts of radiolabel incorporated in
reactions with either MBP-Rep68
D
or MBP-Rep78 (Fig. 3)
varied among different reactions and between different extracts
and may not reflect distinctions in activity between the two
recombinant proteins. In contrast, MBP-Rep68
D
NTP resulted
in relatively little incorporation of [
32P]dCMP (Fig. 3A, lanes
1, 4, and 7). The nucleoside triphosphate (NTP)-binding
mu-tant of MBP-Rep68
D
has been previously characterized and
shown to bind to the ITR and
D
ITR but not to have helicase or
endonuclease activities associated with wild-type Rep (6). The
activities of MBP-Rep68
D
NTP were found to be similar to
those of the NTP mutant of Rep68, K340H (26, 27). In the
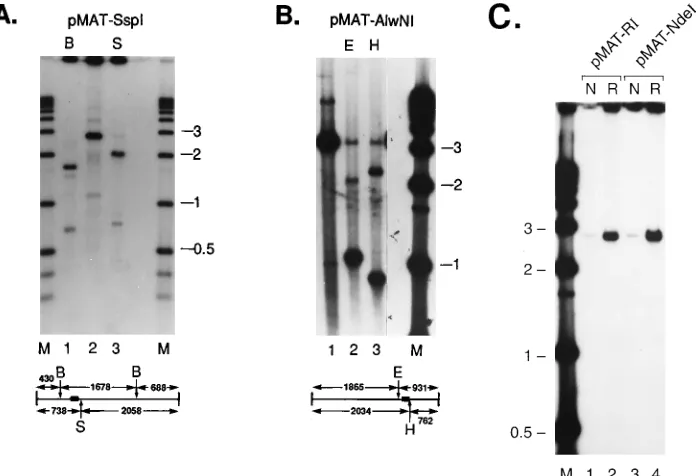
FIG. 5. Replication using linearized plasmids as substrates. pMAT50 was linearized prior to incubation with MBP-Rep68D. Following extraction of protein, the DNA was digested with the restriction enzymes indicated. (A) pMAT50 linearized with SspI. Lanes: 1, postreplication digestion with BglI (B); 2, no postreplication digestion; 3, postreplication digestion with SphI (S); M, 1-kb size markers (Gibco-BRL) that were 39end labeled. A diagram of SspI-linearized pMAT50 is shown underneath the autoradiograph, with the relative positions of the BglI and SphI sites indicated. The 110-bp P1 element is represented by the filled rectangle. SspI and BglI digestion of pMAT50 generated three fragments, respectively, of 1,678, 688, and 430 bp. The 430-bp fragment remained unlabeled. SspI and SphI digestion produced two fragments, respectively, of 2,058 and 738 bp. Both fragments were labeled. (B) pMAT50 linearized with AlwNI prior to the replication reaction. Lanes: 1, no postreplication digestion; 2, postreplication digestion with EcoRI (E); 3, postreplication digestion with HinDIII (H); M, 1-kb size markers (Gibco-BRL) that were 39end labeled. The diagram beneath the autoradiograph indicates the relative positions of EcoRI-, HindIII-, and AlwNI-cut sites. The position of the P1 element is represented by the filled rectangle. (C) MBP-Rep68Drequirement for the replication of pMAT50. The substrate, pMAT50, was linearized prior to the replication reaction with either EcoRI (lanes 1 and 2) or NdeI (lanes 3 and 4). MBP-Rep68DNTP was included in the reaction mixtures labeled N; MBP-Rep68Dwas includedin the lanes labeled R. Lane M, 1-kb size markers (Gibco-BRL) that were 39end labeled.
on November 9, 2019 by guest
http://jvi.asm.org/
presence of either MBP-Rep68
D
or MBP-Rep78 and a
plas-mid containing wild-type AAV DNA, both rescued and
circu-lar forms of pAV2 were detected (Fig. 3A, lanes 2 and 3).
The requirement for Rep and either the AAV ITR or the P1
element for specific replication is in contrast to results
ob-tained using a DNA sequence lacking the P1 element or AAV
ori (pHisD) (28). Very little incorporation was detected in
reactions with pHisD as a template and in which either
MBP-Rep68
D
NTP, MBP-Rep68
D
, or MBP-Rep78 was added (Fig.
3A, lanes 7, 8, and 9, respectively). The labeling of pMAT50
was dependent on the addition of MBP-Rep68
D
or
MBP-Rep78 to the reaction. Either pUC19 (Fig. 3A, lane 10) or
pBluescript (data not shown) (7) was unable to serve as a
template. As an additional control for Rep effects on known
oris, plasmid pSVoriAAV was used as a template for the in
vitro replication reaction (6). This plasmid consists of the SV40
ori cloned into a plasmid containing an AAV genome lacking
both ITRs. Plasmid pAV2 consists of the wild-type AAV
ge-nome. Little or no specific incorporation of [
32P]dCMP was
detected with pSVoriAAV template reactions (Fig. 3B, lanes 1,
2, and 3). The presence of two viral ITRs in pAV2 resulted in
radiolabeled linear and rescued products as previously
de-scribed (11, 40, 41). These results indicate that Rep-dependent
replication is likely to involve DNA recognition by Rep.
Fur-ther support of specificity is provided by the negative results
obtained with a larger plasmid (pHisD) (Fig. 3A, lanes 7, 8,
and 9) or a heterologous origin of replication derived from
SV40 DNA (pSVoriAAV) (Fig. 3B, lanes 1, 2, and 3).
Esti-mating the extent of radiolabel incorporated into pAV2 and
pMAT50 indicates that the P1 element serves almost as
effi-ciently as the two viral oris in pAV2 (Fig. 3A, lanes 2 and 5 and
lanes 3 and 6). This is consistent with a previous report that
demonstrated that a plasmid containing the AD
9
portion of the
AAV ITR (Fig. 2) stimulated incorporation of [
32P]dCMP into
acid-precipitable material with similar efficiency as pAV2 (6).
That is, using the levels of P-32 incorporation as an indicator of
DNA synthesis, a nonhairpinned ori element is utilized with
results similar to those with a hairpinned ori.
pMAT50 replication products are
Dpn
I and
Mbo
I resistant.
The extent of DNA synthesis can be established by resistance
to DpnI endonuclease. This has much greater activity when
both adenosines in the recognition sequence are methylated.
Such methylation is not performed by eukaryotic methylases.
Therefore, conversion to DpnI resistance indicates that
hemi-methylation or lack of hemi-methylation is a result of one or two
rounds of DNA replication in a eukaryotic host, respectively,
whereas the restriction endonuclease MboI will cleave DNA
only when both adenosines in the enzyme recognition site are
unmethylated. Thus, two rounds of replication will yield
radio-labeled products that are DpnI resistant and MboI sensitive.
However, one round of replication generates radiolabeled
products that are both DpnI and MboI resistant as a result of
hemimethylation of the duplex DNA. The results indicate that
a band of mobility corresponding to the open circular form
(
'
3.2 kb) of pMAT50 is resistant to either MboI (Fig. 4A,
lanes 2 and 3) or DpnI (Fig. 4A, lanes 8 and 9). Linear forms
of pMAT50 appear to be produced as a consequence of
diges-tion with either endonuclease. The locadiges-tions of DpnI-MboI
recognition sites are unsymmetrically distributed around
pUC19, and no sites are within the P1 element inserted into
the unique Sma site (Fig. 4B). The simplest explanation for the
formation of linear products after DpnI digestion is that DNA
synthesis proceeds at least 80% around the circular template
and terminates prior to completion of the full-length product.
The DpnI site at position 277 would then be bimethylated and
sensitive to cleavage. Similarly, the minor amount of linear-size
product present in the MboI-treated samples could be
ex-plained by inefficient second-round synthesis terminating
within 1 kb of initiation. The open circular form of the 3.6-kb
band was confirmed by restriction endonuclease digestion of
the isolated product.
pMAT50 replication is asymmetric.
To determine the
direc-tionality of pMAT50 replication, linearized templates were
utilized. The template pMAT50 was digested to completion
with either SspI (Fig. 5A) or AlwNI (Fig. 5B) (shown
diagram-matically in Fig. 4B). Following the replication reaction, the
SspI-linearized DNA was digested with BglI or SphI, and the
AlwNI-linearized template was digested with HindIII or EcoRI
FIG. 6. (A) Labeled-strand specificity determined by hybridization to single-stranded DNA. Approximately 100 ng of DNA was applied to each slot and adsorbed to a Zeta-Probe membrane (Bio-Rad). Each strip contains three slots: duplex pBluescript SK (2) (pBst), single-stranded pBluescript SK (1), and (2) DNA.32
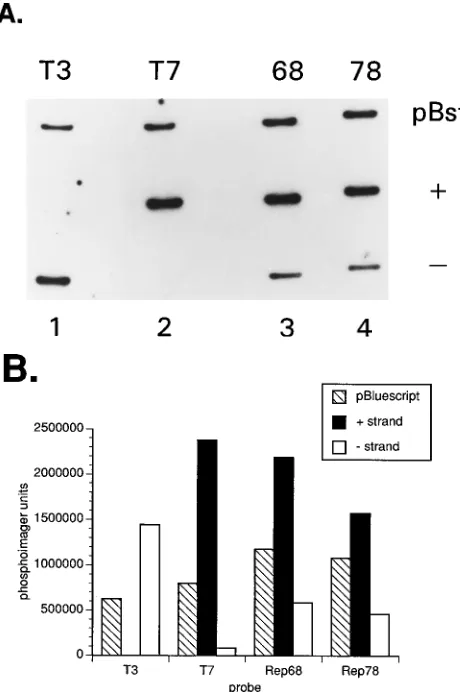
P-59-end labeled oligonucleotides specific to the T3 or T7 promoters were used as probes to confirm strand specificity. T3, 59-ATTAACCCTCACTA AAGGGA; T7, 59-TAATACGACTCACTATAGGG (strips 1 and 2, respective-ly). Open-circular DNA radiolabeled in the replication reaction was agarose gel purified, electroeluted, and used as a hybridization probe. Probes derived from MBP-68Dand MBP-Rep78 were used for hybridizations of strips 3 and 4. E. coli DH11S (Gibco-BRL) transformed with either pBluescript SK (1) or (2) were infected with helper phage M13KO7 to produce single-stranded pBluescript. The hybridization medium consisted of 7% SDS, 0.1 M NaPi(pH 7.4), 1 mM EDTA, and 0.1 mg of denatured salmon sperm DNA per ml. The high-stringency wash solution was 0.23SSC (13SSC is 0.15 M NaCl plus 0.015 M sodium citrate)– 0.1% SDS. The T3 and T7 probes were hybridized at 428C, and the strips were washed at room temperature. The MBP-Rep68Dand MBP-Rep78 probes were hybridized at 658C, and the strips were washed at 658C. (B) Measurement of radioactivity associated with the samples shown in panel A. Activities were determined by phosphoimaging, and the results are graphically displayed.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.62.292.72.418.2](Fig. 5). The pattern of labeled fragments from each set can
best be explained by predominantly unidirectional replication
originating from within the P1 element and proceeding to the
5
9
end of the template strand. The BglI-digested products of
the SspI-linearized template resulted in radiolabeling two of
the three fragments. The unlabeled fragment of 430 bp is
located proximally to the SspI site. Digestion with SphI
gener-ated two fragments, of 2,058 and 738 bp, and both fragments
were labeled. The lack of incorporated label into the 430-bp
SspI-BglI fragment, together with the labeled 738-bp SspI-SphI
fragment, indicates that replication most likely originates from
the region between the BglI and SphI sites.
Results consistent with these data were obtained with
Al-wNI-linearized substrate (Fig. 5B). The replication products
were digested with either HindIII or EcoRI, which have unique
sites flanking the P1 insert. The predominantly labeled smaller
fragment indicates a strong bias in replication direction. The
relatively low amount of label incorporated into the larger
fragment may be the result of nonspecific or repair activity.
The specificity of linearized DNA replication for
MBP-Rep68
D
is demonstrated by the inability to use
MBP-Rep68
D
NTP (Fig. 5C) or MBP-Lacz (data not shown). These
proteins provide a control for the presence of copurified
bac-terial proteins which might have activity on the substrates in
the reaction. These results imply that binding per se of
MBP-Rep68
D
or MBP-Rep78 to a functional ori is not sufficient for
replication. Thus, the incorporation of [
a
-
32P]dCMP into
plas-mid-size DNA is dependent on the presence of the P1
se-quence in cis and MBP-Rep68
D
in trans.
Corroborating evidence for asymmetrical replication is
pro-vided by the use of the labeled open-circular replication
prod-ucts as probes against single-stranded DNA derived from
res-cued pBluescript SK(
1
) or (
2
). The hybridization results
shown in Fig. 6 indicate that there is preferential labeling of
one strand of DNA. The T3 and T7 probes demonstrate the
specificity of the target DNA (Fig. 6A, lanes 1 and 2). The
radiolabeled DNA derived from the MBP-Rep68
D
- and
MBP-Rep78-containing reaction mixtures both hybridized
preferen-tially to the plus strand of pBluescript. PhosphorImager
anal-ysis of the gel established that the hybridization signal to the
plus strand is approximately 2.5-fold greater than the signal to
the minus strand (Fig. 6B). The observed bias in the direction
of replication is likely to be a low estimate, since labeling of
pMAT50 by nonspecific repair activities contributes to the
hybridization signal of both the plus and the minus strands.
Labeling of minus strands of pMAT50 is as predicted from the
results obtained with the SspI- or AlwNI-linearized pMAT50
templates.
Rep68 binding is accompanied by site-specific endonuclease
activity and 5
*
-covalent attachment.
The mechanism of
termi-nal resolution of the AAV ITR requires that Rep cleave one
strand of duplex DNA at a defined site within the ITR that is
a fixed distance from the Rep-binding motif. If replication of
pMAT50 is initiated by a similar mechanism, then formation of
a covalent intermediate of MBP-Rep68
D
–DNA would be
pre-dicted. Such a protein-DNA complex would be stable upon
SDS-PAGE and would be distinct from free probe or
nonco-FIG. 7. Covalent linkage activity of MBP Rep protein to P1. (A) Diagram ofkey features of P1 sequence. The Rep-binding motif (GCTC)3and the region homologous to the AAV TRS are indicated. (B) Radiolabeled P1 probes. Probes were prepared as follows. The EcoRI-to-HindIII fragment, which includes the P1 element, was radiolabeled with32P on either strand at the HindIII site (59
HindIII [59D3] or 39HindIII [39D3]). The probes were incubated in core buffer (27) with either MBP-Rep78 or MBP-Rep68DNTP. ATP and MgCl2were in-cluded when indicated. The positions of free probe and probe covalently at-tached to MBP-Rep78 are indicated. The products were fractionated by electro-phoresis on a 10% polyacrylamide gel containing SDS. (C) Effect of increasing protein concentration on covalent bond formation between MBP-Rep78 and P1 oligonucleotide. Synthetic complementary oligonucleotides containing 57 bp of P1 sequence were uniquely 39end labeled and examined for their ability to form a covalent bond with MBP-Rep78 as described elsewhere (see reference 8 and Materials and Methods). The products were fractionated on a 4 to 20% gradient polyacrylamide gel containing SDS (Bio-Rad). The positions of the free probe P1 and the covalent Rep78:P1 complex are indicated. Lanes: 1, no MBP-Rep78; 2, empty lane; 3 to 9, increasing concentrations of MBP-Rep78 as fol-lows: 0.0068 (lane 3), 0.0136 (lane 4), 0.028 (lane 5), 0.056 (lane 6), 0.0840 (lane 7), 0.112 (lane 8), and 0.140 (lane 9) mg. Molecular mass standards are in kilodaltons. (D) No added Rep control. Each end of the fragment diagrammed in panel A was uniquely end labeled at the positions indicated. RI, EcoRI site; D3, HindIII site. The reaction mixtures were treated with or without proteinase K as indicated (0.4% SDS–0.74 mg of proteinase K per ml at 558C for 20 min). The reaction mixtures were incubated for 30 min at 378C and fractionated on a 10% polyacrylamide gel containing 0.1% SDS. The positions of the free probes
are shown. Positions of molecular mass standards are on the right in kilodaltons. Following electrophoresis, the gel was fixed, dried, and exposed for autoradiog-raphy. (E) Experiment similar to that described in panel D, except that 0.8mg of MBP-Rep78 was included in each reaction. Following incubation, half of the reaction mixture was treated with proteinase K when indicated. The mobilities of the free probes and complexed probes are indicated. Molecular mass standards are in kilodaltons.
on November 9, 2019 by guest
http://jvi.asm.org/
valent protein-DNA complexes. The cloned P1 element was
excised from pUC19 and uniquely end labeled for use as a
substrate in the TRS endonuclease assay (Fig. 7A). The
lower-mobility band consists of 3
9
-end-labeled DNA covalently
linked to protein according to a previous characterization (7)
(Fig. 7B). Complementary oligonucleotides that corresponded
to the P1 sequence were synthesized (see Materials and
Meth-ods) and used to determine whether the TRS endonuclease
activity was responsive to increasing MBP-Rep78
concentra-tions. The extent of covalent protein-DNA production was
proportional to the the concentration of MBP-Rep78 in the
system (Fig. 7C). The cleavage site may correspond to a 5- out
of 6-nt match of the TRS of the AAV ITR.
The reaction mixtures were treated with proteinase K to
ascertain whether the lower-mobility bands that were
identi-fied as complex (Fig. 7B and C) were composed of
protein-DNA. The strand-specific complex that formed with the 3
9
-end-labeled HindIII site was sensitive to the protease
treatment (Fig. 7E [sample 3
9
D3]), whereas proteinase
treat-ments of reactions with no added Rep were unaffected (Fig.
7D [sample 3
9
D3]). These results are consistent with previous
data that established that covalent linkage of Rep to DNA
occurs as a consequence of endonuclease activity (see below)
(7, 13).
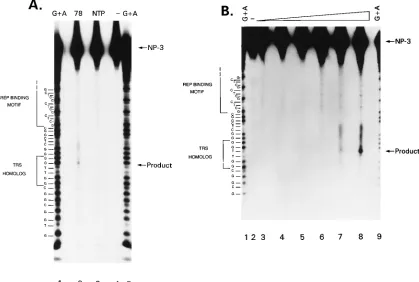
Site-specific endonuclease activity of MBP-Rep78 on P1.
If
Rep initiated cellular DNA synthesis by a mechanism
analo-gous to that of AAV DNA replication, then a critical early step
[image:7.612.97.516.74.356.2]in this process is nicking a single strand of DNA at a defined
site. Previous studies have demonstrated that MBP-Rep
pro-teins could bind to linear duplex substrates with affinities
sim-ilar to those of hairpinned ITR substrates (7). However, the
endonuclease activity of Rep was 50- to 100-fold less efficient
on the nonhairpinned substrates (6). To test whether
MBP-Rep could nick at a site within the P1 element, oligonucleotide
NP-3 was 5
9
end labeled, annealed to unlabeled NP-1, and
incubated with MBP-Rep78, MBP-Rep68
D
NTP (Fig. 8A,
lanes 2 and 3), or MBP-Rep68
D
(data not shown). The
posi-tion of the full-length NP-3 is indicated. A purine-specific
sequencing ladder (Fig. 8A, lanes 1 and 5) allows precise
map-ping of the cleavage products. The most prominent band
pro-duced by incubation with MBP-Rep78 is indicated by the
ar-row labeled product. A fragment of this size could be produced
by cleavage of NP-3 within the TRS homolog motif and
cor-responds to cleavage predominantly between the two
thymi-dines. A fragment of this size is not observed either with the
NTP mutant or with no added protein (Fig. 8A, lanes 3 and 4).
Similar reactions were performed with increasing amounts
of MBP-Rep78 (Fig. 8B). The intensity of the product band is
proportional to the amount of MBP-Rep78 in the reaction.
Other cleavage products become more pronounced also as the
concentration of Rep increases. This imprecision may be an
inherent property of Rep or may be due to the nature of the
Rep protein that is employed, i.e., Rep that is bacterially
ex-pressed as a fusion protein.
FIG. 8. Rep endonuclease cleavage of P1.32
P-59-end-labeled oligonucleotide NP-3 was annealed with unlabeled NP-1 (see Materials and Methods). Approximately 20 ng of duplex oligonucleotides was used as a substrate in each 20-ml reaction mixture. Following incubation, the reaction mixtures were processed as described and fractionated on an 8% sequencing gel. Following electrophoresis, the gel was fixed, dried, and exposed to X-ray film for autoradiography. The positions of full-length substrate and cleavage product are indicated by the arrows labeled NP-3 and product. The sequence of the oligonucleotide is on the left, with the Rep-binding motif and TRS homolog bracketed. (A) Approximately 0.7mg of the indicated protein was added. Lanes: 1 and 5, purine-specific sequencing reaction; 2, MBP-Rep78; 3, MBP-Rep68DNTP; 4, no added protein. (B) Dose response of endonuclease activity. Increasing amounts of MBP-Rep78 correspond to amounts of product generated. The reactions were performed as described for panel A. Lanes: 1 and 9; purine-specific sequencing reaction; 2, no added protein; 3 to 8, 0.09, 0.18, 0.4, 0.7, 1.4, and 2.8mg, respectively.
on November 9, 2019 by guest
http://jvi.asm.org/
DISCUSSION
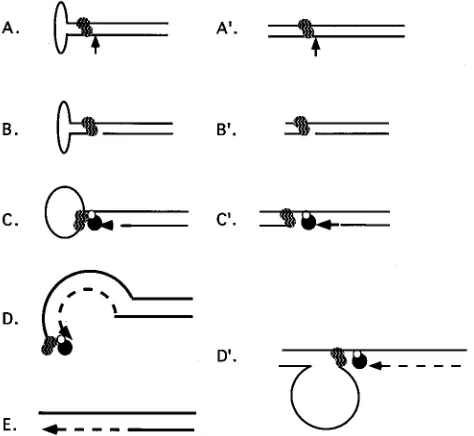
The identification of a Rep-responsive, human ori provides
the basis for a model of DNA leading-strand synthesis. A
process involving unidirectional replication neatly dissects two
complex and interrelated replication reactions, i.e.,
leading-and lagging-strleading-and DNA synthesis. The model for Rep68- or
Rep78-dependent replication of DNA containing the P1 (or
D
ITR) origin is derived from the known in vitro activities of
the Rep proteins (Fig. 9). These include sequence-specific
binding (6, 7, 42), strand- and site-specific endonuclease
activ-ity (12), helicase activactiv-ity (12), and stimulation of replication in
vitro from an AAV ori (6, 24). The Rep-binding motif has been
determined and occurs in both the AAV ITR and P1 (Fig. 2)
(6, 42). A dissociation constant of
'
10
210M has been
deter-mined for MBP-Rep68
D
binding to hairpin wild-type ITR or
linear
D
57ITR (7). DNA synthesis initiates from the 3
9
-OH
generated by the endonuclease activity of Rep at the TRS
within P1 independently of RNA priming. This is analogous to
AAV DNA replication. The asymmetry of the TRS with
re-spect to the Rep-binding motif constrains elongation from a
single site on one strand, i.e., unidirectional. This model
pre-dicts displacement of the nontemplate strand as the replication
complex proceeds (Fig. 9D
9
). These replication intermediates
were not detected, presumably because of the sensitivity of
single-stranded DNA to nucleases in the cell extract.
The cleavage at the TRS involves the formation of a stable
Rep-thymidine:3
9
-DNA intermediate (13). The
ATP-depen-dent helicase activity of Rep68 and Rep78 (12) may obviate the
need for cellular helicases to promote unwinding of the DNA
for elongation. The covalently attached Rep molecule is then
released by an undetermined mechanism. The nucleoprotein
intermediate is stable, as demonstrated by in vitro assays (13),
although Rep has not been detected in the mature AAV virion.
The model of AAV Rep protein-mediated replication of
pMAT50 appears analogous to rolling-circle replication
mod-els described for Staphylococcus aureus plasmid pT181 (35) or
bacteriophage
f
X174 (10). The parallel with staphylococcal
pT181 replication is so striking that the components of the
models are interchangeable. The staphylococcal RepD protein
has origin specific-binding activity and single-strand,
site-spe-cific endonuclease activity that results in covalent attachment
of RepD to the 5
9
end of the nick via a phosphotyrosine
linkage. Unidirectional replication initiates from the free
3
9
-OH and proceeds around the plasmid. The staphylococcal
RepD protein may remain covalently attached to the 5
9
end of
the displaced strand throughout replication. The similarity to
the bacterial system provides a precedent for AAV Rep68- or
Rep78-dependent replication of a circular template. The many
similarities between the two systems suggest independent
con-vergent evolution of this mode of replication. Alternatively,
evolutionary conservation of protein functions could account
for the similarities observed.
Targeted integration of AAV DNA into human chromosome
19.
AAV DNA integrates into a small, defined region of
hu-man chromosome 19 in cultured cells at a frequency of
ap-proximately 70% (19, 30). The integration junctions are
dis-tinct at the molecular level among the latently infected cell
lines analyzed. Comparison of AAV DNA sequence with
AAVS1 sequence confirmed that integration occurred by
non-homologous recombination (17, 30). The interaction of Rep
with either chromosome 19-derived DNA (42) or AAV ITR (6,
12, 13, 26) strengthens the argument for Rep involvement in
targeted integration. The data presented here indicate that
Rep is required and allow a model to be formulated for
tar-geted integration. The process of AAV DNA integration now
appears to be the result of limited DNA synthesis initiated by
binding of Rep to the P1 element within AAVS1. The
recom-bination mechanism may involve subunit exchange between
Rep complexes associated with each substrate. The
Rep-bind-ing motif has been found within other genes (GenBank release
80). However, the requirement for a properly positioned TRS
would decrease the probability of occurrence in random
se-quence to
#
6
3
10
211, thereby defining a unique sequence.
A recombination model involving limited DNA synthesis is
supported by previous descriptions of proviral structures (19,
30, 31). For some latently infected cell lines, duplication of
cellular sequences adjacent to the provirus has been described.
Significantly, all of the characterized integration events were
asymmetrically distributed with respect to the P1 element. This
recombination model may constitute a new recombination
pathway that utilizes functions intrinsic to the cell.
In vitro reconstitution of DNA synthesis using cloned P1 or
D
ITR as template, MBP-Rep, and purified cellular proteins
may be possible. The large quantities of MBP-Rep available
provide an opportunity to identify cellular components
in-volved in leading-strand synthesis without the additional
com-plication of lagging-strand synthesis. The involvement of Rep
in specialized cellular DNA synthesis provides the basis for a
targeted integration model of AAV DNA integration into
hu-man chromosome 19.
ACKNOWLEDGMENTS
We especially thank Mark Challberg, Charlotte McGuinness, Ken-neth Berns, Bernard Moss, and Nancy Nossal for helpful discussions and comments. We are grateful to Jay Chiorini, Roland Owens, and Sirrka Kyo¨stio¨ for critical reading and suggestions that were useful in the preparation of the manuscript.
[image:8.612.60.297.72.290.2]This work was supported in part by NHLBI CRADA 91-02 with Genetic Therapy, Inc., Gaithersburg, Md.
FIG. 9. Comparison of AAV terminal resolution with a model of Rep-de-pendent plasmid replication. (A to E) Terminal resolution of the AAV genome; (A9to E9) Rep-dependent replication from a non-ITR origin, e.g., P1; (A and A9) Rep binding to its recognition sequence adjacent to a properly positioned endonuclease site (vertical arrow); (B and B9) single-stranded cleavage at the endonuclease site and covalent attachment of Rep to the 59end of the nick; (C and C9) assembled Pol complex (closed and open circles) extending the nontem-plate strand from the 39-OH of the nick; (D) extension of the ITR by DNA leading-strand synthesis; (D9) extension of the nonviral origin by leading-strand DNA synthesis.
on November 9, 2019 by guest
http://jvi.asm.org/
REFERENCES
1. Bergsma, D. J., D. M. Olive, S. W. Hartzell, and K. N. Subramanian. 1982. Territorial limits and functional anatomy of the simian virus 40 replication origin. Proc. Natl. Acad. Sci. USA 79:381–385.
2. Berns, K. I. 1990. Parvovirus replication. Microbiol. Rev. 54:316–329. 3. Brewer, B. J. 1994. Intergenic DNA and the sequence requirements for
replication initiation in eukaryotes. Curr. Opin. Genet. Dev. 4:196–202. 4. Carter, B. J. 1992. Adeno-associated virus vectors. Curr. Opin. Biotechnol.
3:533–539.
5. Challberg, M. D., and T. J. Kelly. 1989. Animal virus DNA replication. Annu. Rev. Biochem. 58:671–717.
6. Chiorini, J. A., M. D. Weitzman, R. A. Owens, E. Urcelay, B. Safer, and R. M.
Kotin.1994. Biologically active Rep proteins of adeno-associated virus type 2 produced as fusion proteins in Escherichia coli. J. Virol. 68:797–804. 7. Chiorini, J. A., S. M. Wiener, S. R. M. Kyo¨stio¨, R. A. Owens, R. M. Kotin,
and B. Safer.1994. Sequence requirements for stable binding and function of Rep68 on the adeno-associated virus type 2 inverted terminal repeats. J. Virol. 68:7448–7457.
8. Collins, K. L., A. A. Russo, B. Y. Tseng, and T. J. Kelly. 1993. The role of the 70 kDa subunit of human DNA polymerase alpha in DNA replication. EMBO J. 12:4555–4566.
9. DeLucia, A. L., B. A. Lewton, R. Tjian, and P. Tegtmeyer. 1983. Topography of simian virus 40 A protein-DNA complexes: arrangement of pentanucle-otide interaction sites at the origin of replication. J. Virol. 46:143–150. 10. Eisenberg, E., J. F. Scott, and A. Kornberg. 1976. An enzyme system for
replication of duplex circular DNA: the replicative form of phagefX174. Proc. Natl. Acad. Sci. USA 73:1594–1597.
11. Hong, G., P. Ward, and K. I. Berns. 1992. In vitro replication of adeno-associated virus DNA. Proc. Natl. Acad. Sci. USA 89:4673–4677. 12. Im, D.-S., and N. Muzyczka. 1990. The AAV origin binding protein is an
ATP-dependent site-specific endonuclease with DNA helicase activity. Cell
61:447–457.
13. Im, D.-S., and N. Muzyczka. 1989. Factors that bind to adeno-associated virus terminal repeats. J. Virol. 63:3095–3104.
14. Jones, K. A., and R. Tjian. 1984. Essential contact residues within SV40 large T antigen binding sites I and II identified by alkylation-interference. Cell
36:155–162.
15. Kelly, T. J. 1991. DNA replication in mammalian cells: insights from the SV40 model system. Harvey Lect. 85:173–188.
16. Kelman, Z., and M. O’Donnell. 1994. DNA replication: enzymology and mechanisms. Curr. Opin. Genet. Dev. 4:185–195.
17. Kotin, R. M., R. M. Linden, and K. I. Berns. 1992. Characterization of a preferred site on human chromosome 19q for integration of adeno-associ-ated virus DNA by non-homologous recombination. EMBO J. 11:5071–5078. 18. Kotin, R. M., J. C. Menninger, D. C. Ward, and K. I. Berns. 1991. Mapping and direct visualization of a region-specific viral DNA integration site on chromosome 19q13-qter. Genomics 10:831–834.
19. Kotin, R. M., M. Siniscalco, R. J. Samulski, X. Zhu, L. Hunter, C. A.
Laughlin, S. McLaughlin, N. Muzyczka, M. Rocchi, and K. I. Berns.1990. Site-specific integration by adeno-associated virus. Proc. Natl. Acad. Sci. USA 87:2211–2215.
20. Laughlin, C. A., J. D. Tratschin, H. Coon, and B. J. Carter. 1983. Cloning of infectious adeno-associated virus genomes in bacterial plasmids. Gene 23: 65–73.
21. Li, J. J., and T. J. Kelly. 1985. Simian virus 40 DNA replication in vitro: specificity of initiation and evidence for bidirectional replication. Mol. Cell. Biol. 5:1238–1246.
22. Li, J. J., K. W. Peden, R. A. Dixon, and T. Kelly. 1986. Functional organi-zation of the simian virus 40 origin of DNA replication. Mol. Cell. Biol.
6:1117–1128.
23. Muzyczka, N. 1992. Use of adeno-associated virus as a general transduction
vector for mammalian cells. Curr. Top. Microbiol. Immunol. 158:97–129. 24. Ni, T.-H., X. Zhou, D. M. McCarty, I. Zolotukhin, and N. Muzyczka. 1994.
In vitro replication of adeno-associated virus DNA. J. Virol. 68:1128–1138. 25. Olivo, P. D., N. J. Nelson, and M. D. Challberg. 1988. Herpes simplex virus DNA replication: the UL9 gene encodes an origin binding protein. Proc. Natl. Acad. Sci. USA 85:5414–5418.
26. Owens, R. A., J. P. Trempe, N. Chejanovsky, and B. J. Carter. 1991. Adeno-associated virus rep proteins produced in insect and mammalian expression systems: wild-type and dominant-negative mutant proteins bind to the viral replication origin. Virology 184:14–22.
27. Owens, R. A., M. D. Weitzman, S. R. Kyo¨stio¨, and B. J. Carter.1993. Identification of a DNA-binding domain in the amino terminus of adeno-associated virus Rep proteins. J. Virol. 67:997–1005.
28. Reid, L. H., E. G. Shesley, H.-S. Kim, and O. Smithies. 1991. Cotransfor-mation and gene targeting in mouse embryonic stem cells. Mol. Cell. Biol.
11:2769–2777.
29. Samulski, R. J., L. S. Chang, and T. Shenk. 1987. A recombinant plasmid from which an infectious adeno-associated virus genome can be excised in vitro and its use to study viral replication. J. Virol. 61:3096–3101. 30. Samulski, R. J., X. Zhu, X. Xiao, J. D. Brook, D. E. Housman, N. Epstein,
and L. A. Hunter. 1991. Targeted integration of adeno-associated virus (AAV) into human chromosome 19. EMBO J. 10:3941–3950. (Erratum,
11:1228, 1992.)
31. Shelling, A. N., and M. G. Smith. 1994. Targeted integration of transfected and infected adeno-associated virus vectors containing the neomycin resis-tance gene. Gene Ther. 2:1–5.
32. Stillman, B., S. P. Bell, A. Dutta, and Y. Marahrens. 1992. DNA replication and the cell cycle. CIBA Found. Symp. 170:147–156.
33. Stillman, B. W., and Y. Gluzman. 1985. Replication and supercoiling of simian virus 40 DNA in cell extracts from human cells. Mol. Cell. Biol.
5:2051–2060.
34. Stow, N. D. 1982. Localization of an origin of replication within the TRs/IRs repeated region of the herpes simplex virus type 1 genome. EMBO J. 1:863– 867.
35. Thomas, C. D., D. F. Balson, and W. V. Shaw. 1990. In vitro studies of the iniation of Staphylococcal plasmid replication. J. Biol. Chem. 265:5519–5530. 36. Tjian, R. 1978. The binding site of SV40 DNA for a T-antigen related
protein. Cell 13:165–179.
37. Tsurimoto, T., and B. Stillman. 1991. Replication factors required for SV40 DNA replication in vitro. I. DNA structure-specific recognition of a primer-template junction by eukaryotic DNA polymerases and their accessory pro-teins. J. Biol. Chem. 266:1950–1960.
38. Tsurimoto, T., and B. Stillman. 1991. Replication factors required for SV40 DNA replication in vitro. II. Switching of DNA polymerase alpha and delta during initiation of leading and lagging strand synthesis. J. Biol. Chem.
266:1961–1968.
39. Waga, S., and B. Stillman. 1994. Anatomy of a DNA replication fork re-vealed by reconstitution of SV40 DNA replication in vitro. Nature (London)
369:207–212.
40. Ward, P., and K. I. Berns. 1991. In vitro rescue of an integrated hybrid adeno-associated virus/simian virus 40 genome. J. Mol. Biol. 218:791–804. 41. Ward, P., E. Urcelay, R. Kotin, B. Safer, and K. Berns. 1994.
Adeno-associated virus DNA replication in vitro: activation by a maltose binding protein/Rep 68 fusion protein. J. Virol. 68:6029–6037.
42. Weitzman, M. D., S. R. M. Kyo¨stio¨, R. M. Kotin, and R. A. Owens.1994. Adeno-associated virus (AAV) rep proteins mediate complex formation between AAV DNA and the human integration site. Proc. Natl. Acad. Sci. USA 91:5808–5817.
43. Wobbe, C. R., F. B. Dean, Y. Murakami, J. A. Borowiec, P. Bullock, and J.
Hurwitz.1987. In vitro replication of DNA containing either the SV40 or the polyoma origin. Philos. Trans. R. Soc. Lond. B Biol. Sci. 317:439–453.