Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Residues Critical for Duck Hepatitis B Virus Neutralization Are
Involved in Host Cell Interaction
CLAIRE SUNYACH,1CHRISTINE ROLLIER,1MAGDALENA ROBACZEWSKA,1,2CHRISTELLE
BOREL,1LUC BARRAUD,1ALAN KAY,1CHRISTIAN TRE´PO,1HANS WILL,3ANDLUCYNA COVA1* Unite´ de Recherche sur les Virus des He´patites, les Re´trovirus Humains, et les Pathologies Associe´es, Institut National
de la Sante´ et de la Recherche Me´dicale 271, 69424 Lyon Cedex 03, France1; Molecular Diagnostics Division, Faculty of Biotechnology, University of Gdansk, 80-822 Gdansk, Poland2; and Heinrich-Pette-Institut fu¨r
Experimentelle Virologie und Immunologie, D-20251 Hamburg, Germany3 Received 3 August 1998/Accepted 14 December 1998
To date, no detailed analysis of the neutralization properties of duck hepatitis B virus (DHBV) has been reported, and it is not clear whether any of the known neutralization epitopes correspond to the viral receptor binding site or to sequences involved in the cell entry pathway. We demonstrate here that antibodies directed against two overlapping peptides (amino acids 83 to 97 and 93 to 107), covering the sequences of most DHBV pre-S neutralizing epitopes, both inhibit virus binding to primary duck hepatocytes and neutralize virus
infectivity. An extensive mutagenesis of the motif 88WTP90, which is the shortest sequence of the epitope
recognized by the virus-neutralizing monoclonal antibody (MAb) 900 was performed in order to define the amino acids involved in these interactions. Single point mutations within this epitope affected neither virus replication nor infectivity but abolished virus neutralization by MAb 900 completely. Interestingly, mutants with two and three consecutive residue replacements (SIP and SIH) within this epitope retained replication competence but were no longer infectious. The loss of infectivity of SIH and SIP mutant particles was
associated with significantly reduced binding to primary duck hepatocytes and could be rescued by trans
complementation with wild-type pre-S protein. Taken together, these results indicate that each amino acid of
the DHBV pre-S sequence88WTP90is critical for recognition by the neutralizing MAb 900 and that
replace-ment of the first two or all three residues strongly reduces virus interaction with hepatocytes and abrogates
infectivity. These data imply that the motif88WTP90contains key residues which are critical for interaction
with both the neutralizing MAb and the host cell.
Duck hepatitis B virus (DHBV) provides a very useful model system with which the role of the hepadnavirus envelope pro-teins in infection, as well as in virus neutralization by antibod-ies, can be studied. Two major cocarboxy-terminal proteins have been identified within the DHBV envelope: a large (L, 36 kDa) and a small (S, 18 kDa) surface protein, both required for viral infectivity. The major DHBV neutralization epitopes map within the pre-S region of L protein, which is thought to me-diate virus-hepatocyte interaction. During the last few years, a number of studies have shown that this pre-S domain takes part in the early steps of DHBV infection and that a 180-kDa carboxypeptidase D protein (gp180), a putative component of the cell receptor protein complex, is suspected of mediating this interaction (3, 13, 14, 16, 28, 30). A domain spanning about one-half of the DHBV pre-S region, mapped to amino acids (aa) 43 to 108 by Ishikawa et al. (13) and to aa 30 to 115 by Urban et al. (30), has been identified as being involved in binding to gp180 and in interaction with the cell surface re-ceptor. In addition, an inner subdomain, the deletion of which abolishes virus interaction with gp180, has been defined within aa 87 to 102 by Tong et al. (28) and aa 85 to 115 by Breiner et al. (3). This region is a part of the highly conserved DHBV envelope protein domain ranging from aa 58 to 107, which contains several neutralization epitopes, i.e., type II (5, 32), type IV (5, 32), M900 (4), and SD20 (4, 17). However, no detailed analysis of the neutralization properties of DHBV has
been reported, and it is not clear whether the same residues are involved in both virus neutralization and interaction with the host cell receptor. In this regard, neutralization of viral infectivity by antibodies is a complex and as yet poorly under-stood phenomenon. Recent studies on the functional domains of proteins suggest that neutralization sites and virus attach-ment sites are often distinct. Therefore, the binding of neu-tralizing antibodies does not necessarily directly block attach-ment to the host cell, as shown for a number of different viruses such as feline immunodeficiency virus (23), feline leukemia virus (26) and influenza virus (22). The neutralizing antibody may interfere with recognition of a host cell by a steric hin-drance, by aggregation of virions, or by inducing a conforma-tional change. There are only a few documented examples of residues within neutralization epitopes which are also involved in the attachment of the virus to its cellular receptor (for a review, see reference 10).
One experimental approach for defining residues involved in neutralization is to select mutant viruses which are able to escape neutralization by monoclonal antibodies (MAbs) and to identify the modified amino acids. Using this approach, we have previously reported the in vivo selection of DHBV pre-S neutralization escape variants by using a murine MAb (MAb 900) which recognizes an epitope mapped between aa 83 and 90 within the DHBV pre-S region (27). These variants har-bored point mutations both at proline 90, within this epitope, and at a distance, at position 5, which almost completely abol-ished virus recognition by neutralizing MAb without affecting viral infectivity (27). However, the selection of neutralization-resistant variants is a strategy which cannot characterize every amino acid in an epitope, since any mutations which interfere
* Corresponding author. Mailing address: INSERM U 271, 151 Cours Albert Thomas, F-69424 Lyon Cedex 03, France. Phone: (33) 472681981. Fax: (33) 472681971. E-mail: [email protected].
2569
on November 9, 2019 by guest
http://jvi.asm.org/
with receptor recognition would lead to noninfectious virus and would thus not be detected in such a study.
In the present study, we have investigated the involvement of specific DHBV pre-S residues in both antibody-mediated neu-tralization and interaction with the host cell by using two com-plementary approaches. First, with a binding test and antisera to synthetic peptides covering the sequence of the clustered DHBV immunodominant epitopes (aa 83 to 107), we show that this domain is involved in virus neutralization and inter-action with hepatocytes. Second, site-directed mutagenesis within the minimum sequence (88WTP90) recognized by the well-characterized DHBV-neutralizing MAb 900 shows that the substitution of a single amino acid is sufficient to abolish virus neutralization but not infectivity, whereas the replace-ment of two or three residues strongly reduces virus-hepato-cyte interaction and abolishes infectivity.
MATERIALS AND METHODS
Antibodies.The rabbit polyclonal antiserum (DPS) used in this study was raised against the first 131 aa of the recombinant DHBV L envelope protein (17). The MAb 900, generated with the same DHBV pre-S domain polypeptide, recognizes a neutralization epitope which was previously characterized and mapped between amino acids I83and Q90(3). The MAb 7C-12, kindly provided
by J. Pugh, is directed against the aa 266 to 275 on the S domain of the DHBV surface proteins (23, 24).
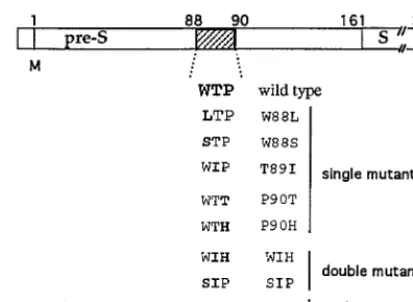
Generation of pre-S mutants.Mutagenesis was performed by two rounds of PCR as previously described (1) by using the wild-type DHBV genome se-quenced by Mandart et al. (21). The mutations introduced led to the replacement of one, two, or three residues within the epitope88WTP90(Fig. 1), which is the
minimum sequence recognized by MAb 900 in PEPSCAN analysis (4). All mutations (except P90T) were designed to avoid introduction of modifications in the overlapping polymerase amino acid sequence. To analyze the impact of mutations on viral replication and infectivity, PCR-mutated fragments were subcloned into an infectious pCMV-DHBV genome (7) by using the appropriate restriction enzyme sites. All mutations were confirmed by sequencing with the Sequenase Version 2.0 DNA sequencing Kit (USB; Amersham). For envelope protein complementation, a pCI–pre-S/S vector was constructed in which the expression of the entire pre-S/S sequence is driven by the early cytomegalovirus promoter. The DHBV DNA fragment (nucleotides 801 to 1785) encoding the pre-S/S gene was PCR amplified and cloned into the polylinker of the pCI expression vector (Promega, Lyon, France).
LMH cell transfection and detection of mutant virus.The avian hepatoma LMH cell line was transfected by the calcium phosphate method with 20mg of DNA plasmid per 100-mm dish as described previously (27). For cotransfection assays the vector pCI–pre-S/S, which expresses DHBV L protein, was used as a helper plasmid to provide wild-type L protein. A total of 20mg of mutated pCMV-DHBV and pCI–pre-S/S plasmids (4:1 ratio) was used. Culture media were collected daily between days 5 and 7 posttransfection, clarified by centrif-ugation at 1,0003gfor 10 min, and concentrated by polyethylene glycol (PEG) precipitation or ultracentrifugation on 10 to 20% sucrose gradients as previously described (27). The presence of mutations was confirmed by PCR and direct
infected with 2310 VGE/well of wild-type virus or DHBV pre-S mutants, preincubated overnight either with L15 culture medium (controls) or with mu-rine or rabbit antibody as described earlier (18). The release of viral particles into cell culture media was monitored for 8 days by quantitative dot blot hybridiza-tion. At the end of the culture, PDHs were harvested, and intracellular viral DNA was analyzed by Southern blot as previously described (18). All infection and neutralization assays were performed in duplicate and reproduced twice by using two series of inocula prepared independently.
In vivo infectivity of DHBV mutants.Three-day-old DHBV-free Pekin duck-lings were intravenously inoculated with quantified wild-type or mutant DHBV inocula concentrated from transfected LMH cells (23108VGE/duck). Viremia
was followed every day during the first week and then every 2 days for the two following weeks as described previously (8, 27).
Immunoblotting.The expression of DHBV L protein was monitored in trans-fected cells or PDHs by Western blotting with rabbit polyclonal anti-DHBV pre-S serum (diluted 1:1,000), anti-peroxidase conjugate (diluted 1:1,000; Bio-sys), and the ECL chemiluminescence detection kit (Amersham) as reported previously (17, 27).
Analysis of DNA from infected primary hepatocytes and duck livers.Primary hepatocytes were collected in lysis buffer containing 0.5% sodium dodecyl sulfate (SDS) at day 10 postinfection and assayed for viral replicative intermediates. After proteinase K digestion (100 mg/ml, 4 h at 37°C), the total DNA was extracted as previously described (2, 27). The same procedure was applied to homogenized duck liver samples. Equal amounts of DNA from each sample were analyzed by electrophoresis through a 1% agarose gel, transferred to a nylon membrane, and hybridized with a32P-labelled full-length genomic DHBV DNA
probe as previously described (2, 11).
Peptide synthesis and preparation of anti-peptide antibodies.Two overlap-ping peptides PS1 (aa 83 to 97; IPQPQWTPEEDQKAR) and PS2 (aa 93 to 107; DQKAREAFRRYQEER) were synthesized in solid phase by using 9-fluoroe-nylmethoxycarbonyl technology and chemically coupled to a carrier protein, keyhole limpet hemocyanin, by the method of Liu (20). Two rabbits were im-munized separately with 200mg of each peptide followed by three boosts at days 14, 28, and 56 by using standard protocols (Neosystem, Strasbourg, France). The immune response to the peptide was evaluated by determining the endpoint titer of antibodies that reacted with recombinant DHBV pre-S protein in an enzyme-linked immunosorbent assay (ELISA) as described previously (4).
Assay to detect DHBV particles bound to PDHs.The assay we have used is essentially the one described by Pugh et al. (25). Briefly, PDHs were prepared as described above and plated at high density in six-well plates. Two or three days after plating, cells were washed with phosphate-buffered saline (PBS) and incu-bated (1 h 30 min at room temperature) with DHBV diluted in Opti-MEM medium (Life Technologies) or preincubated overnight with different rabbit anti-preS sera (DPS, anti-PS1, and anti-PS2) diluted to a similar anti-DHBV pre-S titer as assessed by ELISA. After three washes, anti-S MAb 7C-12 (diluted 1/100 in PBS–5% normal duck serum [NDS]) was added to the cells and incu-bated under the same conditions. Cells were washed again and then incuincu-bated for 30 min with125I-labeled goat anti-mouse immunoglobulin G (IgG) (Biosys)
diluted in PBS–5% NDS. After three washes, cells were lysed with PBS–1% SDS, and the cell-associated radioactivity was determined in a gamma counter.
RESULTS
Antibodies to DHBV neutralization epitopes inhibit DHBV
binding to PDHs.First, we determined whether virus binding
[image:2.612.67.274.70.221.2]to PDHs is affected by anti-DHBV pre-S antibodies against recombinant pre-S protein or synthetic peptides. The binding assay was performed by measuring the amount of PDH-asso-ciated viral particles after incubation with DHBV-positive duck serum, detected by using the MAb 7C-12, which is known to specifically detect DHBV S protein (25) and I125-labeled goat anti-mouse IgG. The results found with PDHs incubated with DHBV-positive serum showed that DHBV binds to PDH, FIG. 1. Schematic representation of amino acid substitutions in the pre-S
domain of the DHBV L envelope protein. The mutant names refer to the location and the nature of the amino acid changes within the88WTP90 neutral-ization epitope recognized by MAb 900.
on November 9, 2019 by guest
http://jvi.asm.org/
whereas no significant binding was observed when hepatocytes were incubated with DHBV-negative (mock) serum (Fig. 2). In addition, the cell-associated radioactivity was proportional to the amounts of viral particles used (results not shown). Taken together, these results indicate that this assay allows the de-tection of PDH-associated DHBV particles.
Preincubation of viral particles with nonimmunized rabbit serum has no effect on binding, whereas preincubation with the recombinant DHBV pre-S polypeptide (aa 1 to 131) antiserum (DPS) reduced binding by 75% (Fig. 2). Both of the two an-tipeptide sera, which recognize part of the pre-S region thought to be involved in recognition of the hepatocyte recep-tor, namely, aa 83 to 97 (PS1) and aa 93 to 107 (PS2), reduced binding by about 65%. Simultaneous addition of both antipep-tide sera reduced the binding of viral particles even further to levels similar to those observed with the recombinant DHBV pre-S polypeptide antiserum directed against almost the entire pre-S region. These data suggest that either the major hepa-tocyte binding domain of the pre-S protein or a critical part of it is located between aa 83 and 107.
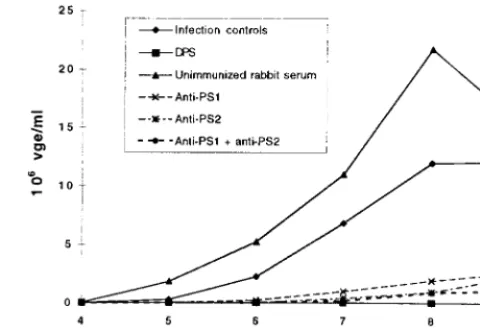
Next, we tested whether the polyclonal antibodies against the recombinant pre-S protein or the two peptides are izing. The in vitro neutralizing test showed that 100% neutral-ization was achieved by using DPS, whereas serum from a nonimmunized rabbit was not neutralizing (Fig. 3). Both PS1 (aa 83 to 97) and PS2 (93 to 107) antisera are highly neutral-izing, since a decrease by 89 and 94%, respectively, of released virions was observed in PDHs infected with DHBV preincu-bated with these sera (Fig. 3). Taken together, these results indicate that antibodies elicited to the aa 83 to 107 domain within DHBV pre-S both inhibit virus binding to PDHs and neutralize DHBV infectivity.
DHBV pre-S mutants are replication competent but differ in
their infectivity.To define which, if any, of the amino acids
within this domain plays a role in virus infectivity, we chose to introduce into the pCMV-DHBV plasmid 1-, 2-, or 3-amino-acid replacements in the 88WTP90 motif recognized by the highly neutralizing MAb 900 (Fig. 1). From LMH cells trans-fected with all eight mutated plasmids, designated W88L, W88S, T89I, P90H, P90T, WIH, SIH, and SIP, and the wild type, a progressive release of viral DNA into the cell culture
medium was observed (Fig. 4A). That this represents secreted viral particles was further confirmed by the detection of similar amounts of the 35- to 36-kDa DHBV pre-S/S proteins in the concentrated culture medium of cells transfected either with the mutants or with the wild-type plasmid (Fig. 4B). To ensure that all of the mutations were maintained and that all of the mutants are replication competent and really secreted envel-oped DNA-containing viral particles, we have immunoprecipi-tated viral particles from supernatants of transfected cells after DNase I treatment and confirmed, by sequencing of the entire DHBV pre-S region (nucleotides 628 to 1485), that the DHBV DNAs had the same sequence as the respective constructions used for the transfection (data not shown).
[image:3.612.58.292.70.206.2]To test the infectivity of the secreted viral particles, trans-fected LMH cell culture media were concentrated with PEG FIG. 2. Binding of DHBV to PDHs is inhibited by antibodies to
neutraliza-tion epitopes. PDH monolayers were incubated with concentrated DHBV-pos-itive duck serum (DHBV) or uninfected duck serum (Mock). To test the capacity of DHBV pre-S antisera to inhibit binding, virus was preincubated with sera from a nonimmunized rabbit, DHBV pre-S polypeptide (aa 1 to 131) rabbit antiserum (DPS), or83I-R97(PS1), or/and93D-R107(PS2) peptide antisera. All antisera were diluted to similar anti-DHBV pre-S titers, as assessed by ELISA. Bound virus was detected with anti-S MAb 7C-12 and125I-labeled goat anti-mouse IgG as described in Materials and Methods. The counts obtained for control DHBV are represented as 100%.
FIG. 3. DHBV infectivity is neutralized in vitro by PS1 and PS2 peptide antisera. PDHs were infected with DHBV-positive duck serum (23108VGE/ well), preincubated with L15 medium (infection controls), unimmunized rabbit serum, DPS, or PS1 (83I-R97), PS2 (93D-R107), or PS1 and PS2 rabbit antisera. The release of viral particles into the supernatants was quantified by dot blot hybridization as described in Materials and Methods. The means of duplicate determinations are presented.
FIG. 4. DHBV pre-S mutations do not affect virus replication capacity. pCMV-DHBV DNA plasmids containing each mutation presented in Fig. 1 were used to transfect the LMH hepatoma cell line. (A) Virus release into the trans-fected cell supernatants was monitored by dot blot hybridization as described in Materials and Methods. Lane 1, wild type; lane 2, W88L; lane 3, W88S; lane 4, T89I; lane 5, P90H; lane 6, P90T; lane 7, WIH; lane 8, SIP; lane 9, SIH. (B) Viral particles from the culture supernatants were concentrated and analyzed by im-munoblotting. Proteins were probed with rabbit anti-DHBV pre-S antibody (DPS). Lane 1, wild type; lane 2, W88L; lane 3, W88S; lane 4, T89I; lane 5, P90H; lane 6, P90T; lane 7, WIH; lane 8, SIP; lane 9, SIH; lane 10, negative control. For comparison, circulating viral particles concentrated from DHBV-positive duck serum were loaded into lane 11.
on November 9, 2019 by guest
http://jvi.asm.org/
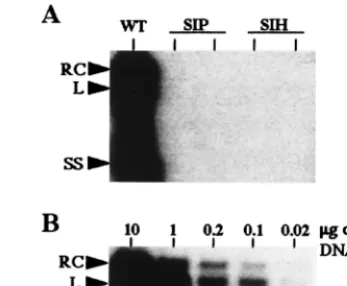
[image:3.612.313.553.74.240.2]and used to infect the PDHs. It was important to ascertain that the infection with different mutant or wild-type viruses was normalized to the same number of enveloped particles. To this end, we have separated the completely enveloped virions from unenveloped cores by CsCl equilibrium ultracentrifugation of LMH cell supernatants. Quantification of the DHBV DNA in all fractions by dot blot hybridization and pre-S protein by immunoblotting (data not shown) indicated that approximately 60% of the viral DNA sedimented at a density of 1.18 g/ml corresponding to the density of enveloped particles and 40% of the viral DNA sedimented as unenveloped cores at a density of 1.3 g/ml, a finding consistent with previous observations (2). The same respective proportions were found to be similar for wild-type and different mutant virus preparations. The single-amino-acid mutants W88L, W88S, P90T, and P90H and the double-mutant WIH were infectious in vitro, and the virus titers were similar to those obtained for the wild-type DHBV. The mutant T89I was also infectious, but the DNA levels in the cell culture medium were about fourfold lower (Table 1). How-ever, the SIP and SIH mutants, which have 2- and 3-amino-acid changes, respectively, failed to infect hepatocyte cultures, as shown by the absence of detectable viral proteins and DNA signals in cell culture lysates and media (Table 1 [see also Fig. 7] and data not shown). Since the PDH infection assay has a limited sensitivity and may not reflect all aspects of an in vivo infection, we also examined the infectivity of these two mutants and of all the other mutants in ducklings. Nine groups of 3-day-old ducklings were infected with concentrated viral par-ticles from transfected LMH cells. As expected from the in vitro results, all single mutants and the WIH double mutant were infectious in vivo (Table 1). In contrast, no DHBV DNA was detected in the sera of ducklings infected with the SIP and SIH mutants (Table 1). Moreover, no intrahepatic DNA was detected in the corresponding livers of these ducklings by Southern blot hybridization (Fig. 5A). This technique allows the detection of viral DNA in as little as 0.02mg of total liver
DNA (Fig. 5B). The direct sequencing of the DHBV pre-S DNA fragments amplified from viremic sera indicated that all mutations were maintained after one in vivo passage (data not shown). Altogether, these results demonstrate that all single-amino-acid mutants tested and one double mutant (WIH) with the last 2 aa changed in the WTP motif do not or only mar-ginally affect viral infectivity in vitro or in vivo. In contrast, changing of the first 2 (SIP mutant) or all 3 aa (SIH mutant) of the WTP motif abrogates DHBV infectivity, which may imply a function of these amino acids in receptor recognition or downstream infection events.
Neutralization of DHBV by MAb 900 is abolished by the
WTP mutations.To investigate the impact of DHBV
neutral-ization by MAb 900 on those mutants that retain infectivity, we performed an in vitro neutralization assay. As illustrated in Fig. 6 and summarized in Table 1, all mutants were fully neu-tralized in vitro, when incubated with the highly neutralizing DPS polyclonal antibody but none were neutralized by MAb 900. From these results we conclude that each of the residues of the WTP motif is essential for the neutralization of virus infectivity by MAb 900. The neutralization of DHBV mutants by DPS is in agreement with our previous studies which showed that this polyclonal antibody recognizes, in addition to the MAb 900 epitope, other neutralization epitopes within the DHBV pre-S protein (27).
Wild-type DHBV L protein trans-complements mutant
in-fectivity. The two mutants SIP and SIH were found to be
noninfectious. We examined whether their infectivity could be restored by providing the wild-type DHBV L protein intrans
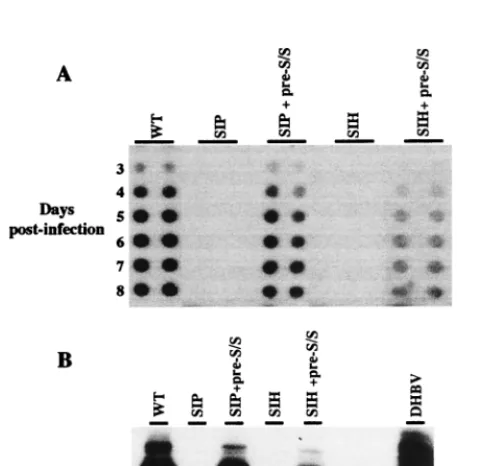
[image:4.612.341.515.74.217.2]by cotransfecting LMH cells with SIP- or SIH-encoding plas-mids and a wild-type L protein expression vector. The virus stocks obtained were then used to infect PDHs as described above. Cotransfection of LMH cells with the SIH or SIP mu-tant and pCI–pre-S/S plasmid resulted in the release of viral particles which infected PDH, as judged by detection of both DHBV DNA in PDH media (Fig. 7A) and L proteins within cell lysates (Fig. 7B). This indicates that the infectivity of both mutants can be rescued by the wild-type envelope protein and implies that the corresponding mutations affect the pre-S pro-tein domain function only.
FIG. 5. SIP and SIH mutant viruses are not infectious in vivo. Viral particles produced by LMH cells transfected with pCMV-DHBV plasmids bearing the double (SIP) or triple (SIH) substitutions were used to infect ducklings. (A) Intrahepatic DHBV DNA replicative forms were analyzed by Southern blotting 3 weeks after inoculation in the liver from one duckling inoculated with wild-type DHBV (control) and two ducklings inoculated with either SIP or SIH mutant viruses. (B) Sensitivity of intrahepatic DHBV DNA detection in successive di-lutions of the total liver DNA from a DHBV-positive liver. Arrows indicate the relaxed circular (RC), linear (L), and single-stranded (SS) DHBV DNA forms.
WIH 1 42 3/3 1 2
SIP 1 0 0/4 NTe NT
SIH 1 0 0/3 NT NT
aFive days after LMH cell transfection with pCMV-DHBV plasmids, DHBV
DNA and proteins were detected in cell lysates and supernatants as described in Materials and Methods.
bFollowing PDH infection with supernatants from transfected LMH cells,
released virions were quantified in culture medium as described in Materials and Methods.
cNumber of ducks positive for DHBV DNA in serum and liver/total number
of inoculated ducks.
dFor the in vitro neutralization assay, PDHs were infected with virus
prein-cubated with rabbit anti-pre-S serum (DPS) or MAb 900. Significant neutraliza-tion (1) is taken to be a 50% or greater reduction in the supernatant viral DNA level.2, Absence of neutralization.
eNT, not tested for neutralization since found to be noninfectious.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.52.293.92.232.2]Binding of noninfectious mutant virus to PDHs is affected.
The data presented above indicate that the lack of infectivity of the SIH and SIP mutants is due to the mutant DHBV L envelope protein. It is not clear whether this interferes
nega-tively with the binding of the mutant viral particles to the cellular receptor or with a subsequent step in infection. In order to examine the first possibility, we performed a binding assay with SIP and SIH mutant viral particles. In PDHs treated with equivalent amounts of mutant or wild-type particles, 60% fewer mutant pre-S particles were bound (Fig. 8), indicating that the amino acid changes significantly reduce but do not abolish the binding of the mutant particles to PDHs.
DISCUSSION
In the present study, we have demonstrated that residues 88 to 90 within the DHBV L envelope protein are critical for virus neutralization and are involved in the interaction with the host cell. This is of particular interest, since the roles of hepadna-virus envelope protein in the earliest events of infection (i.e., virus interaction with the host cell membrane, penetration, and disassembly) and virus neutralization are still poorly under-stood. Our data are consistent with and extend previous data and speculation about the type of pre-S sequences involved in receptor interaction by pointing to the involvement of a short-amino-acid motif in the pre-S region. A large portion (aa 30 to 115) of DHBV L protein has recently been shown to interact with carboxypeptidase gp180, which is thought to be part of a cellular receptor for avian hepadnaviruses (3, 30). Within this pre-S domain, a recombinant polypeptide covering the resi-dues 87 to 102 has been shown to bind specifically to a gp180 in an in vitro binding test (28). Since aa 87 to 102 harbor three major neutralization epitopes (4, 5, 6, 17, 32), it has been speculated that the sequences comprising the neutralization epitopes are also involved in the interaction with hepatocyte (28). However, the nature of viral epitopes involved in neu-tralization has been analyzed in detail for only some viruses, and it has been shown that they may be either identical to or distinct from the receptor binding site (10, 22, 23, 26). In this regard, our data are of particular interest since they provide the first experimental support for the hypothesis that some residues involved in DHBV neutralization are also involved in the early stages of interaction with the host cell.
[image:5.612.109.237.72.261.2]We found that rabbit antibodies elicited by two overlapping peptides (I83-R97) and (D93-R107) covering the cluster of three DHBV pre-S neutralizing epitopes (aa 83 to 107) were able to both inhibit virus binding to primary duck hepatocytes and neutralize virus infectivity. To define these interactions at the
[image:5.612.311.552.73.215.2]FIG. 6. Mutations within the WTP epitope abolish in vitro neutralization of DHBV by MAb 900. PDH cultures were infected with wild-type DHBV or P90H mutant particles preincubated either with L15 culture medium (controls), with MAb 900 ascites fluid, or with DPS polyclonal antibody as described in Materials and Methods. Viral release into the culture medium was monitored from days 5 to 10 postinfection by dot blot hybridization.
[image:5.612.52.292.403.636.2]FIG. 7. Wild-type (WT) DHBV L proteintrans-complements SIP and SIH mutant infectivity. LMH cells were transfected with pCMV-DHBV plasmids bearing the double (SIP) or triple (SIH) substitutions in the presence or absence of pCI–pre-S/S vector, which expresses wild-type L protein. PDH cultures were infected with viral particles produced by transfected LMH cells. (A) The viral release into cell supernatants was monitored from days 3 to 8 postinfection by dot blot hybridization. (B) Cells were harvested 8 days postinfection, and the L protein was detected in the cell lysate by immunoblotting with rabbit anti-DHBV pre-S antibody. For comparison, circulating viral particles concentrated from DHBV-positive duck serum were loaded in the right lane. The arrow shows the position of the 36-kDa DHBV L protein.
FIG. 8. Binding of SIP and SIH mutant virus to PDH is reduced. PDH monolayers were incubated with wild type (WT), SIP, or SIH viral particles produced by transfected LMH cells as described in Materials and Methods. The results are expressed as the percentages of counts obtained with wild-type DHBV particles, after substraction of the values obtained with concentrated medium from mock-transfected LMH cells.
on November 9, 2019 by guest
http://jvi.asm.org/
DHBV L protein (2). It is therefore probable that the reduc-tion of viral secrereduc-tion observed after infecreduc-tion of primary he-patocyte cultures with the mutant T89I is not due to an impact on phosphorylation.
By contrast, analysis of the SIP and SIH mutants has re-vealed that these double and triple amino acid substitutions abrogate DHBV infectivity in vitro and in vivo without affect-ing their replication capacity. The wild-type-like replication capacity of these two mutant plasmids in transfected LMH cells is consistent with the fact that none of the mutations introduced altered the P-protein sequence. Moreover, the wild-type-like efficiency of viral particle secretion of these mu-tants indicates that no other transcription regulatory sequences are affected by these mutations. Therefore, our results strongly suggest that these mutations affect early stages of virus infec-tion only. This suggesinfec-tion is corroborated by complementainfec-tion assays that showed that the DHBV L envelope protein pro-vided intransis able to restore the infectivity of the mutants. The question remained whether the mutations affect the initial step of specific binding to hepatocyte membranes or subsequent steps in viral penetration into the duck hepato-cytes. Recent data provide evidence that gp180, a Golgi-resi-dent protein, is involved in the first step of avian hepadnavirus uptake (3). This initial step involves the interaction of gp180 with a defined subdomain of DHBV pre-S which has been mapped between aa 85 and 115 by using recombinant DHBV pre-S polypeptides in an in vitro infection competition assay (3, 30). Interestingly, within this pre-S region, a short sequence spanning residues 85 to 96 has been recently identified as an absolutely essential element for receptor interaction (31). Us-ing a completely different approach, i.e., site-directed mu-tagenesis, we have demonstrated that mutants which harbor the replacement of residues 88 and 89 within this short amino acid stretch completely lose their infectivity in PDH cultures and in ducklings. These observations are of particular interest since they provide the first evidence that the portion of the DHBV pre-S protein which is prerequisite for gp180 recogni-tion contains the residues which are essential for in vivo initi-ation of DHBV infection. However, the direct interaction of the noninfectious mutants SIP and SIH with gp180 was not investigated in the present study. The binding assay we have developed showed that these mutants bind to PDHs with an efficiency decreased by 60% compared to that of wild-type particles. In spite of this significant decrease in mutant binding, it is unclear why the 40% of residual binding does not allow viral infection. Considering that gp180 is a Golgi-resident pro-tein which cycles to and from plasma membrane (3), it is difficult to evaluate the number of binding sites at the hepato-cyte surface in our binding test. Further study of the interac-tion of SIP and SIH mutants with recombinant gp180 protein will provide a definite answer as to what extent, if at all, the lack of infectivity of the SIH and SIP mutants is due to their inability to recognize and/or strongly attach to the hepatocyte
action with both the host cell and the neutralizing MAb, al-though this does not mean that other portions of the DHBV L protein are not involved in the recognition of the cellular receptor or in virus neutralization. In this regard, it has been recently demonstrated that recombinant polypeptides covering a large portion (aa 30 to 115) of DHBV or heron hepatitis B virus (HHBV) pre-S region compete equally well for interac-tion with gp180, despite a 50% difference in their amino acid sequences, suggesting that the three-dimensional structure of DHBV pre-S is important for interaction with the cellular receptor (30). The three-dimensional structure of DHBV pre-S seems also important for virus neutralization, since we have previously shown that P90within the 83–90 epitope stud-ied here and a distantly located residue, P5, are both involved in DHBV neutralization (27). For HBV it has been demon-strated, by using the random peptide library approach, that an anti-pre-S1 MAb which prevents virus binding to liver mem-branes is also able to recognize distantly located pre-S2 epitopes, suggesting that the hepatocyte-binding domain of the surface proteins is conformationally dependent (9). The use of combinatorial libraries to define mimotopes recognized by an-ti-DHBV pre-S MAbs will be of interest in defining the tertiary structures involved in DHBV neutralization and/or the cell entry pathway.
ACKNOWLEDGMENTS
C.S. was the recipient of fellowships from Institut Pasteur Lyon and the Ligue Nationale Contre le Cancer. This work was supported in part by the Institut National de la Sante´ et de la Recherche Me´dicale, the French Association de la Recherche pour le Cancer (ARC), and Ligue Nationale de Recherche sur le Cancer.
We thank S. Urban for sharing unpublished observations. We are grateful to C. Jamard for expert assistance with animals.
REFERENCES
1.Baowei, C., and A. E. Przyla.1994. An efficient site-directed mutagenesis method based on PCR. BioTechniques17:657–659.
2.Borel, C., C. Sunyach, O. Hantz, C. Tre´po, and A. Kay.1998. Phosphoryla-tion of DHBV pre-S: identificaPhosphoryla-tion of the major site of phosphorylaPhosphoryla-tion and effects of mutations on the virus life cycle. Virology242:90–98.
3.Breiner, K., S. Urban, and H. Schaller.1998. Carboxypeptidase D (gp180), a Golgi-resident protein, functions in the attachement and entry of avian hepatitis B viruses. J. Virol.72:8098–8104.
4.Chassot, S., V. Lambert, A. Kay, C. Godinot, B. Roux, C. Trepo, and L. Cova.
1993. Fine mapping of neutralization epitopes on duck hepatitis B virus (DHBV) pre-S protein using monoclonal antibodies and overlapping pep-tides. Virology192:217–223.
5.Cheung, R. C., W. Robinson, P. L. Marion, and H. B. Greenberg.1989. Epitope mapping of neutralization monoclonal antibodies against duck hep-atitis B virus. J. Virol.63:2445–2451.
6.Cheung, R. C., D. E. Trujillo, W. Robinson, H. B. Greenberg, and P. L. Marion.1990. Epitope-specific antibody response to the surface antigen of duck hepatitis B virus in infected ducks. Virology176:546–552.
7.Condreay, L. D., C. E. Aldrich, L. Coastes, W. S. Mason, and T. Wu.1990. Efficient duck hepatitis B virus production by avian tumor cell line. J. Virol.
63:2445–3258.
8.Cova, L., C. P. Wild, R. Mehrotra, V. Turusov, T. Shirai, V. Lambert, C.
on November 9, 2019 by guest
http://jvi.asm.org/
Jacquet, L. Tomatis, C. Trepo, and R. Montesano.1990. Contribution of aflatoxin B1 and hepatitis B virus infection in the induction of liver tumours in ducks. Cancer Res.50:2156–2163.
9.D’Mello, F., C. D. Partidos, M. W. Steward, and C. R. Howard.1997. Definition of the primary structure of hepatitis B virus pre-S hepatocyte binding domain using random peptide libraries. Virology237:319–326. 10. Dimmock, N. J.1993. Immunoglobulin G neutralization by inhibition of
attachment of virus to the cell, p. 3–9.InNeutralization of animal viruses. Springer-Verlag, Berlin, Germany.
11. Fourel, I., P. Gripon, O. Hantz, L. Cova, V. Lambert, C. Jacquet, K. Wa-tanabe, J. Fox, C. Guillouzo, and C. Trepo.1989. Prolonged duck hepatitis B virus replication in duck hepatocytes cocultivated with rat epithelial cells: a useful system for antiviral testing. Hepatology10:186–191.
12. Grgacic, E. V. L., and D. A. Anderson.1994. The large surface protein of duck hepatitis B virus is phosphorylated in the pre-S domain. J. Virol.
68:7344–7350.
13. Ishikawa, T., K. Kuroki, R. Lenhoff, J. Summers, and D. Ganem.1994. Analysis of a host cell surface glycoprotein to the pre-S protein of duck hepatitis B virus. Virology202:1061–1064.
14. Klingmu¨ller, U., and H. Schaller.1993. Hepadnavirus requires interaction between the viral pre-S domain and a specific hepatocellular receptor. J. Vi-rol.67:7414–7422.
15. Kuroki, K., R. Cheung, P. L. Marion, and D. Ganem.1994. A cell surface protein that bind avian hepatitis B virus particles. J. Virol.68:2091–2096. 16. Kuroki, K., F. Eng, T. Ishikawa, C. Turck, F. Harada, and D. Ganem.1995.
gp180, a host cell glycoprotein that binds duck hepatitis B virus particles, is encoded by a member of the carboxypeptidase gene family. J. Biol. Chem.
270:15022–15028.
17. Lambert, V., D. Fernholz, R. Sprengel, I. Fourel, G. Deleage, G. Wildner, C. Peyret, C. Trepo, L. Cova, and H. Will.1990. Virus-neutralizing monoclonal antibody to a conserved epitope on the duck hepatitis B virus pre-S protein. J. Virol.64:1290–1297.
18. Lambert, V., S. Chassot, A. Kay, C. Tre´po, and L. Cova.1991.In vivo neutralization of duck hepatitis B virus by antibodies specific to the N-terminal portion of pre-S protein. Virology185:446–450.
19. Lenhoff, R. J., and J. Summers.1994. Coordinate regulation of replication and virus assembly by the large envelope protein of an avian hepadnavirus. J. Virol.68:4565–4571.
20. Liu, F. T.1979. New procedures for preparation and isolation of conjugates
of proteins and a synthetic copolymer of D-amino-acids and immunochemi-cal characterization of such conjugates. Biochemistry18:690–697. 21. Mandart, E., A. Kay, and F. Galibert.1984. Nucleotide sequence of a cloned
duck hepatitis B virus genome: comparison with woodchuck and human hepatitis B virus sequences. J. Virol.49:782–792.
22. Outlav, M. C., and N. J. Dimmock.1990. Mechanisms of neutralization of influenza virus on mouse tracheal epithelial cells by mouse monoclonal polymeric IgA and polyclonal IgM directed against the viral hemagglutinin. J. Gen. Virol.71:69–76.
23. Pancino, G., and P. Sonigo.1997. Retention of viral infectivity after extensive mutation of the highly conserved immunodominant domain of the feline immunodeficiency virus envelope. J. Virol.71:4339–4346.
24. Pugh, J. C., and H. Simmons.1994. Duck hepatitis B virus infection of muscovy duck hepatocytes and nature of resistance in vivo. J. Virol.68:2487– 2494.
25. Pugh, J. C., D. Qu, S. W. Mason, and H. Simmons.1995. Susceptibility to duck hepatitis B virus infection is associated with the presence of cell surface receptor sites that efficiently bind viral particles. J. Virol.69:4814–4822. 26. Ramsey, I. K., N. Spibey, and O. Jarret.1998. The receptor binding site of
feline leukemia virus surface glycoprotein is distinct from the site involved in virus neutralization. J. Virol.72:3268–3277.
27. Sunyach, C., S. Chassot, C. Jamard, A. Kay, C. Trepo, and L. Cova.1997.In vivoselection of duck hepatitis B virus pre-S variants which escape from neutralization. Virology234:291–299.
28. Tong, S., J.-S. Li, and J. R. Wands.1995. Interaction between duck hepatitis B virus and a 170-kilodalton cellular protein is mediated through a neutral-izing epitope of the pre-S region and occurs during viral infection. J. Virol.
69:7106–7112.
29. Tuttleman, J. S., J. C. Pugh, and J. W. Summers.1986. In vitro experimental infection of primary duck hepatocytes cultures with duck hepatitis B virus. J. Virol.58:17–25.
30. Urban, S., K. M. Breiner, F. Fehler, U. Kingmu¨ller, and H. Schaller.1998. Avian hepatitis B virus infection is initiated by the species independent interaction of a distinc pre-S subdomain with its cellular receptor. J. Virol.
72:8089–8097.
31. Urban, S.Personal communication.
32. Yuasa, S., R. C. Cheung, Q. Pham, W. S. Robinson, and P. L. Marion.1991. Peptide mapping of neutralizing and nonneutralizing epitopes of duck hep-atitis B virus pre-S polypeptide. Virology181:14–21.